
Near-Infrared Spectroscopy (NIRS) as a Method for Biological Sex Discrimination in the Endangered Houston Toad (Anaxyrus houstonensis)

 ,
,
Abstract
:1. Introduction
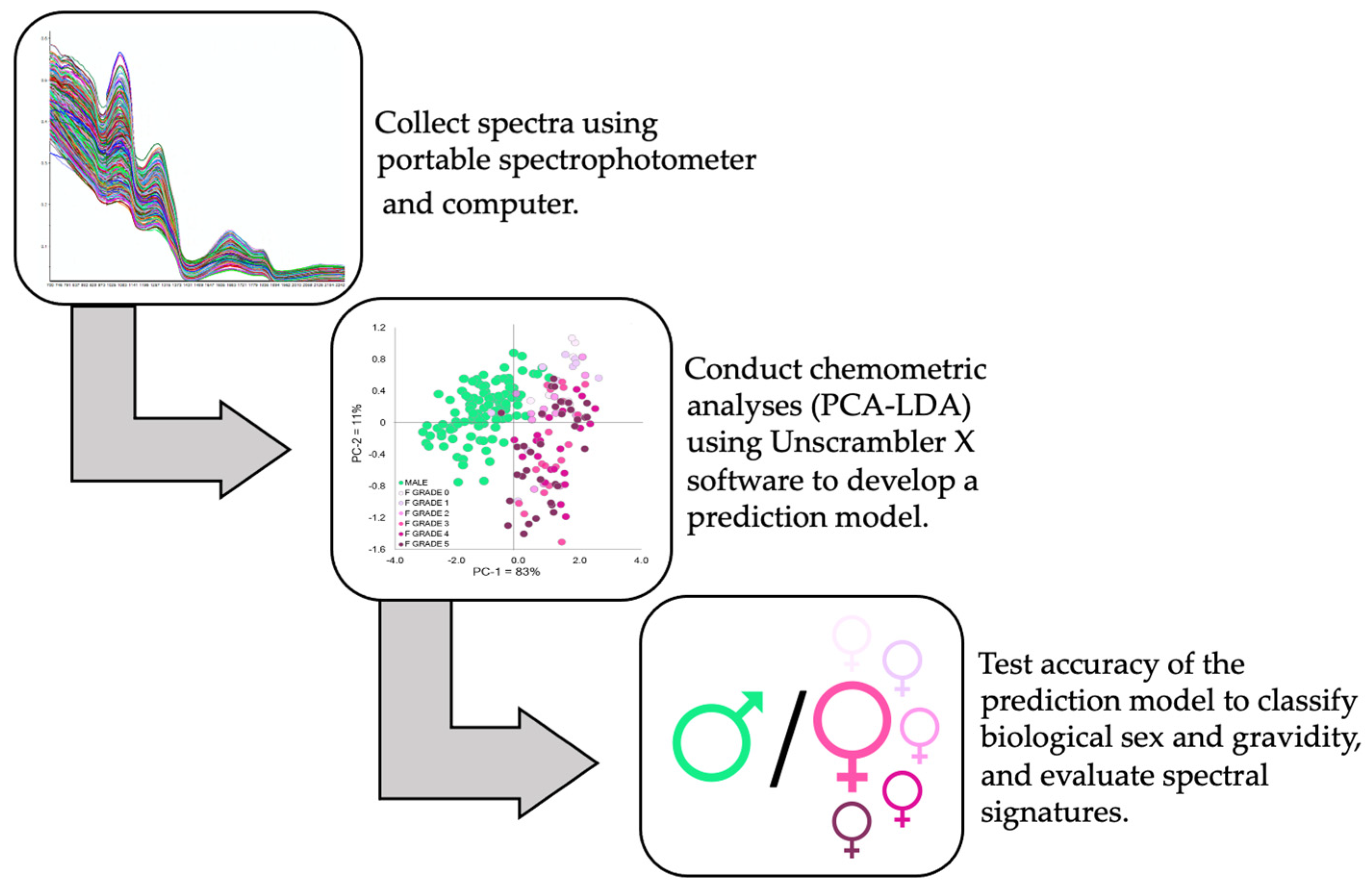
