Evaluation of a Virus Neutralisation Test for Detection of Rift Valley Fever Antibodies in Suid Sera




Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Animal Inoculations
2.3. Sera
2.4. Additional Performance Measures
2.5. Serological Tests
2.5.1. ELISA
2.5.2. Virus Neutralisation Test
2.6. Statistics
3. Results
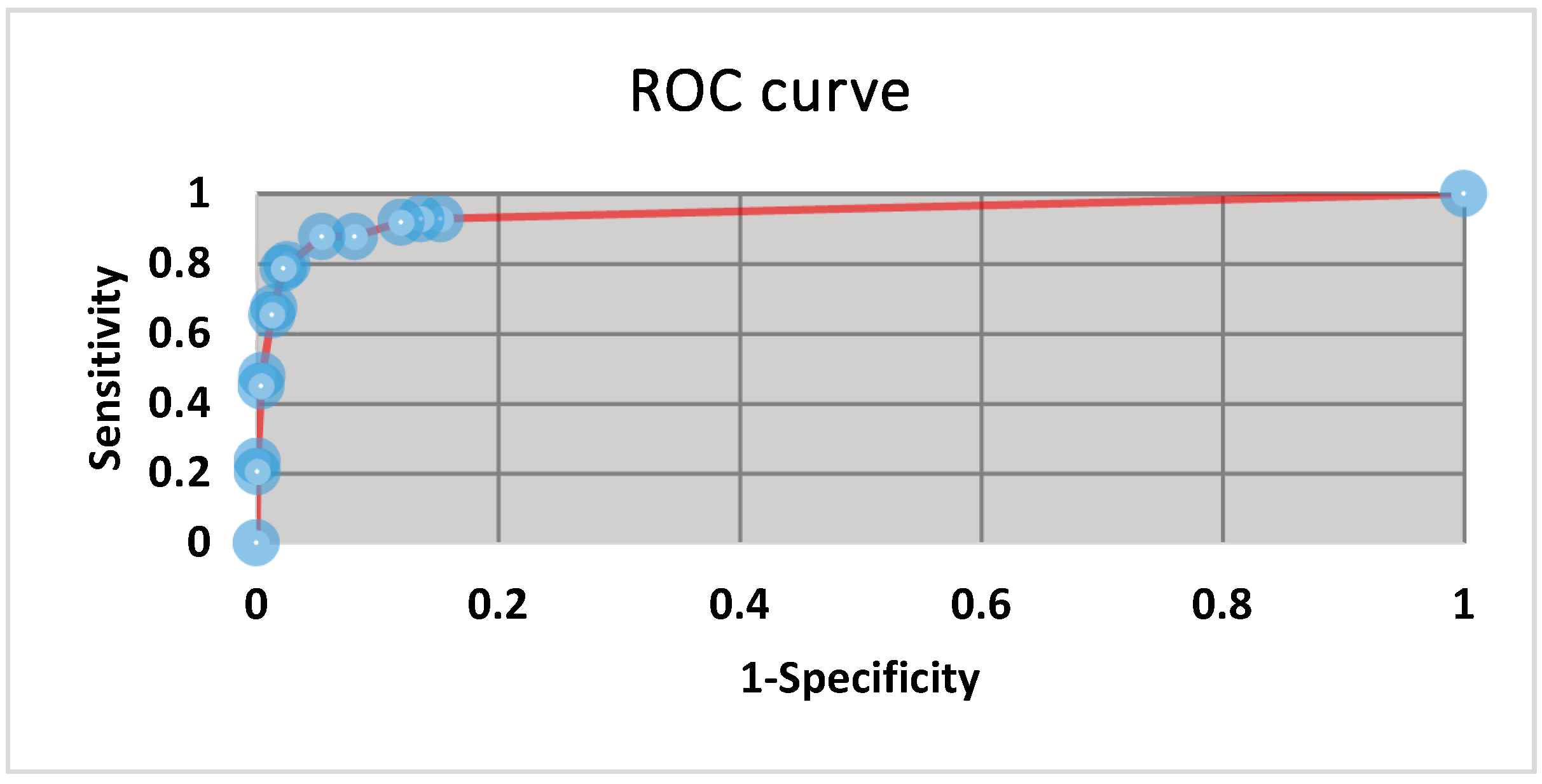
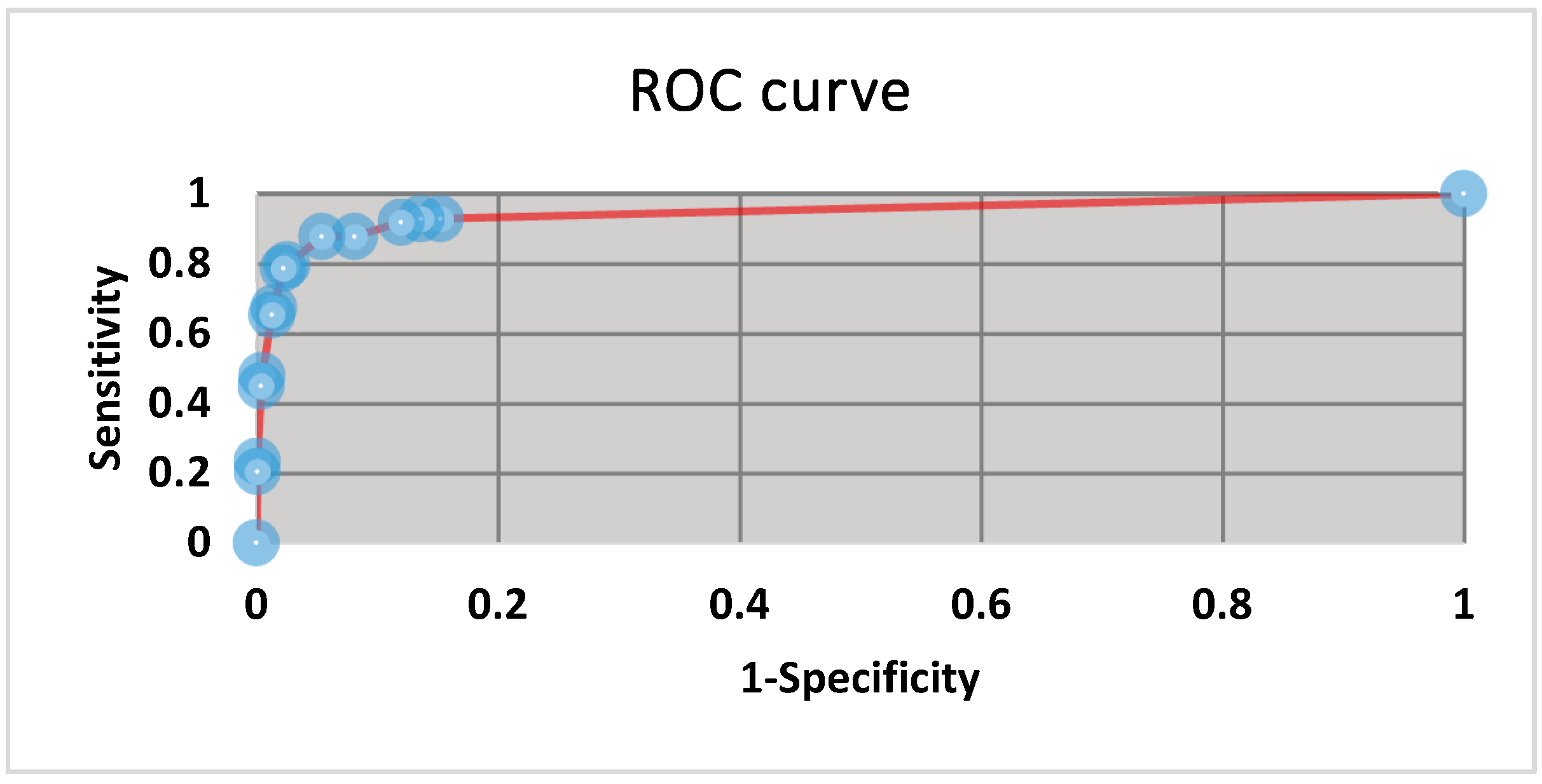
3.1. Operating Range and Thresholds
3.2. Analytical Sensitivity and Specificity
3.3. Diagnostic Sensitivity and Specificity
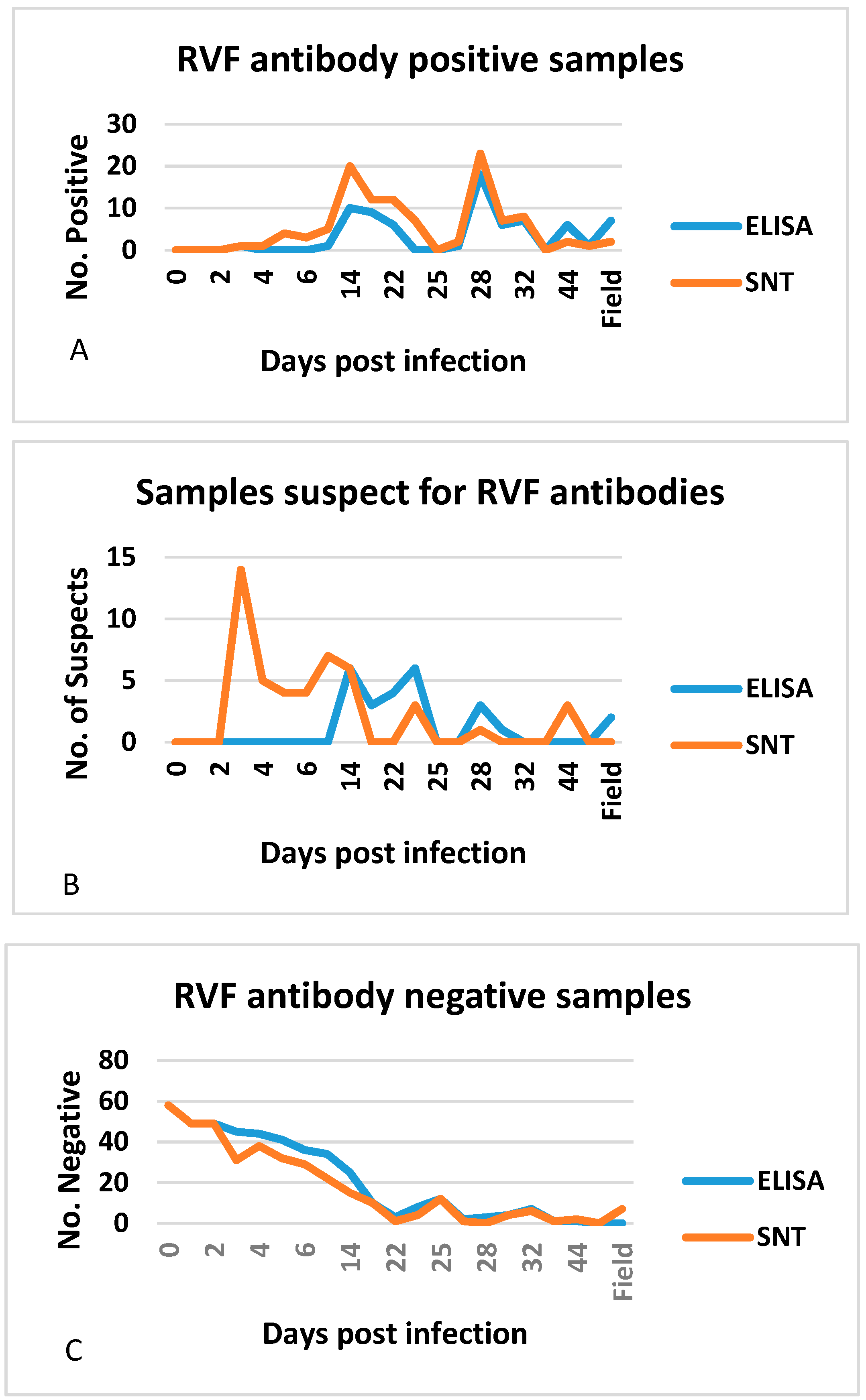
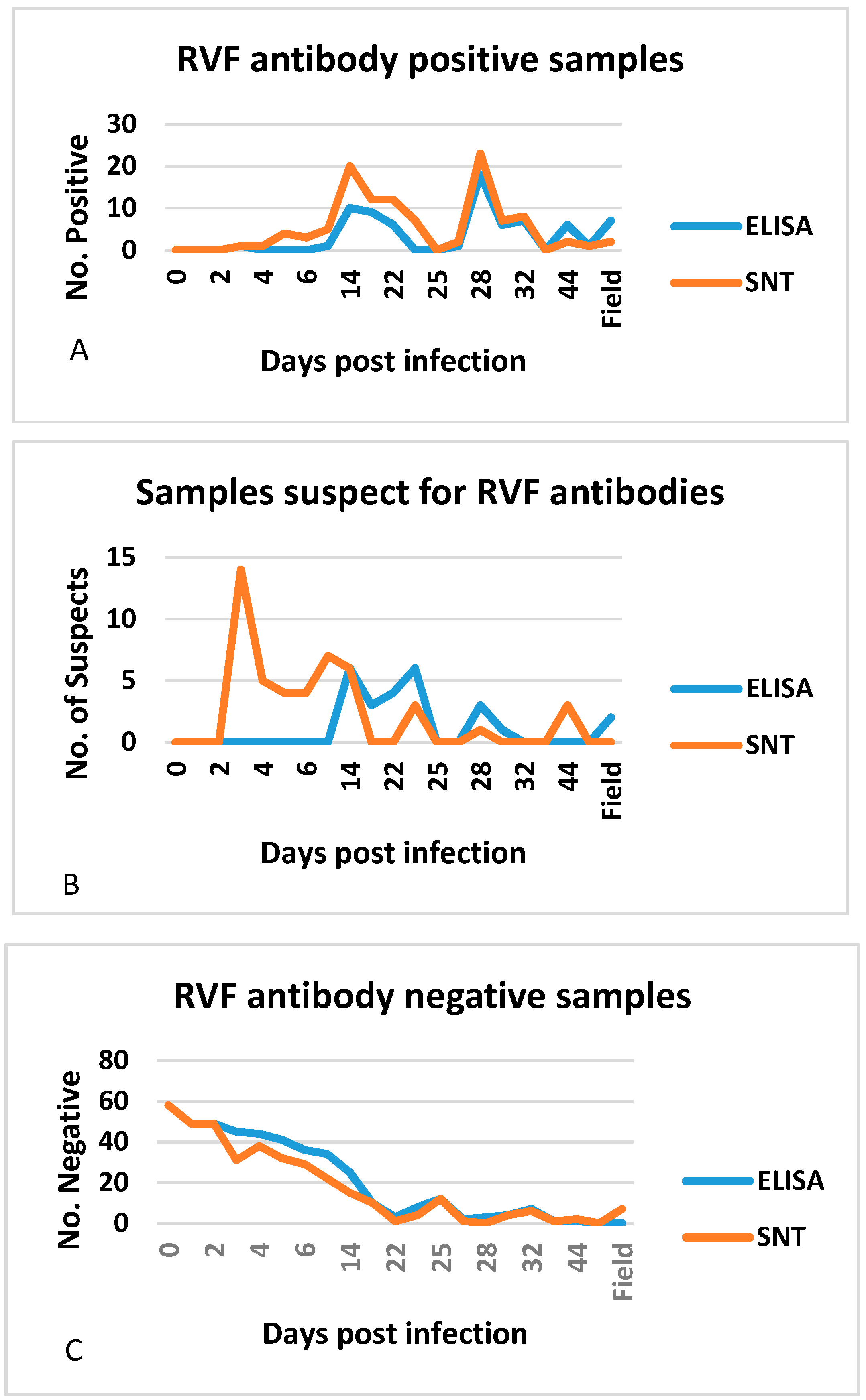
3.4. Additional Performance Measures
3.5. Statistics and Fitness for Purpose
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Daubney, R.; Hudson, J.R. Enzootic hepatitis or Rift Valley fever: An undescribed virus disease of sheep cattle and man from east Africa. J. Pathol. Bacteriol. 1931, 34, 545–579. [Google Scholar] [CrossRef]
