Abstract
Climate change has intensified the increase in irrigated crops to solve frequent droughts, but part of the stress continues due to heat waves, and for these systems, there is a lack of in-depth discussion about their damage and strategies to minimize this damage. The damage caused by high temperatures may be exacerbated in crops with a nutritional disorder of nitrogen, but optimized management of this nutrient can help mitigate the effects of this stress. This merits further debate, as it would be a sustainable strategy without risk to the environment and at the same time could induce greater plant tolerance to heat stress. This review will address the relevance of rising temperatures and their isolated effects on crop nutrition and productivity and the role of optimized nitrogen use in mitigating this stress and propose future perspectives for this research that could help researchers improve thermotolerance. Nitrogen plays an essential role in plant metabolism, inducing the production of proteins from photosynthesis, boosting primary and secondary plant metabolism and, consequently, the production and defense systems of the plant. Studies indicate that adequate nitrogen supplementation can increase plant resilience to high temperatures, improving water use efficiency and promoting the synthesis of heat shock proteins. In addition, new strategies in nitrogen fertilizer management, such as the use of nitrification inhibitors and biofertilizers, have demonstrated greater efficiency in the use of nitrogen, reducing environmental losses, and, consequently, they could have an impact on stress mitigation. Although nitrogen can mitigate the negative effects of heat waves on plants, there are still gaps in the knowledge about the underlying biochemical and physiological mechanisms involved and whether the doses of N used in research are really optimal for maximizing the plant’s defense system against stress. The future outlook is that optimal N management will become important not only to guarantee high yields but also to minimize heat wave losses by strengthening the plant’s defense mechanisms to deal with this stress. In the context of climate change, a better understanding of the benefits of N will help to better manage heat stress towards more sustainable agriculture.
1. Introduction
Climate change has intensified globally, causing severe impacts on agriculture, especially through heat waves that damage the productivity of various crops [1]. Modern agriculture faces the challenge of maintaining or increasing productivity in increasingly adverse environmental conditions, including abiotic stresses such as drought, salinity, and, especially, extreme heat. This phenomenon has been a growing concern for farmers and scientists, as prolonged exposure to high temperatures can cause irreversible physiological damage to plants, resulting in lower crop yield and quality [2]. Each species has an optimal temperature range for growth and development, and with climate change, an increase of up to 2 °C or more in the global average temperature is predicted in the coming decades [1,3]. Therefore, when this increase can exceed the maximum tolerated limit of the species depending on the growing region, heat stress occurs, negatively impacting crop productivity and quality.
In this context, proper management of plant nutrition has emerged as one of the main strategies for mitigating the negative effects of heat waves on agricultural productivity. Among the essential elements for plant growth, development, and production, nitrogen plays a crucial role, being one of the main elements responsible for the synthesis of hundreds of vital organic compounds, such as proteins, nucleic acids, and chlorophyll [4,5]. The relationship between nitrogen nutrition and the ability of plants to tolerate heat stress has attracted increasing attention from researchers because it can help maintain essential physiological functions, such as photosynthesis and water use efficiency [6]. An increase in air temperature will induce heat stress in plants only when the air temperature exceeds the upper threshold temperature or upper thermal limit, which vary between species or cultivars and between the age and phenological stage of the plant, and it can cause the leaf expansion and vegetative development of plants to come to a standstill or to reduce to a very low level [7]. It should be noted, however, that the upper base temperature is unlikely to be reached under field conditions, but with the acceleration in the frequency of climate change, this is being modified. However, there are cases in which an increase in air temperature, if it does not reach the upper thermal threshold of the species studied and depending on the growing region, can be beneficial for the plant.
Nitrogen metabolism is intimately linked to plant responses to thermal stress. Under high-temperature conditions, plants activate defense mechanisms, such as the production of heat shock proteins (HSPs) and the activation of the antioxidant system, which help protect cells from heat-induced damage [8]. Nitrogen plays a fundamental role in these processes, as it is essential for the synthesis of amino acids—precursors of HSPs—and for maintaining redox balance, which is crucial for the activity of antioxidant enzymes [9,10]. Moreover, adequate N availability can improve cellular membrane stability and photosynthetic efficiency, reducing the negative effects of thermal stress by reinforcing the plant’s defense system [6].
Although there are several studies exploring the interaction between nitrogen nutrition and heat stress conditions, many gaps remain regarding the underlying biochemical and physiological mechanisms. In addition, the optimization of nitrogen fertilization practices, aimed at increasing plant resilience in a climate change scenario, requires further investigation, especially with the use of improved sources of controlled release fertilizers, nitrification inhibitors and the use of biofertilizers. However, basic information on N management, such as timing of application and appropriate doses, is the fruit of research that needs to be updated for each species and growing region in different provinces and countries [4] and, consequently, will ensure optimum plant nutrition, including N, enhancing the benefits of stress mitigation.
In this scenario, it is important to have a more in-depth discussion about the response of plants when subjected to high temperatures and the progress of research indicating the role of optimal N in the plant in increasing the tolerance of crops subjected to high temperatures, with the other production factors being optimized. With the pressure of climate change, the use of irrigation in crops has increased, but this is not enough to resolve this pressure, due to the increases in air temperature. This problem can be aggravated in crops with N deficiency, which is common in the world’s different soils.
This review should also help to identify gaps in the role of N in mitigating high temperatures in irrigated crops, and it discusses the challenges and opportunities to be employed in order to contribute to the development of more resilient and sustainable agricultural practices.
2. Advances in Abiotic Stresses in Agriculture: High Temperature Alone, Without Additional Stress
Defined as exposure to temperatures above the upper threshold temperature or upper thermal limit for plant development, high-temperature-induced thermal stress results in the inhibition of photosynthesis, damage to cellular membranes, and impairment of essential metabolic activities, which lead to lower growth rates and, consequently, reduced productivity [11]. In a high-carbon-emissions scenario, global temperatures could rise between 3.3 °C and 5.7 °C by the end of the century [1]. To put the impact of this projection into context, the last time the Earth experienced a temperature rise of 2.5 °C above pre-industrial levels was more than three million years ago. The State of the Global Climate 2023 report describes that 2023 was the hottest year ever recorded, with the global average near-surface temperature 1.45 °C above pre-industrial levels [12,13]. In addition, the report estimates an 80% probability that in at least one of the next five years, the annual global temperature will temporarily exceed the 1.5 °C limit, reinforcing the urgency of mitigating climate impacts for agriculture and other vulnerable sectors, especially due to the intensification of extreme events such as heat waves and droughts (Figure 1).
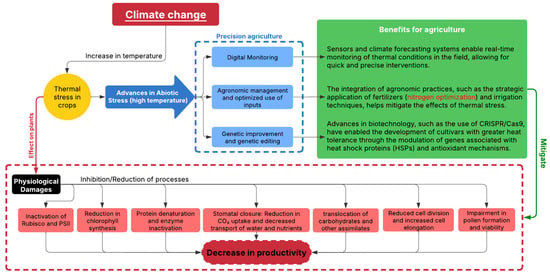
Figure 1.
Integrated scheme on high-air-temperature-induced thermal stress, its effects on plants, and research advances to mitigate stress impacts, thereby strengthening agricultural systems.
