

The Optimum Substrate Moisture Level to Enhance the Growth and Quality of Arugula (Eruca sativa)




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growing Conditions
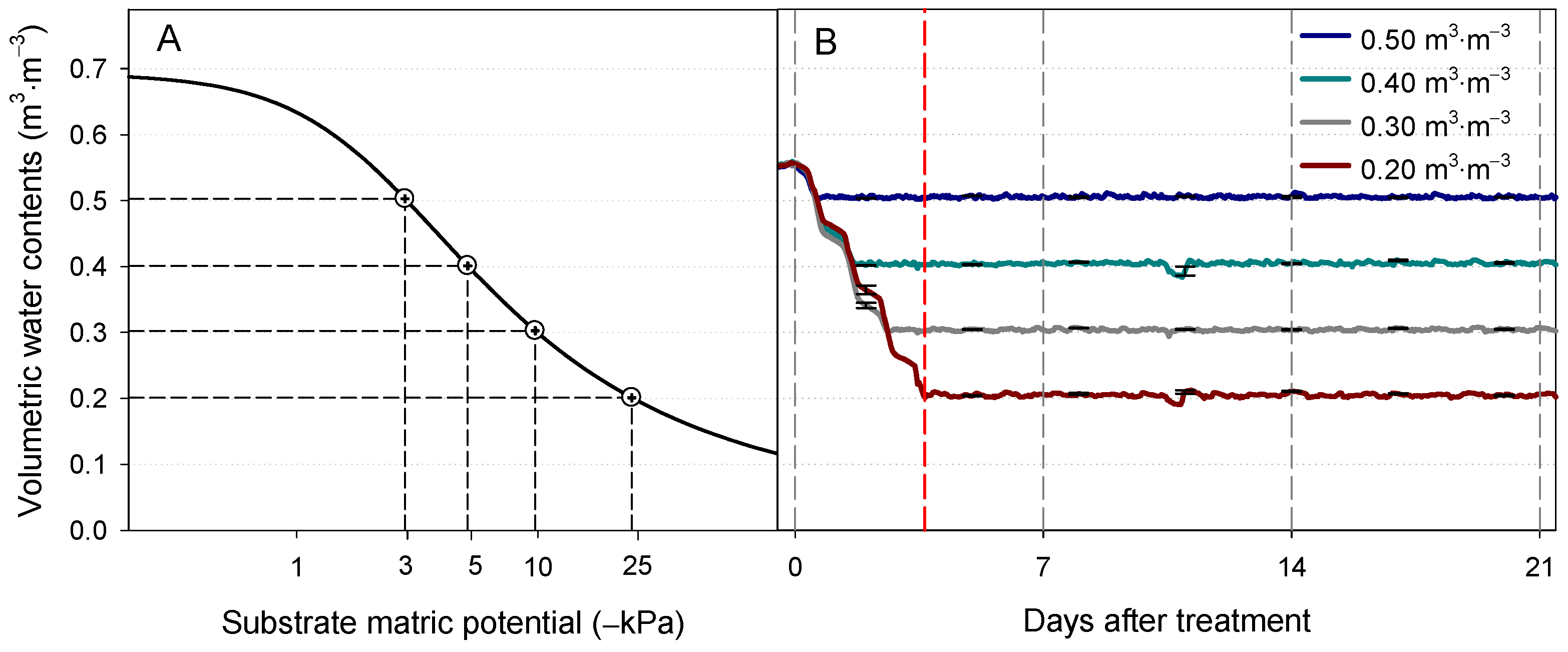
2.2. Irrigation Treatment via FDR Sensor-Based Automated Irrigation System
2.3. Growth and Physiological Measurements
2.4. Nitrate Content Measurement
2.5. Glucosinolate Content Analysis
2.6. Experimental Design and Statistical Analysis
3. Results
3.1. VWC Changes with the Automated Irrigation System
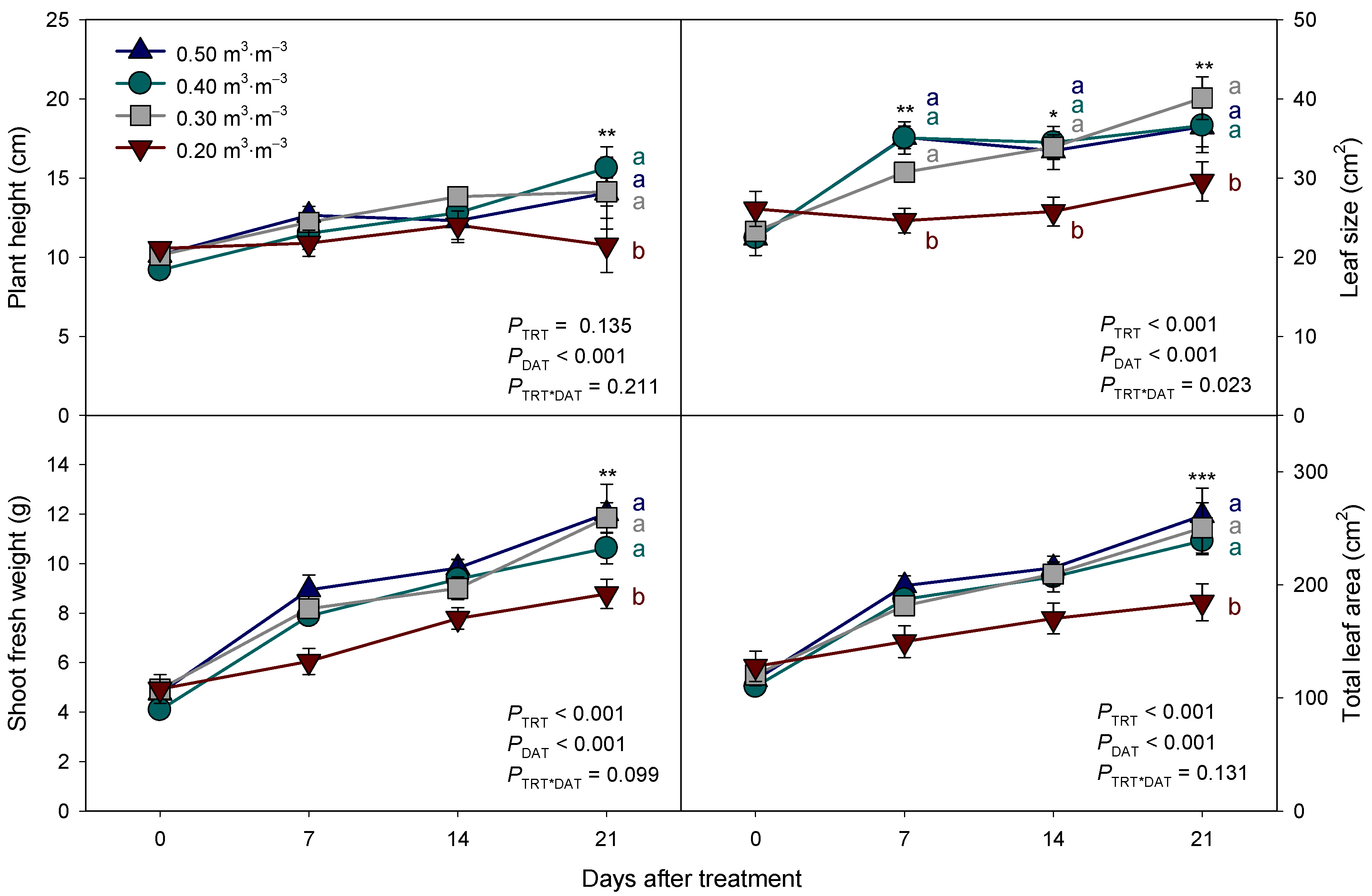
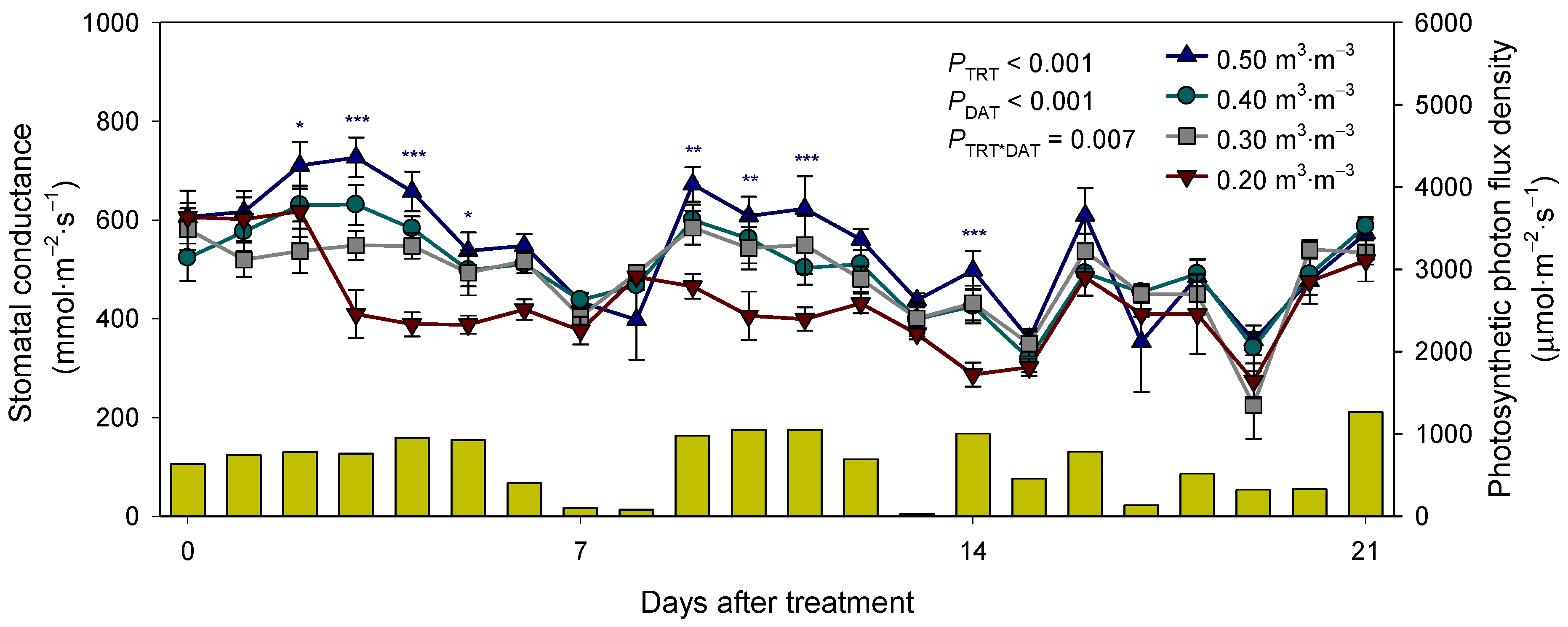
3.2. Growth and Leaf Physiological Parameters
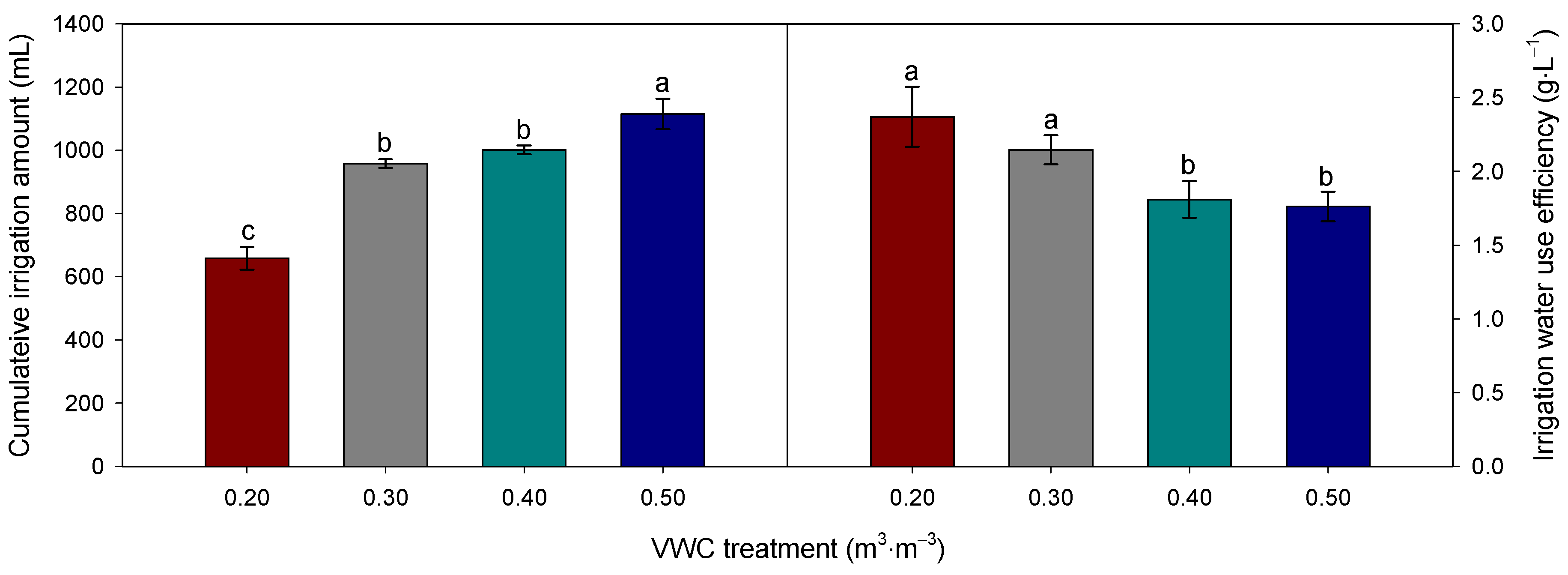
3.3. Water Use and Water Use Efficiency
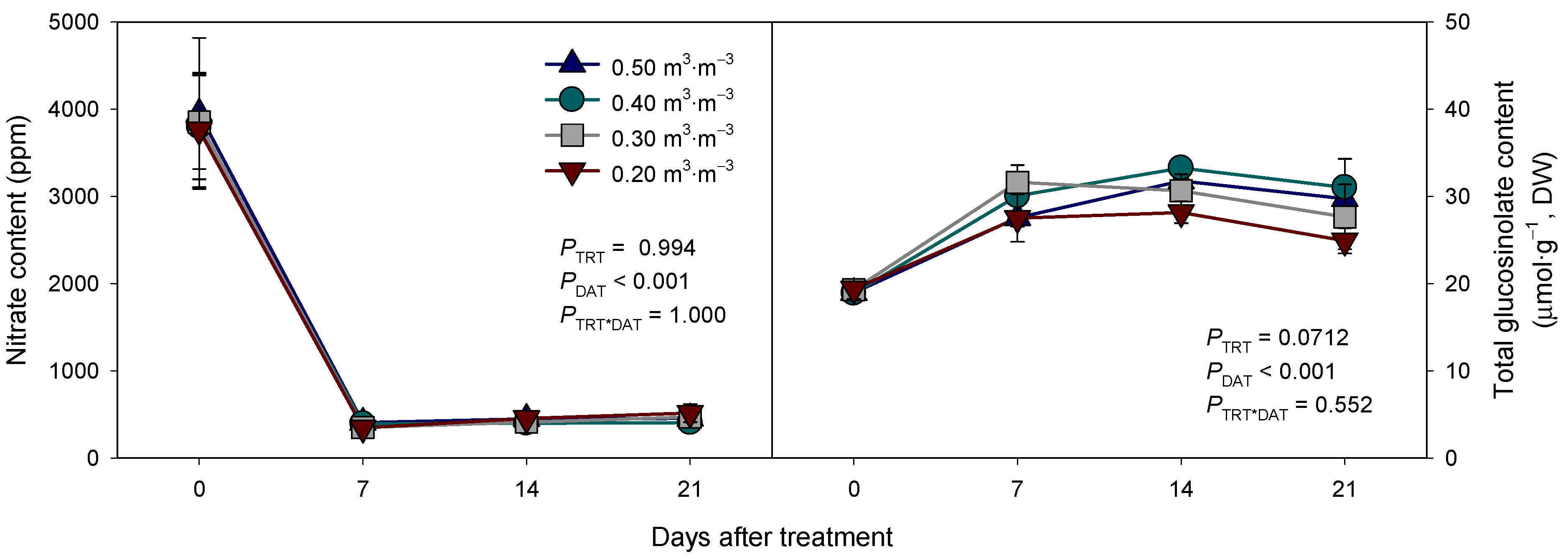
3.4. Changes in the Nitrate and Glucosinolate Content in Arugula by VWC Treatments
4. Discussion
4.1. Growth and Physiological Responses to VWC Treatment
4.2. VWC Effects on Arugula Quality
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Barazani, O.; Ziffer-Berger, J. Eruca sativa, a Tasty Salad Herb with Health-Promoting Properties. In Medicinal and Aromatic Plants of the Middle-East. Medicinal and Aromatic Plants of the World; Yaniv, Z., Dudai, N., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 2, pp. 269–279. [Google Scholar] [CrossRef]
- Matev, G.; Dimitrova, P.; Petkova, N.; Ivanov, I.; Mihaylova, D. Antioxidant activity and mineral content of rocket (Eruca sativa) plant from Italian and Bulgarian. J. Microbiol. Biotechnol. Food Sci. 2018, 8, 759. [Google Scholar] [CrossRef]
