PHB Production in Biofermentors Assisted through Biosensor Applications †



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Plasmid Preparation for E. coli Transformation
2.2. Culture Media and Conditions
2.3. Amplification and Cloning of phaCAB Genes in E. coli BL21(DE3)
2.4. Analytical Procedures during Biofermentor Use
2.4.1. Quantification of Reducing Sugars
2.4.2. Cell Dry Weight (CDW)
2.4.3. Monitoring the Production of PHB by Nile Blue Staining
2.4.4. PHB/PHA Quantification and Extraction
3. Results and Discussion
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| PHA | poylhydroxyalanoate |
| PHB | polyhydroxybutyrate |
| PHB-V | mixed type PHA, containing hydroxyvalerate |
| EIS | electrochemical impedance spectroscopy |
| FTIR | Fourier Transformation-IR Spectrometry |
| SERS | Surface Enhanced Rama Spectroscopy |
References
- Dietrich, D.; Illman, B.; Crooks, C. Differential sensitivity of polyhydroxyalkanoate producing bacteria to fermentation inhibitors and comparison of polyhydroxybutyrate production from Burkholderia cepacia and Pseudomonas pseudoflava. BMC Res. Notes 2013, 6, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Guevara-Martínez, M.; Sjöberg Gällnö, K.; Sjöberg, G.; Jarmander, J.; Perez-Zabaleta, M.; Quillaguamán, J.; Larsson, G. Regulating the production of (R)-3-hydroxybutyrate in Escherichia coli by N or P limitation. Front. Microbiol. 2015, 6, 844. [Google Scholar] [CrossRef] [PubMed]
- El-sayed, A.A.; Abdel Hafez, A.M.; Hemmat Abdelhady, M.; Khodair, T.A. Production of Polyhydroxybutyrate (PHB) using batch and two-stage batch culture strategies. Aust. J. Basic Appl. Sci. 2009, 3, 617–627. [Google Scholar]
- Chen, G.Q.; Page, W.J. Production of poly-b-hydroxybutyrate by Azotobacter vinelandii in a two-stage fermentation process. Biotechnol. Tech. 1997, 11, 347–350. [Google Scholar] [CrossRef]
- Singhaboot, P.; Kaewkannetra, P. A higher in value biopolymer product of polyhydroxyalkanoates (PHAs) synthesized by Alcaligenes latus in batch/repeated batch fermentation processes of sugar cane juice. Ann. Microbiol. 2015, 65, 2081–2089. [Google Scholar] [CrossRef]
