A Modified Protocol for the Isolation, Culture, and Characterization of Human Smooth Muscle Cells from the Umbilical Cord




Abstract
1. Introduction
2. Methods
2.1. Solutions
2.2. Collection of Tissue Sample
2.3. Dissection of the Umbilical Cord
- The cord was removed from the collection solution, and a clean transverse cut was made across the two ends of the cord.
- To obtain a higher yield of cells, a long segment of the cord (25–30 cm) was used.
- The cord was cut into 3–5 cm long segments, and they were kept submerged in PBS in a sterile plastic Petri dish.
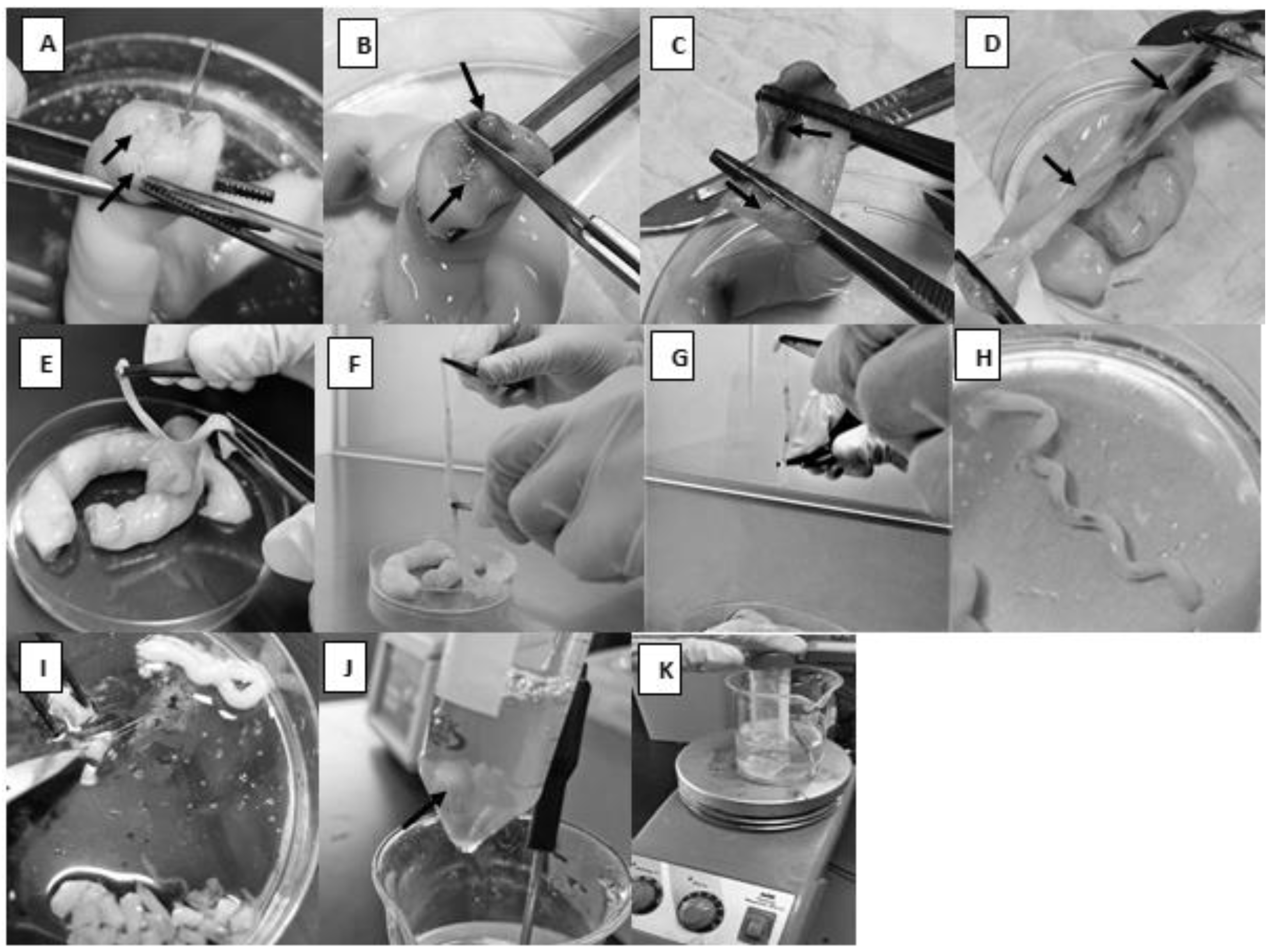
- Using forceps, one segment was held in a way that allows for visualizing the two arteries and the vein (Figure 1A).
