Differential Involvement of Hippocampal Angiotensin 1 Receptors in Learning and Memory Processes in Bulbectomized Rats
{kind=link}
{kind=link}
Abstract
:Introduction
Materials and methods
Animals
Surgical procedures
Behavioral methods
Statistical analysis
Results
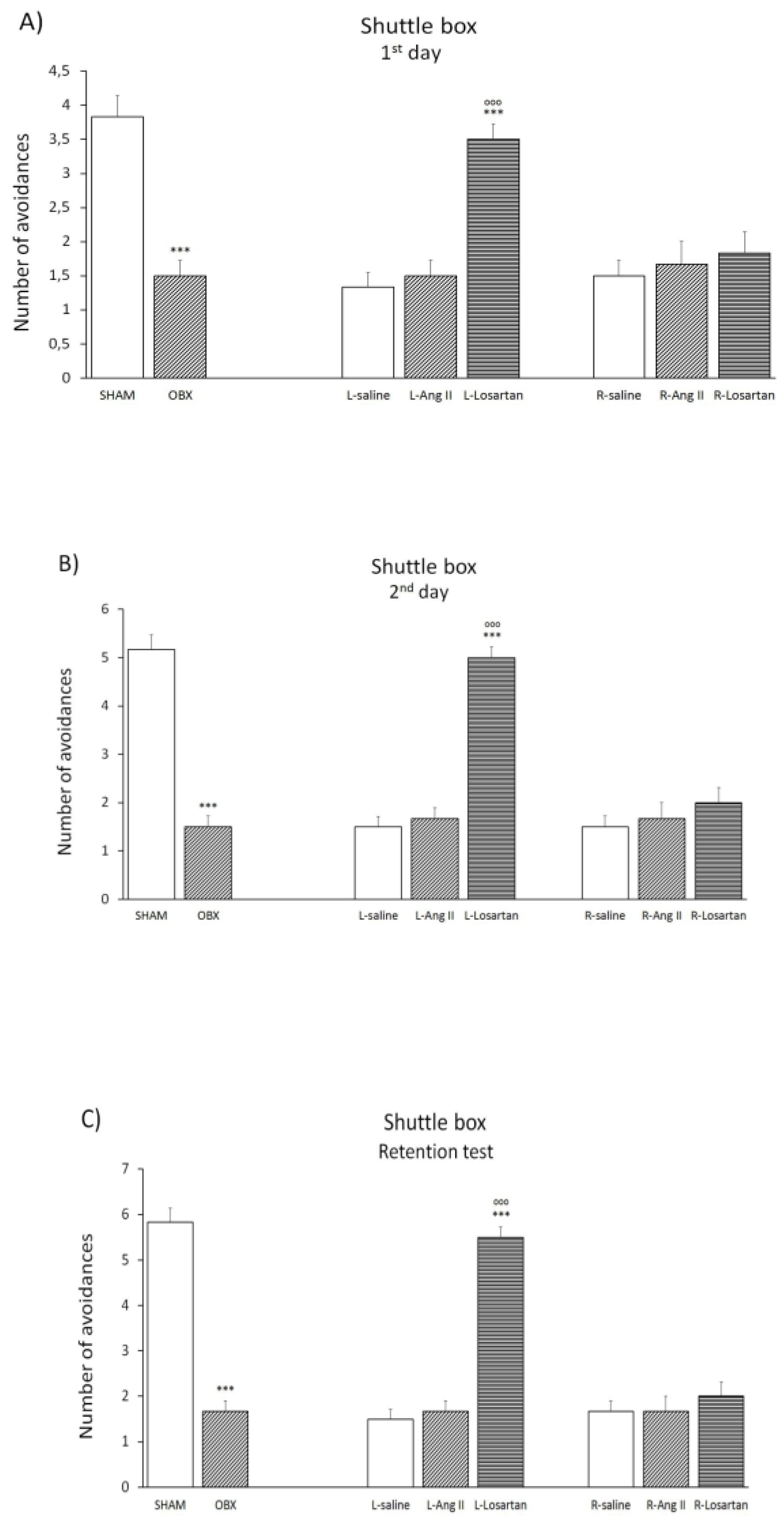
Shuttle box test
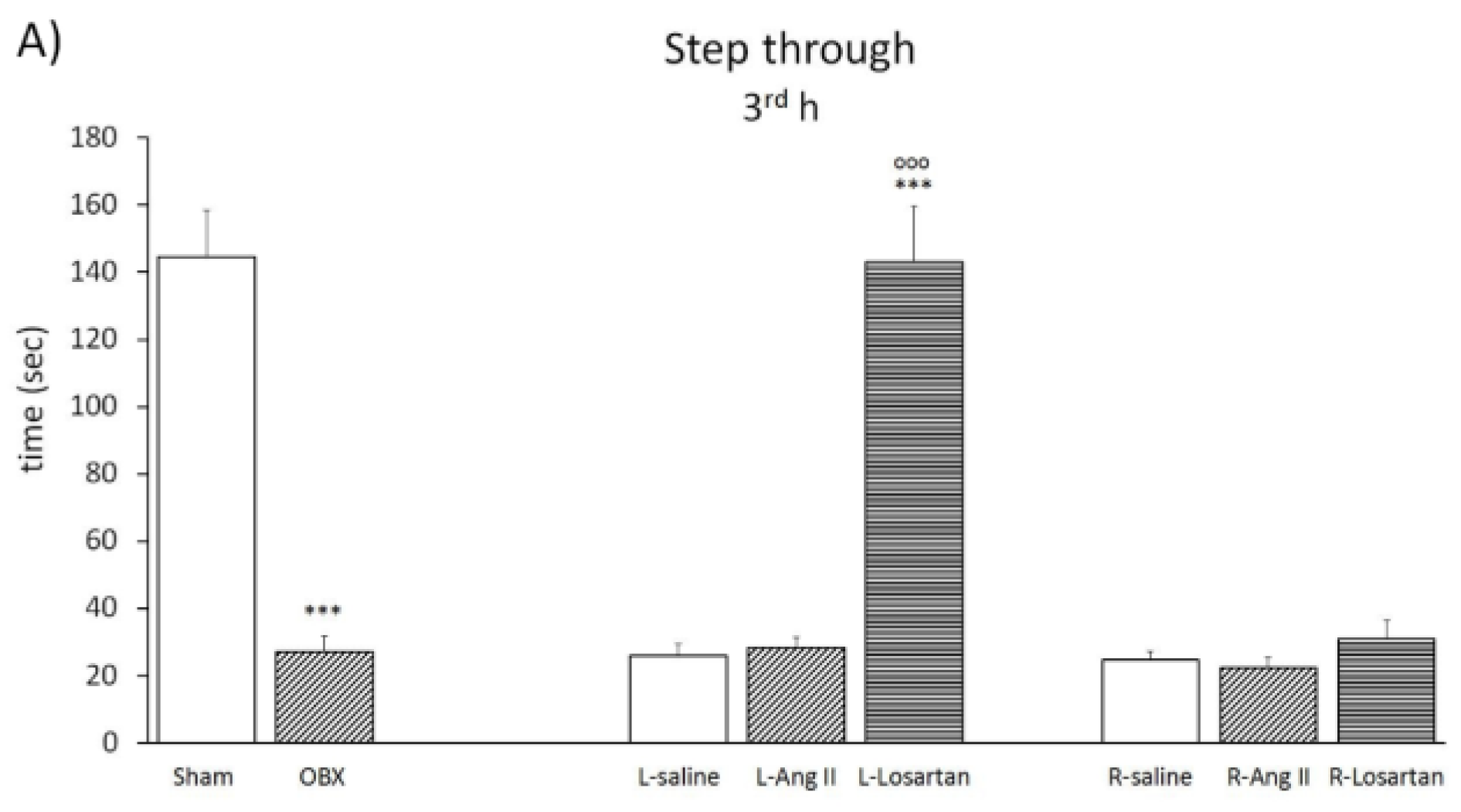
Step through test
Discussion
References
- von Bohlen und Halbach, O.; Albrecht, D. The CNS renin-angiotensin system. Cell Tissue Res. 2006, 326, 599–616. [Google Scholar] [CrossRef] [PubMed]
- de Gasparo, M.; Catt, K.J.; Inagami, T.; Wright, J.W.; Unger, T. International union of pharmacology. XXIII. The angiotensin II receptors. Pharmacol Rev. 2000, 52, 415–472. [Google Scholar] [PubMed]
- Santos, R.A.; Simoes e Silva, A.C.; Maric, C.; Silva, D.M.; Machado, R.P.; de Buhr, I.; Heringer-Walther, S.; Pinheiro, S.V.; Lopes, M.T.; Bader, M.; et al. Angiotensin-(1–7) is an endogenous ligand for the G protein- coupled receptor Mas. Proc. Natl. Acad. Sci USA. 2003, 100, 8258–8263. [Google Scholar] [CrossRef]
- Wright, J.W.; Harding, J.W. Brain angiotensin receptor subtypes AT1, AT2, and AT4 and their functions. Regul Pept. 1995, 59, 269–295. [Google Scholar] [CrossRef]
- von Bohlen und Halbach, O.; Albrecht, D. Mapping of angiotensin AT1 receptors in the rat limbic system. Regul Pept 1998, 78, 51–6. [Google Scholar] [CrossRef]
- Wright, J.W.; Harding, J.W. Brain renin- angiotensin--a new look at an old system. Prog Neurobiol. 2011, 95, 49–67. [Google Scholar] [CrossRef] [PubMed]
- Tashev, R.; Stefanova, M. Hippocampal asymmetry in angiotensin II modulatory effects on learning and memory in rats. Acta Neurobiol Exp. 2015, 75, 48–59. [Google Scholar] [CrossRef]
- de Souza, F.A.; Sanchis-Segura, C.; Fukada, S.Y.; de Bortoli, V.C.; Zangrossi HJr de Oliveira, A.M. Intracerebroventricular effects of angiotensin II on a step-through passive avoidance task in rats. Neurobiol Learn Mem. 2004, 81, 100–3. [Google Scholar] [CrossRef]
- Belcheva, I.; Ternianov, A.; Georgiev, V. Lateralized learning and memory effects ofangiotensin II microinjected into the rat CA1 hippocampal area. Peptides. 2000, 21, 407–411. [Google Scholar] [CrossRef]
- Belcheva, I.; Chobanova, M.; Georgiev, V. Differential behavioral effects of angiotensin II microinjected unilaterally into the CA1 hippocampal area. Regul Pept. 1998, 74, 67–71. [Google Scholar] [CrossRef]
- Belcheva, I.; Georgiev, V.; Chobanova, M.; Hadjiivanova, C. Behavioral effects of angiotensin II microinjected into CA1 hippocampal area. Neuropeptides. 1997, 31, 60–4. [Google Scholar] [CrossRef] [PubMed]