2. Materials and Methods
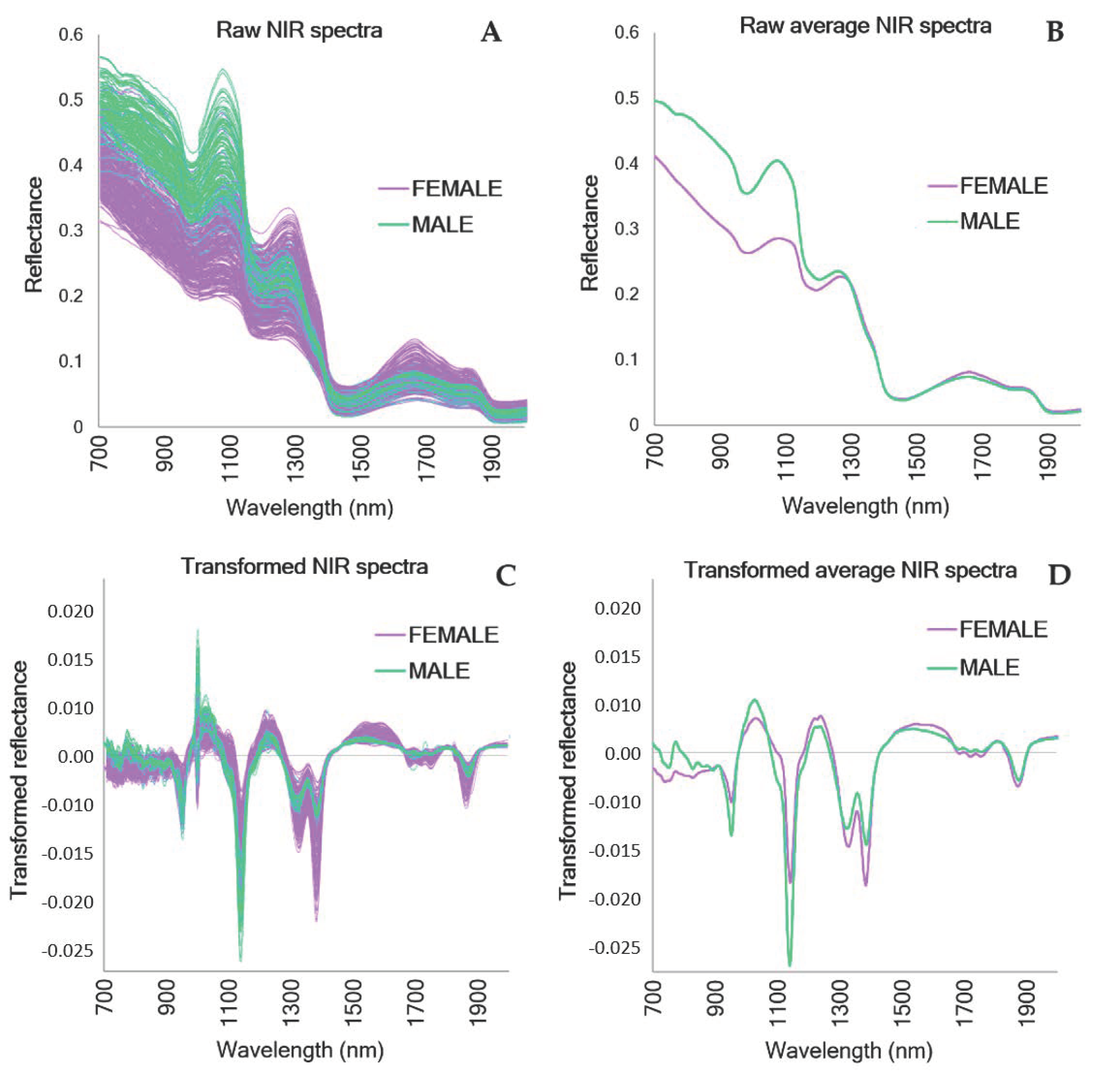
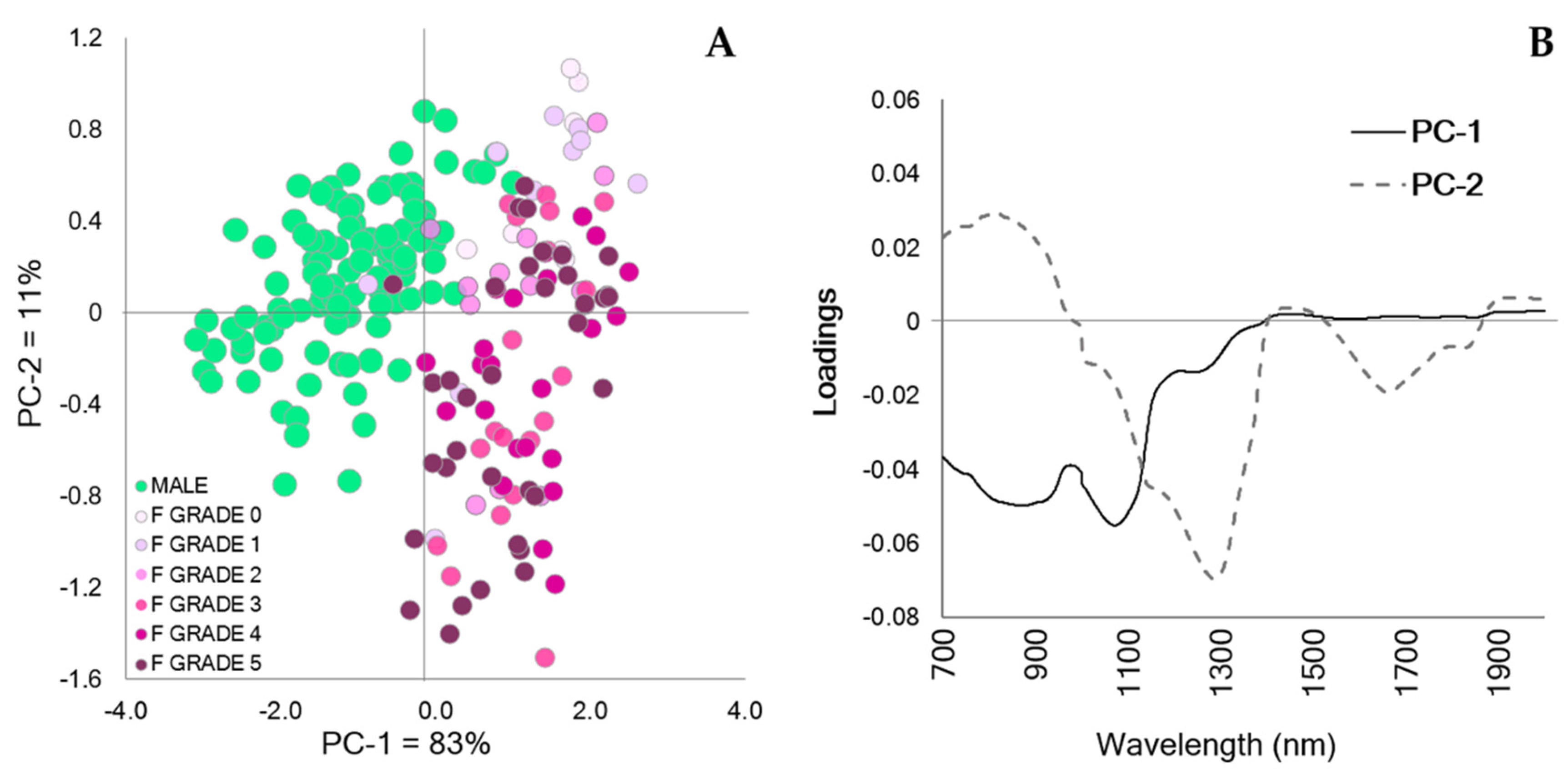
