
Short-Term Responsive Mating Intentions Increase with Estradiol and Testosterone across the Menstrual Cycle: Scale Development and an Observational Study


Abstract
1. Introduction
2. Materials and Methods
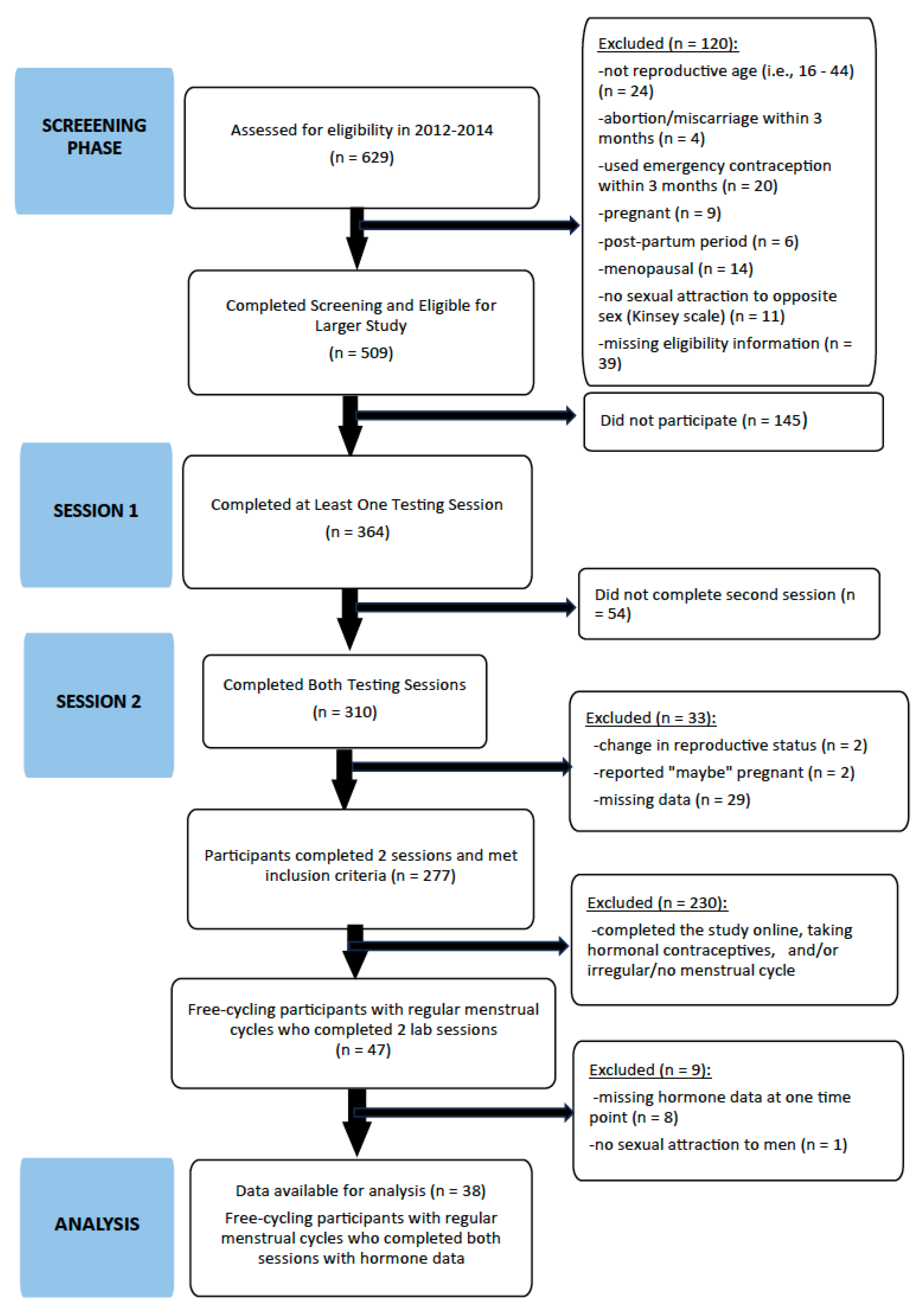
2.1. Participants
2.2. Measures
2.2.1. Demographics
2.2.2. Proactive and Responsive Mating Strategies Scales (PARMSS)
2.2.3. Mating Vignettes
2.2.4. Hormone Assays
2.3. Procedure
2.3.1. Recruitment and Screening
2.3.2. Lab Sessions: Time 1 and Time 2
2.3.3. Statistical Analyses
3. Results
3.1. Factor Analyses, Reliability, and Validity of the PARMSS
3.2. Descriptive Data
3.3. Correlations and Partial Correlations
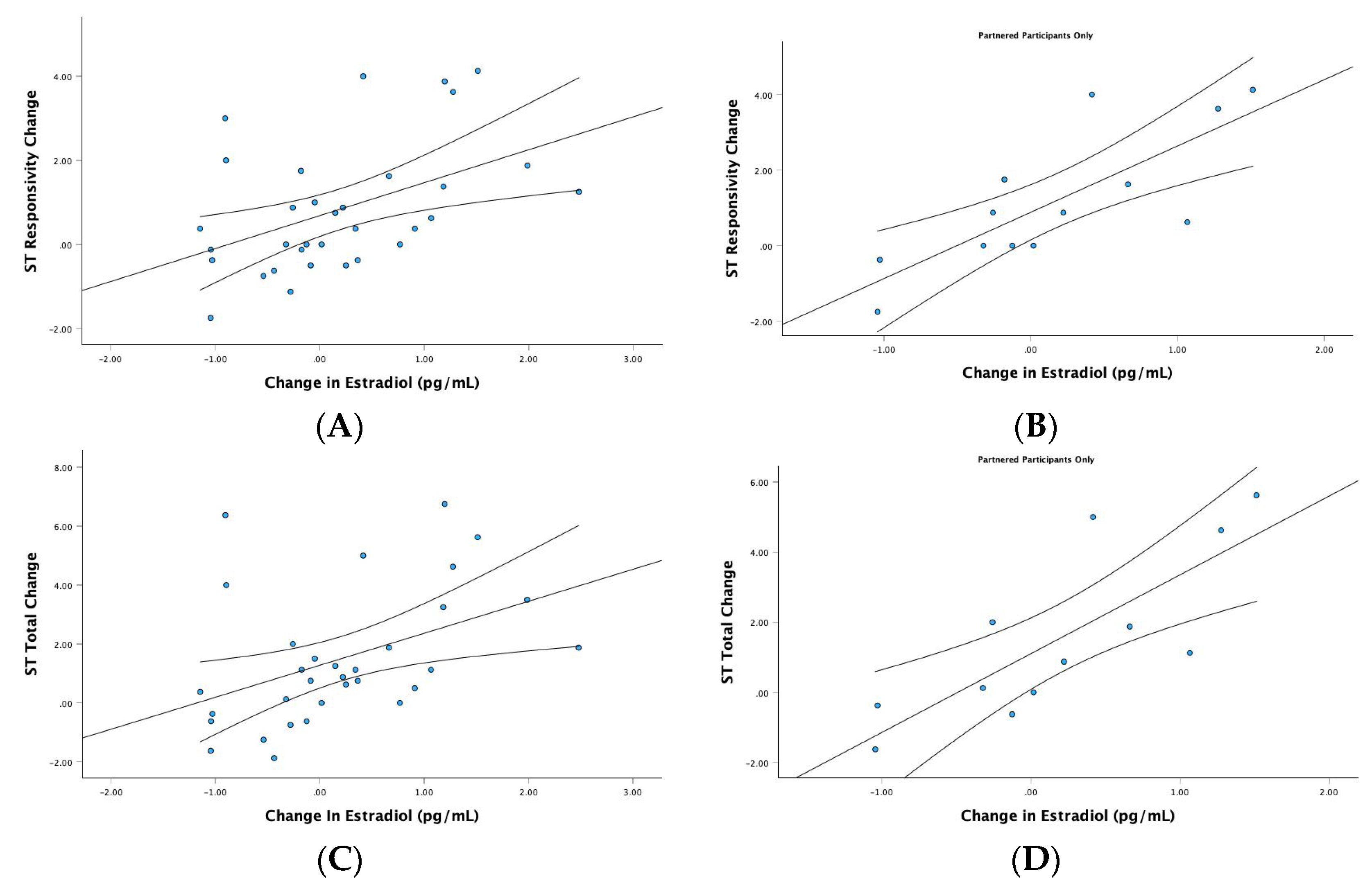
3.3.1. Estradiol
3.3.2. Progesterone
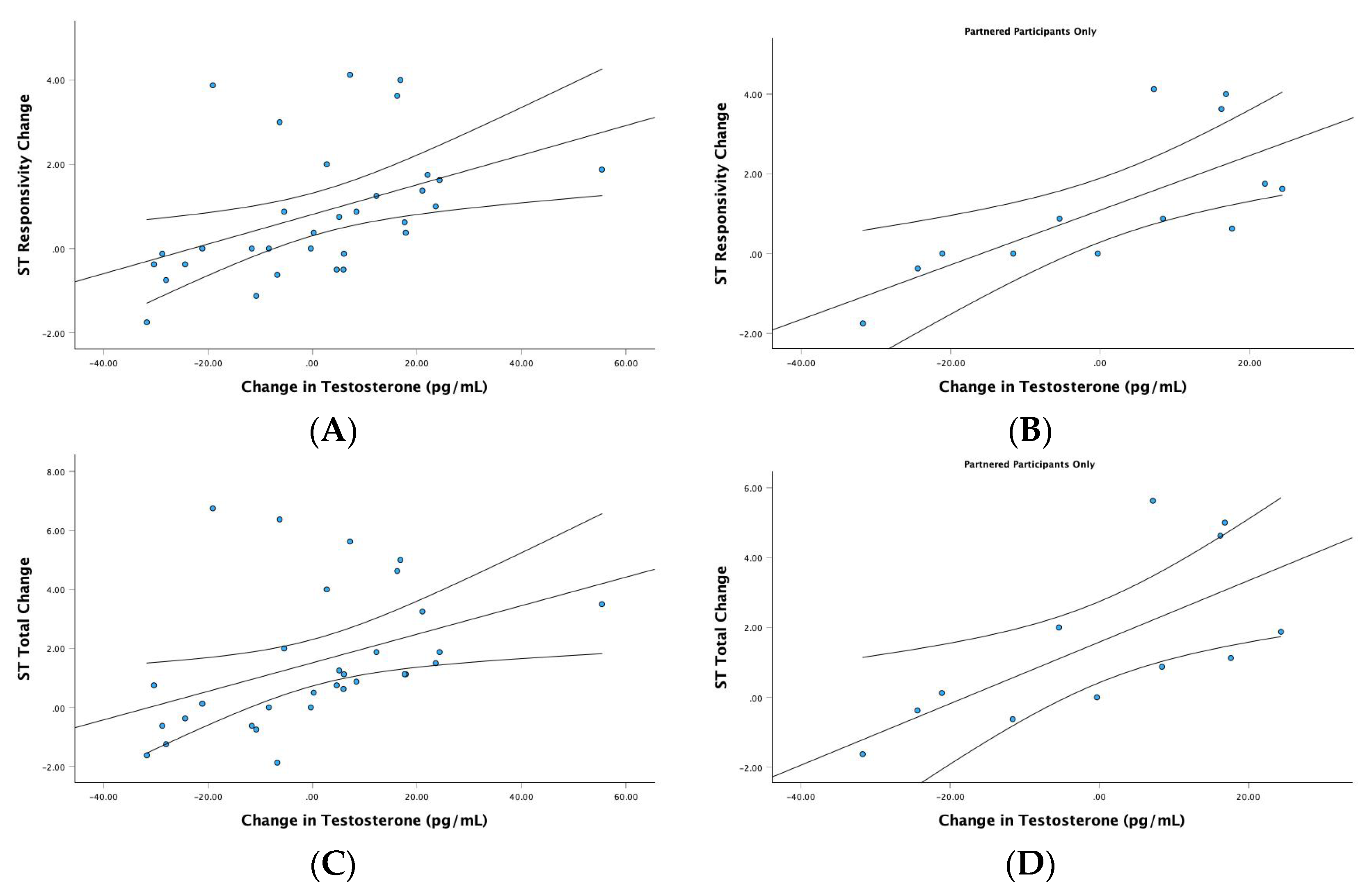
3.3.3. Testosterone
3.3.4. Exploratory Examination of Relationship Status
3.4. Regressions
4. Discussion
4.1. Preliminary Psychometric Support for the PARMSS
4.2. A Stimulatory Effect of Estradiol on ST Mating Intentions across the Cycle
4.3. A Stimulatory Effect of Testosterone on Female ST Mating Intentions
4.4. No Association between Progesterone and Mating Intentions
4.5. Strengths, Limitations, and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
- [ ]
- I have gone through menopause and do not get a period
- [ ]
- I am not currently menstruating because I am currently lactating or breast feeding
- [ ]
- I never have my period
- [ ]
- I have not had my period in the last three months
- [ ]
- Some months I get my period and some months I don’t
- [ ]
- I usually get my period every month, but it is irregular and I cannot predict when it will start
- [ ]
- I usually get my period within two to three days of when I expect it
- [ ]
- My period is like clockwork; the same number of days elapse between periods
Appendix B
- Give this person your phone number if asked
- Ask this person for their number
- Return a smile or eye contact from this person
- Smile, or initiate eye contact with, this person
- Dance with this person if asked
- Ask this person to dance with you
- Allow this person to buy you a drink
- Buy this person a drink or ask this person to buy you a drink
- Allow this person to kiss you
- Initiate kissing with this person
- Accept a ride home from this person if offered
- Offer this person a ride home or ask this person for a ride home
- Allow this person to initiate any sexual activity
- Initiate any sexual activity with this person
- Allow this person to initiate sex
- Initiate sex with this person
Appendix C
References
- Bancroft, J.; Graham, C.A. The varied nature of women’s sexuality: Unresolved issues and a theoretical approach. Horm. Behav. 2011, 59, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Regan, P.C. Rhythms of desire: The association between menstrual cycle phases and female sexual desire. Can. J. Hum. Sex. 1996, 5, 145–156. [Google Scholar]