- Garg, G.; Sharma, V. Eruca sativa (L.): Botanical description, crop improvement, and medicinal properties. J. Herbs Spices Med. Plants 2014, 20, 171–182. [Google Scholar] [CrossRef]
- Vo, Q.V.; Trenerry, C.; Rochfort, S.; Wadeson, J.; Leyton, C.; Hughes, A.B. Synthesis and anti-inflammatory activity of indole glucosinolates. Bioorg. Med. Chem. 2014, 22, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Ku, K.M.; Kim, M.J.; Jeffery, E.H.; Kang, Y.H.; Juvik, J.A. Profiles of glucosinolates, their hydrolysis products, and quinone reductase inducing activity from 39 arugula (Eruca sativa Mill.) accessions. J. Agric. Food Chem. 2016, 64, 6524–6532. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Talalay, P. Antioxidant functions of sulforaphane: A potent inducer of phase II detoxication enzymes. Food Chem. Toxicol. 1999, 37, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Agerbirk, N.; Olsen, C.E.; Nielsen, J.K. Seasonal variation in leaf glucosinolates and insect resistance in two types of Barbarea vulgaris ssp. arcuata. Phytochemistry 2001, 58, 91–100. [Google Scholar] [CrossRef]
- Engelen-Eigles, G.; Holden, G.; Cohen, J.D.; Gardner, G. The effect of temperature, photoperiod, and light quality on gluconasturtiin concentration in watercress (Nasturtium officinale R. Br.). J. Agric. Food Chem. 2006, 54, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Shawon, R.A.; Kang, B.S.; Lee, S.G.; Kim, S.K.; Ju Lee, H.J.; Katrich, E.; Gorinstein, S.; Ku, Y.G. Influence of drought stress on bioactive compounds, antioxidant enzymes and glucosinolate contents of Chinese cabbage (Brassica rapa). Food Chem. 2020, 308, 125657. [Google Scholar] [CrossRef] [PubMed]