- Germain, L.; Chouinard, G. Captopril treatment of major depression with serial measurements of blood cortisol concentrations. Biol Psychiatry 1989, 25, 489–493. [Google Scholar] [CrossRef]
- Bondy, B.; Baghai, T.C.; Zill, P.; Schule, C.; Eser, D.; Deiml, T.; Zwanzger, P.; Ella, R.; Rupprecht, R. Genetic variants in the angiotensin I-converting-enzyme (ACE) and angiotensin II receptor (AT1) gene and clinical outcome in depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2005, 29, 1094–1099. [Google Scholar] [CrossRef] [PubMed]
- Heck, A.; Lieb, R.; Ellgas, A.; Pfister, H.; Lucae, S.; Erhardt, A.; Himmerich, H.; Horstmann, S.; Kloiber, S.; Ripke, S.; Müller-Myhsok, B.; Bettecken, T.; Uhr, M.; Holsboer, F.; Ising, M. Polymorphisms in the angiotensin- converting enzyme gene region predict coping styles in healthy adults and depressed patients. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2009, 150B, 104–114. [Google Scholar] [CrossRef]
- López-León, S.; Janssens, A.C.; Tiemeier, H.; Hofman, A.; Aulchenko, Y.S.; Snijders, P.J.; Claes, S.; Oostra, B.A.; van Duijn, C.M. Angiotensinogen M235T polymorphism and symptoms of depression in a population-based study and a family-based study. Psychiatr. Genet. 2008, 18, 162–166. [Google Scholar] [CrossRef]
- Wilson, W.; Voigt, P.; Bader, M.; Marsden, C.A.; Fink, H. Behaviour of the transgenic (mREN2)27 rat. Brain Res. 1996, 729, 1–9. [Google Scholar] [PubMed]
- Krsková, L.; Vrabcová, M.; Zeman, M. Effect of melatonin on exploration and anxiety in normotensive and hypertensive rats with high activity of renin-angiotensin system. Neuro Endocrinol. Lett. 2007, 28, 295–301. [Google Scholar]
- Saavedra, J.M.; Sánchez-Lemus, E.; Benicky, J. Blockade of brain angiotensin II AT1 receptors ameliorates stress, anxiety, brain inflammation and ischemia: therapeutic implications. Psychoneuroendocrinology. 2011, 36, 1–18. [Google Scholar] [CrossRef]
- Gard, P.R.; Mandy, A.; Sutcliffe, M.A. Evidence of a possible role of altered angiotensin function in the treatment, but not etiology, of depression. Biol. Psychiatry. 1999, 45, 1030–1034. [Google Scholar] [CrossRef]
- Weber, M.A. Angiotensin-II receptor blockers for hypertension and heart failure: quality of life and outcomes. Manag Care Interface. 2005, 18, 47–54. [Google Scholar]