- Islam, R.M.; Bell, R.J.; Green, S.; Page, M.J.; Davis, S.R. Safety and efficacy of testosterone for women: A systematic review and meta-analysis of randomised controlled trial data. Lancet Diabetes Endocrinol. 2019, 7, 754–766. [Google Scholar] [CrossRef]
- Maseroli, E.; Vignozzi, L. Are endogenous androgens linked to female sexual function? A systemic review and meta-analysis. J. Sex. Med. 2022, 19, 553–568. [Google Scholar] [CrossRef]
- Gildersleeve, K.; Haselton, M.G.; Fales, M.R. Do women’s mate preferences change across the ovulatory cycle? A meta-analytic review. Psychol. Bull. 2014, 140, 1205–1259. [Google Scholar] [CrossRef]
- Barrett, E.S.; Tran, V.; Thurston, S.W.; Frydenberg, H.; Lipson, S.F.; Thune, I.; Ellison, P.T. Women who are married or living as married have higher salivary estradiol and progesterone than unmarried women. Am. J. Hum. Biol. 2015, 27, 501–507. [Google Scholar] [CrossRef]
- Kuzawa, C.; Gettler, L.T.; Huang, Y.; McDade, T.W. Mothers have lower testosterone than non-mothers: Evidence from the Philippines. Horm. Behav. 2010, 57, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.B. Reexploring the concept of sexual desire. J. Sex. Marital. Ther. 2002, 28, 39–51. [Google Scholar] [CrossRef]
- Levine, S.B. The nature of sexual desire: A clinician’s perspective. Arch. Sex. Behav. 2003, 32, 279–285. [Google Scholar] [CrossRef]
- Rellini, A.H.; Stratton, N.; Tonani, S.; Santamaria, V.; Brambilla, E.; Nappi, R.E. Differences in sexual desire between women with clinical versus biochemical signs of hyperandrogenism in polycystic ovarian syndrome. Horm. Behav. 2013, 63, 65–71. [Google Scholar] [CrossRef]
- Beach, F.A. Sexual attractivity, proceptivity, and receptivity in female mammals. Horm. Behav. 1976, 7, 105–138. [Google Scholar] [CrossRef] [PubMed]
- Pfaus, J.G.; Kippin, T.E.; Coria-Avila, G.A. What can animal models tell us about human sexual response? Annu. Rev. Sex. Res. 2003, 14, 1–63. [Google Scholar] [PubMed]
- de Jonge, F.H.; Eerland, E.M.; Van de Poll, N.E. The influence of estrogen, testosterone and progesterone on partner preference, receptivity and proceptivity. Physiol. Behav. 1986, 37, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.A.; Pfeifle, J.K. Hormonal control of receptivity, proceptivity and sexual motivation. Physiol. Behav. 1983, 30, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, C.R. Sexual behavior of free ranging rhesus monkeys (Macaca mulatta). I. Specimens, procedures and behavioral characteristics of estrus. J. Comp. Psychol. 1942, 33, 113–142. [Google Scholar] [CrossRef]
- Wallen, K. Influence of female hormonal state on rhesus sexual behavior varies with space for social interaction. Science 1982, 217, 375–376. [Google Scholar] [CrossRef]
- Young, W.C.; Orbison, W.D. Changes in selected features of behaviour in pairs of oppositely sexed chimpanzees during the sexual cycle and after ovariectomy. J. Comp. Physiol. Psychol. 1944, 37, 107–143. [Google Scholar] [CrossRef]