Cleary defining the potential behavior of air temperature in future scenarios is crucial for determining the research needed to mitigate future damages and to estimate physiological and nutritional losses in plants. To this end, predictive model data available in global digital climate atlases, such as WorldClim and CHELSA, can be utilized. For example, projections from the study by Kufa et al. (2022) [14] indicate that, in Representative Concentration Pathway (RCP) scenarios—specifically RCP 2.6 and RCP 8.5—air temperatures are expected to increase by approximately +2 °C to +6 °C by mid-century. Similarly, the holistic approach described by Illangasingha et al. (2023) [15], using results from Global Climate Models (GCMs), confirms that warming trends are robust across different regions, with substantial increases in both mean and extreme temperatures. These models are fed by extensive datasets from satellites, weather stations, and historical climate records; their results are rigorously downscaled and bias-corrected, ensuring that the projected temperature increases are reliable for evaluating plant responses [14,15]. Based on these projections, the potential response of plants under elevated temperatures is defined as one that may include root expansion, limited nutrient mineralization, and impaired enzymatic activities crucial for N assimilation. These changes lead to reduced photosynthetic efficiency and overall plant growth. However, optimal N fertilization can mitigate these negative effects by enhancing protein synthesis, activating antioxidant defenses, and stabilizing cellular membranes and plant metabolism.
An innovative system for carrying out these studies would be the T-FACE (Temperature–Free-Air-Controlled Enhancement) system, which is an innovative tool that simulates, in an open field, the impacts of temperature increases predicted for the future, making it possible to study plant responses under realistic conditions. Recent studies carried out in Brazil with the forage species Stylosantes capitata Vogel that used T-FACE systems to simulate temperature increases in the field showed that an increase of 2 °C led to an increase in leaf nutrient content, nutrient use efficiency, and biomass allocation in all aerial organs of this forage under well-watered conditions [16,17,18] and favored biological fixation and soil microbial activity, highlighting the adaptation potential of this species [11].
This scenario of a 2 °C increase in ambient temperature would be advantageous for forage production in tropical regions in the future, if proper irrigation management is maintained [19]. However, most studies still use fixed temperatures that do not fluctuate over time, which is normal for rain and/or cloudy days, for example, which limits the extrapolation of results to real climate change scenarios.
In general, in other species, high air temperature is an abiotic stress that represents one of the greatest threats to agricultural productivity, especially if there are associated stresses such as drought and salinity, among others. Stresses in the field can occur in isolation or in combination, so in this type of review the main objective would be to focus on the isolated stress of high temperature and, consequently, whether the other production factors are at optimum levels for the crop.
This type of approach is especially important in intensive field cultivation systems that use all the advanced technologies to maintain optimum water levels in the soil from irrigated systems, with adequate fertilization and phytosanitary management, but unfortunately, it is not possible to control the air temperature, and knowledge of this effect becomes relevant in these cultivation systems. Irrigated areas in the world are increasing in the face of climate change and with the use of high technology, currently covering around 340 million hectares, with projections of expansion that could reach up to 450 million hectares by 2050 to meet growing food demands and mitigate climate stresses [12].
The impact of these factors on plants, depending on the temperature and species, is drastic because it affects essential physiological processes such as photosynthesis, transpiration, and cell metabolism [20]. Heat stress, defined as an increase in ambient temperature beyond optimal levels for plant growth and development [21], induces vulnerability in crops [22]. Recent findings indicate a 4.4% decline in yield for every 1 °C increase beyond 27 °C, with japonica ecotypes exhibiting high sensitivity to air temperature [23], but this is little known across species
The increase in air temperature, especially at night, has a significant impact on a plant’s physiological processes. During the night, there is an increase in the rate of respiration and the plant’s energy expenditure, using a greater part of the photosynthesis products for cell maintenance rather than growth, consequently decreasing grain yield [24]. In addition, this increase in temperature affects the translocation of carbohydrates and nitrogen, decreasing the filling rate and grain mass by intensifying the production of reactive oxygen species (ROS), causing permanent damage to cell membranes by oxidizing lipids, proteins, and nucleic acids [24].
Plants respond to this accumulation of ROS by activating enzymatic antioxidant systems such as superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX) [9] and non-enzymatic systems that help protect proteins and cell membranes from oxidative and heat stress [25]. A plant’s non-enzymatic mechanism for mitigating heat stress occurs by activating complex signaling networks, which include HSPs, transcription factors, and plant hormones. HSPs play a fundamental role in protecting cellular proteins from heat-induced denaturation, helping to maintain proper protein conformation and preventing protein aggregation [26]. Among the transcription factors activated by heat are the Heat Shock Factors (HSFs), which regulate the expression of genes involved in plant adaptation to heat stress [8,27]. Thus, the regulatory function of HSFs is fundamental for the induction of HSPs and for coordinating responses that promote thermotolerance. Wang et al. (2023) [28] indicated that the SUMO protein (Small Ubiquitin-like Modifier), when covalently bound to lysine residues on the target protein TaHsfA1, acts as an ON/OFF switch, dynamically regulating the activation of heat stress response genes in wheat. This evidences a sophisticated post-translational control mechanism that adjusts the heat response, a process referred to as SUMOylation. Moreover, Huang et al. (2023) [29] demonstrated that HSFA1a modulates heat stress responses by altering the chromatin architecture, which optimizes interactions between enhancers and promoters of HSP genes, thereby promoting a rapid and robust response to heat.
Although these mechanisms can minimize damage, the persistence of heat stress often exceeds the plant’s defense capacity, resulting in cellular dysfunction [30,31].
These negative events of high temperature decrease the efficiency of photosynthesis [32] and productivity [2]. This decrease in photosynthesis is also attributed to the inactivation of Rubisco and photosystem II (PSII) [21] due to the degradation of proteins associated with this system, which are particularly sensitive to heat [33]. In addition, stomatal closure, a common response to heat, limits CO2 absorption and directly affects carbohydrate production [34]. Thermal acclimatization involves adjusting cell metabolism, modifying the composition of cell membranes for greater stability, and regulating defense genes [35] in C4 plants due to their anatomy and specialized photosynthetic pathway, which allows for more efficient CO2 fixation in hot environments [36]. In contrast, C3 plants, such as rice and wheat, have higher yield losses at high temperatures due to their lower photosynthetic efficiency (Fv/Fm) under heat stress [37].
The impact of heat stress on plant reproductive development is critical. During the flowering phase, high temperatures can cause flower abortion, reduced pollen viability, and fertilization failure [38]. This results in low seed formation and, consequently, a drop in productivity, especially in wheat, which is highly sensitive to heat during anthesis and grain filling [39].
The effect of increased temperature is directly related to plant nutrition. Ref. [40], using transcriptome analysis, indicated in a study with Panicum virgatum (switchgrass) that heat stress suppresses the expression of more than 3000 genes, including many involved in N metabolism, especially in the assimilation of this nutrient, causing damage to biomass accumulation.
Studies in rice cultivation have indicated that heating alone by 0.7 °C compared to room temperature reduces yield by redirecting N to vegetative parts to the detriment of grains [41].
Rising temperatures can reduce the activity of enzymes that are essential for the incorporation of nutrients into plants, such as nitrate reductase (NR) and glutamine synthetase (GS) [3,42], especially in areas where underdoses of nitrogen fertilizers are used. This problem is critical in regions with limited resources, where the high cost of fertilizers restricts their use, directly affecting plant nutrition and their productive capacity [3,43]. In conditions of nitrogen deficiency, plants invest in the production of secondary organic compounds, such as flavonoids and phenolic compounds, to strengthen their antioxidant defenses against oxidative stress. However, this occurs at the expense of energy that could be directed towards growth and production, damaging primary metabolism and productivity [44,45].
