Potential Intermediate Hosts for Coronavirus Transmission: No Evidence of Clade 2c Coronaviruses in Domestic Livestock from Ghana

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Selection
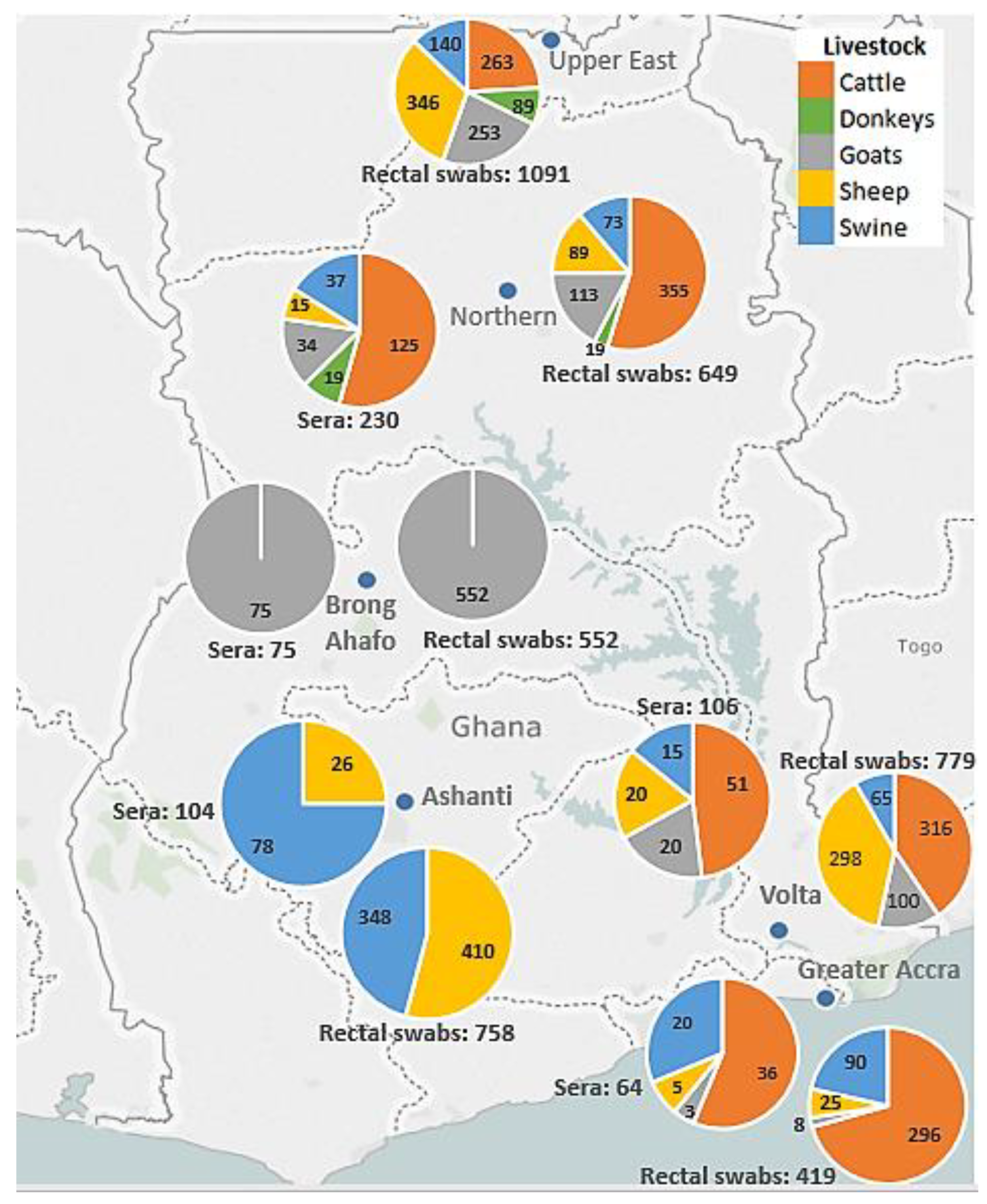
2.2. Sample and Data Collection
2.3. Ethical Issues
2.4. Viral RNA Extraction
2.5. PCR for Clade 2c Coronaviruses
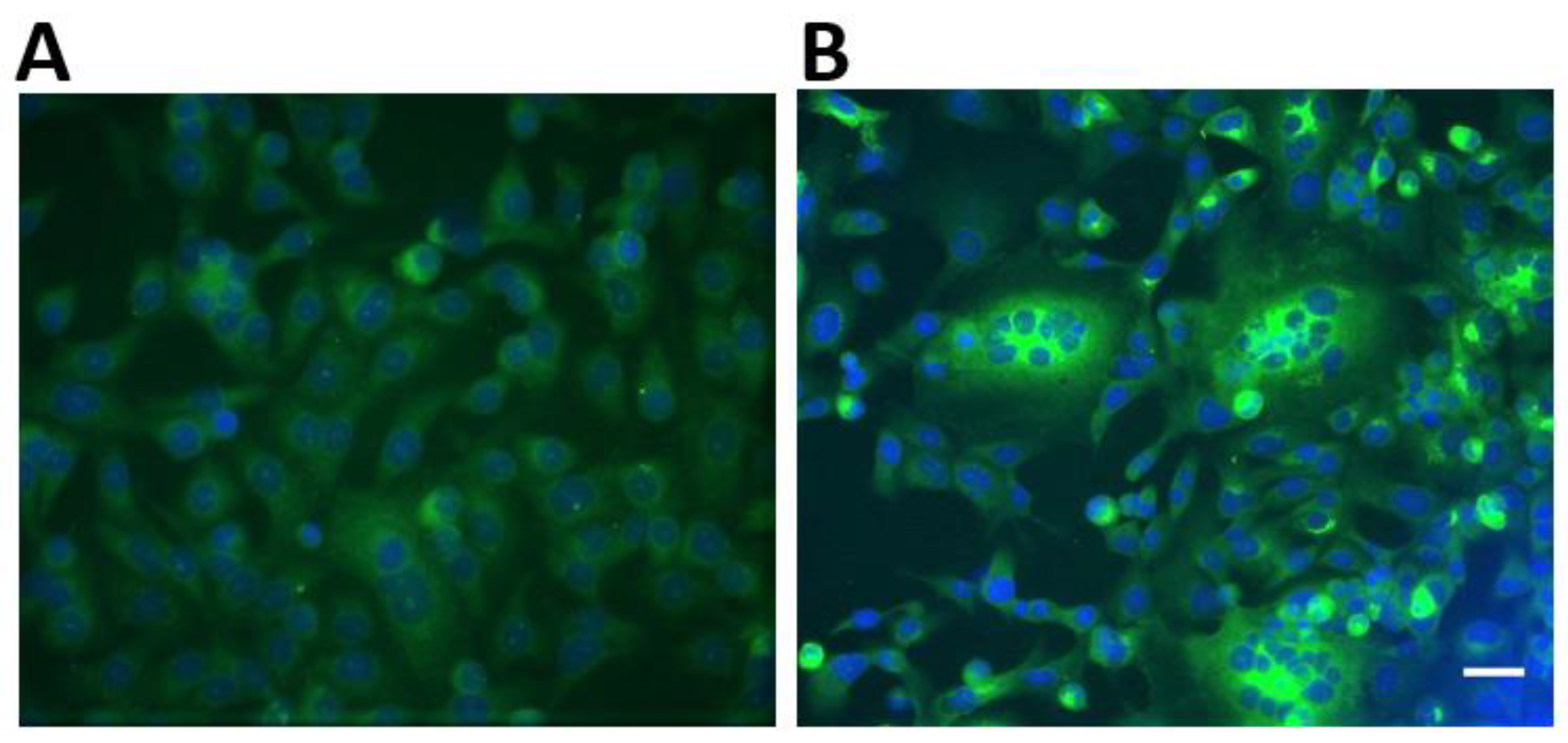
2.6. Recombinant Immunofluorescence Assay for Spike Protein of Clade 2c Coronaviruses
2.7. Determination of Minimum Detectable Seroprevalence
3. Results
3.1. Locations of Livestock and Bats
3.2. Possible Contact between Bats and Livestock
3.3. Laboratory Testing of Livestock Samples
3.4. Minimum Detectable Seroprevalence
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Woo, P.C.; Lau, S.K.; Huang, Y.; Yuen, K.-Y. Coronavirus diversity, phylogeny and interspecies jumping. Exp. Biol. Med. 2009, 234, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- De Groot, R.; Ziebuhr, J.; Poon, L.; Woo, P.; Talbot, P.; Rottier, P.; Holmes, K.; Baric, R.; Perlman, S.; Enjuanes, L. Taxonomic Proposal to the ICTV Executive Committee: Revision of the Family Coronaviridae; ICTV: Mountain View, CA, USA, 2008. [Google Scholar]
- Drosten, C.; Günther, S.; Preiser, W.; van der Werf, S.; Brodt, H.-R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, A.R.; et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Eng. J. Med. 2003, 348, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Zheng, B.; He, Y.; Liu, X.; Zhuang, Z.; Cheung, C.; Luo, S.; Li, P.; Zhang, L.; Guan, Y.; et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Field, H. Bats and emerging zoonoses: Henipaviruses and SARS. Zoonoses Public Health 2009, 56, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Zaki, M.A.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, A.R. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Eng. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Haagmans, L.B.; al Dhahiry, S.H.; Reusken, C.B.; Raj, V.S.; Galiano, M.; Myers, R.; Godeke, G.-J.; Jonges, M.; Farag, E.; Diab, A.; et al. Middle East respiratory syndrome coronavirus in dromedary camels: An outbreak investigation. Lancet Infect. Dis. 2014, 14, 140–145. [Google Scholar] [CrossRef]
- Azhar, I.E.; El-Kafrawy, S.A.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Hashem, A.M.; Madani, T.A. Evidence for camel-to-human transmission of MERS coronavirus. N. Eng. J. Med. 2014, 370, 2499–2505. [Google Scholar] [CrossRef] [PubMed]
- Memish, A.Z.; Mishra, N.; Olival, K.J.; Fagbo, S.F.; Kapoor, V.; Epstein, J.H.; AlHakeem, R.; al Asmari, M.; Islam, A.; Kapoor, A.; et al. Middle East respiratory syndrome coronavirus in bats, Saudi Arabia. Emerg. Infect. Dis. 2013, 19. [Google Scholar] [CrossRef] [PubMed]
- Alagaili, N.A.; Briese, T.; Mishra, N.; Kapoor, V.; Sameroff, S.C.; de Wit, E.; Munster, V.J.; Hensley, L.E.; Zalmout, I.S.; Kapoor, A.; et al. Middle East respiratory syndrome coronavirus infection in dromedary camels in Saudi Arabia. MBio 2014, 5, e00884-14. [Google Scholar] [CrossRef] [PubMed]
- Menachery, V.D.; Yount, B.L., Jr.; Debbink, K.; Agnihothram, S.; Gralinski, L.E.; Plante, J.A.; Graham, R.L.; Scobey, T.; Ge, X.-Y.; Donaldson, E.F.; et al. A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence. Nat. Med. 2015, 21, 1508–1513. [Google Scholar] [CrossRef] [PubMed]
- Pfefferle, S.; Oppong, S.; Drexler, J.F.; Gloza-Rausch, F.; Ipsen, A.; Seebens, A.; Muller, M.A.; Annan, A.; Vallo, P.; Adu-Sarkodie, Y.; et al. Distant relatives of severe acute respiratory syndrome coronavirus and close relatives of human coronavirus 229E in bats, Ghana. Emerg. Infect. Dis. 2009, 15, 1377–1384. [Google Scholar] [CrossRef]