- Pavlatou, M.G.; Mastorakos, G.; Lekakis, I.; Liatis, S.; Vamvakou, G.; Zoumakis, E.; Papassotiriou, I.; Rabavilas, A.D.; Katsilambros, N.; Chrousos, G.P. Chronic administration of an angiotensin II receptor antagonist resets the hypothalamic- pituitary-adrenal (HPA) axis and improves the affect of patients with diabetes mellitus type 2: preliminary results. Stress. 2008, 11, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Braszko, J.J.; Karwowska-Polecka, W.; Halicka, D.; Gard, P.R. Captopril and enalapril improve cognition and depressed mood in hypertensive patients. J. Basic Clin. Physiol. Pharmacol. 2003, 14, 323–343. [Google Scholar] [CrossRef]
- Hertzman, M.; Adler, L.W.; Arling, B.; Kern, M. Lisinopril may augment antidepressant response. J. Clin. Psychopharmacol. 2005, 25, 618–620. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.P.; Wrynn, A.S.; Leonard, B.E. The olfactory bulbectomized rat as a model of depression: An update. Pharmacol Ther. 1997, 74, 299–316. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Leonard, B.E. The olfactory bulbectomized rat as a model of depression. Neurosci Biobehav Rev. 2005, 29, 627–647. [Google Scholar] [CrossRef]
- Mucignat-Caretta, C.; Bondí, M.; Caretta, A. Time course of alterations after olfactory bulbectomy in mice. Physiol Behav. 2006, 89, 637–43. [Google Scholar] [CrossRef]
- Tashev, R.; Ivanova, M.; Toromanov, T.; Marinov, M.; Belcheva, S.; Belcheva, I. Olfactory bulbectomy impairs active and passive avoidance learning in rats. Compt. Rend. Acad. Bulg. Sci. 2010, 63, 617–620. [Google Scholar]
- Borre, Y.; Bosman, E.; Lemstra, S.; Westphal, K.G.; Olivier, B.; Oosting, R.S. Memantine partly rescues behavioral and cognitive deficits in an animal model of neurodegeneration. Neuropharmacology. 2012, 62, 2010–7. [Google Scholar] [CrossRef]
- Douma, T.N.; Borre, Y.; Hendriksen, H.; Olivier, B.; Oosting, R.S. Simvastatin improves learning and memory in control but not in olfactory bulbectomized rats. Psychopharmacology. 2012, 216, 537–44. [Google Scholar] [CrossRef]
- Yehuda, S.; Rabinovitz, S. Olfactory bulbectomy as a putative model for Alzheimer’: The protective role of essential fatty acids. Pharma Nutrition 2014, 2, 12–18. [Google Scholar] [CrossRef]
- Hozumi, S.; Nakagawasai, O.; Tan-No, K.; Niijima, F.; Yamadera, F.; Murata, A.; Arai, Y.; Yasuhara, H.; Tadano, T. Characteristics of changes in cholinergic function and impairment of learning and memory-related behavior induced by olfactory bulbectomy. Behav Brain Res. 2003, 138, 9–15. [Google Scholar] [CrossRef]
- Aleksandrova, I.; Kuyichkin, V.; Kashparov, I.; Medvinskaya, N.; Nesterova, I.; et al. Increased level of beta-amyloid in the brain of bulbectomized mice. Biochemistry (Mosc). 2004, 69, 176–80. [Google Scholar] [CrossRef]