In contrast, plants with optimal doses of nitrogen can invest in both the synthesis of defense compounds and compounds necessary for growth, ensuring better resilience and productivity under heat stress [4,46,47]. In view of the damage to plant metabolism caused by rising temperatures, it is essential to explore strategies that optimize nitrogen nutrition as a way of mitigating the effects of heat stress in challenging agricultural environments. Recent advances have enabled the development of thermotolerant cultivars through genetic improvement, which, although time-consuming, provides a longer-lasting effect, and the use of biostimulants, a rapidly deployable technology with immediate effects on the plant as it promotes the synthesis of heat shock proteins, thereby strengthening plant defense mechanisms [27]. Moreover, sustainable management practices, such as mulching, which reduces soil temperature and conserves moisture, and the implementation of agrivoltaic systems, which offer shading and protection against thermal extremes, demonstrate potential to mitigate the effects of thermal stress, enhancing photosynthetic efficiency and nutrient uptake [48]. These strategies have contributed to increasing crop resilience and promoting more sustainable agriculture in the face of global warming.
3. Physiological and Molecular Mechanisms of the Interaction Between Nitrogen and Thermal Stress
For plants to benefit from N, fertilization is necessary because most of the world’s soils do not have the quantities of the nutrient to meet plant demand, as it is the element most required by plants [4]. N in the soil predominates in an organic form, which is why it is important to mineralize this nutrient in the soil to increase the inorganic forms that can be assimilated by plants, which is highly dependent on temperature. The ideal temperature accelerates the nitrogen mineralization process, increasing the availability of ammonium (NH4+) and nitrate (NO3−) in the soil. De Jesus et al. (2024) [49], when evaluating different organic fertilizers, observed that N mineralization was more efficient at temperatures between 20 °C and 30 °C, and above this range, the rate of mineralization and the synchrony between N release and plant demand can be modified (Figure 2).
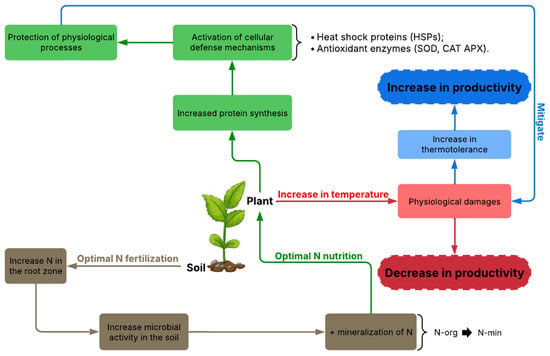
Figure 2.
Integrated scheme of nitrogen nutrition in mitigating thermal stress.
In addition, excessive heat above the upper threshold temperature of the species can interrupt cell division and cell elongation [22] and inhibit root development by compromising the absorption of water and nutrients [20]. In a study with tomato plants, Giri et al. (2017) [43] observed that increasing the temperature (35/30 °C to 42/37 °C day/night) significantly decreased the concentrations of nitrate transporter proteins (NRT1, NRT2) and ammonium transporter proteins (AMT1) in the roots, directly affecting the plant’s ability to absorb N.
In another study with maize, grown under field conditions [47], it was observed that heat stress with temperatures above 35 °C reduced the integrity of cell membranes, photosynthesis, and grain yield. The authors add that hybrids that exhibited less membrane damage maintained a higher yield under these adverse conditions, suggesting genetic variability in terms of heat resistance.
In another study with plants from the poaceae group, Mohammed and Tarpley (2009) [50] pointed out that high night temperatures (32 °C) in rice plants grown in a controlled environment cause a significant reduction in grain yield (90%) due to lower pollen germination and spikelet fertility, although leaf photosynthesis was not directly affected in this study. Furthermore, the nitrogen content in the leaves was also reduced, which contributed to a decrease in spikelet fertility and affected the distribution of N and the fertility of the rice, with direct implications for grain yield.
Another form of nutritional damage to the plant caused by high air temperatures is linked to damage to photosynthesis, which consequently decreases the plants’ ability to use nitrogen effectively [51]. In legumes such as peas, heat stress (grown initially at 21 °C and, after 21 days, at 28 °C) promoted apical senescence in the nodules, negatively affecting N fixation [52], although the authors did not assess the nutritional status of the plant.
Another study was carried out with two fast- and slow-growing poinsettias under different conditions of temperature and nitrogen availability [53]. The authors observed that the high temperature of 30 °C, compared to 20 °C, increased the respiration rates of roots and leaves with a lower fraction of available carbon in the fast-growing species Bromus hordeaceus, almost halving the relative growth rate of this plant. In contrast, the slow-growing species Bromus erectus showed better adaptation to heat, maintaining more stable growth rates under high temperatures due to an increase in nitrogen use efficiency (NUE) under low-nitrogen conditions. It was therefore noted that N limitation combined with rising temperatures affects fast-growing species more severely, although the authors did not assess the plants’ N nutritional status.
Research with maize varieties grown in experimental fields under heat stress evaluated the impact of an increase in air temperature of approximately 6.0 °C and 3.1 °C compared to a control in different study years (2020: 41.9 °C and 35.9 °C; 2021: 40.8 °C and 37.7 °C) on plant physiology [54]. The authors noted that a medium dose of N (180 kg/ha) mitigated the adverse effects of heat, and high doses (270 kg/ha) worsened photosynthetic performance and reduced yields, particularly in heat-sensitive varieties. In this study, it can be seen that the dose of N can help or hinder the effects of high temperatures, but the plant’s nutritional status for N was not observed, i.e., whether the doses studied induced precisely the right nutritional status for N or not.
4. Optimum N Concentration Reduces the Effect of High Temperature Stress
Around the world, climate change has induced heat waves that can cause damage to irrigated crops, but this isolated aspect needs to be discussed more specifically, and review studies are incipient, which raises concerns. The optimized use of N can be an important strategy to increase crop growth and production and strengthen mechanisms of tolerance to heat stress. This hypothesis is relevant for debate based on the research carried out and to indicate possible improvements to strengthen this thesis, given the relevance of this issue for world agriculture, which is being affected by climate change.
One study investigated the impact of heat stress (temperatures: maximum: 36 °C; minimum: 26 °C) on wheat plants grown in pots under optimum water levels and with different doses of N compared to ambient-temperature conditions (temperatures: maximum: 26 °C; minimum: 16 °C) [42]. The authors observed negative impacts of heat stress, with a decrease in photosynthetic rate on the 12th day of stress compared to room temperature. In N-deficient plants, the increase in temperature resulted in a decrease in N accumulation and grain protein yield. However, during this same period of stress, the application of medium doses of N (1.48 g N per pot) in relation to the low dose increased N absorption and the activity of the enzymes NR and GS and grain protein yield. Leghari et al. (2024) [55], in experiments conducted in wheat production systems in China and Pakistan, observed that the application of 207 kg N/ha in Tongzhou (China) maintained productivity and NUE while reducing N losses, suggesting it to be the optimal rate for the region. In contrast, in Faisalabad (Pakistan), the application of 125 kg N/ha resulted in increased productivity but was accompanied by greater N losses and a reduction in NUE.
Although N is one of the main macronutrients involved in plant growth and development, its efficient assimilation and role in mitigating thermal stress depend directly on its interaction with other nutrients, such as phosphorus (P) and potassium (K).