- Ikegami, T.; Makino, S. The pathogenesis of Rift Valley fever. Viruses 2011, 3, 493–519. [Google Scholar] [CrossRef] [PubMed]
- Maes, P.; Alkhovsky, S.V.; Bào, Y.; Beer, M.; Birkhead, M.; Briese, T.; Buchmeier, M.J.; Calisher, C.H.; Charrel, R.N.; Choi, I.R.; et al. Taxonomy of the family Arenaviridae and the order Bunyavirales: Update 2018. Arch. Virol. 2018, 163, 2295–2310. [Google Scholar] [CrossRef] [PubMed]
- Coetzer, J.A. Brain teratology as a result of transplacental virus infection in ruminants. J. S. Afr. Vet. Assoc. 1980, 51, 153–157. [Google Scholar] [PubMed]
- Arishi, H.M.; Aqeel, A.Y.; Al Hazmi, MM. Vertical transmission of fatal Rift Valley fever in a newborn. Ann. Trop. Paediatr. 2006, 26, 251–253. [Google Scholar] [CrossRef]
- Adam, I.; Karsany, M.S. Case report: Rift Valley Fever with vertical transmission in a pregnant Sudanese woman. J. Med. Virol. 2008, 80, 929. [Google Scholar] [CrossRef] [PubMed]
- Balenghien, T.; Cardinale, E.; Chevalier, V.; Elissa, N.; Failloux, AB.; Nipomichene, T.N.J.J.; Nicolas, G.; Rakotoharinome, V.M.M.; Zumbo, B. Towards a better understanding of Rift Valley fever epidemiology in the south-west of the Indian Ocean. Vet. Res. 2013, 44, 78. [Google Scholar] [CrossRef]
- Linthicum, K.J.; Davies, F.G.; Kairo, A.; Bailey, C.L. Rift Valley fever virus (family Bunyaviridae, genus Phlebovirus): Isolations from Diptera collected during an inter-epizootic period in Kenya. J. Hyg. 1985, 95, 197–209. [Google Scholar] [CrossRef]