- Pellegrino, L.; Cushman, A. A stereotaxic atlas of the rat brain; Appleton-Century-Crofts: New York, 1967. [Google Scholar]
- Morgan, J.M.; Routtenberg, A. Ang II injected into the neostriatum after learning disrupts retention performance. Science. 1977, 196, 87–9. [Google Scholar] [CrossRef] [PubMed]
- Braszko, J.J.; Wiœniewski, K. Effect of angiotensin II and saralasin on motor activity and the passive avoidance behavior of rats. Peptides. 1988, 9, 475–9. [Google Scholar] [CrossRef] [PubMed]
- Braszko, J.J. AT(2) but not AT(1)receptor antagonism abolishes angiotensin II increase of the acquisition of conditioned avoidance responses in rats. Behavioural Brain Research 2002, 131, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Karwowska-Polecka, W.; Kułakowska, A.; Wiśniewski, K.; Braszko, J.J. Losartan influences behavioural effects of angiotensin II (3-7) in rats. Pharmacol Res. 1997, 36, 275–83. [Google Scholar] [CrossRef]
- Wright, J.; Miller-Wing, A.; Shaffer, M.; Higginson, C.; Wright, D.; Hanesworth, J.; Harding, J. Angiotensin II(3-8) (ANG IV) hippocampal binding: potential role in the facilitation of memory. Brain Res Bull. 1993, 32, 497–502. [Google Scholar] [CrossRef]
- Kerr, D.; Bevilaqua, L.; Bonini, J.; Rossato, J.; Köhler, C.; Medina, J.; Izquierdo, I.; Cammarota, M. Angiotensin II blocks memory consolidation through an AT2 receptor-dependent mechanism. Psychopharmacology (Berl). 2005, 179, 529–35. [Google Scholar] [CrossRef]
- Bonini, J.S.; Bevilaqua, L.R.; Zinn, C.G.; Kerr, D.S.; Medina, J.H.; Izquierdo, I.; Cammarota, M. Angiotensin II disrupts inhibitory avoidance memory retrieval. HormBehav. 2006, 50, 308–13. [Google Scholar] [CrossRef]
- Braszko, J. The contribution of AT1 and AT2 angiotensin receptors to its cognitive effects. Acta Neurobiol Exp (Wars). 1996, 56, 49–54. [Google Scholar] [CrossRef]
- Kułakowska, A.; Karwowska, W.; Wiśniewski, K.; Braszko, J.J. Losartan influences behavioural effects of angiotensin II in rats. Pharmacological Research. 1996, 34, 109–15. [Google Scholar] [CrossRef] [PubMed]
- Raghavendra, V.; Chopra, K.; Kulkarni, S.K. Comparative studies on the memory enhancing actions of captopril and losartan in mice using inhibitory shock avoidance paradigm. Neuropeptides. 2001, 35, 65–9. [Google Scholar] [CrossRef] [PubMed]
- DeNoble, V.J.; DeNoble, K.F.; Spencer, K.R.; Chiu, A.T.; Wong, P.C.; Timmermans, B.M. Non-peptide angiotensin II receptor antagonist and angiotensin-converting enzyme inhibitor: effect on a rennin induced deficit of a passive avoidance response in rats. Brain Res. 1991, 561, 230–5. [Google Scholar] [CrossRef] [PubMed]
