
The Myosin Myocardial Mesh Interpreted as a Biological Analogous of Nematic Chiral Liquid Crystals †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Background: Materials and Methods
2.1. Ethics Statement
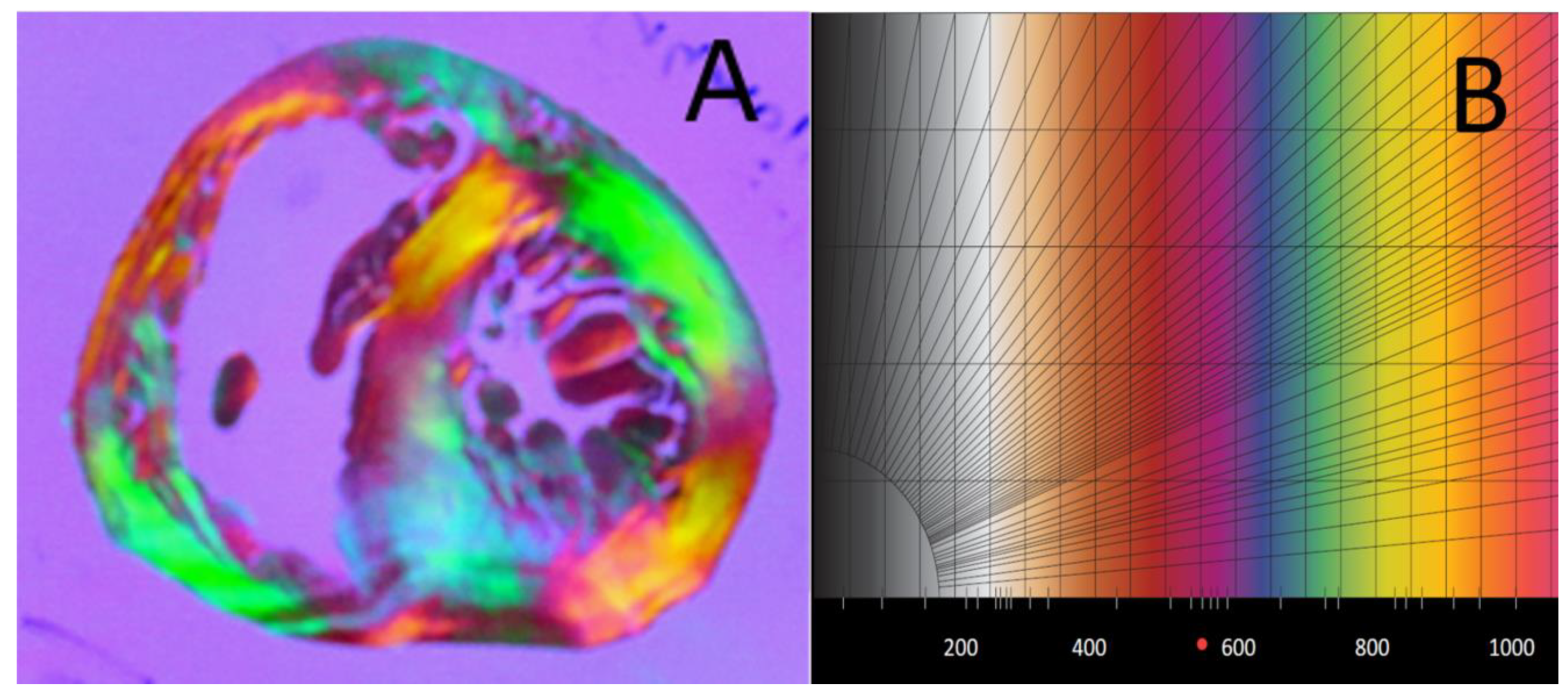
2.2. Polarised Light Imaging (PLI) and Histological Preparation
- -
- the azimuth angle (az), that is the angle between the east–west axis of the optical bench stage and the projection of the measured mean orientation of a few thousands of myocardial cells in an elementary point of the image (voxel) on the stage plane (0°–180°);
- -
- and the elevation angle (el), that is the angle corresponding to the obliquity of the voxel principal orientation, with respect to the plane of the section; in other words, it is the way the voxel principal orientation escapes from the section plane (−90° to +90°).