- Sarabi, B.; Ghaderi, N.; Ghashghaie, J. Light-emitting diode combined with humic acid improve the nutritional quality and enzyme activities of nitrate assimilation in rocket (Eruca sativa (Mill.) Thell.). Plant Physiol. Biochem. 2022, 187, 11–24. [Google Scholar] [CrossRef]
- European Union (EU) Commission Regulation. 2023/915 of 25 April 2023 on maximum levels for certain contaminants in food and repealing Regulation (EC). Off. J. Eur. Union 2023, L119, 103–157. [Google Scholar]
- Jin, J.; Koroleva, O.A.; Gibson, T.; Swanston, J.; Magan, J.; Zhang, Y.; Rowland, I.R.; Wagstaff, C. Analysis of phytochemical composition and chemoprotective capacity of rocket (Eruca sativa and Diplotaxis tenuifolia) leafy salad following cultivation in different environments. J. Agric. Food Chem. 2009, 57, 5227–5234. [Google Scholar] [CrossRef]
- Jasper, J.; Wagstaff, C.; Bell, L. Growth temperature influences postharvest glucosinolate concentrations and hydrolysis product formation in first and second cuts of rocket salad. Postharvest Biol. Technol. 2020, 163, 111157. [Google Scholar] [CrossRef] [PubMed]
- Signore, A.; Bell, L.; Santamaria, P.; Wagstaff, C.; Van Labeke, M.C. Red light is effective in reducing nitrate concentration in rocket by increasing nitrate reductase activity, and contributes to increased total glucosinolates content. Front. Plant Sci. 2020, 11, 604. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, E.M.; Giovanelli, L.B.; Delazari, F.T.; dos Santos, M.L.; Pereira, S.B.; da Silva, D.J. Arugula production as a function of irrigation depths and potassium fertilization. Rev. Bras. Eng. Agric. Ambient. 2017, 21, 197–202. [Google Scholar] [CrossRef]