- Everitt, B.J.; Herbert, J.; Hamer, J.D. Sexual receptivity of bilaterally adrenalectomized female rhesus monkeys. Physiol. Behav. 1972, 8, 409–415. [Google Scholar] [CrossRef]
- Johnson, D.F.; Phoenix, C.H. Hormonal control of female sexual attractiveness, proceptivity and receptivity in rhesus monkeys. J. Comp. Physiol. Psychol. 1976, 90, 473–483. [Google Scholar] [CrossRef]
- Young, W.C. The hormones and mating behaviour. In Sex and Internal Secretions; Young, W.C., Ed.; Bailliere, Tindall and Cox: London, UK, 1961; pp. 1173–1239. [Google Scholar]
- Michael, R.P.; Keverne, E.B.; Zumpe, D.; Bonsall, R.W. Neuroendocrine factors in the control of primate behaviour. Rec. Prog. Horm. Res. 1972, 28, 665–706. [Google Scholar]
- Wallen, K.; Winston, L.A.; Gaventa, S.; Davis-DaSilva, M.; Collins, D.C. Periovulatory changes in female sexual behavior and patterns of ovarian steroid secretion in group-living rhesus macaques. Horm. Behav. 1984, 18, 431–450. [Google Scholar] [CrossRef]
- Warner, L.H. A study of sex behaviour in the white rat by means of the obstruction method. Comp. Psychol. Mong. 1927, 4, 1–68. [Google Scholar]
- Baum, M.J.; Everitt, B.J.; Herbert, J.; Keverne, E.B. Hormonal basis of proceptivity and receptivity in female primates. Arch. Sex. Behav. 1977, 6, 173–192. [Google Scholar] [CrossRef] [PubMed]
- Rowell, T.E. Female reproduction cycles and social behaviour in primates. Adv. Stud. Behav. 1972, 4, 69–105. [Google Scholar]
- Frye, C.A.; Bayon, L.E.; Pursnani, N.K.; Purdy, R.H. The neurosteroids, progesterone and 3α,5α-THP, enhance sexual motivation, receptivity, and proceptivity in female rats. Brain Res. 1998, 808, 72–83. [Google Scholar] [CrossRef]
- Tennent, B.J.; Smith, E.R.; Davidson, J.M. The effects of estrogen and progesterone on female rat proceptive behavior. Horm. Behav. 1980, 14, 65–75. [Google Scholar] [CrossRef]
- Beyer, C.; Vidal, N.; Mijares, A. Probable role of aromatization in the induction of estrous behaviour by androgens in the ovariectomized rabbit. Endocrinology 1970, 87, 1386–1389. [Google Scholar] [CrossRef]
- Keverne, E.B. Sexual receptivity and attractiveness in the female rhesus monkey. In Advances in the Study of Behaviour; Rosenblatt, J.S., Ed.; Academic Press: New York, NY, USA, 1976; Volume 7, pp. 155–200. [Google Scholar]
- Trimble, M.R.; Herbert, J. The effect of testosterone or oestradiol upon the sexual and associated behaviour of the adult female rhesus monkey. J. Endocrinol. 1968, 42, 171–185. [Google Scholar] [CrossRef]
- Beach, F.A.; Leboeuf, B.J. Coital behaviour in dogs: I. Preferential mating in the bitch. Anim. Behav. 1967, 15, 546–558. [Google Scholar] [CrossRef]
- Cochran, C.G. Proceptive patterns of behavior throughout the menstrual cycle in female rhesus monkeys. Behav. Neural Biol. 1979, 27, 342–353. [Google Scholar] [CrossRef]
- Bancroft, J.; Sherwin, B.B.; Alexander, G.M.; Davidson, D.W.; Walker, A. Oral contraceptives, androgens, and the sexuality of young women: I. A comparison of sexual experience, sexual attitudes, and gender role in oral contraceptive users and nonusers. Arch. Sex. Behav. 1991, 20, 105–120. [Google Scholar] [CrossRef]