Phosphorus is essential for energy transfer in plant cells, serving as a key component of ATP (adenosine triphosphate), which provides the energy required for various metabolic reactions. This energetic function directly impacts N assimilation, as the nitrate absorbed by the roots must be reduced to ammonium to be incorporated into cellular metabolism. This process occurs in two stages catalyzed by the enzymes nitrate reductase (NR) and nitrite reductase (NiR), which require large amounts of ATP-derived energy [3,4]. P deficiencies reduce this conversion, limiting the availability of assimilable N. The ammonium produced is incorporated into the carbon skeleton via the GS and glutamate synthase (GOGAT), both of which depend on ATP. Under thermal stress conditions, the demand for HSPs increases, necessitating an efficient supply of both N and P to support this accelerated protein synthesis [4,46]. Additionally, P is involved in electron transport during photosynthesis, which is critical for maintaining the integrity of the photosynthetic system under high temperatures. Kayoumu et al. (2023) [56], while assessing the effects of low P availability on cotton grown hydroponically, observed a direct effect in the increased production of ROS and, consequently, a decrease in pigment content, net photosynthetic rate, PSII content, ATP, and NADP(H). They also noted an impairment in the electron transport from the donor side of PSII to the acceptor side of PSI, with a greater impact on the PSII donor-side performance than on PSII overall or the PSI acceptor side. P limitation under thermal stress can reduce water use efficiency, compromising cellular homeostasis and N assimilation.
Although not part of structural organic compounds like N and P, K is essential for the activation of more than 60 enzymes involved in plant metabolism. K regulates the activity of NR, NiR, and GS, which are fundamental for converting absorbed N into usable organic compounds [4]. Under high temperatures, the stability of these enzymes may be compromised, making adequate K supply essential to maintaining their functionality. Furthermore, K is the primary cation responsible for maintaining cellular turgor and controlling stomatal opening, playing a fundamental role in osmotic regulation and protection against cellular damage. During thermal stress, transpiration regulation is critical to prevent excessive dehydration and oxidative stress. K deficiency can increase plant vulnerability to heat, as it affects membrane stability and reduces antioxidant capacity [57]. Moreover, K modulates the transcription of genes involved in protein synthesis—especially those related to HSPs—and its role in activating SOD, CAT, and APX strengthens the antioxidant systems that protect cells from heat-induced damage [57]. Sarwar et al. (2019) [58], in a study with cotton, observed that K application enhanced chlorophyll content, net photosynthetic rate, water relations, and overall crop productivity.
In addition to the interactions discussed among N, P, and K, it is essential to explore the dynamic interactions between N and other nutrients such as calcium (Ca) and sulfur (S). During the absorption process, Ca plays a critical role in maintaining the integrity of cellular membranes, thereby facilitating the uptake of nutrients, including N, which is particularly relevant under high-temperature scenarios [59]. Yang et al. (2019) [60], while studying seedlings of different rice genotypes in a controlled environment, demonstrated that Ca2+ mediated signaling can modulate NO3− uptake and improve N assimilation even under thermal stress, favoring both root system expansion and the microbial activity responsible for N mineralization. In parallel, S plays a crucial metabolic role as a key component in the synthesis of sulfur-containing amino acids, which are essential for protein formation, thereby collaborating with N metabolism and the formation of HSPs that are fundamental for cellular protection against heat-induced damage [4]. Mir et al. (2024) [61] evidenced that increased S assimilation improves N metabolism, enhances antioxidant production, and contributes to maintaining redox balance under stress conditions. Thus, the integration of a balanced nutritional management strategy that considers the interactions between N and other nutrients—notably P, K, Ca, and S—can collectively potentiate the beneficial effects of N. Careful integrated nutrient management can mitigate the impacts of thermal stress, promoting greater resilience and productivity in plants.
In research with wheat grown in pots, Fan et al. (2022) [3] studied the relationship between N application and temperature in the reproductive phase of the plant. Initially, they observed that under low-N conditions and high temperatures during grain filling, there was a decrease in photosynthesis, translocation of photoassimilates, and plant yield. The authors concluded that the strategic application of nitrogen before heat stress helps prepare plants to face high temperatures, increasing sucrose content and the activity of the enzyme sucrose-phosphate synthase (SPS) in the leaves, promoting NUE and protecting yield by increasing the accumulation of sugars and proteins in the grains. This improvement indicated that optimal N helps the plant to better withstand heat stress, minimizing yield losses in the crop.
A study was carried out focusing on recovery from heat stress in tomato plants grown in pots under different doses of N [44]. The authors observed in plants under heat stress treatment (40/30 °C—morning/evening) compared to the control temperature (30/20 °C—morning/evening) that a lack of N use caused a decrease in chlorophyll fluorescence intensity and a decrease in Fv/Fm in tomato leaves. However, they observed that plants under moderate doses of N (1.95 g N per plant) grown under heat stress showed an increase in fluorescence intensity, while high doses of the nutrient (3.25 g N per plant) negatively affected the Fv/Fm of heat-stressed plants. However, the authors did not monitor the effect of treatments on the plants’ N accumulation.
In research under pot conditions, Hu et al. (2024) [46] studied the cultivation of rice cultivars subjected to doses of N at the panicle initiation stage and under conditions of high temperature (37/27 °C—morning/evening) and at a control temperature (31/27 °C—morning/evening) without additional stress. The authors observed that plants grown under a low dose of N (1.20 g N per pot) subjected to high temperatures experienced a reduction in spikelet fertility, abnormalities in anther development, and a reduction in pollen viability and, consequently, a reduction in grain yield. However, with the optimum application of nitrogen (2.52 g N per pot), this damage from the high temperature was mitigated in the plant, as it increased pollen viability, anther integrity, and spikelet fertility and, consequently, grain yield.
One study evaluated the response of Agrostis stolonifera (bentgrass) grown in pots and subjected heat stress (38/28 °C—morning/evening) and under different doses of N [62] with optimal irrigation and fertilization. The authors observed that in plants without the use of N and grown under high temperatures, there was a decrease in Fv/Fm and an increase in the rate of electrolyte leakage in the shoots (ShEL). However, at the optimum dose of N (7.5 kg N ha−1 applied biweekly until a total of 65 kg/ha), compared to the plant without N, heat stress was mitigated, as there was an increase in Fv/Fm and cell integrity. These results indicate that N in adequate doses acts to mitigate high temperatures by maintaining photosynthesis and reducing cell damage by regulating the production of HSPs, such as HSP101 and HSP70, which protect plants against the deleterious effects of heat. However, it was not possible to understand the level of stress caused by the increase in temperature due to the lack of a room-temperature treatment and the losses or gains induced by the treatments on N accumulation in the plant
In a study evaluating how different doses N (0.5, 2 and 8 mM) affect photosynthesis (PSI and PSII) in rice plants grown in hydroponics under ambient (27/22 °C—morning/evening) and high (35/30 °C—morning/evening) temperatures [63]. The authors observed that plants grown under a low N concentration (0.5 mM) and subjected to a high temperature compared to room temperature experienced a decrease in the efficiency of PSII use and the plant’s photosynthetic efficiency. However, plants under heat stress and grown in a nutrient solution with an optimum concentration of N (8 mM) experienced an increase in photosynthetic efficiency. In this way, the study suggests that increasing the supply of N helps to maintain the balance between light absorption and that dissipated as heat (NPQ), which is essential for protecting the photosynthetic system under conditions of heat stress, although the effects of the treatments on N absorption by the plants are not known.