2.3. Alternative Optical Methods of Studying 3D Myocyte Organisation
2.4. Mapping
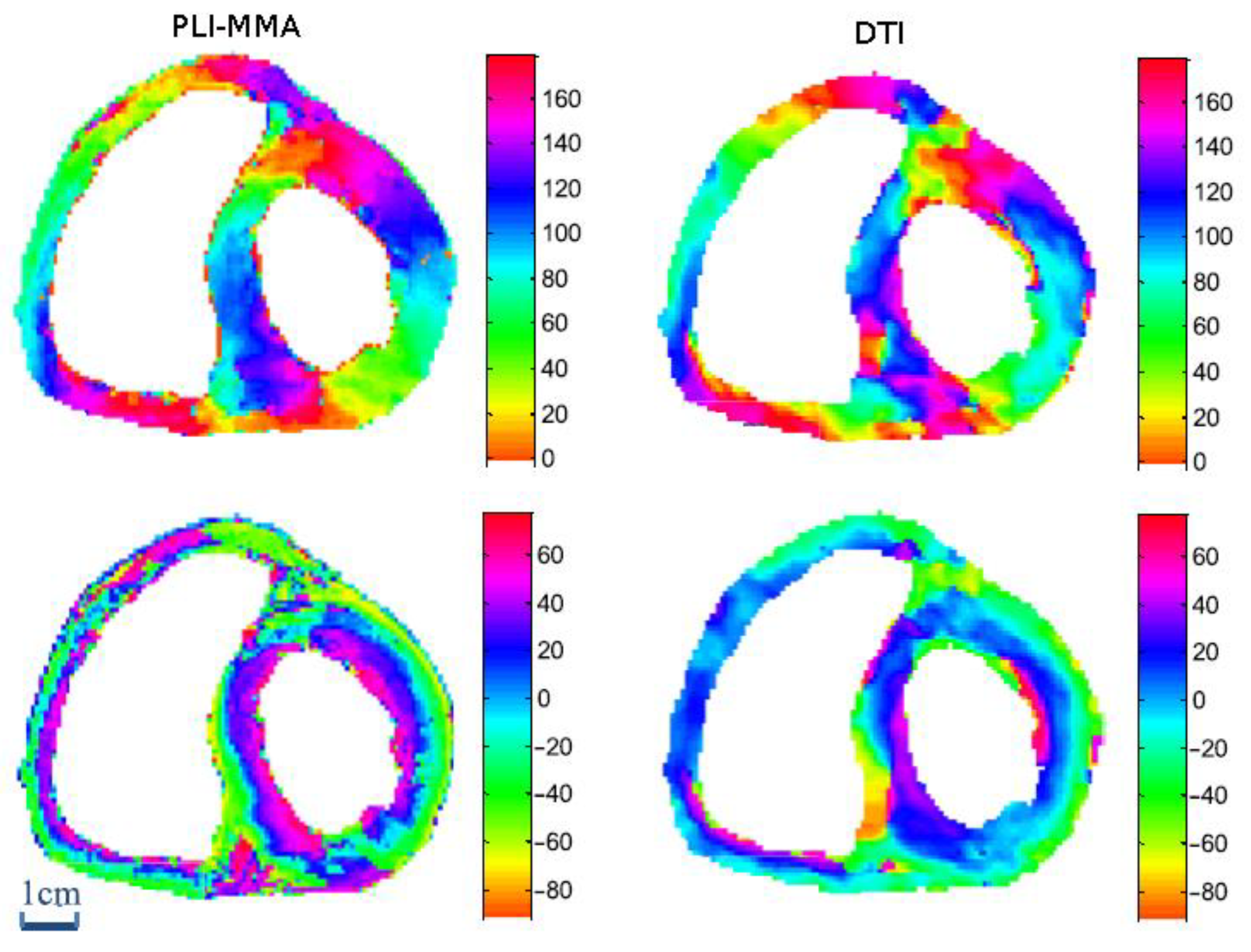
2.5. Comparison between PLI-MMA and Diffusion Tensor Imaging (DTI) Measurements
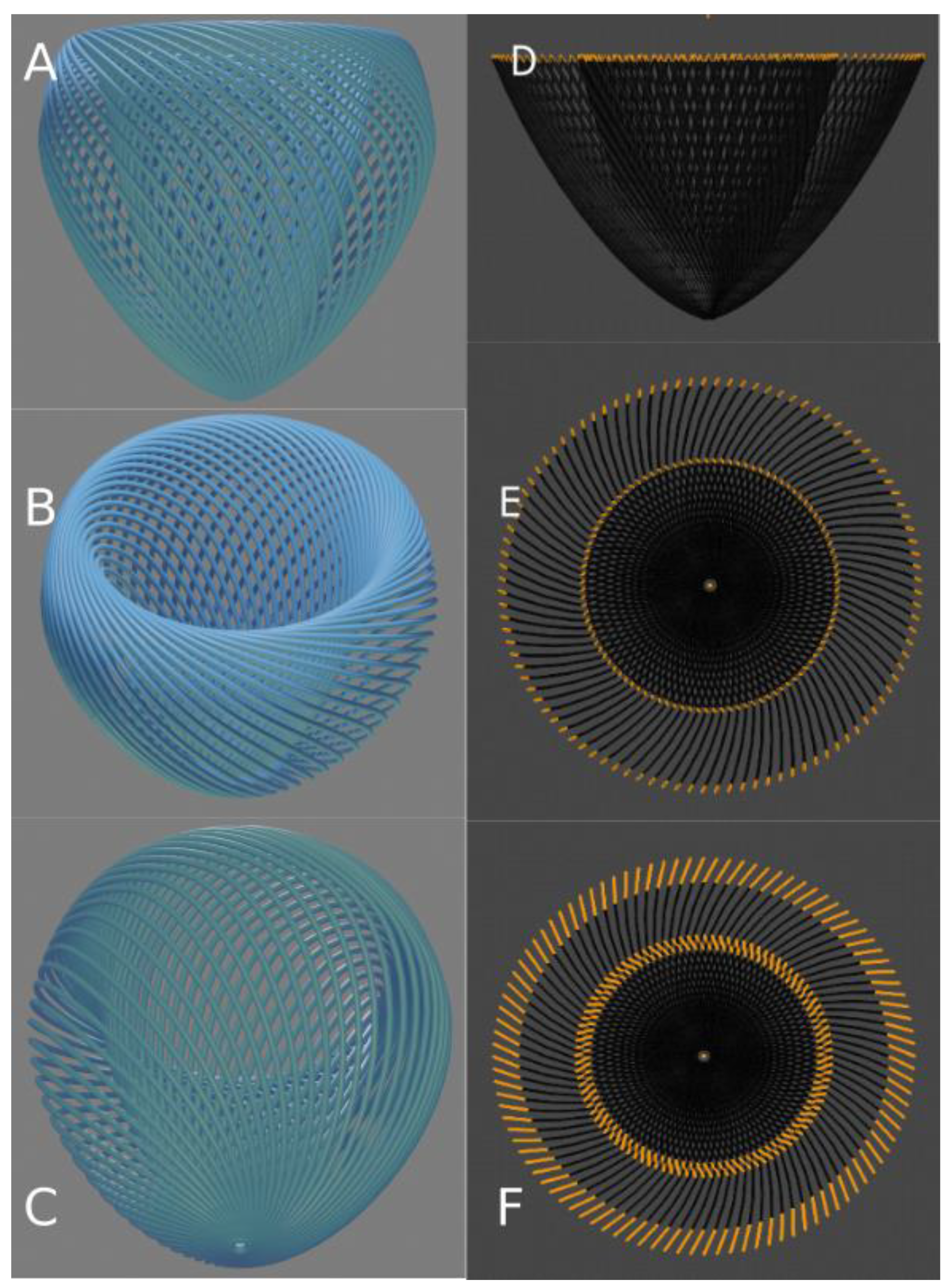
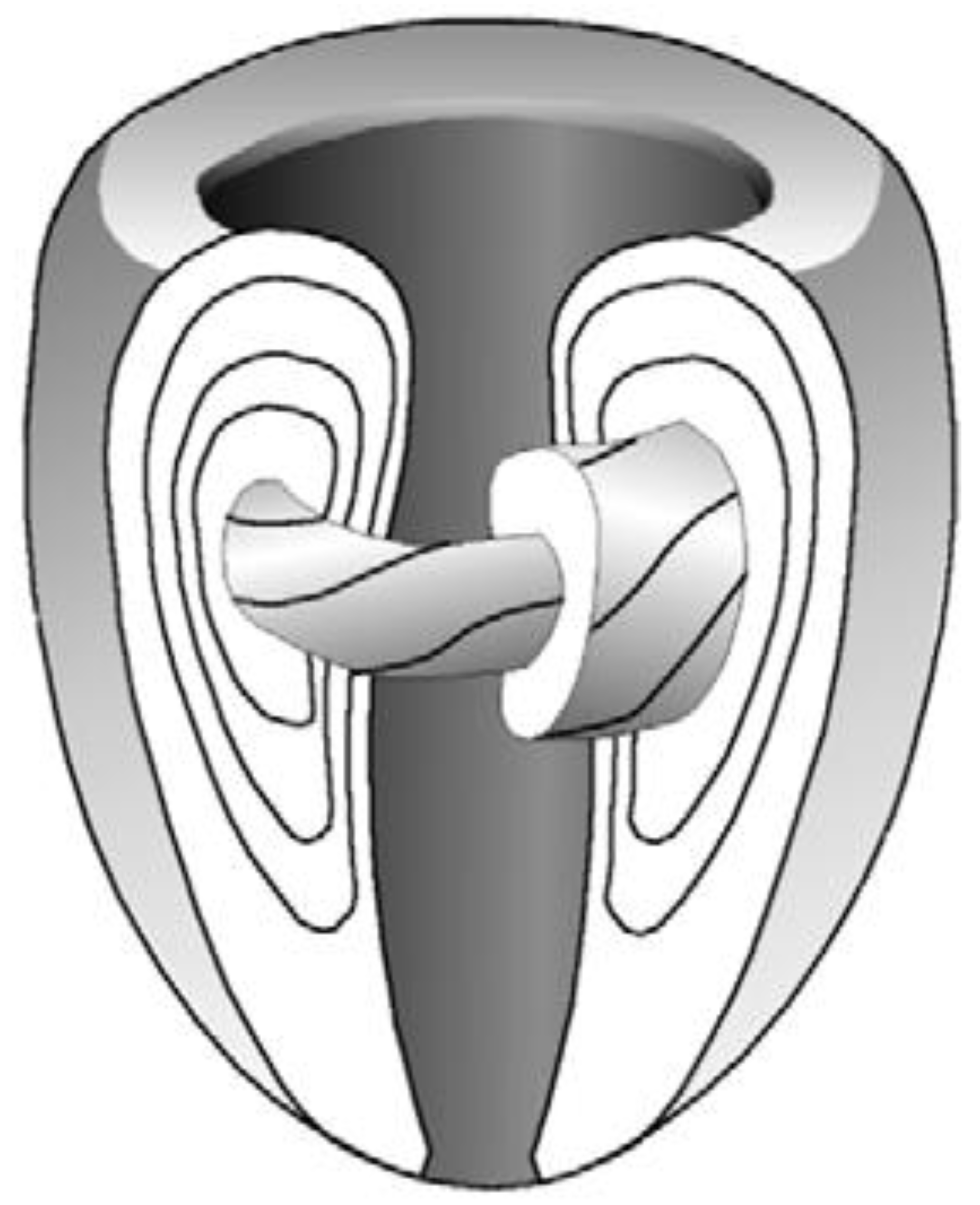
3. The Insights of 1979 Streeter’s Conjecture: “Fibres Run like Geodesics on a Nested Set of Toroidal Bodies of Revolution”
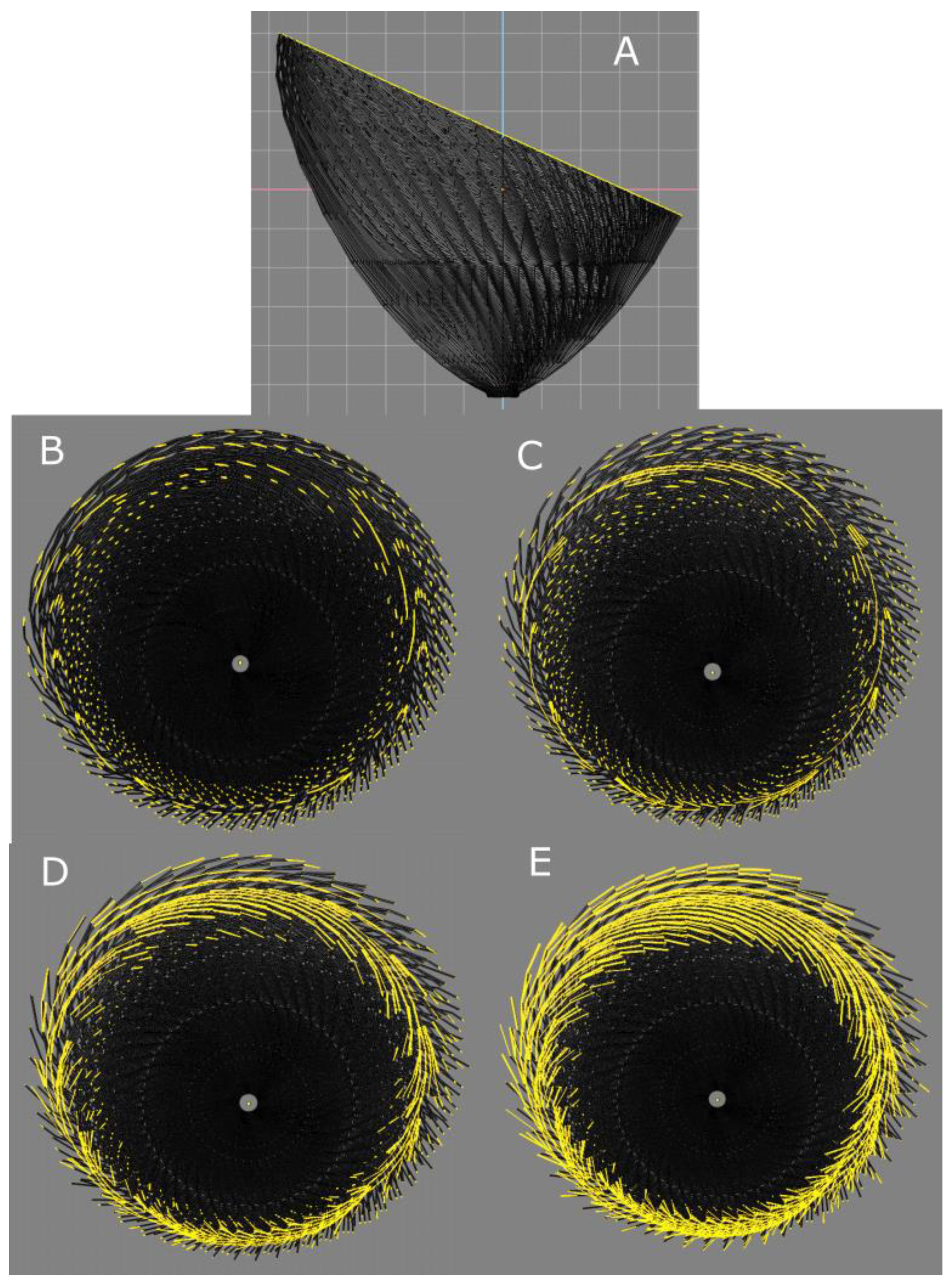
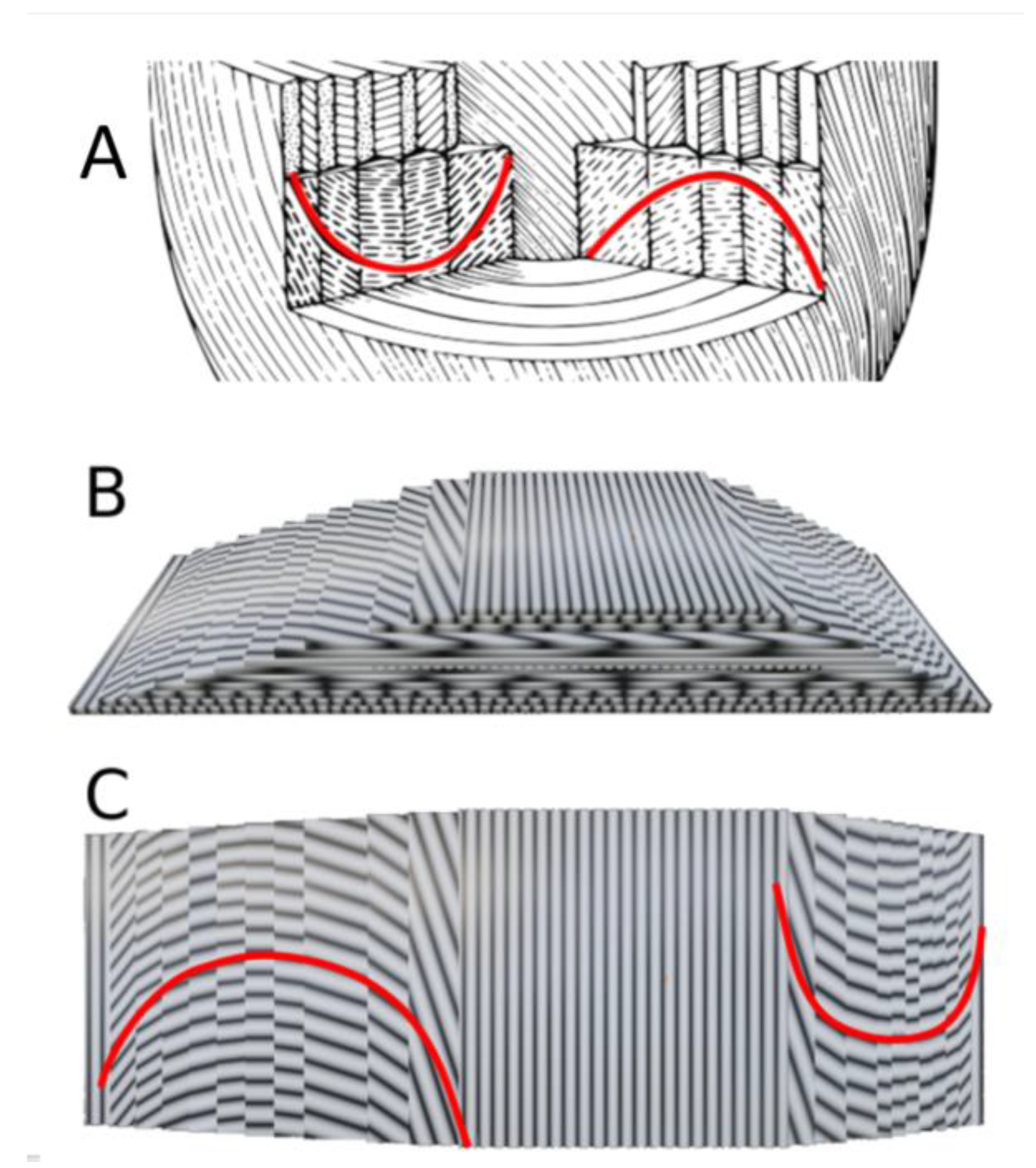
3.1. Validation by PLI-MMA of 1979 Streeter’s Conjecture
3.2. The Integrative Value of Nested Tori Streeter’s Model
3.3. Prospective Value of Streeter’s Nested Tori Model
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Stephenson, R.; Agger, P.; Omann, C.; Sanchez-Quintana, D.; Jarvis, J.; Anderson, R. Resolving the True Ventricular Mural Architecture. J. Cardiovasc. Dev. Dis. 