- Blake, K.R.; Dixson, B.J.W.; O’Dean, S.M.; Denson, T.F. No compelling positive association between ovarian hormones and wearing red clothing when using multinomial analyses. Horm. Behav. 2017, 90, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Grebe, N.M.; Gangestad, S.W.; Garver-Apgar, C.E.; Thornhill, R. Women’s luteal phase sexual proceptivity and the functions of extended sexuality. Psychol. Sci. 2013, 24, 2106–2110. [Google Scholar] [CrossRef] [PubMed]
- Basson, R. The female sexual response: A different model. J. Sex. Marital. Ther. 2000, 26, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Wood, W.; Kressel, L.; Joshi, P.D.; Louie, B. Meta-analysis of menstrual cycle effects on women’s mate preferences. Emot. Rev. 2014, 6, 229–249. [Google Scholar] [CrossRef]
- Adams, D.B.; Gold, A.R.; Burt, A.D. Rise in female-initiated sexual activity at ovulation and its suppression by oral contraceptives. N. Engl. J. Med. 1978, 299, 1145–1150. [Google Scholar] [CrossRef] [PubMed]
- Cantú, S.M.; Simpson, J.A.; Griskevicius, V.; Weisberg, Y.J.; Durante, K.M.; Beal, D.J. Fertile and selectively flirty: Women’s behavior toward men changes across the ovulatory cycle. Psychol. Sci. 2013, 25, 431–438. [Google Scholar] [CrossRef]
- Marcinkowska, U.M.; Shirazi, T.; Mijas, M.; Roney, J.R. Hormonal underpinnings of the variation in sexual desire, arousal and activity throughout the menstrual cycle—A multifaceted approach. J. Sex. Res. 2023, 60, 1297–1303. [Google Scholar] [CrossRef]
- Roney, J.R.; Simmons, Z.L. Hormonal predictors of sexual motivation in natural menstrual cycles. Horm. Behav. 2013, 63, 636–645. [Google Scholar] [CrossRef]
- Arslan, R.C.; Schilling, K.M.; Gerlach, T.M.; Penke, L. Using 26,000 diary entries to show ovulatory changes in sexual desire and behavior. J. Pers. Soc. Psychol. 2021, 121, 410–431, Erratum in J. Pers. Soc. Psychol. 2023, 125, 1238. [Google Scholar] [CrossRef]
- Atukorala, K.R.; Silva, W.; Amarasiri, L.; Fernando, D.M.S. Changes in serum testosterone during the menstrual cycle—An integrative systematic review of published literature. Gynecol. Reprod. Endocrinol. Metab. 2020, 3, 9–20. [Google Scholar] [CrossRef]
- Reed, B.G.; Carr, B.R. The Normal Menstrual Cycle and the Control of Ovulation. Endotext. 2018. Available online: https://www.ncbi.nlm.nih.gov/books/NBK279054 (accessed on 7 June 2024).
- Bullivant, S.B.; Sellegren, S.A.; Stern, K.; Spencer, N.A.; Jacob, S.; Mennella, J.A.; McClintock, M.K. Women’s sexual desire during the menstrual cycle: Identification of the sexual phase by noninvasive measurement of luteinizing hormone. J. Sex. Res. 2004, 41, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Matteo, S.; Rissman, E.F. Increased sexual activity during the midcycle portion of the human menstrual cycle. J. Sex. Res. 1984, 18, 249–255. [Google Scholar] [CrossRef]