In general, the underlying benefits of N in increasing tolerance to high temperatures observed may be related to its effects on the expression of genes related to thermotolerance and the maintenance of osmotic balance [64] and the consequent improvement in the antioxidant defense system in terms of the production of antioxidants [65], water use efficiency [51,65], and stabilizing cell function [6]. We have also found that supplying the plant with N at high temperatures induces strategies in the reproductive phase such as increasing the viability of pollen grains, ensuring fertilization and setting of the flower [46], as it synthesizes amino acids such as proline and GABA, minimizing the action of ROS and favoring pollen grain viability [10].
The interaction between nitrogen nutrition and high temperature above the species’ optimum level, even if there is no other associated stress, is complex and highly dependent on many factors relating to the plant and its adaptive mechanisms of action to the stress and the growing environment
This review showed that most of the studies evaluating the combination of N and air temperature were carried out in pot conditions, with only three studies on field crops [11,38,47] in crops with optimum irrigation and fertilization.
Pot experiments are valuable for obtaining data under highly controlled conditions, where factors such as irrigation, temperature, and fertilization can be rigorously adjusted. However, this approach has limitations compared to natural field conditions. In pots, the restricted soil volume limits root development, thereby reducing nutrient uptake and the interactions between roots and the soil’s rich microbiota that occur in field environments. These interactions are critical for N mineralization, as microbial activity converts soil nitrogen into plant-absorbable forms. Additionally, the microclimate in pot experiments tends to be highly homogeneous, whereas field-exposed plants face diurnal and seasonal variations in temperature, humidity, solar radiation, and interactions with diverse biotic factors. These variations directly influence nutrient dynamics and NUE. Guo et al. (2022) [66], while evaluating maize development under nitrogen doses in field conditions, demonstrated that root architecture and nitrogen uptake efficiency are significantly superior in environments that promote robust root system development, higher microbial diversity, and variable climatic conditions, all of which enhance nutrient absorption, including nitrogen. Thus, while pot experiments remain indispensable for elucidating physiological mechanisms under controlled conditions, extrapolating these results to agricultural practice requires caution. Field studies more accurately replicate real-world cultivation conditions by simultaneously interacting with multiple biotic and abiotic factors. These interactions directly affect plant nitrogen absorption and metabolism, ultimately shaping crop performance in the field.
Another concern is that most of the doses of N studied were not monitored based on the accumulation or assessment of the nutritional status for N, hindering further discussion to better understand the effects of temperature and/or doses of N on plant nutrition and, consequently, on physiology and the accumulation of dry mass and/or productivity. These difficulties for the authors are due to the lack of information on adequate leaf N levels for crops, especially in the field, which can be useful for monitoring the nutritional status of the crop.
In a scenario of global warming, it is imperative to invest in new basic and applied studies, especially at the field level, to develop agronomic practices that consider the synergy between N management and thermal fluctuations, promoting agriculture that is more resilient to extreme heat. However, there are situations where, depending on the species, a moderate increase in temperature of around 2 °C can favor plant nutrition and crop yields, especially in the group of forage plants grown in tropical regions.
Faced with the pressure of climate change, there has been an increase in irrigated and well-fertilized crops around the world, but this review shows that this will not be enough to reverse the damage, as we have seen in most species that the increase in temperature is likely to be a significant climate problem in this technified agro-ecosystem. However, it was discussed that the proper use of N can be an efficient strategy for boosting production whilst at the same time favoring the plant’s defense system against this thermal stress, but there is room for progress that needs to be considered from a future perspective (Table 1).

Table 1.
Optimal nitrogen concentration for different crop types worldwide. These values are initial references and should be adjusted based on soil analysis, management history, and the specific climate conditions of each region. It is worth noting that in leguminous crops (such as soybeans), biological nitrogen fixation reduces the need for high nitrogen fertilization.
5. Prospects
The use of more efficient N sources according to climatic conditions [49,64] minimizes the environmental impacts associated with NO3− leaching and greenhouse gas emissions [6] and should improve the efficiency of nitrogen fertilization. For example, slow-release fertilizers can reduce volatilization and leaching losses by up to 40% compared to conventional fertilizers such as urea and/or nitrification inhibitors [75]. The use of crop rotation, including green manures, contributes to improving the natural cycling of N [76]. This research should be encouraged in order to enhance sustainable crop production and resilience to heat stress.
In most of the world’s growing regions, there is a lack of technical recommendations for nitrogen fertilization based on research. This prevents optimum nitrogen nutrition in the plant, and as a result, the scientific robustness of studies evaluating N and temperature is hampered. The optimum doses of N depend on the expected productivity and the response class of the growing region, i.e., in areas with a high response class, the doses of N are higher compared to areas with a low response class of the plant to N. Systems with a high response class refer to areas with soils of optimum fertility but that are often cultivated with grasses and/or sandy-textured soils and/or irrigated areas. In addition, low-response-class systems refer to areas cultivated with legumes and/or areas left fallow for a long time or pasture areas with low soil fertility. This basic information for experimentation involving N is essential, but most studies do not present these indicators, as mentioned above. Therefore, more basic studies are needed to establish updated nitrogen fertilization programs from research to better exploit the benefits of N and favor the plant’s tolerance to heat waves.
Most studies involving N and temperature in irrigated systems use nitrogen fertilizer in a solid form rather than fertigation. This strategy allows for a more uniform distribution of nutrients and a reduction in N losses, since the nutrient is applied directly to the root zone of the plants and is versatile and can be used in the field or in greenhouses [77]. Therefore, more research is needed using N fertigation at different air temperatures.
The retention of chlorophyll and the longevity of the photosynthetic apparatus for plant adaptation under high temperature is a relevant process, and therefore, more research is needed to silence genes associated with chlorophyll degradation. For example, suppression of the NYC gene delays the senescence of crops that have catalyzed the breakdown of chlorophyll for the conversion of chlorophyll-b into chlorophyll-a [78,79]. Genetic engineering studies, which is the science that competes in the incorporation of an individual gene into a plant, should be encouraged, and this research would allow for faster genetic gain than conventional breeding methods. Gene editing techniques, such as CRISPR/Cas9 technology, offer the possibility of precisely modifying genes associated with N uptake and heat stress tolerance, fostering the development of more resilient and climate-adapted cultivars. According to Chen et al. (2019) [80], these approaches enable the targeted modification of molecular pathways, facilitating the improvement of critical agronomic traits without introducing undesired exogenous elements. Concurrently, the integration of high-resolution phenomics platforms with machine learning models has enabled the collection and analysis of vast phenotypic datasets, which can be correlated with environmental variables such as temperature and N availability. Gill et al. (2022) [81] further highlight that this combination allows for the creation of predictive models to optimize real-time fertilizer application, adjusting rates in response to dynamic environmental conditions and maximizing NUE. This technological integration represents a promising strategy for precision agriculture, enhancing both productivity and sustainability in agricultural production systems.
Optimal N metabolism is highly dependent on various nutrients, such as S, P, K, Mo, and Fe, so it is vital to ensure optimum nutrition from these elements to boost N’s contribution to increasing the thermotolerance of crops, so these interactions need to be better discussed.