- A sterile scalpel was used to make a sagittal section that separates the two arteries from each other so that the vein would be cut longitudinally (Figure 1B).
- Each half was held with blunt, straight stainless-steel forceps, and they were ripped apart by pulling each half in the opposite direction (Figure 1C,D).
- Using pointed forceps, the thick-walled artery was picked up and ripped carefully from the surrounding Wharton’s jelly (Figure 1E). It would not be a problem if the whole artery was not captured since it would be cut into smaller pieces at a later step. To make sure a segment of the artery, not Wharton’s jelly, would be specifically picked, note the spiral appearance of the artery (Figure 1H).
- The isolated arteries were submerged in PBS in a new sterile Petri dish.
2.4. Cell Dissociation and Primary Culture of VSMCs
- Culture media and solutions were prepared and pre-warmed to 37 °C in a water bath.
- As much surrounding connective tissue as possible was removed from around the arteries (Figure 1F); then, the arteries were immersed in PBS in a Petri dish to keep them moist.
- To avoid contamination with other cell types, such as fibroblasts, adventitia was removed mechanically using forceps (Figure 1G). Note: the prevention of contamination with HUVECs is described in step 9 below.
- The arteries were washed with PBS and transferred to a new container.
- The arteries were cut into 1–3 mm pieces (Figure 1I).
- The tissue pieces were placed in a sterile 50 mL conical tube.
- A sterile magnetic stir bar was added to the conical tube and followed by approximately 10 mL of 0.3% collagenase type II to give a ratio of tissue to enzyme of 1:5 (w/v) (Figure 1J).
- The conical tube was placed inside a 250 mL beaker containing 100 mL distilled water on a heating magnetic stirrer. The temperature was continuously set to 37 °C (Figure 1K). The mechanical movement of the magnetic stir bar inside the conical tube should enhance cell separation from the tissue.
- After two hours, the solution was passed through a sterile sieve (an autoclaved stainless-steel strainer, 1 mm pore size) to remove any remaining tissue pieces. Based on our experience, when a 70 µm cell strainer (Corning, NY, USA) was used, the solution containing the cells would not pass through easily because it was highly viscous. In addition, the 2 h period was sufficient to yield a good number of VSMCs from tunica media without reaching tunica intima, which encompasses HUVECs that should be trapped by the strainer and, thus, prevents the contamination of the primary VSMC culture with HUVECs (see also the Supplementary Materials).
- The cells were pelleted by centrifugation at 1120× g for 10 min at 4 °C.
- Except for the last 2 mL, the supernatant was aspirated and discarded.
- The pellet was resuspended in 5 mL of complete medium and plated in a gelatin-coated well (0.5 mL coating solution) of a 6-well plate.
- The plate was incubated at 37 °C in a humidified incubator at 5% CO2 for 5 days. On the third day, and without disturbing the culture, 1 mL of complete medium was added gently to nourish the cells.
- After the 5-day incubation period, the media with unattached cells were removed and refreshed with 2 mL fresh media. The red blood cells (RBCs) were discarded by gently washing the cultured cells with warm serum-free M199 medium, and the incubation was continued as usual until confluency.
- The cells were split into a gelatin-coated culture flask as described below.
2.5. Subculturing
- The culture medium was aspirated, and the cells were washed with pre-warmed, sterile PBS to remove any traces of serum.