- Corman, M.V.; Eckerle, I.; Memish, Z.A.; Liljander, A.M.; Dijkman, R.; Jonsdottir, H.; Ngeiywa, K.J.J.; Kamau, E.; Younan, M.; Al Masri, M.; et al. Link of a ubiquitous human coronavirus to dromedary camels. Proc. Natl. Acad. Sci. USA 2016, 113, 9864–9869. [Google Scholar] [CrossRef] [PubMed]
- Crossley, M.B.; Mock, R.E.; Callison, S.A.; Hietala, S.K. Identification and characterization of a novel alpaca respiratory coronavirus most closely related to the human coronavirus 229E. Viruses 2012, 4, 3689–3700. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Shi, M.; Chommanard, C.; Queen, K.; Zhang, J.; Markotter, W.; Kuzmin, I.V.; Holmes, E.C.; Tong, S. Surveillance of bat coronaviruses in Kenya identifies relatives of human coronaviruses NL63 and 229E and their recombination history. J. Virol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Drosten, C.; Meyer, B.; Müller, M.A.; Corman, V.M.; Al-Masri, M.; Hossain, R.; Madani, H.; Sieberg, A.; Bosch, B.J.; Lattwein, E.; et al. Transmission of MERS-coronavirus in household contacts. N. Eng. J. Med. 2014, 371, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Liljander, A.; Meyer, B.; Jores, J.; Müller, M.A.; Lattwein, E.; Njeru, I.; Bett, B.; Drosten, C.; Corman, V.M. MERS-CoV antibodies in humans, Africa, 2013–2014. Emerg. Infect. Dis. 2016, 22, 1086–1089. [Google Scholar] [CrossRef] [PubMed]
- Reusken, C.; Ababneh, M.; Raj, V.; Meyer, B.; Eljarah, A.; Abutarbush, S.; Godeke, G.; Bestebroer, T.; Zutt, I.; Müller, M.; et al. Middle East respiratory syndrome coronavirus (MERS-CoV) serology in major livestock species in an affected region in Jordan, June to September 2013. Euro Surveill. 2013, 12, 20662. [Google Scholar] [CrossRef]
- Hemida, M.; Perera, R.; Wang, P.; Alhammadi, M.; Siu, L.; Li, M.; Poon, L.; Saif, L.; Alnaeem, A.; Peiris, M. Middle East Respiratory Syndrome (MERS) coronavirus seroprevalence in domestic livestock in Saudi Arabia, 2010 to 2013. Euro Surveill. Bull. 2012, 18, 20659. [Google Scholar] [CrossRef]