2018, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Dejea, H.; Bonnin, A.; Cook, A.C.; Garcia-Canadilla, P. Cardiac multi-scale investigation of the right and left ventricle ex vivo: A review. Cardiovasc. Diagn. Ther. 2020, 10, 1701–1717. [Google Scholar] [CrossRef]
- Sosnovik, D.E. Magnetic Resonance-Based Characterization of Myocardial Architecture. Heart Fail. Clin. 2021, 17, 85–101. [Google Scholar] [CrossRef]
- Michel-Levy, A.; Lacroix, A. Les Minéraux Des Roches; Librairie polytechnique: Paris, France, 1888; p. 334. [Google Scholar]
- Canham, P.B.; Finlay, H.M.; Whittaker, P.; Starkey, J. The tunica muscularis of human brain arteries: Three-dimensional measurements of alignment of the smooth muscle mechanical axis, by polarized light and the universal stage. Neurol. Res. 1986, 8, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S. Appendix III An introduction to biological polarization microscopy. In Video Microscopy; Springer: Berlin/Heidelberg, Germany, 1986; pp. 477–509. [Google Scholar]
- Haskell, R.C.; Carlson, F.D.; Blank, P.S. Form birefringence of muscle. Biophys. J. 1989, 56, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Usson, Y.; Parazza, F.; Jouk, P.-S.; Michalowicz, G. Method for the study of the three-dimensional orientation of the nuclei of myocardial cells in fetal human heart by means of confocal scanning laser microscopy. J. Microsc. 1994, 174, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Jouk, P.-S.; Usson, Y.; Michalowicz, G.; Parazza, F. Mapping of the orientation of myocardial cells by means of polarized light and confocal scanning laser microscopy. Microsc. Res. Tech. 1995, 30, 480–490. [Google Scholar] [CrossRef]
- Jouk, P.-S.; Usson, Y.; Michalowicz, G.; Grossi, L. Three-dimensional cartography of the pattern of the myofibres in the second trimester fetal human heart. Anat. Embryol. 2000, 202, 103–118. [Google Scholar] [CrossRef]