- Ryu, H.W.; Hahn, S.K.; Chang, Y.K.; Chang, H.N. Production of poly(3-hydroxybutyrate) by high cell density fed-batch culture of Alcaligenes eutrophus with phospate limitation. Biotechnol. Bioeng. 1997, 55, 28–32. [Google Scholar] [CrossRef]
- Urtuvia, V.; Villegas, P.; González, M.; Seeger, M. Bacterial production of the biodegradable plastics Polyhydroxyalkanoates. Int. J. Biol. Macromol. 2014, 70, 208–213. [Google Scholar] [CrossRef]
- Nikel, P.I.; de Almeida, A.; Melillo, E.C.; Galvagno, M.A.; Pettinari, M.J. New recombinant Escherichia coli strain tailored for the production of poly(3-hydroxybutyrate) from agroindustrial by-products. Appl. Environ. Microbiol. 2006, 72, 3949–3954. [Google Scholar] [CrossRef]
- Saranya, V.; Shenbagarathai, R. Production and characterization of PHA from recombinant E. coli harbouring phac1 gene of indigenous Pseudomonas sp. ldc-5 using molasses. Braz. J. Microbiol. 2011, 42, 1109–1118. [Google Scholar] [CrossRef]
- Li, Z.J.; Shi, Z.Y.; Jian, J.; Guo, Y.Y.; Wu, Q.; Chen, G.Q. Production of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) from unrelated carbon sources by metabolically engineered Escherichia coli. Metab. Eng. 2010, 12, 352–359. [Google Scholar] [CrossRef]
- Zhou, X.Y.; Yuan, X.X.; Shi, Z.Y.; Meng, D.C.; Jiang, W.J.; Wu, L.P.; Chen, J.C.; Chen, G.Q. Hyperproduction of poly(4-hydroxybutyrate) from glucose by recombinant Escherichia coli. Microb. Cell Fact. 2012, 11. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Zhang, Y.; Yuan, Q.; Liu, Q.; Li, Y.; Wang, Z.; Ma, H.; Chen, T.; Zhao, X. Metabolic engineering of Escherichia coli for poly(3-hydroxybutyrate) production via threonine bypass. Microb. Cell Fact. 2015, 14, 185. [Google Scholar] [CrossRef] [PubMed]
- Mahishi, L.H.; Tripathi, G.; Rawal, S.K. Poly(3-hydroxybutyrate) (PHB) synthesis by recombinant Escherichia coli harbouring Streptomyces aureofaciens PHB biosynthesis genes: Effect of various carbon and nitrogen sources. Microbiol. Res. 2003, 158, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lin, Z.; Liu, Q.; Li, Y.; Wang, Z.; Ma, H.; Chen, T.; Zhao, X. Engineering of serine-deamination pathway, Entner-Doudoroff pathway and pyruvate dehydrogenase complex to improve poly(3-hydroxybutyrate) production in Escherichia coli. Microb. Cell Fact. 2014, 13, 172. [Google Scholar] [CrossRef]