- dos Santos, S.K.; da Silva Gomes, D.; dos Santos, L.W.O.; de Azevedo Soares, V.; Dantas, E.F.O.; Henschel, J.M.; Batista, D.S. Exogenous carnitine mitigates the deleterious effects of mild-water stress on arugula by modulating morphophysiological responses. J. Plant Growth Regul. 2023, 42, 4073–4082. [Google Scholar] [CrossRef]
- Jones, H.G. Monitoring plant and soil water status: Established and novel methods revisited and their relevance to studies of drought tolerance. J. Exp. Bot. 2007, 58, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Malladi, A.; van Iersel, M.W. Physiological and molecular responses to drought in petunia: The importance of stress severity. J. Exp. Bot. 2012, 63, 6335–6345. [Google Scholar] [CrossRef] [PubMed]
- An, S.K.; Lee, H.B.; Kim, J.; Kim, K.S. Efficient water management for cymbidium grown in coir dust using a soil moisture sensor-based automated irrigation system. Agronomy 2020, 11, 41. [Google Scholar] [CrossRef]
- Nam, S.; Kang, S.; Kim, J. Maintaining a constant soil moisture level can enhance the growth and phenolic content of sweet basil better than fluctuating irrigation. Agric. Water Manag. 2020, 238, 106203. [Google Scholar] [CrossRef]
- Nemali, K.S.; Montesano, F.; Dove, S.K.; van Iersel, M.W. Calibration and performance of moisture sensors in soilless substrates: ECH2O and Theta probes. Sci. Hortic. 2007, 112, 227–234. [Google Scholar] [CrossRef]
- de Boodt, M.; Verdonck, O. The physical properties of the substrates in horticulture. Acta Hortic. 1972, 26, 37–44. [Google Scholar] [CrossRef]