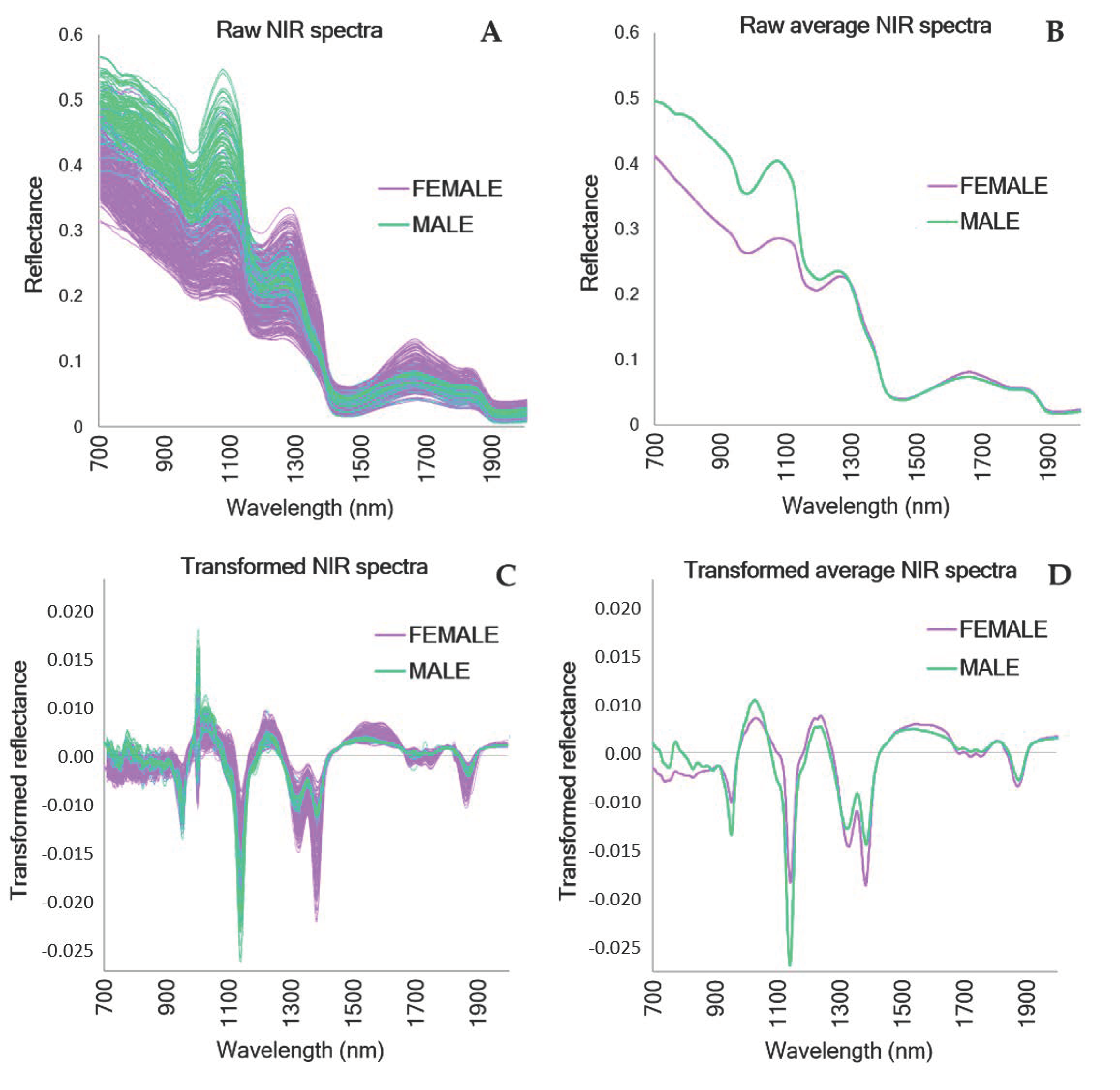
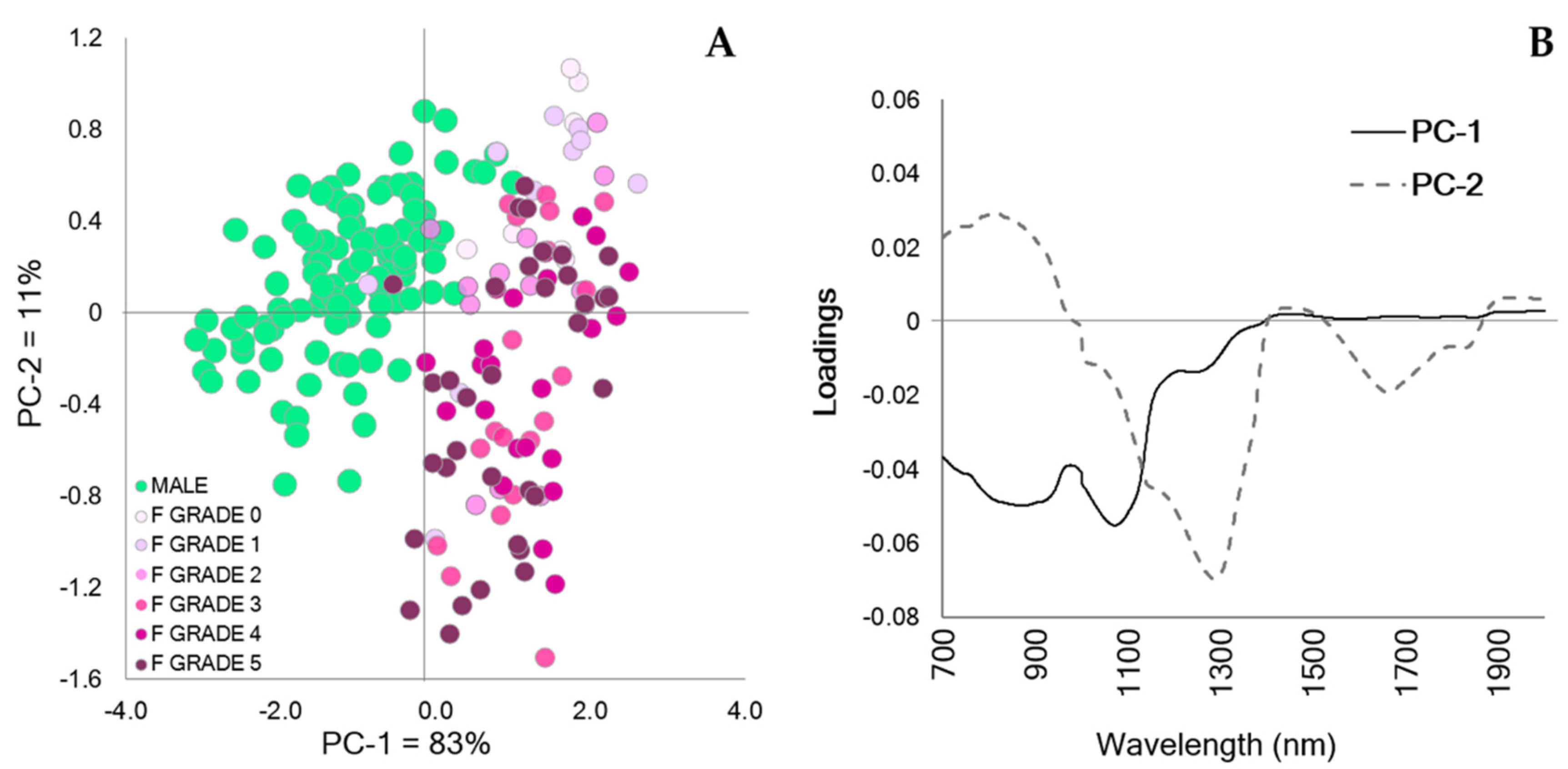