- Meyer, B.; García-Bocanegra, I.; Wernery, U.; Wernery, R.; Sieberg, A.; Müller, M.A.; Drexler, J.F.; Drosten, C.; Eckerle, I. Serologic assessment of possibility for MERS-CoV infection in equids. Emerg. Infect. Dis. 2015, 21, 181–182. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.; Wang, P.; Gomaa, M.; El-Shesheny, R.; Kandeil, A.; Bagato, O.; Siu, L.; Shehata, M.; Kayed, A.; Moatasim, Y. Seroepidemiology for MERS coronavirus using microneutralisation and pseudoparticle virus neutralisation assays reveal a high prevalence of antibody in dromedary camels in Egypt, June 2013. Euro Surveill. 2013, 18, 20574. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.M.; Corman, V.M.; Jores, J.; Meyer, B.; Younan, M.; Liljander, A.; Bosch, B.J.; Lattwein, E.; Hilali, M.; Musa, B.E.; et al. MERS coronavirus neutralizing antibodies in camels, Eastern Africa, 1983–1997. Emerg. Infect. Dis. 2014, 20, 2093–2095. [Google Scholar] [CrossRef] [PubMed]
- Annan, A.; Baldwin, H.J.; Corman, V.M.; Klose, S.M.; Owusu, M.; Nkrumah, E.E.; Badu, E.K.; Anti, P.; Agbenyega, O.; Meyer, B.; et al. Human betacoronavirus 2c EMC/2012–related viruses in bats, Ghana and Europe. Emerg. Infect. Dis. 2013, 19, 456. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.; Müller, M.; Costabel, U.; Timm, J.; Binger, T.; Meyer, B.; Kreher, P.; Lattwein, E.; Eschbach-Bludau, M.; Nitsche, A.; et al. Assays for laboratory confirmation of novel human coronavirus (hCoV-EMC) infections. Eurosurveillance 2012, 17, 20334. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Müller, M.A.; Corman, V.M.; Reusken, C.B.; Ritz, D.; Godeke, G.J.; Lattwein, E.; Kallies, S.; Siemens, A.; van Beek, J.; et al. Antibodies against MERS coronavirus in dromedaries, United Arab Emirates, 2003 and 2013. Emerg. Infect. Dis. 2014, 20, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Binger, T.; Annan, A.; Drexler, J.F.; Müller, M.A.; Kallies, R.; Adankwah, E.; Wollny, R.; Kopp, A.; Heidemann, H.; Dei, D.; et al. A novel rhabdovirus isolated from the straw-colored fruit bat Eidolon helvum, with signs of antibodies in swine and humans. J. Virol. 2015, 89, 4588–4597. [Google Scholar] [CrossRef] [PubMed]