- Pre-warmed trypsin-EDTA solution was added (1 mL per 25 cm2 culture flasks or 2 mL for 75 cm2 culture flasks) to cover the cells, and the flask was placed in a humidified incubator at 5% CO2 and 37 °C for 2–4 min. The flask could be examined under a microscope to ensure complete cell detachment. This can also be facilitated by tapping the side of the flask. Note that VSMCs detach relatively slowly and, thus, may take a longer time to fully detach (approximately 10 min).
- Serum-containing medium (double the amount of trypsin-EDTA solution) was added to stop the action of trypsin. To break up any cell clumps, the cell suspension was gently pipetted four to six times.
- The cells were transferred into new gelatin-coated culture flasks at a split ratio of 1:3, and sufficient serum-containing medium was added to the new flasks (5 mL in a 25 cm2 flask and 10 mL in a 75 cm2 flask). Every 2–3 days, half of the culture medium was replaced by a fresh one until confluency. Note: the growth rate of the cells may decline and phenotypic changes become evident beyond passage 10, and therefore, further subculturing of the cells is not recommended.
- For experimental protocols, growth medium can be replaced with either low-serum, low-ECGF, or serum-free medium for up to 24 h, but not later, since the cells may exhibit an altered apoptotic phenotype.
2.6. Characterization of VSMCs
2.7. Cryopreservation