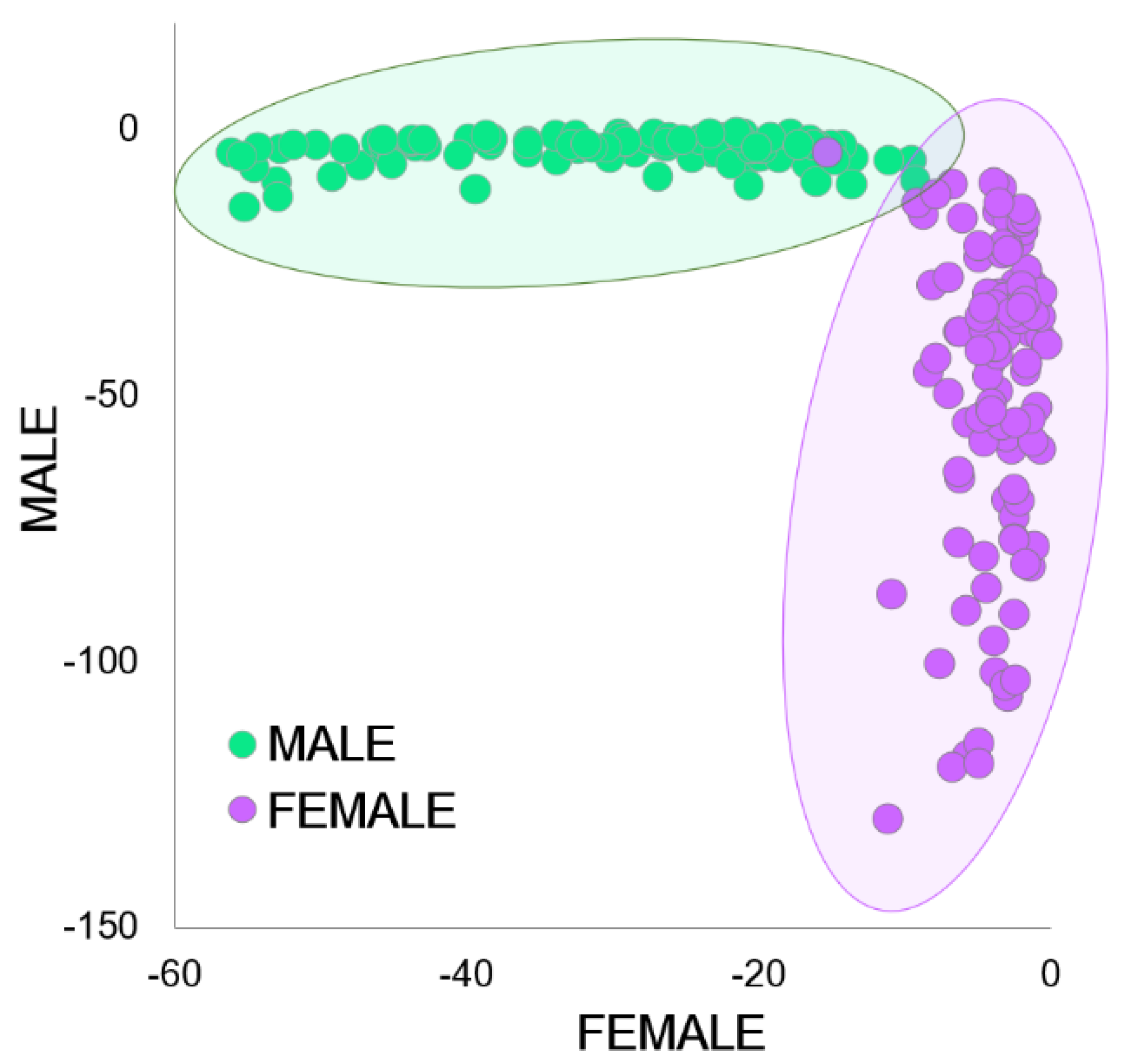
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 30 November 2021).