- Raviv, M.; Leith, J.H.; Bar-Tal, A. Soilless Culture: Theory and Practice; Elsevier: Boston, MA, USA, 2019; pp. 79–84. [Google Scholar]
- Chae, S.H.; Lee, O.N.; Park, H.Y.; Ku, K.M. Seasonal effects of glucosinolate and sugar content determine the pungency of small-type (Altari) radishes (Raphanus sativus L.). Plants 2022, 11, 312. [Google Scholar] [CrossRef]
- Al-Yasi, H.; Attia, H.; Alamer, K.; Hassan, F.; Ali, E.; Elshazly, S.; Siddique, K.H.M.; Hessini, K. Impact of drought on growth, photosynthesis, osmotic adjustment, and cell wall elasticity in damask rose. Plant Physiol. Biochem. 2020, 150, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Cornic, G. Drought stress inhibits photosynthesis by decreasing stomatal aperture –not by affecting ATP synthesis. Trends Plant Sci. 2000, 5, 187–188. [Google Scholar] [CrossRef]
- Liang, G.; Liu, J.; Zhang, J.; Guo, J. Effects of drought stress on photosynthetic and physiological parameters of tomato. J. Am. Soc. Hortic. Sci. 2020, 145, 12–17. [Google Scholar] [CrossRef]
- Huang, B.L.; Li, X.; Liu, P.; Ma, L.; Wu, W.; Zhang, X.; Li, Z.; Huang, B. Transcriptomic analysis of Eruca vesicaria subs. sativa lines with contrasting tolerance to polyethylene glycol-simulated drought stress. BMC Plant Biol. 2019, 19, 419. [Google Scholar] [CrossRef]
- Taiz, L.; Møller, I.M.; Murphy, A.; Peer, W.A. Fundamentals of Plant Physiology; Sinauer Associates: Sunderland, MA, USA, 2018; pp. 537–560. [Google Scholar]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought stress in plants: An overview. In Drought Stress Tolerance in Plants; Hossain, M.A., Wani, S., Bhattacharjee, S., Burritt, D., Tran, L.S.P., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 1, pp. 1–33. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Cavarianni, R.L.; Cecílio Filho, A.B.C.; Cazetta, J.O.; May, A.; Corradi, M.M. Nutrient contents and production of rocket as affected by nitrogen concentrations in the nutritive solution. Sci. Agric. 2008, 65, 652–658. [Google Scholar] [CrossRef]
- Fontana, E.; Nicola, S. Traditional and soilless culture systems to produce corn salad (Valerianella olitoria L.) and rocket (Eruca sativa Mill.) with low nitrate content. J. Food Agric. Environ. 2009, 7, 405–410. [Google Scholar]
- Kim, S.J.; Chiami, K.; Ishii, G. Effect of ammonium: Nitrate nutrient ratio on nitrate and glucosinolate contents of hydroponically-grown rocket salad (Eruca sativa Mill.). Soil Sci. Plant Nutr. 2006, 52, 387–393. [Google Scholar] [CrossRef]
- Omirou, M.; Papastefanou, C.; Katsarou, D.; Papastylianou, I.; Passam, H.C.; Ehaliotis, C.; Papadopoulou, K.K. Relationships between nitrogen, dry matter accumulation and glucosinolates in Eruca sativa Mills. The applicability of the critical NO3-N levels approach. Plant Soil 2012, 354, 347–358. [Google Scholar] [CrossRef]
- Barickman, T.C.; Ku, K.-M.; Sams, C.E. Differing precision irrigation thresholds for kale (Brassica oleracea L. var. acephala) induces changes in physiological performance, metabolites, and yield (Brassica oleracea L. var. acephala). Environ. Exp. Bot. 2020, 180, 104253. [Google Scholar] [CrossRef]