- Cesário, M.T.; Raposo, R.S.; de Almeida, M.C.M.D.; van Keulen, F.; Ferreira, B.S.; da Fonseca, M.M.R. Enhanced bioproduction of poly-3-hydroxybutyrate from wheat straw lignocellulosic hydrolysates. New Biotechnol. 2014, 31, 104–113. [Google Scholar] [CrossRef]
- Haas, C.; Steinwandter, V.; de Apodaca, E.D.; Maestro Madurga, B.; Smerilli, M.; Dietrich, T.; Neureiter, M. Production of PHB from chicory roots—Comparison of three Cupriavidus necator strains. Chem. Biochem. Eng. Q. 2015, 29, 99–112. [Google Scholar] [CrossRef]
- TransBio. EU Framework Program FP7 Project. Biotransformation of by-Products from Fruit and Vegetable Processing Industry into Valuable Bioproducts. 2012. Available online: http://www.transbio.eu (accessed on 30 October 2016).
- Wang, F.; Lee, S.Y. High cell density culture of metabolically engineered Escherichia coli for the production of poly(3-hydroxybutyrate) in a defined medium. Biotechnol. Bioeng. 1998, 58, 325–328. [Google Scholar] [CrossRef]
- Ienczak, J.L.; Schmidell, W.; de Aragão, G.M.F. High-cell-density culture strategies for polyhydroxyalkanoate production: A review. J.Ind. Microbiol. Biotechnol. 2013, 40, 275–286. [Google Scholar] [CrossRef]
- Peña, C.; Castillo, T.; García, A.; Millán, M.; Segura, D. Biotechnological strategies to improve production of microbial poly-(3-hydroxybutyrate): A review of recent research work. Microb. Biotechnol. 2014, 7, 278–293. [Google Scholar] [CrossRef]
- Leong, Y.K.; Show, P.L.; Ooi, C.W.; Ling, T.C.; Lan, J.C. Current trends in polyhydroxyalkanoates (PHAs) biosynthesis: Insights from the recombinant Escherichia coli. J. Biotechnol. 2014, 180, 52–65. [Google Scholar] [CrossRef]
- Kumar, A.; Srivastava, J.K.; Mallick, N.; Singh, A.K. Commercialization of bacterial cell factories for the sustainable production of polyhydroxyalkanoate thermoplastics: Progress and prospects. Recent Patents Biotechnol. 2015, 9, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Anthony, R.J.; Sathish, A.; Sims, R.C.; Miller, C.D. Effects of wastewater microalgae harvesting methods on polyhydroxybutyrate production. Bioresour. Technol. 2014, 156, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Putman, R.J.; Inan, K.; Sal, F.A.; Sathish, A.; Smith, T.; Nielsen, C.; Sims, R.C.; Miller, C.D. Polyhydroxybutyrate production using a wastewater microalgae based media. Algal Res. 2015, 8, 95–98. [Google Scholar] [CrossRef]