- Pievani, T. The sixth mass extinction: Anthropocene and the human impact on biodiversity. Rend. Lincei 2014, 25, 85–93. [Google Scholar] [CrossRef]
- Harfoot, M.B.J.; Johnston, A.; Balmford, A.; Burgess, N.D.; Butchart, S.H.M.; Dias, M.P.; Hazin, C.; Hilton-Taylor, C.; Hoffmann, M.; Isaac, N.J.B.; et al. Using the IUCN Red List to map threats to terrestrial vertebrates at global scale. Nat. Ecol. Evol. 2021, 5, 1510–1519. [Google Scholar] [CrossRef] [PubMed]
- Burger, I.; Julien, A.R.; Kouba, A.J.; Counsell, K.R.; Krebs, J.; Kouba, C.K. Linking in-situ and ex-situ populations of threatened amphibians through genome banking. Conserv. Sci. Pract. 2021, 3, e525. [Google Scholar] [CrossRef]
- Cayuela, H.; Besnard, A.; Bonnaire, E.; Perret, H.; Rivoalen, J.; Miaud, C.; Joly, P. To breed or not to breed: Past reproductive status and environmental cues drive current breeding decisions in a long-lived amphibian. Oecologia 2014, 176, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Wilczynski, W.; Allison, J.D.; Marler, C.A. Sensory pathways linking social and environmental cues to endocrine control regions of amphibian forebrains. Brain. Behav. Evol. 1993, 42, 252–264. [Google Scholar] [CrossRef]
- McDonough, C.E.; Martin, M.W.; Vance, C.K.; Cole, J.A.; Kouba, A.J. Frequency of exogenous hormone therapy impacts spermiation in male Fowler’s toad (Bufo fowleri). Reprod. Fertil. Dev. 2016, 28, 995–1003. [Google Scholar] [CrossRef]
- Guy, E.L.; Martin, M.W.; Kouba, A.J.; Cole, J.A.; Kouba, C.K. Evaluation of different temporal periods between hormone-induced ovulation attempts in the female Fowler’s toad Anaxyrus fowleri. Conserv. Physiol. 2020, 8, coz113. [Google Scholar] [CrossRef]
- Clulow, J.; Pomering, M.; Herbert, D.; Upton, R.; Calatayud, N.; Clulow, S.; Mahony, M.J.; Trudeau, V.L. Differential success in obtaining gametes between male and female Australian temperate frogs by hormonal induction: A review. Gen. Comp. Endocrinol. 2018, 265, 141–148. [Google Scholar] [CrossRef]
- Vance, C.K.; Kouba, A.J.; Willard, S.T. Near Infrared Spectroscopy Applications in Amphibian Ecology and Conservation: Gender and Species Identification. NIR News 2014, 25, 10–15. [Google Scholar] [CrossRef]
- Kupfer, A. Sexual size dimorphism in caecilian amphibians: Analysis, review and directions for future research. Zoology 2009, 112, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Graham, K.M.; Kouba, A.J.; Langhorne, C.J.; Marcec, R.M.; Willard, S.T. Biological sex identification in the endangered dusky gopher frog (Lithobates sevosa): A comparison of body size measurements, secondary sex characteristics, ultrasound imaging, and urinary hormone analysis methods. Reprod. Biol. Endocrinol. 2016, 14, 41. [Google Scholar] [CrossRef] [Green Version]
- Hogan, L.A.; Lisle, A.T.; Johnston, S.D.; Goad, T.; Robertston, H. Adult and juvenile sex identification in threatened monomorphic geocrinia frogs using fecal steroid analysis. J. Herpetol. 2013, 47, 112–118. [Google Scholar] [CrossRef]
- Germano, J.M.; Molinia, F.C.; Bishop, P.J.; Bell, B.D.; Cree, A. Urinary hormone metabolites identify sex and imply unexpected winter breeding in an endangered, subterranean-nesting frog. Gen. Comp. Endocrinol. 2012, 175, 464–472. [Google Scholar] [CrossRef]
- Ruiz-Fernández, M.J.; Jiménez, S.; Fernández-Valle, E.; García-Real, M.I.; Castejón, D.; Moreno, N.; González-Soriano, J. Sex Determination in Two Species of Anuran Amphibians by Magnetic Resonance Imaging and Ultrasound Techniques. Animals 2020, 10, 2142. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-García, A.; Roco, Á.S.; Bullejos, M. Sex Differentiation in Amphibians: Effect of Temperature and Its Influence on Sex Reversal. Sex. Dev. 2021, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Silla, A.; Kouba, A.; Heatwhole, H. Reproductive Technologies and Biobanking as Tools for the Conservation of Amphibians; CSIRO Publishing: Clayton, Australia, 2021; in press. [Google Scholar]