- Audain, D.P. Simulation De L’imagerie En Lumière Polarisée: Application à L’étude De L’architecture Des “Fibres” Du Myocarde Humain. Ph.D. Thesis, Grenoble-Alpes University, Grenoble, France, 2014. [Google Scholar]
- Desrosiers, P.A.; Michalowicz, G.; Jouk, P.-S.; Usson, Y.; Zhu, Y. Modeling of the Optical Behavior of Myocardial Fibers in Polarized Light Imaging. In Statistical Atlases and Computational Models of the Heart Imaging and Modelling Challenges; Camara, O., Mansi, T., Pop, M., Rhode, K., Sermesant, M., Young, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 235–244, (Lecture Notes in Computer Science). [Google Scholar]
- Sun, T.; Liu, T.; He, H.; Wu, J.; Ma, H. Distinguishing anisotropy orientations originated from scattering and birefringence of turbid media using Mueller matrix derived parameters. Opt. Lett. 2018, 43, 4092–4095. [Google Scholar] [CrossRef]
- Li, P.; Lv, D.; He, H.; Ma, H. Separating azimuthal orientation dependence in polarization measurements of anisotropic media. Opt. Express 2018, 26, 3791–3800. [Google Scholar] [CrossRef]
- Qi, J.; He, H.; Ma, H.; Elson, D.S. Extended polar decomposition method of Mueller matrices for turbid media in reflection geometry. Opt. Lett. 2017, 42, 4048–4051. [Google Scholar] [CrossRef]
- Agger, P.; Stephenson, R.S. Assessing Myocardial Architecture: The Challenges and Controversies. J. Cardiovasc. Dev. Dis. 2020, 7, 47. [Google Scholar] [CrossRef]
- Cabral, B.; Leedom, L.C. Imaging Vector Fields using Line Integral Convolution. In Proceedings of the 20th Annual Conference on Computer Graphics and Interactive Techniques, SIGGRAPH, Anaheim, CA, USA, 2–6 August 1993; pp. 263–270. [Google Scholar]