- Du, G.T.; Hu, M.; Mei, Z.L.; Wang, C.; Liu, G.J.; Hu, M.; Long, Y.; Miao, M.X.; Chang Li, J.; Hong, H. Telmisartan treatment ameliorates memory deficits in streptozotocin-induced diabetic mice via attenuating cerebral amyloidosis. J Pharmacol Sci. 2014, 124, 418–26. [Google Scholar] [CrossRef]
- Mogi, M.; Li, J.M.; Tsukuda, K.; Iwanami, J.; Min, L.J.; Sakata, A.; Fujita, T.; Iwai, M.; Horiuchi, M. Telmisartan prevented cognitive decline partly due to PPAR- gamma activation. Biochem Biophys Res Commun. 2008, 375, 446–9. [Google Scholar] [CrossRef]
- Inaba, S.; Iwai, M.; Furuno, M.; Tomono, Y.; Kanno, H.; Senba, I.; Okayama, H.; Mogi, M.; Higaki, J.; Horiuchi, M. Continuous activation of renin-angiotensin system impairs cognitive function in renin/angiotensinogen transgenic mice. Hypertension. 2009, 53, 356–62. [Google Scholar] [CrossRef]
- Tedesco, M.A.; Ratti, G.; Mennella, S.; Manzo, G.; Grieco, M.; Rainone, A.C.; Iarussi, D.; Iacono, A. Comparison of losartan and hydrochlorothiazide on cognitive function and quality of life in hypertensive patients. Am J Hypertens. 1999, 12, 1130–1134. [Google Scholar] [CrossRef]
- Fogari, R.; Mugellini, A.; Zoppi, A.; Derosa, G.; Pasotti, C.; Fogari, E.; Preti, P. Influence of losartan and atenolol on memory function in very elderly hypertensive patients. Journal of Human Hypertension. 2003, 17, 781–785. [Google Scholar] [CrossRef]
- Ge, J.; Barnes, N.M. Alterations in angiotensin AT1 and AT2 receptor subtype levels in brain regions from patients with neurodegenerative disorders. Eur J Pharmacol. 1996, 297, 299–306. [Google Scholar] [CrossRef]
- Savaskan, E.; Hock, C.; Olivieri, G.; Bruttel, S.; Rosenberg, C.; Hulette, C.; Müller-Spahn, F. Cortical alterations of angiotensin converting enzyme, angiotensin II and AT1 receptor in Alzheimer’s dementia. Neurobiol Aging. 2001, 22, 541–546. [Google Scholar] [CrossRef]
- Tasset, I.; Medina, F.J.; Peña, J.; Jimena, I.; Del Carmen Muñoz, M.; et al. Olfactory bulbectomy induced oxidative and cell damage in rat: protective effect of melatonin. Physiol Res. 2010, 59, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Myint, A.M.; Steinbusch, H.W.; Goeghegan, L.; Luchtman, D.; Kim, Y.K.; Leonard, B.E. Effect of the COX-2 inhibitor celecoxib on behavioural and immune changes in an olfactory bulbectomized rat model of depression. Neuroimmunomodulation 2007, 14, 65–71. [Google Scholar] [CrossRef]