- Durante, K.M.; Li, N.P. Oestradiol level and opportunistic mating in women. Biol. Lett. 2009, 5, 179–182. [Google Scholar] [CrossRef]
- PPersky, H.; Dreisbach, L.; Miller, W.R.; O’brien, C.P.; Khan, M.A.; Lief, H.I.; Charney, N.; Strauss, D. The relation of plasma androgen levels to sexual behaviors and attitudes of women. Psychosom. Med. 1982, 44, 305–319. [Google Scholar] [CrossRef]
- Prasad, A.; Mumford, S.L.; Louis, G.M.B.; Ahrens, K.A.; Sjaarda, L.A.; Schliep, K.C.; Schisterman, E.F. Sexual activity, endogenous reproductive hormones and ovulation in premenopausal women. Horm. Behav. 2014, 66, 330–338. [Google Scholar] [CrossRef]
- Wåhlin-Jacobsen, S.; Pedersen, A.T.; Kristensen, E.; Læssøe, N.C.; Lundqvist, M.; Cohen, A.S.; Hougaard, D.M.; Giraldi, A. Is there a correlation between androgens and sexual desire in women? J. Sex. Med. 2015, 12, 358–373. [Google Scholar] [CrossRef] [PubMed]
- Wallen, K. Desire and ability: Hormones and the regulation of female sexual behavior. Neurosci. Biobehav. Rev. 1990, 14, 233–241. [Google Scholar] [CrossRef]
- van Anders, S.M.; Hamilton, L.D.; Schmidt, N.; Watson, N.V. Associations between testosterone secretion and sexual activity in women. Horm. Behav. 2007, 51, 477–482. [Google Scholar] [CrossRef]
- Tao, L.; Duan, Z.; Liu, Y.; Hou, H.; Zhang, X. Correlation of sexual dysfunction with sex hormone and estrogen receptor gene polymorphism in Chinese Han women with epilepsy. Epilepsy Res. 2021, 169, 106527. [Google Scholar] [CrossRef]
- Macdowall, W.G.; Clifton, S.; Palmer, M.J.; Tanton, C.; Copas, A.J.; Lee, D.M.; Mitchell, K.R.; Mercer, C.H.; Sonnenberg, P.; Johnson, A.M.; et al. Salivary testosterone and sexual function and behavior in men and women: Findings from the Third British National Survey of Sexual Attitudes and Lifestyles (Natsal-3). J. Sex. Res. 2022, 59, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Islam, R.M.; Skiba, M.A.; Bell, R.J.; Davis, S.R. Associations between androgens and sexual function in premenopausal women: A cross-sectional study. Lancet Diabetes Endocrinol. 2020, 8, 693–702. [Google Scholar] [CrossRef] [PubMed]
- van Stein, K.R.; Strauß, B.; Brenk-Franz, K. Ovulatory shifts in sexual desire but not mate preferences: An LH-test-confirmed, longitudinal study. Evol. Psychol. 2019, 17, 1474704919848116. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.C.; Hahn, A.C.; Fisher, C.I.; Wang, H.; Kandrik, M.; DeBruine, L.M. General sexual desire, but not desire for uncommitted sexual relationships, tracks changes in women’s hormonal status. Psychoneuroendocrinology 2018, 88, 153–157. [Google Scholar] [CrossRef]
- Grebe, N.M.; Emery Thompson, M.; Gangestad, S.W. Hormonal predictors of women’s extra-pair vs. in-pair sexual attraction in natural cycles: Implications for extended sexuality. Horm. Behav. 2016, 78, 211–219. [Google Scholar] [CrossRef]