- Cozzolino, D. The sample, the spectra and the maths-The critical pillars in the development of robust and sound applications of vibrational spectroscopy. Molecules 2020, 25, 3674. [Google Scholar] [CrossRef]
- Grabska, J.; Huck, C.W. Near-Infrared Spectroscopy in Bio-Applications. Molecules 2020, 25, 2948. [Google Scholar]
- Osborne, B.G. Near-Infrared Spectroscopy in Food Analysis. In Encyclopedia of Analytical Chemistry: Applications, Theory and Instrumentation; 2000; pp. 1–14. [Google Scholar] [CrossRef]
- Sarraguça, M.C.; Lopes, J.A. Quality control of pharmaceuticals with NIR: From lab to process line. Vib. Spectrosc. 2009, 49, 204–210. [Google Scholar] [CrossRef]
- Vance, C.K.; Tolleson, D.R.; Kinoshita, K.; Rodriguez, J.; Foley, W.J. Near infrared spectroscopy in wildlife and biodiversity. J. Near Infrared Spectrosc. 2016, 24, 1–25. [Google Scholar] [CrossRef]
- Foley, W.J.; McIlwee, A.; Lawler, I.; Aragones, L.; Woolnough, A.P.; Berding, N. Ecological applications of near infrared reflectance spectroscopy—A tool for rapid, cost-effective prediction of the composition of plant and animal tissues and aspects of animal performance. Oecologia 1998, 116, 293–305. [Google Scholar] [CrossRef]
- Zouboulis, C.C. The skin as an endocrine organ. Derm. Endocrinol. 2009, 1, 250–252. [Google Scholar] [CrossRef]
- Hammerson, G.; Shepard, D. Anaxyrus houstonensis. Available online: https://www.iucnredlist.org/species/3170/9651352# (accessed on 30 November 2021).
- Gottschalk, J.S. United States list of endangered native fish and wildlife. Fed. Regist. 1970, 35, 16047–16048. [Google Scholar]
- Kennedy, J.P. Spawning season and experimental hybridization of the Houston toad, Bufo houstonensis. Herpetologica 1962, 17, 239–245. [Google Scholar]
- Moore, M.C. Application of organization-activation theory to alternative male reproductive strategies: A review. Syst. Biol. 1991, 25, 278–289. [Google Scholar] [CrossRef]
- Emerson, S.B. Phylogenies and physiological processes—The evolution of sexual dimorphism in Southeast Asian frogs. Syst. Biol. 1996, 45, 278–289. [Google Scholar] [CrossRef]
- Santos-Rivera, M.; Feeney, R.Z.; Julien, A.R.; Guy, E.; Gillis, A.; Zhang, H.X.; Kouba, A.J.; Vance, C. Gender discrimination using Near Infrared Reflectance (NIR) Spectroscopy in three caudate species. In Proceedings of the 19th International Council for Near Infrared Spectroscopy Conference (NIR-2019), Gold Coast, Australia, 15–20 September 2019; pp. 423–427. [Google Scholar]
- Belanger, R.M.; Corkum, L.D. Review of aquatic sex pheromones and chemical communication in anurans. J. Herpetol. 2009, 43, 184–191. [Google Scholar] [CrossRef]
- Jacobson, N.L. Breeding dynamics of the Houston toad. Southwest. Nat. 1989, 374–380. [Google Scholar] [CrossRef]
- Graham, K.M.; Langhorne, C.J.; Vance, C.K.; Willard, S.T.; Kouba, A.J. Ultrasound imaging improves hormone therapy strategies for induction of ovulation and in vitro fertilization in the endangered dusky gopher frog (Lithobates sevosa). Conserv. Physiol. 2018, 6, coy020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolleson, D.R.; Randel, R.D.; Stuth, J.W.; Neuendorff, D.A. Determination of sex and species in red and fallow deer by near infrared reflectance spectroscopy of the faeces. Small Rumin. Res. 2005, 57, 141–150. [Google Scholar] [CrossRef]
- Wiedower, E.E.; Kouba, A.J.; Vance, C.K.; Hansen, R.L.; Stuth, J.W.; Tolleson, D.R. Fecal near infrared spectroscopy to discriminate physiological status in giant pandas. PLoS ONE 2012, 7, e38908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canaza-Cayo, A.W.; Cozzolino, D.; Alomar, D.; Quispe, E. A feasibility study of the classification of Alpaca (Lama pacos) wool samples from different ages, sex and color by means of visible and near infrared reflectance spectroscopy. Comput. Electron. Agric. 2012, 88, 141–147. [Google Scholar] [CrossRef]