- Shiloach, J.; Fass, R. Growing E. coli to high cell density—A historical perspective on method development. Biotechnol. Adv. 2005, 23, 345–357. [Google Scholar] [CrossRef]
- De Almeida, A.; Giordano, A.M.; Nikel, P.I.; Pettinari, M.J. Effects of aeration on the synthesis of poly(3-hydroxybutyrate) from glycerol and glucose in recombinant Escherichia coli. Appl. Environ.Microbiol. 2010, 76, 2036–2040. [Google Scholar] [CrossRef] [PubMed]
- Inan, K.; Sal, F.A.; Rahman, A.; Putman, R.J.; Agblevor, F.A.; Miller, C.D. Microbubble assisted polyhydroxybutyrate production in Escherichia coli. BMC Res. Notes 2016, 9, 338. [Google Scholar] [CrossRef]
- Chanprateep, S. Current trends in biodegradable polyhydroxyalkanoates. J. Biosci. Bioeng. 2010, 110, 621–632. [Google Scholar] [CrossRef]
- Sreedevi, S.; Unni, K.; Sajith, S.; Priji, P.; Josh, M.; Benjamin, S. Bioplastics: Advances in polyhydroxybutyrate research. In Advances in Polymer Science; Springer: Berlin, Germany, 2015; pp. 1–30. [Google Scholar]
- Jacquel, N.; Lo, C.-W.; Wei, Y.-H.; Wu, H.-S.; Wang, S.S. Isolation and purification of bacterial poly(3-hydroxyalkanoates). Biochem. Eng. J. 2008, 39, 15–27. [Google Scholar] [CrossRef]
- Penloglou, G.; Chatzidoukas, C.; Kiparissides, C. Microbial production of polyhydroxybutyrate with tailor-made properties: An integrated modelling approach and experimental validation. Biotechnol. Adv. 2012, 30, 329–337. [Google Scholar] [CrossRef]
- Ahmed, A.; Rushworth, J.V.; Hirst, N.A.; Millner, P.A. Biosensors for whole-cell bacterial detection. Clin. Microbiol. Rev. 2014, 27, 631–646. [Google Scholar] [CrossRef]
- Cimaglia, F.; de Lorenzis, E.; Mezzolla, V.; Rossi, F.; Poltronieri, P. Detection of L. monocytogenes in enrichment cultures by immunoseparation and immunosensors. IEEE Sens. 2016, 16, 7045–7052. [Google Scholar] [CrossRef]
- Poltronieri, P.; Mezzolla, V.; Primiceri, E.; Maruccio, G. Biosensors for detection of food pathogens. Foods 2014, 3, 511–526. [Google Scholar] [CrossRef] [PubMed]