- Rinwa, P.; Kumar, A. Quercetin suppress microglial neuroinflammatory response and induce antidepressant-like effect in olfactory bulbectomized rats. Neuroscience 2013, 255, 86–98. [Google Scholar] [CrossRef]
- Song, C.; Zhang, X.Y.; Manku, M. Increased phospholipase A2 activity and inflammatory response but decreased nerve growth factor expression in the olfactory bulbectomized rat model of depression: effects of chronic ethyl-eicosapentaenoate treatment. J Neurosci. 2009, 29, 14–22. [Google Scholar] [CrossRef]
- Saavedra, J. Angiotensin II AT(1) receptor blockers as treatments for inflammatory brain disorders. Clin. Sci. (Lond). 2012, 123, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Pang, T.; Hafko, R.; Benicky, J.; Sanchez-Lemus, E.; Saavedra, J.M. Telmisartan ameliorates glutamate-induced neurotoxicity: roles of AT(1) receptor blockade and PPARγ activation. Neuropharmacology. 2014, 79, 249–61. [Google Scholar] [CrossRef] [PubMed]
- Villapol, S.; Yaszemski, A.K.; Logan, T.T.; Sánchez-Lemus, E.; Saavedra, J.M.; Symes, A.J. Candesartan, an angiotensin II AT₁- receptor blocker and PPAR-γ agonist, reduces lesion volume and improves motor and memory function after traumatic brain injury in mice. Neuropsychopharmacology. 2012, 37, 2817–29. [Google Scholar] [CrossRef]
- Timaru-Kast, R.; Wyschkon, S.; Luh, C.; Schaible, E.V.; Lehmann, F.; Merk, P.; Werner, C.; Engelhard, K.; Thal, S.C. Delayed inhibition of angiotensin II receptor type 1 reduces secondary brain damage and improves functional recovery after experimental brain trauma. Crit Care Med. 2012, 40, 935–944. [Google Scholar] [CrossRef]
- Kasahara, Y.; Taguchi, A.; Uno, H.; Nakano, A.; Nakagomi, T.; Hirose, H.; Stern, D.M.; Matsuyama, T. Telmisartan suppresses cerebral injury in a murine model of transient focal ischemia. Brain Res. 2010, 1340, 70–80. [Google Scholar] [CrossRef]
- Jung, K.; Chu, K.; Lee, S.; Kim, S.; Song, E.; Kim, E.; Park, D.; Sinn, D.; Kim, J.; Kim, M.; Roh, J. Blockade of AT1 receptor reducesapoptosis, inflammation, and oxidative stress in normotensive rats with intracerebral hemorrhage. J Pharmacol Exp Ther. 2007, 322, 1051–8. [Google Scholar] [CrossRef]
- Benicky, J.; Sánchez-Lemus, E.; Honda, M.; Pang, T.; Orecna, M.; Wang, J.; Leng, Y.; Chuang, D.M.; Saavedra, J.M. Angiotensin II AT1 receptor blockade ameliorates brain inflammation. Neuropsychopharmacology. 2011, 36, 857–70. [Google Scholar] [CrossRef] [PubMed]