- Cells at passage 3 or after were detached from a 75 cm2 culture flask (see step 2 of Section 2.5 above). Note: the cell content of a 75 cm2 flask can be frozen into 3 cryovials, if fully confluent.
- Following centrifugation of the cell suspension for 10 min at 1120× g, the supernatant was aspirated, and the cell pellet was resuspended in 900 μL FBS for each cryovial with an additional 100 μL dimethyl sulfoxide (DMSO) and then transferred to a suitable cryovial.
- The cryovial was then placed at 4 °C for 30 min, transferred to −20 °C for 30 min, and, finally, to −80 °C for 2 h before being immersed into liquid nitrogen for long-term (~6 months to 1 year) storage. Alternatively, a “Mr. Frosty” cryovial freezing chamber containing isopropanol was used and placed at −80 °C directly.
- When re-culturing the frozen cells, they were thawed quickly, added to 5 mL M199 medium, and centrifuged at 1120× g for 10 min. The cell pellet was re-suspended and cultured as usual. Otherwise, the thawed cells were added to a 25 or 75 cm2 culture flask and left until they adhered to the surface (not more than 24 h) before renewing the medium.
3. Results
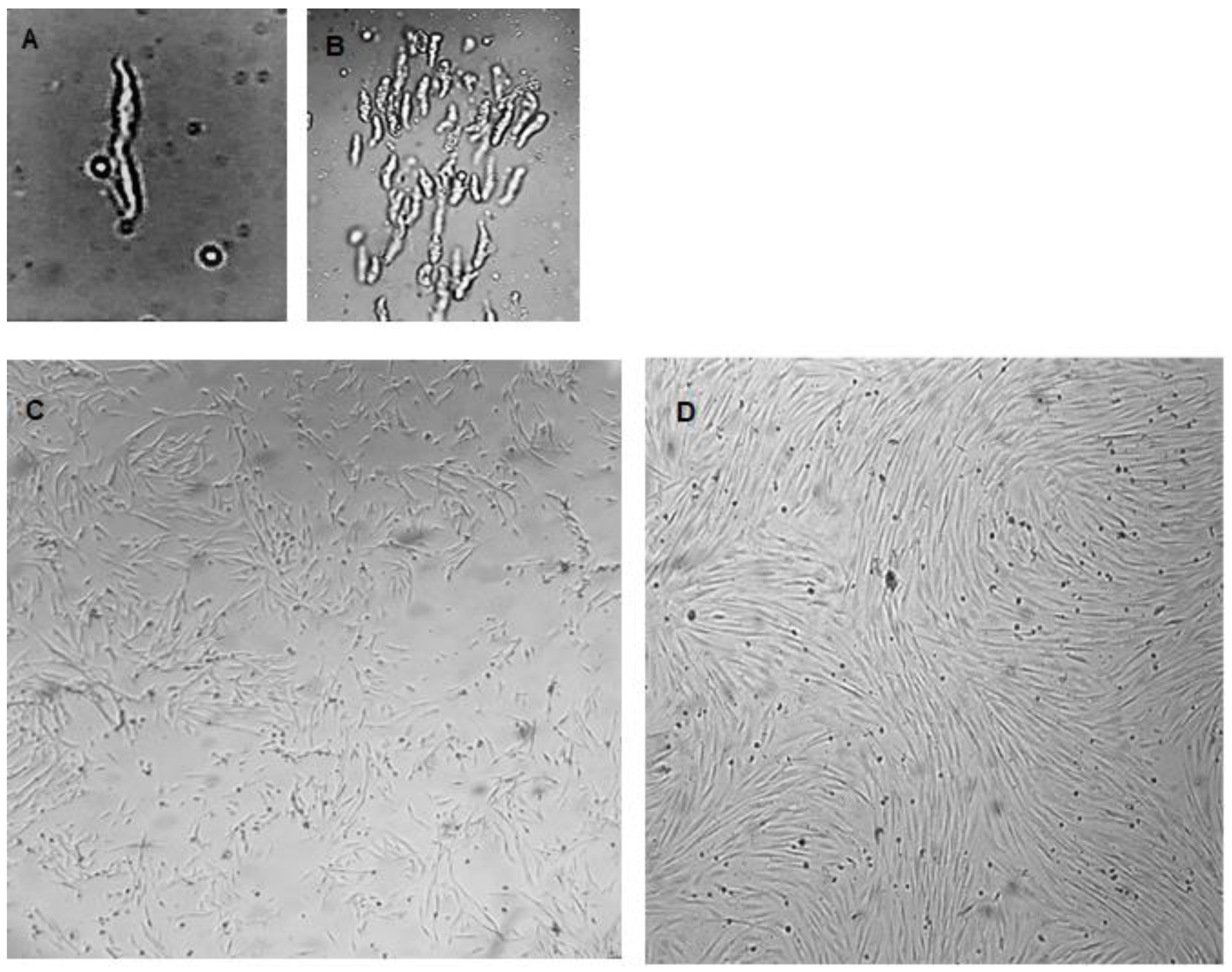
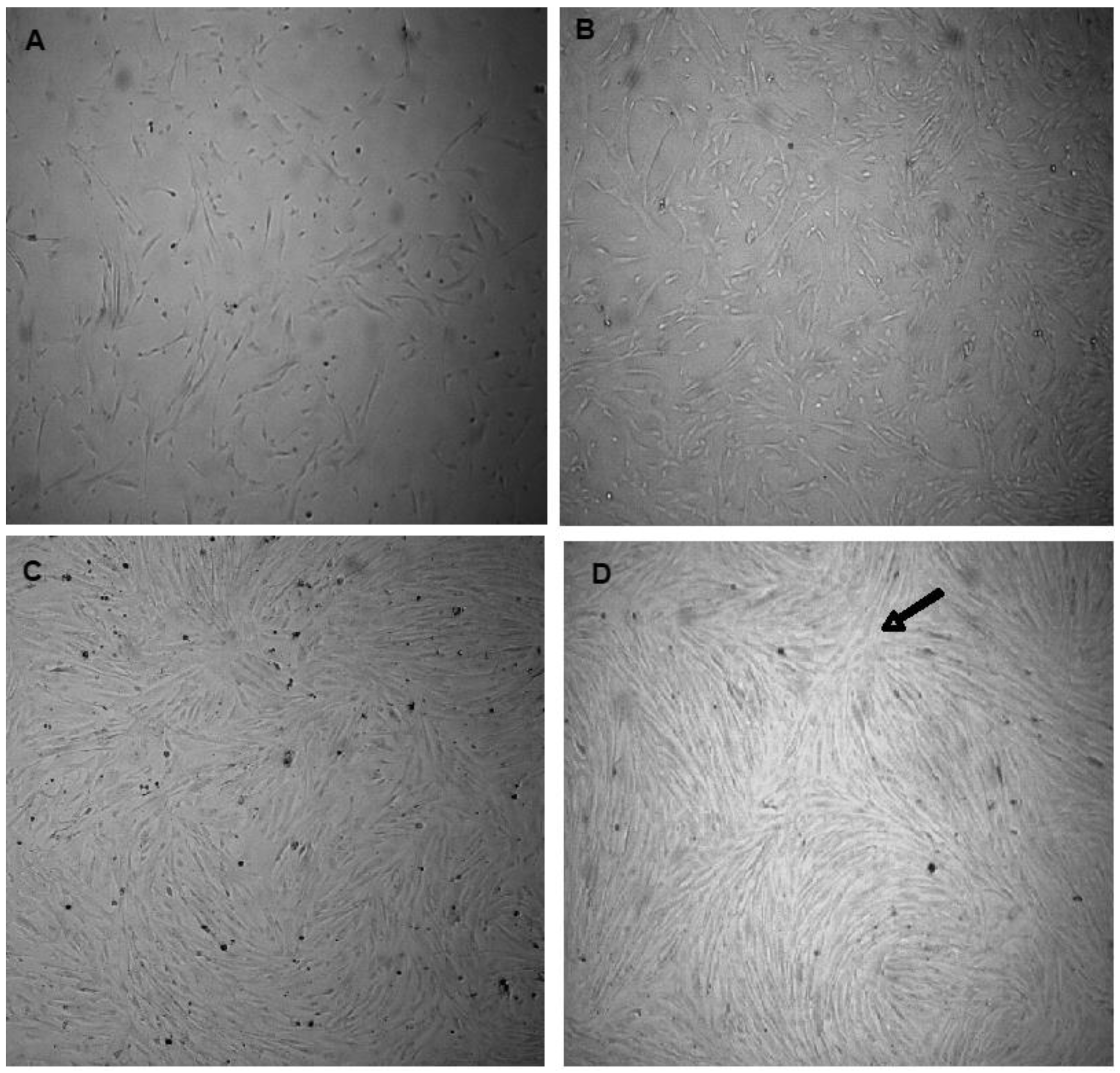
3.1. Cell Isolation and Cultures
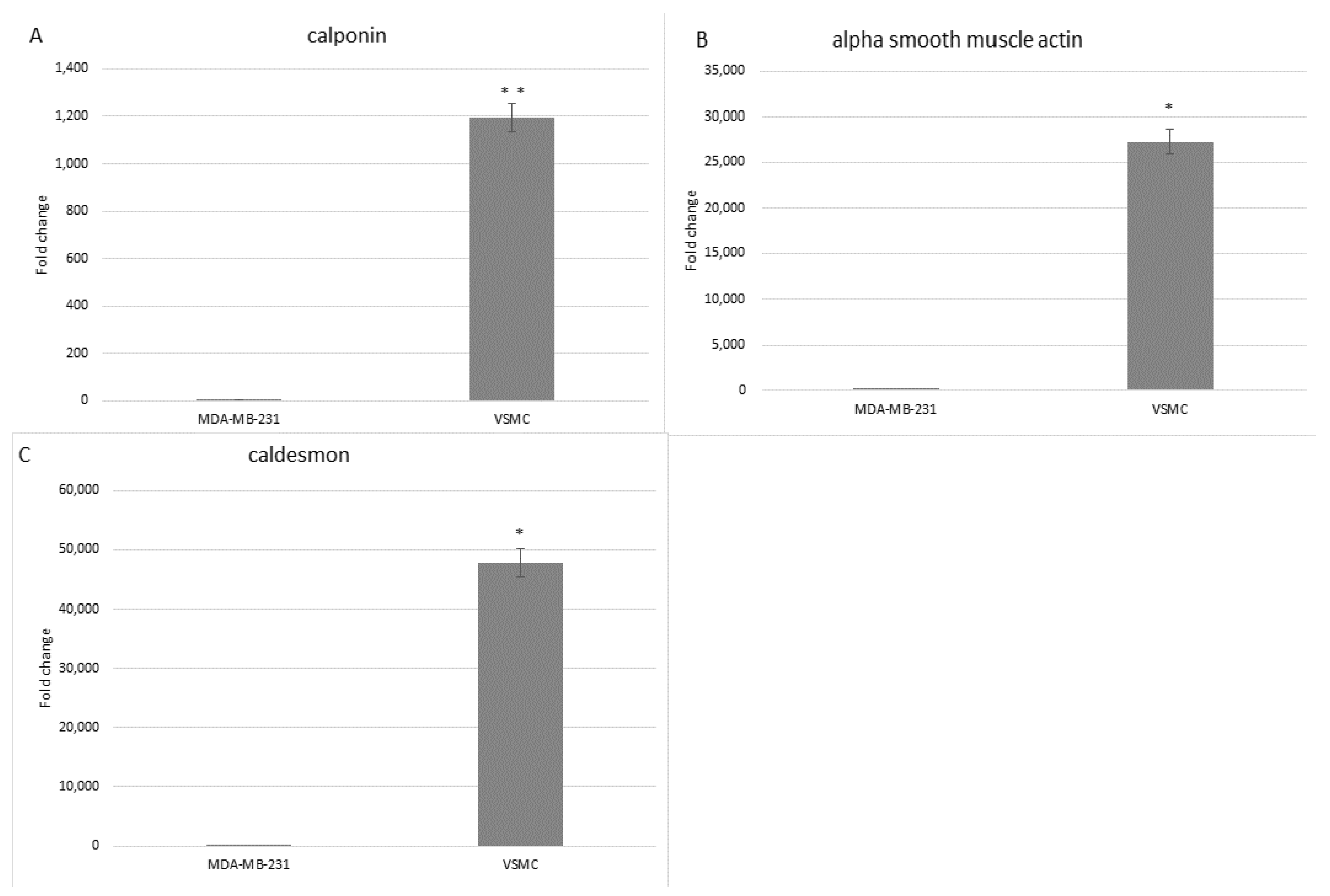
3.2. Characterization