- Hoye, J.B.; Munster, V.J.; Nishiura, H.; Klaassen, M.; Fouchier, R.A. Surveillance of wild birds for avian influenza virus. Emerg. Infect. Dis. 2010, 16, 1827. [Google Scholar] [CrossRef] [PubMed]
- Venette, C.R.; Moon, R.D.; Hutchison, W.D. Strategies and statistics of sampling for rare individuals. Ann. Rev. Entomol. 2002, 47, 143–174. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.; Poon, L.; Gomaa, M.M.; Shehata, M.M.; Perera, R.; Zeid, D.A.; el Rifay, A.S.; Siu, L.Y.; Guan, Y.; Webby, R.J.; et al. MERS coronaviruses in dromedary camels, Egypt. Emerg. Infect. Dis. 2014, 20, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Drosten, C.; Seilmaier, M.; Corman, V.M.; Hartmann, W.; Scheible, G.; Sack, S.; Guggemos, W.; Kallies, R.; Muth, D.; Junglen, S.; et al. Clinical features and virological analysis of a case of Middle East. Respiratory syndrome coronavirus infection. Lancet Infect. Dis. 2013, 13, 745–751. [Google Scholar] [CrossRef]
- Cotton, M.; Watson, S.J.; Kellam, P.; Al-Rabeeah, A.A.; Makhdoom, H.Q.; Assiri, A.; Al-Tawfiq, J.A.; Alhakeem, R.F.; Madani, H.; AlRabiah, F.A.; et al. Transmission and evolution of the Middle East. respiratory syndrome coronavirus in Saudi Arabia: A descriptive genomic study. Lancet 2013, 382, 1993–2002. [Google Scholar] [CrossRef]
- Reusken, B.C.; Haagmans, B.L.; Müller, M.A.; Gutierrez, C.; Godeke, G.-J.; Meyer, B.; Muth, D.; Raj, V.S.; Smith-De Vries, L.; Corman, V.M.; et al. Middle East. respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: A comparative serological study. Lancet Infect. Dis. 2013, 13, 859–866. [Google Scholar] [CrossRef]