- Benicky, J.; Sánchez-Lemus, E.; Pavel, J.; Saavedra, J.M. Anti-inflammatory effects of angiotensin receptor blockers in the brain and the periphery. Cell MolNeurobiol. 2009, 29, 781–92. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, Y.; Hosoya, A.; Yamasaki, N.; Ahmed, H.; Hattori, S.; Eguchi, M.; Yamaguchi, S.; Miyakawa, T.; Hirase, H.; Shigemoto, R. Right-hemispheric dominance of spatial memory in split-brain mice. Hippocampus. 2012, 22, 117–21. [Google Scholar] [CrossRef]
- Shipton, O.A.; El-Gaby, M.; Apergis-Schoute, J.; Deisseroth, K.; Bannerman, D.M.; Paulsen, O.; Kohl, M.M. Left–right dissociation of hippocampal memory processes in mice. Proc Natl AcadSci USA. 2014, 111, 15238–15243. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.M.; Wang, C.; Wang, X.L.; Wang, L.; Chang, C.W.; Wang, P.; Gao, G.D. Correlations between angiotensinase activity asymmetries in the brain and paw preference in rats. Neuropeptides. 2010, 44, 253–9. [Google Scholar] [CrossRef]
- Banegas, I.; Prieto, I.; Alba, F.; Vives, F.; Araque, A.; Segarra, A.B.; Durán, R.; de Gasparo, M.; Ramírez, M. Angiotensinase activity is asymmetrically distributed in the amygdala, hippocampus and prefrontal cortex of the rat. Behav Brain Res. 2005, 156, 321–6. [Google Scholar] [CrossRef]
- Tashev, R.; Ivanova, M. Lateralized hippocampal effects of angiotensin IV on learning and memory in rats. Compt. Rend L'Acad Bulg Sci. 2015, 68, 1309–1316. [Google Scholar]
© 2016 by the authors. 2016 Bogdan Popescu, Oana Păun, Răzvan V. Scăunașu, Cristian Bălălău, Șerban V. Berteșteanu. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roman, T.E.; Ivanova, M.S.; Belcheva, S.P.; Belcheva, I.P. Differential Involvement of Hippocampal Angiotensin 1 Receptors in Learning and Memory Processes in Bulbectomized Rats. J. Mind Med. Sci. 2016, 3, 39-54. https://doi.org/10.22543/2392-7674.1036
Roman TE, Ivanova MS, Belcheva SP, Belcheva IP. Differential Involvement of Hippocampal Angiotensin 1 Receptors in Learning and Memory Processes in Bulbectomized Rats. Journal of Mind and Medical Sciences. 2016; 3(1):39-54. https://doi.org/10.22543/2392-7674.1036
Chicago/Turabian StyleRoman, Tashev E., Margarita S. Ivanova, Stiliana P. Belcheva, and Iren P. Belcheva. 2016. "Differential Involvement of Hippocampal Angiotensin 1 Receptors in Learning and Memory Processes in Bulbectomized Rats" Journal of Mind and Medical Sciences 3, no. 1: 39-54. https://doi.org/10.22543/2392-7674.1036
APA StyleRoman, T. E., Ivanova, M. S., Belcheva, S. P., & Belcheva, I. P. (2016). Differential Involvement of Hippocampal Angiotensin 1 Receptors in Learning and Memory Processes in Bulbectomized Rats. Journal of Mind and Medical Sciences, 3(1), 39-54. https://doi.org/10.22543/2392-7674.1036