- Poltronieri, P.; Cimaglia, F.; de Lorenzis, E.; Chiesa, M.; Mezzolla, V.; Reca, I.B. Protein chips for detection of Salmonella spp. from enrichment culture. Sensors 2016, 16, 574. [Google Scholar] [CrossRef]
- Almaviva, S.; Palucci, A.; Botti, S.; Puiu, A.; Rufoloni, A. Validation of a miniaturized spectrometer for trace detection of explosives by Surface-Enhanced Raman Spectroscopy. Challenges 2016, 7, 14. [Google Scholar] [CrossRef]
- Spiekermann, P.; Rehm, B.H.A.; Kalscheuer, R.; Baumeister, D.; Steinbüchel, A. A sensitive, viable-colony staining method using Nile red for direct screening of bacteria that accumulate polyhydroxyalkanoic acids and other lipid storage compounds. Arch. Microbiol. 1999, 171, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, S.; Davidson, A.; Yang, B.; Wang, Q.; Lin, Q. A MEMS viscometric sensor for continuous glucose monitoring. J. Micromech. Microeng. 2007, 17, 2528–2537. [Google Scholar] [CrossRef]
- Ghoshdastider, U.; Wu, R.; Trzaskowski, B.; Mlynarczyk, K.; Miszta, P.; Gurusaran, M.; Viswanathan, S.; Renugopalakrishnan, V.; Filipek, S. Nano-encapsulation of glucose oxidase dimer by graphene. RSC Adv. 2015, 5, 13570–13578. [Google Scholar] [CrossRef]
- Heinrich, D.; Madkour, M.H.; Al-Ghamdi, M.A.; Shabbaj, I.I.; Steinbüchel, A. Large scale extraction of poly(3-hydroxybutyrate) from Ralstonia eutropha H16 using sodium hypochlorite. AMB Express 2012, 2, 59. [Google Scholar] [CrossRef]
- Naheed, N.; Jamil, N. Optimization of biodegradable plastic production on sugar cane molasses in Enterobacter sp. SEL2. Braz. J. Microbiol. 2014, 45, 417–426. [Google Scholar] [CrossRef]
- Lin, L.L.; Hsu, W. Lactose-induced expression of Bacillus sp. TS-23 amylase gene in E. coli regulated by a T7 promoter. Lett. Appl. Microbiol. 1997, 24, 365–368. [Google Scholar] [CrossRef]