- Raj, S.V.; Mou, H.; Smits, S.L.; Dekkers, D.H.; Müller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.; Zaki, A.; Fouchier, R.A.; et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251. [Google Scholar] [CrossRef] [PubMed]
- Reusken, B.C.; Raj, V.S.; Koopmans, M.P.; Haagmans, B.L. Cross host transmission in the emergence of MERS coronavirus. Curr. Opini. Virol. 2016, 16, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Eckerle, I.; Corman, V.M.; Müller, M.A.; Lenk, M.; Ulrich, R.G.; Drosten, C. Replicative capacity of MERS coronavirus in livestock cell lines. Emerg. Infect. Dis. 2014, 20, 276. [Google Scholar] [CrossRef] [PubMed]
- Doremalen, V.N.; Miazgowicz, K.L.; Milne-Price, S.; Bushmaker, T.; Robertson, S.; Scott, D.; Kinne, J.; McLellan, J.S.; Zhu, J.; Munster, V.J. Host species restriction of Middle East. Respiratory syndrome coronavirus through its receptor, dipeptidyl peptidase 4. J. Virol. 2014, 88, 9220–9232. [Google Scholar] [CrossRef] [PubMed]
- Adney, R.D.; Brown, V.R.; Porter, S.M.; Bielefeldt-Ohmann, H.; Hartwig, A.E.; Bowen, R.A. Inoculation of goats, sheep, and horses with MERS-CoV DOES Not. Result in productive viral shedding. Viruses 2016, 8, 230. [Google Scholar] [CrossRef] [PubMed]
- Gortazar, C.; Reperant, L.A.; Kuiken, T.; de la Fuente, J.; Boadella, M.; Martínez-Lopez, B.; Ruiz-Fons, F.; Estrada-Peña, A.; Drosten, C.; Medley, G.; et al. Crossing the interspecies barrier: Opening the door to zoonotic pathogens. PLoS Pathog. 2014, 10, e1004129. [Google Scholar] [CrossRef] [PubMed]
- Mossoun, A.; Pauly, M.; Akoua-Koffi, C.; Couacy-Hymann, E.; Leendertz, S.A.J.; Anoh, A.E.; Gnoukpoho, A.H.; Leendertz, F.H.; Schubert, G. Contact to non-human primates and risk factors for zoonotic disease emergence in the Taï region, Côte d’Ivoire. EcoHealth 2015, 12, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Drosten, C.; Müller, M.A. Serological assays for emerging coronaviruses: Challenges and pitfalls. Virus Res. 2014, 194, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.T. Extra-limital records of the one-humped camel in west and central Africa. J. Camel Pract. Res. 2014, 21, 115–120. [Google Scholar] [CrossRef]