- Lumley, S.; Horton, D.L.; Hernandez-Triana, L.L.M.; Johnson, N.; Fooks, A.R.; Hewson, R. Rift Valley fever virus: Strategies for maintenance, survival and vertical transmission in mosquitoes. J. Gen. Virol. 2017, 98, 875–887. [Google Scholar] [CrossRef]
- Mbotha, D.; Bett, B.; Kairu-Wanyoike, S.; Grace, D.; Kihara, A.; Wainaina, M.; Hoppenheit, A.; Clausen, P.H.; Lindahl, J. Inter-epidemic Rift Valley fever virus seroconversions in an irrigation scheme in Bura, south-east Kenya. Transbound. Emerg. Dis. 2018, 65, e55–e62. [Google Scholar] [CrossRef]
- Rift Valley Fever (Infection with Rift Valley Fever Virus). Available online: http://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/2.01.18_RVF.pdf (accessed on 12 December 2018).
- Williams, R.; Ellis, C.E.; Smith, S.J.; Potgieter, C.A.; Wallace, D.; Mareledwane, V.E.; Majiwa, P.A. Validation of an IgM antibody capture ELISA based on a recombinant nucleoprotein for identification of domestic ruminants infected with Rift Valley fever virus. J. Virol. Methods 2011, 177, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Kortekaas, J.; Kant, J.; Vloet, R.; Cêtre-Sossah, C.; Marianneau, P.; Lacote, S.; Banyard, A.C.; Jeffries, C.; Eiden, M.; Groschup, M.; et al. European ring trial to evaluate ELISAs for the diagnosis of infection with Rift Valley fever virus. J. Virol. Methods 2013, 187, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Mroz, C.; Gwida, M.; El-Ashker, M.; El-Diasty, M.; El-Beskawy, M.; Ziegler, U.; Eiden, M.; Groschup, M.H. Seroprevalence of Rift Valley fever virus in livestock during inter-epidemic period in Egypt, 2014/15. BMC Vet. Res. 2017, 13, 87. [Google Scholar] [CrossRef]
- GraphPad Prism Version 8.0.1 (244). Available online: //www.graphpad.com/scientific-software/prism/ (accessed on 9 December 2018).