- Elaut, E.; Buysse, A.; De Sutter, P.; Gerris, J.; De Cuypere, G.; T’Sjoen, G. Cycle-related changes in mood, sexual desire, and sexual activity in oral contraception-using and nonhormonal-contraception-using couples. J. Sex. Res. 2016, 53, 125–136. [Google Scholar] [CrossRef]
- Bancroft, J.; Sanders, D.; Davidson, D.; Warner, P. Mood, sexuality, hormones, and the menstrual cycle: III. Sexuality and the role of androgens. Psychosom. Med. 1983, 45, 509–516. [Google Scholar] [CrossRef]
- Graham, C.A.; Bancroft, J.; Doll, H.A.; Greco, T.; Tanner, A. Does oral contraceptive induced reduction in free testosterone adversely affect the sexuality or mood of women? Psychoneuroendocrinology 2007, 32, 246–255. [Google Scholar] [CrossRef]
- Oinonen, K.A. Putting a finger on potential predictors of oral contraceptive side effects: 2D:4D and middle-phalangeal hair. Psychoneuroendocrinology 2009, 34, 713–726. [Google Scholar] [CrossRef]
- Tzalazidis, R.; Oinonen, K.A. Continuum of symptoms in Polycystic Ovary Syndrome (PCOS): Links with sexual behavior and unrestricted sociosexuality. J. Sex. Res. 2021, 58, 532–544. [Google Scholar] [CrossRef]
- Schmidt, P.J.; Martinez, P.E.; Nieman, L.K.; Koziol, D.E.; Thompson, K.D.; Schenkel, L.; Wakim, P.G.; Rubinow, D.R. Premenstrual Dysphoric Disorder symptoms following ovarian suppression: Triggered by change in ovarian steroid levels but not continuous stable levels. Am. J. Psychiatry 2017, 174, 980–989. [Google Scholar] [CrossRef]
- Teatero, M. Women’s Reproductive Experiences (REP) and Hormones: Patterns in Affective, Sexual, and Physical Well-Being. Doctoral Dissertation, Lakehead University, Thunder Bay, ON, Canada, 2016. [Google Scholar]
- Kinsey, A.C.; Pomeroy, W.R.; Martin, C.E. Sexual behavior in the human male. Am. J. Public Health 1948, 93, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.J.; Kirkpatrick, L.A. The structure and measurement of human mating strategies: Toward a multidimensional model of sociosexuality. Evol. Hum. Behav. 2007, 28, 382–391. [Google Scholar] [CrossRef]
- Phillips, M. The Ovulatory Shift: Proceptive and Receptive Mating Behaviours Across the Menstrual Cycle. Doctoral Dissertation, Lakehead University, Thunder Bay, ON, Canada, 2015. [Google Scholar]
- Little, A.C.; Jones, B.C.; Burriss, R.P. Preferences for masculinity in male bodies change across the menstrual cycle. Horm. Behav. 2007, 51, 633–639. [Google Scholar] [CrossRef]
- von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gøtzsche, P.C.; Vandenbroucke, J.P.; STROBE Initiative. The strengthening the reporting of observational studies in epidemiology (STROBE) statement: Guidelines for reporting observational studies. PLoS Med. 2007, 4, e296. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge Academic: New York, NY, USA, 1988. [Google Scholar]
- Tabachnick, B.G.; Fidell, L.S. Using Multivariate Statistics, 5th ed.; Allyn and Bacon: New York, NY, USA, 2007. [Google Scholar]
- Arifin, W.N. Sample Size Calculator. 2024. Available online: http://wnarifin.github.io (accessed on 7 June 2024).