- Anti, P.; Owusu, M.; Agbenyega, O.; Annan, A.; Badu, E.K.; Nkrumah, E.E.; Tschapka, M.; Oppong, S.; Adu-Sarkodie, Y.; Drosten, C. Human–bat interactions in rural West. Africa. Emerg. Infect. Dis. 2015, 21, 1418. [Google Scholar] [CrossRef] [PubMed]

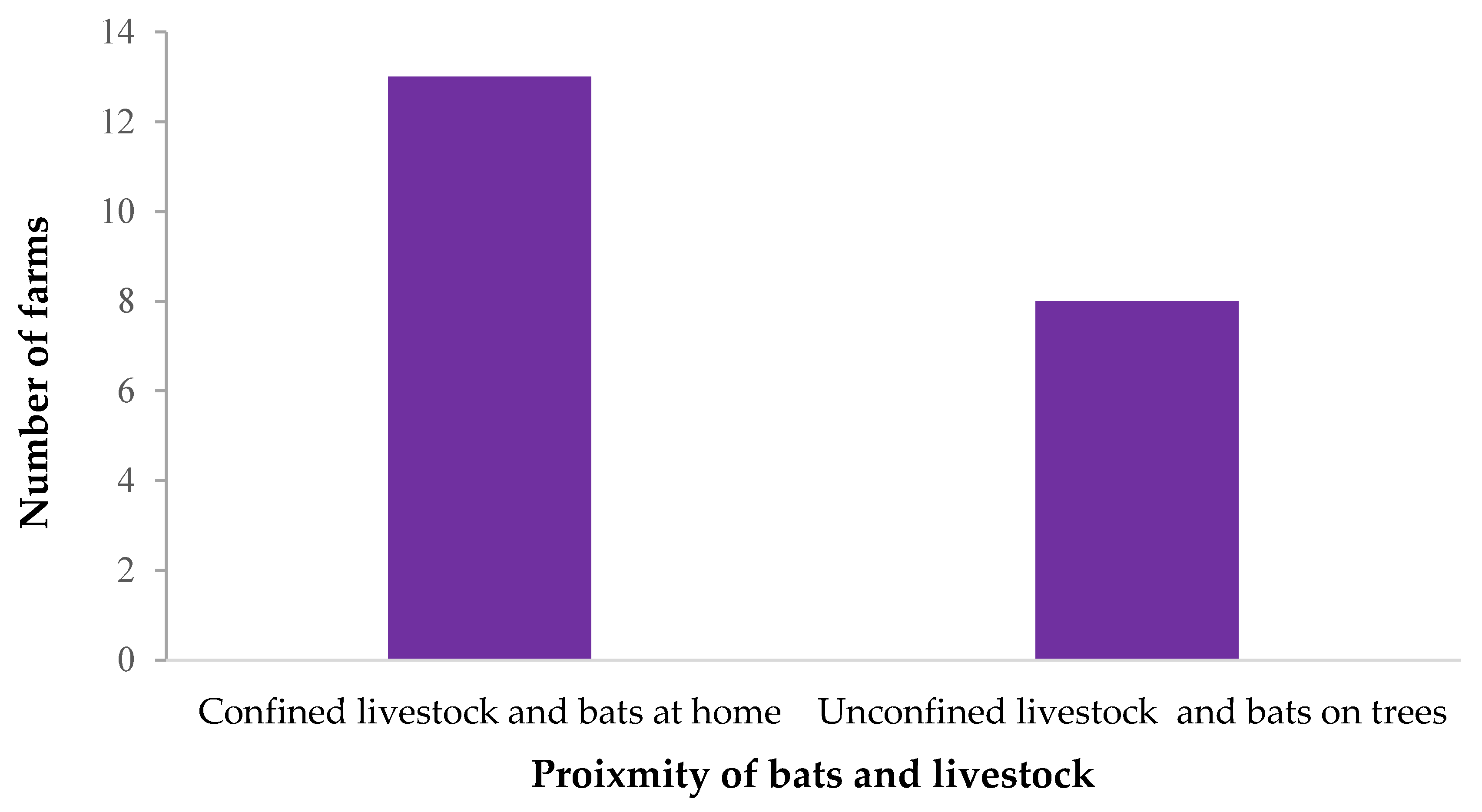

{kind=link}
{kind=link}
{kind=link}
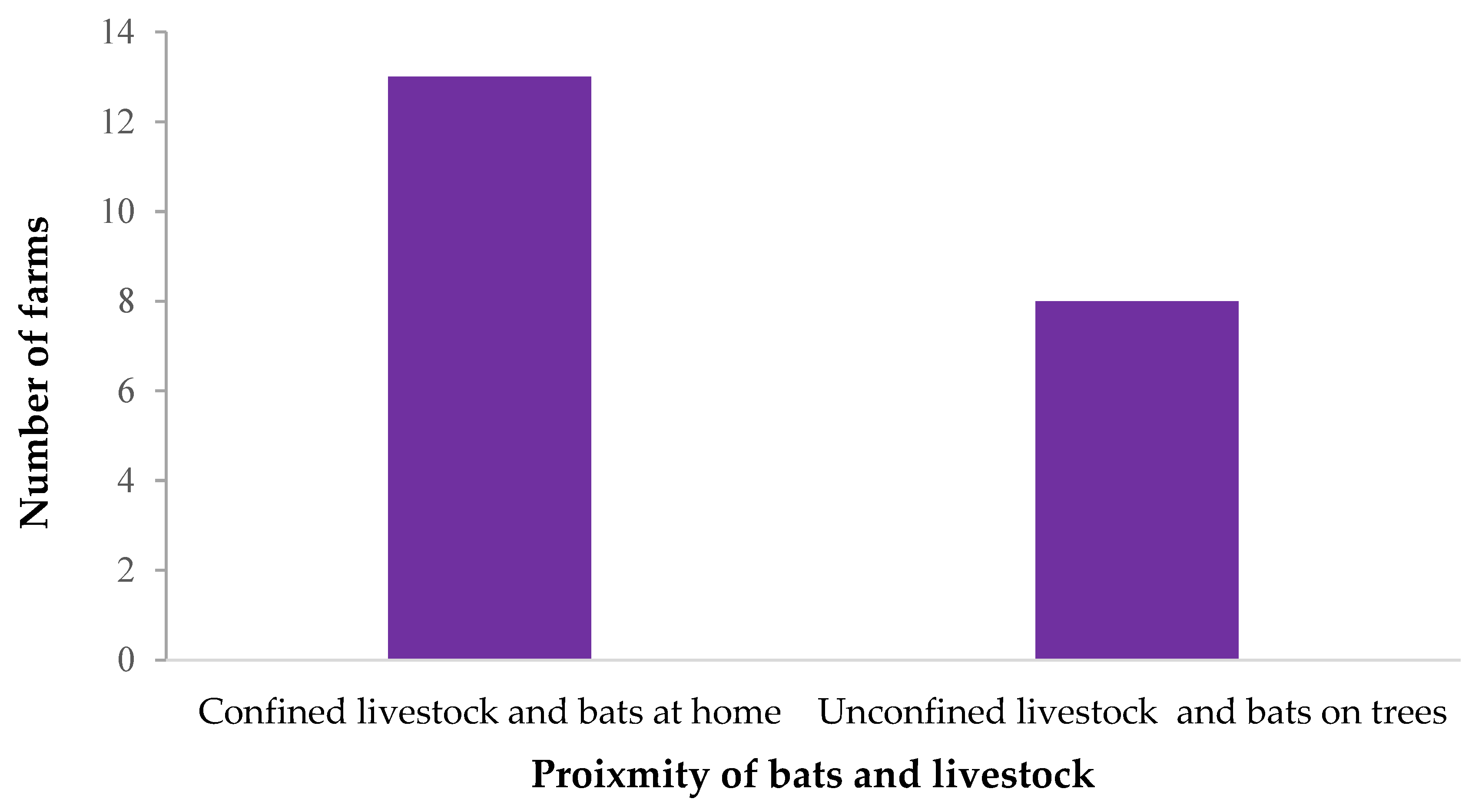{kind=link}
| Farms n = 35 | Number of Rectal Samples per Farm (Percentage) n = 4248 | Number of Serum Samples per Farm (Percentage) n = 592 |
|---|---|---|
| AC | 100 (2.4) | 20 (3.4) |
| AF | 77 (1.8) | 2 (0.3) |
| AGD | 618 (14.5) | 61 (10.3) |
| AGV | 132 (3.1) | 30 (5.1) |
| AGB | 52 (1.2) | - |
| AGO | 134 (3.2) | - |
| AKA | 24 (0.6) | 2 (0.3) |
| AKI | 70 (1.6) | 5 (0.8) |
| AKU | 148 (3.5) | 14 (2.4) |
| AA | 284 (6.7) | 56 (9.5) |
| AI | 193 (4.5) | 108 (18.2) |
| ANY ** | 70 (1.6) | - |
| ATY | 108 (2.5) | - |
| ATA | 85 (2) | - |
| AW | 1 (0) | - |
| AZ | 81 (1.9) | - |
| ESBS | 410 (9.7) | 26 (4.4) |
| IB | 50 (1.2) | 9 (1.5) |
| JO | 99 (2.3) | 10 (1.7) |
| KB | 33 (0.8) | 19 (3.2) |
| KGBS ** | 552 (13) | 75 (12.7) |
| KAS ** | 58 (1.4) | 58 (9.8) |
| KT | 51 (1.2) | - |
| KF | 290 (6.8) | 20 (3.4) |
| LB | 137 (3.2) | - |
| LF | 40 (0.9) | 10 (1.7) |
| MS ** | 28 (0.7) | - |
| NS | 108 (2.5) | - |
| NF | 26 (0.6) | 26 (4.4) |
| PT | 65 (1.5) | 4 (0.7) |
| SY | 23 (0.5) | 3 (0.5) |
| TH | 16 (0.4) | 15 (2.5) |
| WF | 45 (1.1) | 10 (1.7) |
| YA | 17 (0.4) | 3 (0.5) |
| ZM | 23 (0.5) | 6 (1) |
| Animal | Location | Reported on the farm (N = 31) |
|---|---|---|
| n (%) | ||
| Livestock | Only confined close to home (<10 m) | 14 (45.2) |
| Only confined away from home (>10 m) | 2 (6.5) | |