6. Conclusions
This review highlights the importance of nitrogen nutrition for the resilience of plants subjected to heat stress, such as heat waves, which are becoming more frequent due to climate change. Nitrogen plays a central role in plant metabolism, as it can strengthen different defense systems by modulating physiological processes and antioxidant systems, reconciling greater plant resilience to temperature increases and greater productivity. However, there is still a need for advances in understanding the molecular mechanisms that govern the interaction between nitrogen nutrition and temperature, as well as further research into specific fertilization practices for different crops and climatic conditions. In the climate change scenario, a better understanding of the benefits of optimized N will help to better manage the stress caused by rising air temperatures towards more sustainable agriculture.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- IPCC. Climate Change 2021: The Physical Science Basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Bartlett, M.K.; Zhu, S.D.; Scoffoni, C.; Sack, L. The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: A global meta-analysis. Ecol Lett. 2016, 19, 667–675. [Google Scholar]
- Fan, Y.; Lv, Z.; Zhang, Y.; Ma, L.; Qin, B.; Liu, Q.; Zhang, W.; Ma, S.; Ma, C.; Huang, Z. Pre-anthesis night warming improves post-anthesis physiological activity and plant productivity to post-anthesis heat stress in winter wheat (Triticum aestivum L.). Environ Exp Bot. 2022, 197, 104819. [Google Scholar] [CrossRef]
- Prado, R.M. Nitrogen. In Mineral Nutrition of Tropical Plants, 1st ed.; Prado, R.M., Ed.; Springer: Cham, Switzerland, 2021; pp. 69–98. [Google Scholar]
- Maathuis, F.J.M. Physiological functions of mineral macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Spaccarotella, K.; Gido, J.; Samanta, I.; Chowdhary, G. Effects of Heat Stress on Plant-Nutrient Relations: An Update on Nutrient Uptake, Transport, and Assimilation. Int. J. Mol. Sci. 2023, 24, 15670. [Google Scholar] [CrossRef] [PubMed]
- Streck, N.A.; Weiss, A.; Xue, Q.; Baenziger, P.S. Improving predictions of developmental stages in winter wheat: A modified Wang and Engel model. Agric. For. Meteorol. 2003, 115, 139–150. [Google Scholar] [CrossRef]
- Ohama, N.; Sato, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional Regulatory Network of Plant Heat Stress Response. Trends Plant Sci. 2017, 22, 53–65. [Google Scholar] [CrossRef]
- Mittler, R.; Finka, A.; Goloubinoff, P. How do plants feel the heat? Trends Biochem. Sci. 2012, 37, 118–125. [Google Scholar] [CrossRef]
- Santiago, J.P.; Sharkey, T.D. Pollen development at high temperature and role of carbon and nitrogen metabolites. Plant Cell Environ. 2019, 42, 2759–2775. [Google Scholar] [CrossRef]
- Gonzalez-Meler, M.A.; Silva, L.B.C.; Dias-De-Oliveira, E.; Flower, C.E.; Martinez, C.A. Experimental air warming of a Stylosanthes capitata Vogel dominated tropical pasture affects soil respiration and nitrogen dynamics. Front. Plant Sci. 2017, 8, 46. [Google Scholar] [CrossRef]
- World Meteorological Organization. State of the Global Climate 2023; WMO: Geneva, Switzerland, 2024; Available online: https://wmo.int/publication-series/state-of-global-climate-2023 (accessed on 14 November 2024).
- World Bank. Climate-Resilient Irrigation; World Bank: Washington, DC, USA, 2024; Available online: https://www.worldbank.org/en/topic/climate-resilient-irrigation (accessed on 14 November 2024).
- Kufa, C.A.; Bekele, A.; Atickem, A. Impacts of climate change on predicted habitat suitability and distribution of Djaffa Mountains Guereza (Colobus guereza gallarum) using MaxEnt algorithm in Eastern Ethiopian Highland. Glob. Ecol. Conserv. 2022, 35, e02094. [Google Scholar] [CrossRef]
- Illangasingha, S.; Koike, T.; Rasmy, M.; Tamakawa, K.; Matsuki, H.; Selvarajah, H. A holistic approach for using global climate model (GCM) outputs in decision making. J. Hydrol. 2023, 626, 130213. [Google Scholar] [CrossRef]
- Barreto, R.F.; Prado, R.d.M.; Habermann, E.; Viciedo, D.O.; Martinez, C.A. Warming Changes Nutritional Status and Improves Stylosanthes capitata Vogel Growth Only Under Well-Watered Conditions. J. Soil Sci. Plant Nutr. 2020, 20, 1838–1847. [Google Scholar] [CrossRef]
- Viciedo, D.O.; Prado, R.d.M.; Martinez, C.A.; Habermann, E.; Piccolo, M.d.C.; Hurtado, A.C.; Barreto, R.F.; Calzada, K.P. Changes in soil water availability and air-temperature impact biomass allocation and C:N:P stoichiometry in different organs of Stylosanthes capitata Vogel. J. Environ. Manag. 2021, 278 Pt 1, 111540. [Google Scholar] [CrossRef]
- Carvalho, J.M.; Barreto, R.F.; Prado, R.M.; Habermann, E.; Martinez, C.A.; Branco, R.B.F. Elevated [CO₂] and warming increase the macronutrient use efficiency and biomass of Stylosanthes capitata Vogel under field conditions. J. Agro. Crop. Sci. 2020, 206, 597–606. [Google Scholar] [CrossRef]
- Olivera-Viciedo, D.; Prado, R.d.M.; Martinez, C.A.; Habermann, E.; Piccolo, M.d.C.; Calero-Hurtado, A.; Barreto, R.F.; Peña, K. Are the interaction effects of warming and drought on nutritional status and biomass production in a tropical forage legume greater than their individual effects? Planta 2021, 254, 104. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Artmed: Porto Alegre, Brazil, 2017. [Google Scholar]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front Plant Sci. 2013, 4, 273. [Google Scholar]
- Riaz, A.; Thomas, J.; Ali, H.H.; Zaheer, M.S.; Ahmad, N.; Pereira, A. High night temperature stress on rice (Oryza sativa)—Insights from phenomics to physiology. A review. Funct Plant Biol. 2024, 51, FP24057. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Chaudhuri, B.; Roychoudhury, A. Influence of night temperature on rice yield and quality. In Rice Research for Quality Improvement: Genomics and Genetic Engineering; Roychoudhury, A., Ed.; Springer Nature: Singapore, 2020; pp. 579–590. [Google Scholar]
- Gupta, A.; Sarkar, A.K.; Senthil-Kumar, M. Global transcriptional regulation of heat shock proteins in Arabidopsis thaliana under heat stress. Front. Plant Sci. 2023, 14, 1123. [Google Scholar]
- Kotak, S.; Larkindale, J.; Lee, U.; Koskull-Döring, P.V.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar]
- Tian, F.; Hu, X.-L.; Yao, T.; Yang, X.; Chen, J.-G.; Lu, M.-Z.; Zhang, J. Recent advances in the roles of HSFs and HSPs in heat stress response in woody plants. Front. Plant Sci. 2021, 12, 704905. [Google Scholar] [CrossRef]
- Wang, H.; Feng, M.; Jiang, Y.; Du, D.; Dong, C.; Zhang, Z.; Wang, W.; Liu, J.; Liu, X.; Li, S. Thermosensitive SUMOylation of TaHsfA1 Defines a Dynamic ON/OFF Molecular Switch for the Heat Stress Response in Wheat. Plant Cell 2023, 35, 3889–3910. [Google Scholar] [CrossRef]
- Huang, Y.; An, J.; Sircar, S.; Bergis, C.; Lopes, C.D.; He, X.; Da Costa, B.; Tan, F.-Q.; Bazin, J.; Antunez-Sanchez, J.; et al. HSFA1a Modulates Plant Heat Stress Responses and Alters the 3D Chromatin Organization of Enhancer–Promoter Interactions. Nat. Commun. 2023, 14, 469. [Google Scholar] [CrossRef]
- Wang, W.; Hu, B.; Li, A.; Chu, C. NRT1.1s in plants: Functions beyond nitrate transport. J. Exp. Bot. 2020, 71, 4373–4379. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.F.; Chen, Y.; Sun, M.M.; Wang, Y.; Chen, Y.F. The transcription factor NIGT1.2 modulates both phosphate uptake and nitrate influx during phosphate starvation in Arabidopsis and maize. Plant Cell. 2020, 32, 3519–3534. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Sharkey, T.D. Effects of moderate heat stress on photosynthesis: Importance of thylakoid reactions, rubisco deactivation, reactive oxygen species, and thermotolerance provided by isoprene. Plant Cell Environ. 2005, 28, 269–277. [Google Scholar]
- Ma, X.; Bai, L. Elevated CO₂ and reactive oxygen species in stomatal closure. Plants 2021, 10, 410. [Google Scholar] [CrossRef]
- Mathivanan, S. Abiotic Stress-Induced Molecular and Physiological Changes and Adaptive Mechanisms in Plants. Abiotic Stress in Plants; IntechOpen: London, UK, 2021; Available online: https://www.intechopen.com/chapters/73040 (accessed on 17 March 2025).