- Field, A. Uncovering Statistics Using SPSS, 3rd ed.; SAGE Publications: Thousand Oaks, CA, USA, 2009. [Google Scholar]
- Little, A.C.; Jones, B.C.; Penton-Voak, I.S.; Burt, D.M.; Perrett, D.I. Partnership status and the temporal context of relationships influence human female preferences for sexual dimorphism in male face shape. Proc. Biol. Sci. 2002, 269, 1095–1100. [Google Scholar] [CrossRef]
- Shirazi, T.N.; Bossio, J.A.; Puts, D.A.; Chivers, M.L. Menstrual cycle phase predicts women’s hormonal responses to sexual stimuli. Horm. Behav. 2018, 103, 45–53. [Google Scholar] [CrossRef]
- Roney, J.R.; Simmons, Z.L. Within-cycle fluctuations in progesterone negatively predict changes in both in-pair and extra-pair desire among partnered women. Horm. Behav. 2016, 81, 45–52. [Google Scholar] [CrossRef]
- Gangestad, S.W.; Garver-Apgar, C.; Simpson, J.A.; Cousins, A.J. Changes in women’s mate preferences across the ovulatory cycle. J. Pers. Soc. Psychol. 2007, 92, 151–163. [Google Scholar] [CrossRef]
- Larson, C.M.; Pillsworth, E.G.; Haselton, M.G. Ovulatory shifts in women’s attractions to primary partners and other men: Further evidence of the importance of primary partner sexual attractiveness. PLoS ONE 2012, 7, e44456. [Google Scholar] [CrossRef]
- Fink, B.; Penton-Voak, I. Evolutionary psychology of facial attractiveness. Curr. Dir. Psychol. Sci. 2002, 11, 154–158. [Google Scholar] [CrossRef]
- Wardecker, B.M.; Smith, L.K.; Edelstein, R.S.; Loving, T.J. Intimate relationships then and now: How old hormonal processes are influenced by our modern psychology. Adapt. Human. Behav. Physiol. 2015, 1, 150–176. [Google Scholar] [CrossRef]
- Pillsworth, E.G.; Haselton, M.G. Male sexual attractiveness predicts differential ovulatory shifts in female extra-pair attraction and male mate retention. Evol. Hum. Behav. 2006, 27, 247–258. [Google Scholar] [CrossRef]
- Costa, R.M.; Oliveira, G.; Pestana, J.; Costa, D.; Oliveira, R.F. Do psychosocial factors moderate the relation between testosterone and female sexual desire? The role of interoception, alexithymia, defense mechanisms, and relationship status. Adapt. Human. Behav. Physiol. 2019, 5, 13–30. [Google Scholar] [CrossRef]
- Shirazi, T.N.; Self, H.; Dawood, K.; Rosenfield, K.A.; Penke, L.; Carré, J.M.; Ortiz, T.; Puts, D.A. Hormonal predictors of women’s sexual motivation. Evol. Hum. Behav. 2019, 40, 336–344. [Google Scholar] [CrossRef]
- López, H.H.; Hay, A.C.; Conklin, P.H. Attractive men induce testosterone and cortisol release in women. Horm. Behav. 2009, 56, 84–92. [Google Scholar] [CrossRef]
- Goldey, K.L.; van Anders, S.M. Sexy thoughts: Effects of sexual cognitions of testosterone, cortisol, and arousal in women. Horm. Behav. 2011, 59, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, R.S.; Chopik, W.J.; Kean, E.L. Sociosexuality moderates the association between testosterone and relationship status in men and women. Horm. Behav. 2011, 60, 248–255. [Google Scholar] [CrossRef]
- van Anders, S.M.; Watson, N.V. Social neuroendocrinology: Effects of social contexts and behaviors on sex steroids in humans. Hum. Nat. 2006, 17, 212–237. [Google Scholar] [CrossRef]
- Ellison, P.T. Reproductive Ecology and Human Evolution, 1st ed.; Routledge: New York, NY, USA, 2011. [Google Scholar] [CrossRef]
- Dennerstein, L.; Gotts, G.; Brown, J.B.; Morse, C.A.; Farley, T.M.; Pinol, A. The relationship between the menstrual cycle and female sexual interest in women with PMS complaints and volunteers. Psychoneuroendocrinology 1994, 19, 293–304. [Google Scholar] [CrossRef]
- Hsueh, A.J.; Peck, E.J.; Clark, J.H. Progesterone antagonism of the oestrogen receptor and oestrogen-induced uterine growth. Nature 1975, 254, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.M.; McRae-Clark, A.L.; Carlson, S.; Saladin, M.E.; Gray, K.M.; Wetherington, C.L.; McKee, S.A.; Allen, S.S. Determining menstrual phase in human biobehavioral research: A review with recommendations. Exp. Clin. Psychopharmacol. 2016, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Schultheiss, O.C.; Dlugash, G.; Mehta, P.H. Hormone measurement in social neuroendocrinology: A comparison of immunoassay and mass spectroscopy methods. In Routledge International Handbook of Social Neuroendocrinology; Schultheiss, O.C., Mehta, P.H., Eds.; Routledge: New York, NY, USA, 2019; pp. 239–255. [Google Scholar]
- Welker, K.M.; Lassetter, B.; Brandes, C.M.; Prasad, S.; Koop, D.R.; Mehta, P.H. A comparison of salivary testosterone measurement using immunoassays and tandem mass spectrometry. Psychoneuroendocrinology 2016, 71, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Teatero, M.; Oinonen, K. The Proactive and Responsive Sexual Strategies Scales (PARMSS) and Changes in Estradiol, Progesterone, and Testosterone Across the Menstrual Cycle: Two Data Sets. 2024. Available online: https://borealisdata.ca/dataset.xhtml?persistentId=doi:10.5683/SP3/VOJUDS (accessed on 7 June 2024).