| Confined both close and away from home Unconfined | 4 (12.9) 11 (35.5) | |
| Bats | At home only | 0 (0) |
| In trees close to farm only At home and in trees close to the farm Neither at home or in trees close to the farm | 7 (22.6) 16 (51.6) 8 (25.8) | |
| In bat caves | 0 (0) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Duah, P.; Sylverken, A.; Owusu, M.; Yeboah, R.; Lamptey, J.; Oppong Frimpong, Y.; Burimuah, V.; Antwi, C.; Folitse, R.; Agbenyega, O.; et al. Potential Intermediate Hosts for Coronavirus Transmission: No Evidence of Clade 2c Coronaviruses in Domestic Livestock from Ghana. Trop. Med. Infect. Dis. 2019, 4, 34. https://doi.org/10.3390/tropicalmed4010034
El-Duah P, Sylverken A, Owusu M, Yeboah R, Lamptey J, Oppong Frimpong Y, Burimuah V, Antwi C, Folitse R, Agbenyega O, et al. Potential Intermediate Hosts for Coronavirus Transmission: No Evidence of Clade 2c Coronaviruses in Domestic Livestock from Ghana. Tropical Medicine and Infectious Disease. 2019; 4(1):34. https://doi.org/10.3390/tropicalmed4010034
Chicago/Turabian StyleEl-Duah, Philip, Augustina Sylverken, Michael Owusu, Richmond Yeboah, Jones Lamptey, Yaw Oppong Frimpong, Vitus Burimuah, Christopher Antwi, Raphael Folitse, Olivia Agbenyega, and et al. 2019. "Potential Intermediate Hosts for Coronavirus Transmission: No Evidence of Clade 2c Coronaviruses in Domestic Livestock from Ghana" Tropical Medicine and Infectious Disease 4, no. 1: 34. https://doi.org/10.3390/tropicalmed4010034
APA StyleEl-Duah, P., Sylverken, A., Owusu, M., Yeboah, R., Lamptey, J., Oppong Frimpong, Y., Burimuah, V., Antwi, C., Folitse, R., Agbenyega, O., Oppong, S., & Adu-Sarkodie, Y. (2019). Potential Intermediate Hosts for Coronavirus Transmission: No Evidence of Clade 2c Coronaviruses in Domestic Livestock from Ghana. Tropical Medicine and Infectious Disease, 4(1), 34. https://doi.org/10.3390/tropicalmed4010034