- Bennett, R.N.; Mellon, F.A.; Botting, N.P.; Eagles, J.; Rosa, E.A.; Williamson, G. Identification of the major glucosinolate (4-mercaptobutyl glucosinolate) in leaves of Eruca sativa L. (salad rocket). Phytochemistry 2002, 61, 25–30. [Google Scholar] [CrossRef]
- Bell, L.; Lignou, S.; Wagstaff, C. High glucosinolate content in rocket leaves (Diplotaxis tenuifolia and Eruca sativa) after multiple harvests is associated with increased bitterness, pungency, and reduced consumer liking. Foods 2020, 9, 1799. [Google Scholar] [CrossRef] [PubMed]
- D’Antuono, L.F.; Elementi, S.; Neri, R. Exploring new potential health-promoting vegetables: Glucosinolates and sensory attributes of rocket salads and related Diplotaxis and Eruca species. J. Sci. Food Agric. 2009, 89, 713–722. [Google Scholar] [CrossRef]
- Bell, L.; Chadwick, M.; Puranik, M.; Jasper, J.; Tudor, R.; Methven, L.; Wagstaff, C. Genotypes of Eruca vesicaria subsp. sativa grown in contrasting field environments differ on transcriptomic and metabolomic levels, significantly impacting nutritional quality. Front. Plant Sci. 2023, 14, 1218984. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VWC Treatment (m3·m−3) | Plant Height (cm) | Plant Width (cm) | Leaf Size (cm2) | Total Leaf Area (cm2) | Specific Leaf Area (cm2·g−1) | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Root Fresh Weight (g) | Root Dry Weight (g) |
|---|---|---|---|---|---|---|---|---|---|
| 0.50 | 14.1 a | 20.9 | 36.5 a | 281.4 a | 128.8 | 12.0 a | 1.96 ab | 4.4 a | 0.61 |
| 0.40 | 15.6 a | 19.4 | 36.6 a | 238.9 a | 130.1 | 10.6 a | 1.81 b | 3.4 b | 0.51 |
| 0.30 | 14.1 a | 23.0 | 40.1 a | 250.5 a | 126.9 | 11.9 a | 2.05 a | 3.4 b | 0.57 |