3.3. Isolation of HUVECs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, Y.; Sun-Waterhouse, D.; Chen, Y.; Li, F.; Li, D. Epigenetic mechanisms underlying the benefits of flavonoids in cardiovascular health and diseases: Are long non-coding RNAs rising stars? Crit. Rev. Food Sci. Nutr. 2022, 62, 3855–3872. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Jiang, X.; Jiang, X.; Zhao, H. X-inactive-specific transcript: A long noncoding RNA with complex roles in human cancers. Gene 2018, 679, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Siow, R.C.M. Culture of human endothelial cells from umbilical veins. In Human Cell Culture Protocols, 3rd ed.; Mitry, R., Hughes, R., Eds.; Humana press: London, UK, 2012. [Google Scholar]
- Patel, J.J.; Srivastava, S.; Siow, R.C. Isolation, culture, and characterization of vascular smooth muscle cells. Methods Mol. Biol. 2016, 1430, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Cairrão, E.; Santos-Silva, A.J.; Alvarez, E.; Correia, I.; Verde, I. Isolation and culture of human umbilical artery smooth muscle cells expressing functional calcium channels. Vitr. Cell Dev. Biol.—Anim. 2009, 45, 175–184. [Google Scholar] [CrossRef] [PubMed]
- McCallinhart, P.E.; Biwer, L.A.; Clark, O.E.; Isakson, B.E.; Lilly, B.; Trask, A.J. Myoendothelial junctions of mature coronary vessels express notch signaling proteins. Front. Physiol. 2020, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Mescher, A. Junqueira’s Basic Histology: Text and Atlas, 14th ed.; McGraw-Hill: Bloomington, IN, USA, 2015. [Google Scholar]