- Li, M.; Wang, J.; Geng, Y.; Li, Y.; Wang, Q.; Liang, Q.; Qi, Q. A strategy of gene overexpression based on tandem repetitive promoters in Escherichia coli. Microb. Cell. Fact. 2012, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Lee, Y. Metabolic engineering of Escherichia coli for production of enantiomerically pure (R)-(−)-hydroxycarboxylic acids. Appl. Environ. Microbiol. 2003, 69, 3421–3426. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Wang, H.; Fu, X.Z.; Gao, X.; Wu, Q.; Chen, G.Q. Effects of chromosomal gene copy number and locations on polyhydroxyalkanoate synthesis by Escherichia coli and Halomonas sp. Appl. Microbiol. Biotechnol. 2015, 99, 5523–5534. [Google Scholar] [CrossRef] [PubMed]
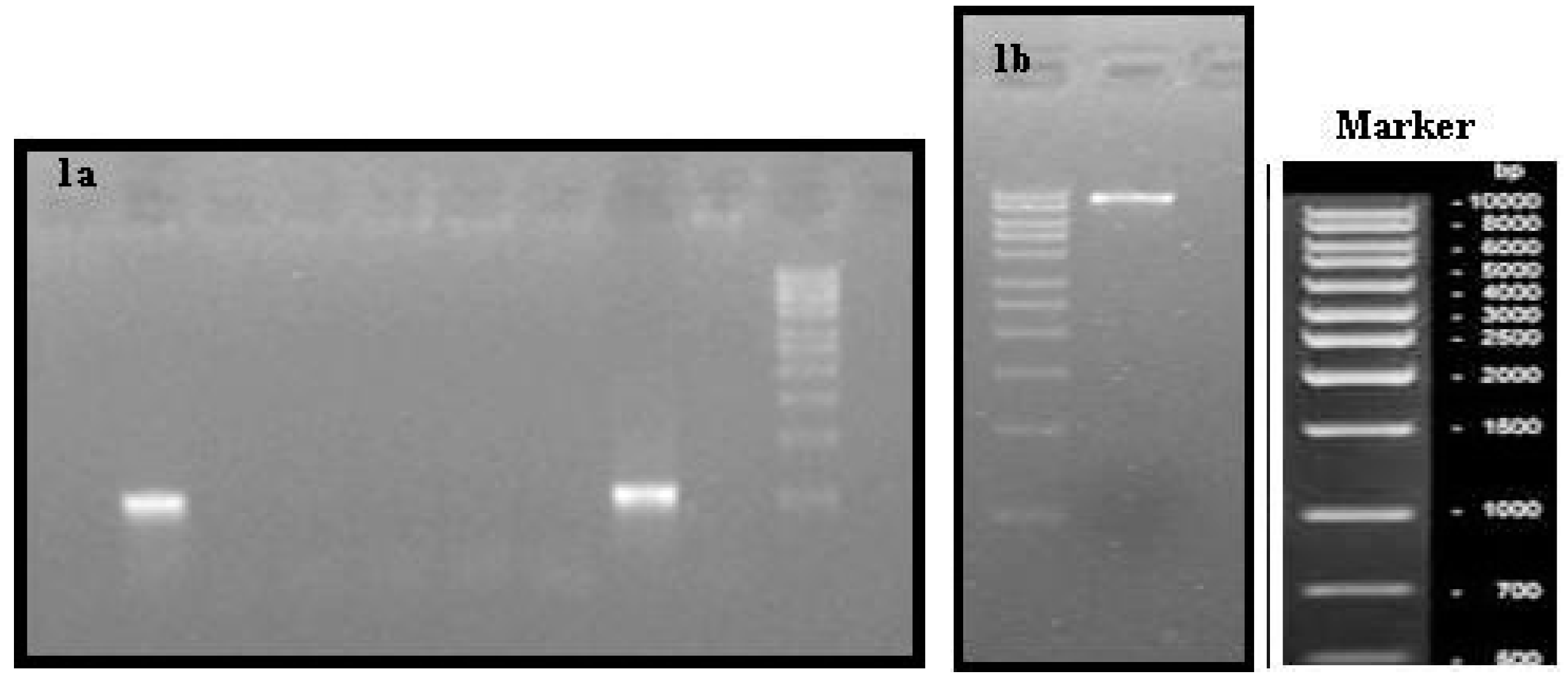
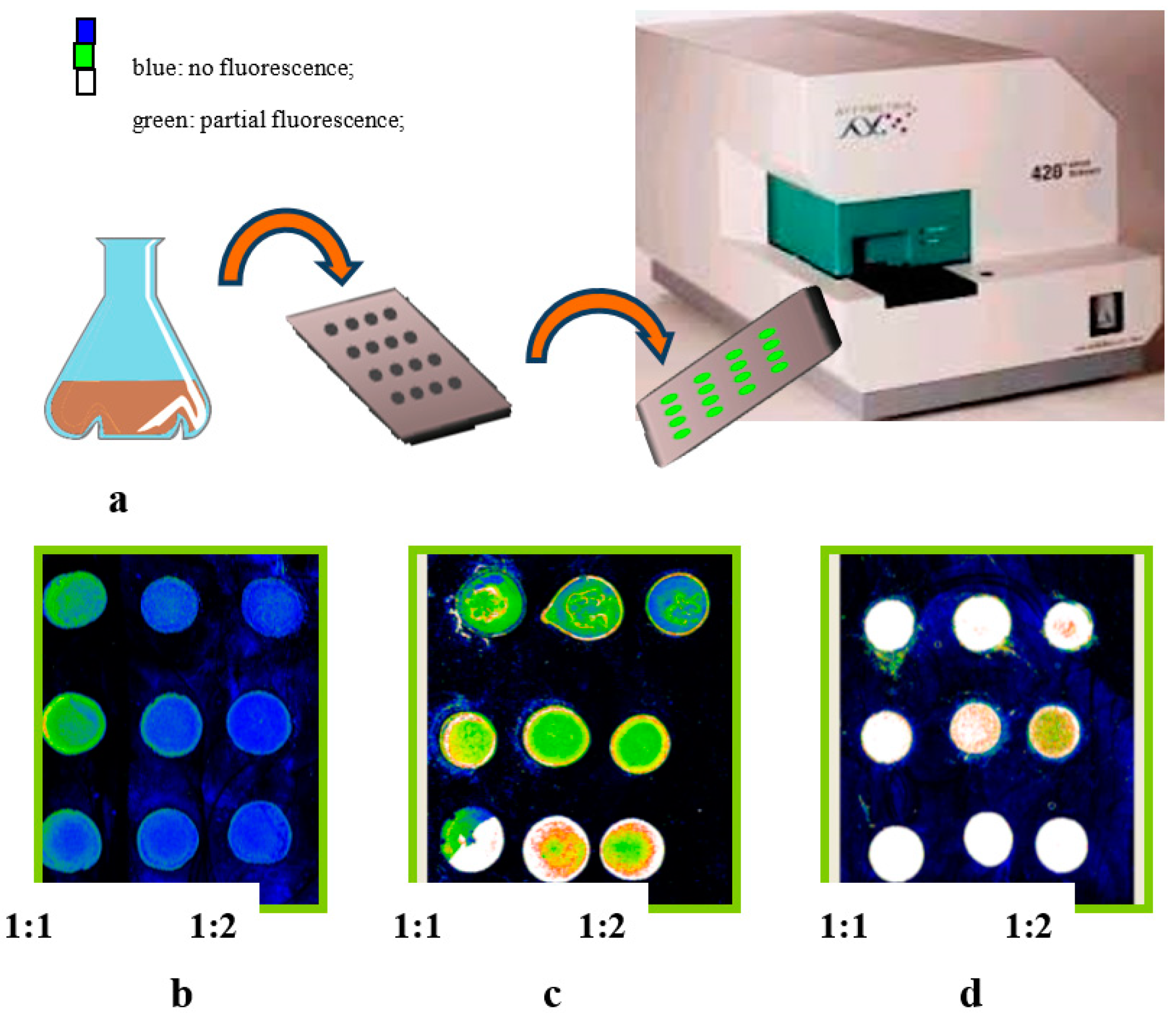


{kind=link}
{kind=link}
{kind=link}
| Composition | Sweet Corn (g/L) | Potato Skin (g/L) | Banana Juice (g/L) |
|---|---|---|---|
| Glucose | 104 | 45 | 90 |
| Fructose | 10 | ≤5 | 82 |
| Maltose | ≤5 | 10 | nd |
| Sucrose | ≤5 | ≤5 | nd |
| Nitrogen | 3.6 | 4 | 6 |
| Citric acid | nd | nd | 3.5 |
| Magnesium | nd | nd | 0.3 |
| Phosphate | nd | nd | 0.25 |
| Fed-Batch Hydrolysate | CDW(g L−1) | PHB (g L−1) |
|---|---|---|
| Sweet Corn Enzymatic hydrolysate 10%: | ||
| Recombinant BL21(DE3) E.coli | 8.2 | 1.7 |
| Empty BL21(DE3) E.coli | 7.9 | / |
| Ralstonia taiwanensis | 4.3 | / |
| Potato Skin Enzymatic hydrolysate 25%: | ||
| Recombinant BL21(DE3)E. coli | 15.3 | 2.9 |
| Empty BL21(DE3) E. coli | 14.9 | / |
| Ralstonia taiwanensis | 9.5 | 0.4 |
| Banana juice 5% hydrolysate: | ||
| Recombinant BL21(DE3) E. coli | 20.6 | 3.9 |
| Empty BL21(DE3) E. coli | 20.2 | / |
| Ralstonia taiwanensis | 15.8 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poltronieri, P.; Mezzolla, V.; D’Urso, O.F. PHB Production in Biofermentors Assisted through Biosensor Applications. Proceedings 2017, 1, 4. https://doi.org/10.3390/ecsa-3-E014
Poltronieri P, Mezzolla V, D’Urso OF. PHB Production in Biofermentors Assisted through Biosensor Applications. Proceedings. 2017; 1(2):4. https://doi.org/10.3390/ecsa-3-E014
Chicago/Turabian StylePoltronieri, Palmiro, Valeria Mezzolla, and Oscar Fernando D’Urso. 2017. "PHB Production in Biofermentors Assisted through Biosensor Applications" Proceedings 1, no. 2: 4. https://doi.org/10.3390/ecsa-3-E014
APA StylePoltronieri, P., Mezzolla, V., & D’Urso, O. F. (2017). PHB Production in Biofermentors Assisted through Biosensor Applications. Proceedings, 1(2), 4. https://doi.org/10.3390/ecsa-3-E014