- Baveja, C.P.; Aggarwa, L.P. Statistical analysis of microbiological diagnostic tests. Indian J. Med. Microbiol. 2017, 35, 184–193. [Google Scholar] [PubMed]
- Simple ROC Curve Analysis. Available online: http://vassarstats.net/roc1.html (accessed on 9 December 2018).
- MedCalc Easy—To—Use Statistical Software Version 18.11. Available online: https://www.medcalc.org/calc/ (accessed on 19 December 2018).
- Baer, A.; Kehn-Hall, K. Viral concentration determination through plaque assays: Using traditional and novel overlay systems. J. Vis. Exp. 2014, 93, 52065. [Google Scholar] [CrossRef]
- Di Gennaro, A.; Lorusso, A.; Casaccia, C.; Conte, A.; Monaco, F.; Savini, G. Serum neutralization assay can efficiently replace plaque reduction neutralization test for detection and quantitation of West Nile virus antibodies in human and animal serum samples. Clin. Vaccine Immunol. 2014, 21, 1460–1462. [Google Scholar] [CrossRef] [PubMed]
- Monaco, F.; Cosseddu, G.M.; Doumbia, B.; Madani, H.; El Mellouli, F.; Jiménez-Clavero, M.A.; Sghaier, S.; Marianneau, P.; Cetre-Sossah, C.; Polci, A.; et al. First external quality assessment of molecular and serological detection of Rift Valley fever in the Western Mediterranean region. PLoS ONE 2015, 10, e0142129. [Google Scholar] [CrossRef]
- Métras, R.; Dommergues, L.; Ortiz, K.; Annequin, M.; Schuler, C.; Roux, P.; Edmunds, J.W.; Keeling, M.J.; Cêtre-Sossah, C.; Cardinale, E. Absence of evidence of Rift Valley fever infection in Eulemur fulvus (Brown Lemur) in Mayotte during an interepidemic period. Vector Borne Zoonotic Dis. 2017, 17, 358–360. [Google Scholar] [CrossRef]
- Scott, G.R. Pigs and Rift Valley Fever. Nature 1963, 200, 919–920. [Google Scholar] [CrossRef] [PubMed]
- Lubisi, B.A.; Mutowembwa, P.B.; Ndouvhada, P.N.; Odendaal, L.; Clift, S.; Bastos, A.D.S.; Penrith, M.-L. Experimental infection of pregnant sows with Rift Valley fever virus. 2019; in prep. [Google Scholar]
- Pepin, M.; Bouloy, M.; Bird, B.H.; Kemp, A.; Paweska, J. Rift Valley fever virus (Bunyaviridae: Phlebovirus): An update on pathogenesis, molecular epidemiology, vectors, diagnostics and prevention. Vet. Res. 2010, 41, 61. [Google Scholar] [CrossRef] [PubMed]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Purpose | Animals | Source | Antibody Status | Days Post Infection | Number Animals: Number of Samples |
|---|---|---|---|---|---|
| Analytical specificity | Sows | Experimental | Negative | Pre-infection | 11:11 |
| Weaners | Experimental | Negative | Pre-infection | 27:27 | |
| Suckling piglets | Experimental | Negative | Pre-infection | 20:20 | |
| Total | 58:58 | ||||
| Analytical sensitivity | Weaners | Experimental: Infected with M21/10 RVFV | Positive | 3, 7, 14, 21 and 28 | 5:5 |
| Newborn piglets | Experimental: Dams infected with M21/10 and M259/09 RVFV | Positive | 22, 28, 32, and 44 | 4:4 | |