- Proudfoot, D.; Shanahan, C. Human vascular smooth muscle cell culture. Methods Mol. Biol. 2012, 806, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Okker-Reitsma, G.H.; Dziadkowiec, I.J.; Groot, C.G. Isolation and culture of smooth muscle cells from human umbilical cord arteries. Vitr. Cell Dev. Biol. 1985, 21, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Kwartler, C.; Zhou, P.; Kuang, S.; Duan, X.; Gong, L.; Milewicz, D. Vascular smooth muscle cell isolation and culture from mouse aorta. Bio-Protocol 2016, 6, e2045. [Google Scholar] [CrossRef]
- Yarbrough, D.; Gerecht, S. Engineering smooth muscle to understand extracellular matrix remodeling and vascular disease. Bioengineering 2022, 9, 449. [Google Scholar] [CrossRef] [PubMed]
- Milutinovic, A.; Suput, D.; Zorc-Pleskovic, R. Pathogenesis of atherosclerosis in the tunica intima, media, and adventitia of coronary arteries: An updated review. Bosn. J. Basic Med. Sci. 2020, 20, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.Y.; Chen, A.Q.; Zhang, H.; Gao, X.F.; Kong, X.Q.; Zhang, J.J. Vascular smooth muscle cells phenotypic switching in cardiovascular diseases. Cells 2022, 11, 4060. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Z.; Li, Z.X.; Zhang, L.L.; Wang, D.; Liu, Y.P. Phenotypic plasticity of vascular smooth muscle cells in vascular calcification: Role of mitochondria. Front. Cardiovasc. Med. 2022, 9, 72836–72852. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Tm (°C) | Reverse Primer | Tm (°C) | References (Origene; CAT) |
|---|---|---|---|---|---|