{kind=link}
{kind=link}
{kind=link}
| Proactive Scale Items | Item Loadings on Factor 1 for Each Context | ||||
|---|---|---|---|---|---|
| ST Vignette | LT Vignette | Picture 1 | Picture 2 | Picture 3 | |
| Ask this person for their number? | 0.758 | 0.884 | 0.899 | 0.904 | 0.908 |
| Smile at, or make eye contact with, this person? | 0.671 | 0.727 | 0.664 | 0.717 | 0.668 |
| Ask this person to dance with you? | 0.817 | 0.886 | 0.878 | 0.902 | 0.881 |
| Buy this person a drink or ask this person to buy you a drink? | 0.809 | 0.869 | 0.872 | 0.874 | 0.894 |
| Initiate kissing with this person? | 0.901 | 0.915 | 0.920 | 0.924 | 0.915 |
| Offer this person a ride home or ask this person for a ride home? | 0.736 | 0.82 | 0.818 | 0.798 | 0.786 |
| Initiate any sexual activity with this person? | 0.882 | 0.887 | 0.901 | 0.903 | 0.881 |
| Initiate sex with this person? | 0.822 | 0.846 | 0.861 | 0.876 | 0.873 |
| Eigenvalue for first factor | 5.158 | 5.86 | 5.85 | 5.984 | 5.842 |
| Percentage of variance accounted for | 64.471 | 73.256 | 73.122 | 74.801 | 73.03 |
| Kaiser–Meyer–Olkin Measure of Sampling Adequacy | 0.884 | 0.912 | 0.907 | 0.896 | 0.886 |
| Bartlett’s Test of Sphericity—Approx. Chi-Square | 2179.997 | 2899.891 | 2943.577 | 3625.036 | 3365.811 |
| df | 28 | 28 | 28 | 28 | 28 |
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| N | 353 | 360 | 359 | 364 | 358 |
| Cronbach’s Alpha | 0.93 | 0.946 | 0.938 | 0.946 | 0.926 |
| Test–Retest Reliability (Pearson Correlation) | 0.82 | 0.796 | |||
| Responsive Scale Items | Item Loadings on Factor 1 for Each Context | ||||
|---|---|---|---|---|---|
| ST Vignette | LT Vignette | Picture 1 | Picture 2 | Picture 3 | |
| Give this person your phone number if asked? | 0.826 | 0.876 | 0.873 | 0.9 | 0.855 |
| Return a smile or eye contact from this person? | 0.716 | 0.781 | 0.729 | 0.765 | 0.723 |
| Dance with this person if asked? | 0.832 | 0.891 | 0.872 | 0.875 | 0.863 |
| Allow this person to buy you a drink? | 0.821 | 0.875 | 0.842 | 0.859 | 0.822 |
| Allow this person to kiss you? | 0.906 | 0.925 | 0.909 | 0.929 | 0.903 |
| Accept a ride home from this person if offered? | 0.752 | 0.816 | 0.775 | 0.795 | 0.77 |
| Allow this person to initiate any sexual activity? | 0.874 | 0.847 | 0.863 | 0.868 | 0.848 |
| Allow this person to initiate sex? | 0.833 | 0.802 | 0.817 | 0.837 | 0.813 |
| Eigenvalue for first factor | 5.405 | 5.817 | 5.602 | 5.848 | 5.462 |
| Percentage of variance accounted for | 67.56 | 72.714 | 70.024 | 73.098 | 68.272 |
| Kaiser–Meyer–Olkin Measure of Sampling Adequacy. | 0.893 | 0.896 | 0.891 | 0.881 | 0.875 |
| Bartlett’s Test of Sphericity—Approx. Chi-Square | 2596.065 | 3182.129 | 2859.574 | 3594.088 | 3009.983 |
| df | 28 | 28 | 28 | 28 | 28 |
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| N | 353 | 360 | 359 | 364 | 358 |
| Cronbach’s Alpha | 0.93 | 0.946 | 0.938 | 0.946 | 0.926 |
| Test–Retest Reliability (Pearson Correlation) | 0.82 | 0.796 | |||
| PARMSS Across Contexts | Estradiol (n = 38) | Progesterone (n = 38) | Testosterone (n = 37) | |||
|---|---|---|---|---|---|---|
| ST proactivity | 0.25 | (0.12) | −0.04 | (−0.26) | 0.24 | (0.14) |
| LT proactivity | 0.17 | (0.20) | −0.04 | (−0.29) | 0.04 | (0.07) |
| Picture proactivity | −0.14 | (−0.33) | −0.01 | (−0.08) | 0.00 | (0.22) |
| ST responsivity | 0.48 ** | (0.34) | 0.01 | (−0.24) | 0.44 * | (0.13) |
| LT responsivity | 0.24 | (0.25) | −0.03 | (−0.34) | 0.07 | (0.10) |
| Picture responsivity | −0.01 | (−0.27) | 0.08 | (−0.02) | 0.15 | (0.30) |
| Total proactivity | 0.09 | (0.04) | −0.03 | (−0.26) | 0.03 | (0.15) |
| Total responsivity | 0.26 | (0.17) | 0.02 | (−0.27) | 0.15 | (0.19) |
| Total picture | −0.13 | (−0.31) | 0.03 | (−0.05) | 0.10 | (0.27) |
| Total ST | 0.44 * | (0.27) | 0.01 | (−0.27) | 0.41 ** | (0.15) |
| Total LT | 0.21 | (0.23) | −0.03 | (−0.33) | 0.04 | (0.09) |
| Total PARMSS | 0.20 | (0.18) | −0.02 | (−0.27) | 0.10 | (0.17) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zugic, K.E.; Teatero, M.L.; Phillips, M.; Oinonen, K.A. Short-Term Responsive Mating Intentions Increase with Estradiol and Testosterone across the Menstrual Cycle: Scale Development and an Observational Study. Sexes 2024, 5, 475-497. https://doi.org/10.3390/sexes5040034
Zugic KE, Teatero ML, Phillips M, Oinonen KA. Short-Term Responsive Mating Intentions Increase with Estradiol and Testosterone across the Menstrual Cycle: Scale Development and an Observational Study. Sexes. 2024; 5(4):475-497. https://doi.org/10.3390/sexes5040034
Chicago/Turabian StyleZugic, Kaitlyn E., Missy L. Teatero, Maggie Phillips, and Kirsten A. Oinonen. 2024. "Short-Term Responsive Mating Intentions Increase with Estradiol and Testosterone across the Menstrual Cycle: Scale Development and an Observational Study" Sexes 5, no. 4: 475-497. https://doi.org/10.3390/sexes5040034
APA StyleZugic, K. E., Teatero, M. L., Phillips, M., & Oinonen, K. A. (2024). Short-Term Responsive Mating Intentions Increase with Estradiol and Testosterone across the Menstrual Cycle: Scale Development and an Observational Study. Sexes, 5(4), 475-497. https://doi.org/10.3390/sexes5040034