| 0.20 | 10.8 b | 19.6 | 29.6 b | 184.7 b | 122.4 | 8.8 b | 1.55 c | 3.1 b | 0.55 |
| p-value | 0.008 | 0.835 | 0.010 | <0.001 | 0.800 | <0.001 | <0.001 | 0.004 | 0.131 |
| Treatment (m3·m−3) | Glucosinolate Amount (μmol·g−1, DW) | ||||||
|---|---|---|---|---|---|---|---|
| Glucoraphanin | Diglucothiobeinin | Glucoibervirin | Glucoerucin | Glucosativin | Dimeric Glucosativin | Total | |
| 0.50 | 6.19 | 1.73 | 0.15 | 8.31 a | 7.30 | 6.04 | 29.7 |
| 0.40 | 4.87 | 2.14 | 0.18 | 8.52 a | 7.87 | 7.44 | 31.0 |
| 0.30 | 4.24 | 1.99 | 0.10 | 6.65 ab | 6.46 | 8.20 | 27.7 |
| 0.20 | 5.01 | 1.60 | 0.06 | 5.27 b | 6.15 | 6.05 | 24.9 |
| p-value | 0.242 | 0.427 | 0.244 | 0.012 | 0.528 | 0.168 | 0.089 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.; An, S.K.; Ku, K.-M.; Kim, J. The Optimum Substrate Moisture Level to Enhance the Growth and Quality of Arugula (Eruca sativa). Horticulturae 2024, 10, 483. https://doi.org/10.3390/horticulturae10050483
Lee K, An SK, Ku K-M, Kim J. The Optimum Substrate Moisture Level to Enhance the Growth and Quality of Arugula (Eruca sativa). Horticulturae. 2024; 10(5):483. https://doi.org/10.3390/horticulturae10050483
Chicago/Turabian StyleLee, Kiram, Seong Kwang An, Kang-Mo Ku, and Jongyun Kim. 2024. "The Optimum Substrate Moisture Level to Enhance the Growth and Quality of Arugula (Eruca sativa)" Horticulturae 10, no. 5: 483. https://doi.org/10.3390/horticulturae10050483
APA StyleLee, K., An, S. K., Ku, K.-M., & Kim, J. (2024). The Optimum Substrate Moisture Level to Enhance the Growth and Quality of Arugula (Eruca sativa). Horticulturae, 10(5), 483. https://doi.org/10.3390/horticulturae10050483