| Sow | Experimental: Infected with M259/09 RVFV | Positive | 61 | 1:1 | |
| Total | 10:10 | ||||
| Diagnostic sensitivity | Suckling piglets | Experimental: Infected with M21/10 and 259/09 RVFV | Positive | 14 | 2:2 |
| Newborn piglets | Experimental: Dams infected with M21/10 and M259/09 RVFV | Positive | 22, 23, 28, 32, 44 | 41:41 | |
| Weaners | Experimental: Infected with M21/10 and 259/09 RVFV | Positive | 3, 7, 14, 21, 27, 28, 30 | 16:30 | |
| Sows | Experimental: Infected with M21/10 and 259/09 RVFV | Positive | 14, 21, 27, 28, 61 | 7:16 | |
| Porcine: various ages | Field | Positive | N/A | 6:6 | |
| Warthogs: Mixed ages | Field | Positive | N/A | 3:3 | |
| Total | 75:98 | ||||
| Diagnostic specificity | Suckling piglets | Experimental: Infected with M21/10 and 259/09 RVFV | Negative | 0– 7, 14, 21 | 20:132 |
| Newborn piglets | Experimental: Dams infected with M21/10 and M259/09 RVFV | Negative | 22, 23, 25, 32, 44 | 29:29 | |
| Weaners | Experimental: Infected with M21/10 and 259/09 RVFV | Negative | 0–7, 14, 21, 28, 30 | 27:167 | |
| Sows | Experimental: Infected with M21/10 and 259/09 RVFV | Negative | 0–7; 14, 21, 22, 27, 34 | 12:104 | |
| Porcine: various ages | Field | Negative | N/A | 725:725 | |
| Warthogs: Mixed ages | Field | Negative | N/A | 97:97 | |
| Total | 910:1254 | ||||
| Grand Total | 985:1352 | ||||
| - | - | ELISA | - | ||
|---|---|---|---|---|---|
| Positive | Suspect | Negative | |||
| VNT | Positive | 61 (4.5%) | 17 (1.3%) | 32 (2%) | 110 (8%) |
| Suspect | 6 (0.5%) | 2 (0.15%) | 39(3%) | 47 (3.5%) | |
| Negative | 6 (0.5%) | 6 (0.5%) | 1183 (88%) | 1195 (88.5%) | |
| 73 (5.5%) | 25 (2%) | 1254 (93%) | 1352 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lubisi, B.A.; Ndouvhada, P.N.; Neiffer, D.; Penrith, M.-L.; Sibanda, D.-R.; Bastos, A.D.S. Evaluation of a Virus Neutralisation Test for Detection of Rift Valley Fever Antibodies in Suid Sera. Trop. Med. Infect. Dis. 2019, 4, 52. https://doi.org/10.3390/tropicalmed4010052
Lubisi BA, Ndouvhada PN, Neiffer D, Penrith M-L, Sibanda D-R, Bastos ADS. Evaluation of a Virus Neutralisation Test for Detection of Rift Valley Fever Antibodies in Suid Sera. Tropical Medicine and Infectious Disease. 2019; 4(1):52. https://doi.org/10.3390/tropicalmed4010052
Chicago/Turabian StyleLubisi, Baratang A., Phumudzo N. Ndouvhada, Donald Neiffer, Mary-Louise Penrith, Donald-Ray Sibanda, and Armanda D.S. Bastos. 2019. "Evaluation of a Virus Neutralisation Test for Detection of Rift Valley Fever Antibodies in Suid Sera" Tropical Medicine and Infectious Disease 4, no. 1: 52. https://doi.org/10.3390/tropicalmed4010052
APA StyleLubisi, B. A., Ndouvhada, P. N., Neiffer, D., Penrith, M.-L., Sibanda, D.-R., & Bastos, A. D. S. (2019). Evaluation of a Virus Neutralisation Test for Detection of Rift Valley Fever Antibodies in Suid Sera. Tropical Medicine and Infectious Disease, 4(1), 52. https://doi.org/10.3390/tropicalmed4010052