- Sage, R.F.; Zhu, X.G. Exploiting the engine of C4 photosynthesis. J. Exp. Bot. 2011, 62, 2989–3000. [Google Scholar] [CrossRef]
- Yamori, W.; Hikosaka, K.; Way, D.A. Temperature response of photosynthesis in C3, C4, and CAM plants: Temperature acclimation and temperature adaptation. Photosynth Res. 2014, 119, 101–117. [Google Scholar] [CrossRef]
- Zinn, K.E.; Tunc-Ozdemir, M.; Harper, J.F. Temperature stress and plant sexual reproduction: Uncovering the weakest links. J. Exp. Bot. 2010, 61, 1959–1968. [Google Scholar]
- Farooq, M.; Wahid, A.; Lee, D.J.; Ito, O.; Siddique, K.H. Advances in drought resistance of rice. Crit. Rev. Plant Sci. 2009, 28, 199–217. [Google Scholar]
- Li, Y.F.; Wang, Y.; Tang, Y.; Kakani, V.G.; Mahalingam, R. Transcriptome analysis of heat stress response in switchgrass (Panicum virgatum L.). BMC Plant Biol. 2013, 13, 153. [Google Scholar] [CrossRef]
- Li, C.; Zhu, J.G.; Sha, L.N.; Zhang, J.S.; Zeng, Q.; Liu, G. Rice (Oryza sativa L.) growth and nitrogen distribution under elevated CO₂ concentration and air temperature. Ecol. Res. 2017, 32, 405–411. [Google Scholar] [CrossRef]
- Ru, C.; Hu, X.; Chen, D.; Song, T.; Wang, W.; Lv, M.; Hansen, N.C. Nitrogen modulates the effects of short-term heat, drought and combined stresses after anthesis on photosynthesis, nitrogen metabolism, yield, and water and nitrogen use efficiency of wheat. Water 2022, 14, 1407. [Google Scholar] [CrossRef]
- Giri, A.; Heckathorn, S.; Mishra, S.; Krause, C. Heat stress decreases levels of nutrient-uptake and assimilation proteins in tomato roots. Plants 2017, 6, 6. [Google Scholar] [CrossRef]
- Li, C.; Yang, Z.; Zhang, C.; Luo, J.; Jiang, N.; Zhang, F.; Zhu, W. Heat stress recovery of chlorophyll fluorescence in tomato (Lycopersicon esculentum Mill.) leaves through nitrogen levels. Agronomy 2023, 13, 2858. [Google Scholar] [CrossRef]
- Gerard, V.A. The role of nitrogen nutrition in high-temperature tolerance of the kelp, Laminaria saccharina (Chromophyta). J. Phycol. 2008, 33, 800–810. [Google Scholar]
- Hu, Q.; Yan, N.; Cui, K.; Li, G.; Wang, W.; Huang, J.; Peng, S. Increased panicle nitrogen application improves rice yield by alleviating high-temperature damage during panicle initiation to anther development. Physiol. Plant. 2024, 176, e14230. [Google Scholar] [CrossRef]
- Tas, T. Physiological and biochemical responses of hybrid maize (Zea mays L.) varieties grown under heat stress conditions. PeerJ 2022, 10, e14141. [Google Scholar] [CrossRef]
- Rossi, G.; Beni, C.; Neri, U. Organic Mulching: A Sustainable Technique to Improve Soil Quality. Sustainability 2024, 16, 10261. [Google Scholar] [CrossRef]
- de Jesus, H.I.; Cassity-Duffey, K.; Dutta, B.; da Silva, A.L.B.R.; Coolong, T. Influence of soil type and temperature on nitrogen mineralization from organic fertilizers. Nitrogen. 2024, 5, 47–61. [Google Scholar]
- Mohammed, A.R.; Tarpley, L. High Nighttime Temperatures Affect Rice Productivity through Altered Pollen Germination and Spikelet Fertility. Agric. For. Meteorol. 2009, 149, 999–1008. [Google Scholar] [CrossRef]
- Valenzuela, H. Optimizing the nitrogen use efficiency in vegetable crops. Nitrogen 2024, 5, 106–143. [Google Scholar] [CrossRef]
- Kitaeva, A.B.; Serova, T.A.; Kusakin, P.G.; Tsyganov, V.E. Effects of Elevated Temperature on Pisum sativum Nodule Development: II-Phytohormonal Responses. Int. J. Mol. Sci. 2023, 24, 17062. [Google Scholar] [CrossRef]
- Colesie, C.; Green, A.T.G.; Budke, J.M.; Lakatos, M.; Raggio, J.; Pintado, A. Differences in Growth-Economics of Fast vs. Slow Growing Grass Species in Response to Temperature and Nitrogen Limitation. BMC Ecol. 2020, 20, 11. [Google Scholar]
- Ma, X.; Cai, F.; Mu, X.; Li, H.; Shao, R.; Li, S.; Xu, J.; Wang, S.; Lu, L.; Zhao, X.; et al. Effects of nitrogen application rate on photosynthetic physiology of maize leaves and yield under high temperature stress at ear stage. J. Plant Nutr. Fertilizers. 2022, 28, 1852–1866. [Google Scholar] [CrossRef]
- Leghari, H.J.; Han, W.; Soomro, A.A.; Shoukat, M.R.; Zain, M.; Wei, Y.; Xu, Q.; Buriro, M.; Bhutto, T.A.; Soothar, R.K.; et al. Navigating water and nitrogen practices for sustainable wheat production by model-based optimization management systems: A case study of China and Pakistan. Agric. Water Manag. 2024, 300, 108917. [Google Scholar] [CrossRef]
- Kayoumu, M.; Iqbal, A.; Muhammad, N.; Li, X.; Li, L.; Wang, X.; Gui, H.; Qi, Q.; Ruan, S.; Guo, R.; et al. Phosphorus Availability Affects the Photosynthesis and Antioxidant System of Contrasting Low-P-Tolerant Cotton Genotypes. Antioxidants 2023, 12, 466. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Hossain, M.S.; Mahmud, J.A.; Hossen, M.S.; Masud, A.A.C.; Moumita Fujita, M. Potassium: A Vital Regulator of Plant Responses and Tolerance to Abiotic Stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef]
- Sarwar, M.; Saleem, M.F.; Ullah, N.; Ali, S.; Rizwan, M.; Shahid, M.R.; Alyemeni, M.N.; Alamri, S.A.; Ahmad, P. Role of mineral nutrition in alleviation of heat stress in cotton plants grown in glasshouse and field conditions. Sci. Rep. 2019, 9, 13022. [Google Scholar] [CrossRef]
- Jia, Y.; Qin, D.; Zheng, Y.; Wang, Y. Finding Balance in Adversity: Nitrate Signaling as the Key to Plant Growth, Resilience, and Stress Response. Int. J. Mol. Sci. 2023, 24, 14406. [Google Scholar] [CrossRef]