| Calponin | CCAACGACCTGTTTGAGAACACC | 58.3 | ATTTCCGCTCCTGCTTCTCTGC | 59.8 | HP205213 |
| α-smooth muscle actin | CTATGCCTCTGGACGCACAACT | 59.3 | CAGATCCAGACGCATGATGGCA | 59.6 | HP205437 |
| Caldesmon | CTGTTCCTGCTGAAGGTGTACG | 57.7 | CCTACCTTCAAGCCAGCAGTTTC | 58.0 | HP234228 |
| GAPDH | GTCTCCTCTGACTTCAACAGCG | 57.4 | ACCACCCTGTTGCTGTAGCCAA | 61.3 | HP205798 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, A.Q.; Abdullah, M.S.; Ahram, M.; Abdalla, S. A Modified Protocol for the Isolation, Culture, and Characterization of Human Smooth Muscle Cells from the Umbilical Cord. Methods Protoc. 2023, 6, 54. https://doi.org/10.3390/mps6030054
Ibrahim AQ, Abdullah MS, Ahram M, Abdalla S. A Modified Protocol for the Isolation, Culture, and Characterization of Human Smooth Muscle Cells from the Umbilical Cord. Methods and Protocols. 2023; 6(3):54. https://doi.org/10.3390/mps6030054
Chicago/Turabian StyleIbrahim, Asmaa Q., Mohammed S. Abdullah, Mamoun Ahram, and Shtaywy Abdalla. 2023. "A Modified Protocol for the Isolation, Culture, and Characterization of Human Smooth Muscle Cells from the Umbilical Cord" Methods and Protocols 6, no. 3: 54. https://doi.org/10.3390/mps6030054
APA StyleIbrahim, A. Q., Abdullah, M. S., Ahram, M., & Abdalla, S. (2023). A Modified Protocol for the Isolation, Culture, and Characterization of Human Smooth Muscle Cells from the Umbilical Cord. Methods and Protocols, 6(3), 54. https://doi.org/10.3390/mps6030054