- Andueza, D.; Alabart, J.L.; Lahoz, B.; Muñoz, F.; Folch, J. Early pregnancy diagnosis in sheep using near-infrared spectroscopy on blood plasma. Theriogenology 2014, 81, 509–513. [Google Scholar] [CrossRef]
- Tallo-parra, O.; Albanell, E.; Carbajal, A.; Manteca, X.; Lopez-bejar, M.; Monclu, L. Prediction of Cortisol and Progesterone Concentrations in Cow Hair Using Near-Infrared Reflectance Spectroscopy (NIRS). Appl. Spectrosc. 2017, 71, 1954–1961. [Google Scholar] [CrossRef]
- Williams, P.; Manley, M.; Antoniszyn, J. Near Infrared Technology: Getting the Best out of Light; African Sun Media: Stellenbosch, South Africa, 2019. [Google Scholar]
- Hackelöer, B.J. The role of ultrasound in female infertility management. Ultrasound Med. Biol. 1984, 10, 35–50. [Google Scholar] [CrossRef]
- Silla, A.J.; Calatayud, N.E.; Trudeau, V.L. Amphibian reproductive technologies: Approaches and welfare considerations. Conserv. Physiol. 2021, 9, coab011. [Google Scholar] [CrossRef] [PubMed]
- Vance, C.K.; Kouba, A.J.; Zhang, H.-X.; Zhao, H.; Wang, Q.; Willard, S.T. Near Infrared Reflectance Spectroscopy Studies of Chinese Giant Salamanders in Aquaculture Production. NIR News 2015, 26, 4–7. [Google Scholar] [CrossRef]
- Passerotti, M.S.; Hesler, T.E.; Bensen, I.M.; Barnett, B.K.; Ballenger, J.C.; Bubley, W.J.; Reichert, M.J.M.; Quattro, J.M. Age estimation of red snapper (Lutjanus camperchanus) using FT-NIR spectroscopy: Feasibility of application to production ageing for management. J. Marine Sci. 2020, 77, 2144–2156. [Google Scholar] [CrossRef]
- Willaert, B.; Bossuyt, F.; Janssenswillen, S.; Adriaens, D.; Baggerman, G.; Matthijs, S.; Pauwels, E.; Proost, P.; Raepsaet, A.; Schoofs, L.; et al. Frog nuptial pads secrete mating season-specific proteins related to salamander pheromones. J. Exp. Biol. 2013, 216, 4139–4143. [Google Scholar] [CrossRef] [Green Version]
- Germano, J.M.; Cree, A.; Bishop, P.J. Ruling out the boys from the girls: Can subtle morphological differences identify sex of the apparently monomorphic frog, Leiopelma pakeka? N. Z. J. Zool. 2011, 38, 161–171. [Google Scholar] [CrossRef]
- Makowsky, R.; Wilson, L.A.; Pauley, T.K. Sexual Dimorphism in the Eastern Hellbender. Herpetol. Conserv. Biol. 2010, 5, 44–48. [Google Scholar]
- Kupfer, A. Sexual Size Dimorphism in Amphibians: An Overview. In Sex, Size and Gender Roles: Evolutionary Studies of Sexual Size Dimorphism; Fairbairn, D.J., Blanckenhorn, W.U., Székely, T., Eds.; Oxford University Press: New York, NY, USA, 2007; pp. 50–59. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | # Spectra | Cal 80% | Val 20% | Ext Val |
| Female | 243 | 107/108 (99.1%) | 26/27 (96.3%) | 95/108 (88.0%) |
| Male | 153 | 107/108 (99.1%) | 27/27 (100.0%) | 18/18 (100.0%) |
| Mean ± SD | 396 | 99.1 ± 0.0% | 98.1 ± 2.6% | 94.0 ± 8.5% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.-D.; Santos-Rivera, M.; Burger, I.J.; Kouba, A.J.; Barber, D.M.; Vance, C.K. Near-Infrared Spectroscopy (NIRS) as a Method for Biological Sex Discrimination in the Endangered Houston Toad (Anaxyrus houstonensis). Methods Protoc. 2022, 5, 4. https://doi.org/10.3390/mps5010004
Chen L-D, Santos-Rivera M, Burger IJ, Kouba AJ, Barber DM, Vance CK. Near-Infrared Spectroscopy (NIRS) as a Method for Biological Sex Discrimination in the Endangered Houston Toad (Anaxyrus houstonensis). Methods and Protocols. 2022; 5(1):4. https://doi.org/10.3390/mps5010004
Chicago/Turabian StyleChen, Li-Dunn, Mariana Santos-Rivera, Isabella J. Burger, Andrew J. Kouba, Diane M. Barber, and Carrie K. Vance. 2022. "Near-Infrared Spectroscopy (NIRS) as a Method for Biological Sex Discrimination in the Endangered Houston Toad (Anaxyrus houstonensis)" Methods and Protocols 5, no. 1: 4. https://doi.org/10.3390/mps5010004
APA StyleChen, L.-D., Santos-Rivera, M., Burger, I. J., Kouba, A. J., Barber, D. M., & Vance, C. K. (2022). Near-Infrared Spectroscopy (NIRS) as a Method for Biological Sex Discrimination in the Endangered Houston Toad (Anaxyrus houstonensis). Methods and Protocols, 5(1), 4. https://doi.org/10.3390/mps5010004