- Yang, J.; Deng, X.; Wang, X.; Wang, J.; Du, S.; Li, Y. The calcium sensor OsCBL1 modulates nitrate signaling to regulate seedling growth in rice. PLoS ONE 2019, 14, e0224962. [Google Scholar] [CrossRef]
- Mir, B.A.; Kumari, R.; Rakhra, G.; Parihar, P.; Singh, R.; Raju, A.D.; Srivastava, P.K.; Prasad, S.M.; Singh, R.; Gulliya, S. Sulfur assimilation and regulation of abiotic stress via OMICS. Plant Stress. 2024, 14, 100630. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, X.; Goatley, M.; Ervin, E. Heat Shock Proteins in Relation to Heat Stress Tolerance of Creeping Bentgrass at Different N Levels. PLoS ONE 2014, 9, e102914. [Google Scholar] [CrossRef]
- Takahashi, Y.; Wada, S.; Noguchi, K.; Miyake, C.; Makino, A.; Suzuki, Y. Photochemistry of Photosystems II and I in Rice Plants Grown under Different N Levels at Normal and High Temperature. Plant Cell Physiol. 2021, 62, 1121–1130. [Google Scholar] [CrossRef]
- Hu, J.; Ma, W.; Wang, Z. Effects of nitrogen addition and drought on the relationship between nitrogen- and water-use efficiency in a temperate grassland. Ecol. Process. 2023, 12, 36. [Google Scholar] [CrossRef]
- Zheng, X.; Zhu, Q.; Liu, Y.; Chen, J.; Wang, L.; Xiu, Y.; Zheng, H.; Lin, S.; Ling, P.; Tang, M. Combined analysis of transcriptome and metabolome provides insights into response mechanism under heat stress in avocado (Persea americana Mill.). Int. J. Mol. Sci. 2024, 25, 10312. [Google Scholar] [CrossRef]
- Guo, S.; Liu, Z.; Zhou, Z.; Lu, T.; Chen, S.; He, M.; Zeng, X.; Chen, K.; Yu, H.; Shangguan, Y.; et al. Root System Architecture Differences of Maize Cultivars Affect Yield and Nitrogen Accumulation in Southwest China. Agriculture 2022, 12, 209. [Google Scholar] [CrossRef]
- Wysokinski, A.; Wysokinska, A.; Noulas, C.; Wysokinska, A. Optimal nitrogen fertilizer rates for soybean cultivation. Agronomy 2024, 14, 1375. [Google Scholar] [CrossRef]
- Malik, T.H.; Lal, S.B.; Wani, N.R.; Amin, D.; Wani, R.A. Effect Of Different Levels Of Nitrogen On Growth And Yield Attributes Of Different Varieties Of Basmati Rice (Oryza sativa L.). Int. J. Sci. Technol. Res. 2014, 3, 444–448. [Google Scholar]
- Wu, B.; Zuo, W.; Yang, P.; Zhang, W. Optimal water and nitrogen management increases cotton yield through improving leaf number and canopy light environment. Field Crops Res. 2023, 290, 108745. [Google Scholar] [CrossRef]
- Desalegn, B.; Kebede, E.; Legesse, H.; Fite, T. Sugarcane productivity and sugar yield improvement: Selecting variety, nitrogen fertilizer rate, and bioregulator as a first-line treatment. Heliyon 2023, 9, e15520. [Google Scholar] [CrossRef]
- Luo, J.; Yang, Z.; Zhang, F.; Li, C. Effect of nitrogen application on enhancing high-temperature stress tolerance of tomato plants during the flowering and fruiting stage. Front. Plant Sci. 2023, 14, 1172078. [Google Scholar] [CrossRef]
- Silva, V.J.; Pedreira, C.G.; Sollenberger, L.E.; da Silva, L.S.; Yasuoka, J.I.; Almeida, I.C. Canopy Height and Nitrogen Affect Herbage Accumulation, Nutritive Value, and Grazing Efficiency of ‘Mulato II’ Brachiariagrass. Crop Sci. 2016, 56, 2054–2061. [Google Scholar]
- Santos, M.E.R.; Carvalho, A.N.; Paiva, A.J.; Carvalho, B.H.R.; Borges, G.S.; da Silva, S.P.; Oliveira, D.M.; de Oliveira, D.H.A.M. Contribution of tiller age category to herbage accumulation of Marandu palisadegrass under two fertilization regimes. Semin. Ciências Agrárias 2022, 43, 211–228. [Google Scholar]
- Ongaratto, F.; Fernandes, M.H.M.D.R.; Dallantonia, E.E.; Lima, L.D.O.; Val, G.A.D.; Cardoso, A.D.S.; Rigobello, I.L.; Campos, J.A.A.; Reis, R.A.; Ruggieri, A.C.; et al. Intensive Production and Management of Marandu Palisadegrass (Urochloa brizantha ‘Marandu’) Accelerates Leaf Turnover but Does Not Change Herbage Mass. Agronomy 2021, 11, 1846. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Y.; Zhong, S.; Meng, O.; Li, Y.; Wang, J.; Gao, Y.; Cui, X. Advances in controlled-release fertilizer encapsulated by organic-inorganic composite membranes. Particuology. 2024, 84, 236–248. [Google Scholar] [CrossRef]
- Smil, V. Harvesting the Biosphere: What We Have Taken from Nature; MIT Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Govindasamy, P.; Muthusamy, S.K.; Bagavathiannan, M.; Mowrer, J.; Jagannadham, P.T.K.; Maity, A.; Halli, H.M.; Sujayananad, G.K.; Vadivel, R.; Das, T.K.; et al. Nitrogen use efficiency—A key to enhance crop productivity under a changing climate. Front. Plant Sci. 2023, 14, 1121073. [Google Scholar] [CrossRef]
- Horie, Y.; Ito, H.; Kusaba, M.; Tanaka, R.; Tanaka, A. Participation of Chlorophyll b Reductase in the Initial Step of the Degradation of Light-Harvesting Chlorophyll a/b-Protein Complexes in Arabidopsis. J. Biol. Chem. 2009, 284, 17449–17456. [Google Scholar]
- Sarker, U.K.; Uddin, R.; Kaysar, S.; Hossain, A.; Somaddar, U.; Saha, G. Exploring relationship among nitrogen fertilizer, yield and nitrogen use efficiency in modern wheat varieties under subtropical condition. Saudi J. Biol. Sci. 2023, 30, 103602. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu. Rev. Plant Biol. 2019, 70, 667–697. [Google Scholar] [CrossRef] [PubMed]
- Gill, T.; Gill, S.K.; Saini, D.K.; Chopra, Y.; de Koff, J.P.; Sandhu, K.S. A Comprehensive Review of High Throughput Phenotyping and Machine Learning for Plant Stress Phenotyping. Phenomics 2022, 2, 156–183. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).