- Yang, F.; Zhu, Y.-M.; Michalowicz, G.; Jouk, P.-S.; Fanton, L.; Viallon, M.; Clarysse, P.; Croisille, P.; Usson, Y. Quantitative comparison of human myocardial fiber orientations derived from DTI and polarized light imaging. Phys. Med. Biol. 2018, 63, 215003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desrosiers, P.A.; Michalowicz, G.; Jouk, P.-S.; Usson, Y.; Zhu, Y. Study of myocardial cell inhomogeneity of the human heart: Simulation and validation using polarized light imaging: Study of myocardial cell inhomogeneity of the human heart. Med. Phys. 2016, 43, 2273–2282. [Google Scholar] [CrossRef]
- Daniel, D.S., Jr. Gross morphology and fiber geometry of the heart. In Handbook of Physiology; Section 2: The cardiovascular system; American physiological society: Bethesda, Williams; Wilkins, Baltimore, 1979; Volume I: The heart, pp. 61–112. [Google Scholar]
- Streeter, D.D.; Spotnitz, H.M.; Patel, D.P.; Ross, J.; Sonnenblick, E.H. Fiber orientation in the canine left ventricle during diastole and systole. Circ. Res. 1969, 24, 339–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krehl, L. Beiträge zur Kenntnis der Füllung und Entleerung des Herzens; Abhandl. Math. Phys. Kl. Königlichen Sachs. Ges. Wiss 29: 341-362: 1891; S. Hirzel Verlag: Leipzig, Germany, 1891. [Google Scholar]
- Lunkenheimer, P.P.; Redmann, K.; Kling, N.; Jiang, X.; Rothaus, K.; Cryer, C.W.; Wübbeling, F.; Niederer, P.; Heitz, P.U.; Yen Ho, S.; et al. Three-dimensional architecture of the left ventricular myocardium. Anat. Rec. A. Discov. Mol. Cell. Evol. Biol. 2006, 288A, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Peskin, C.S. Fiber architecture of the left ventricular wall: An asymptotic analysis. Commun. Pure Appl. Math. 1989, 42, 79–113. [Google Scholar] [CrossRef]
- Jouk, P.-S.; Mourad, A.; Milisic, V.; Michalowicz, G.; Raoult, A.; Caillerie, D.; Usson, Y. Analysis of the fiber architecture of the heart by quantitative polarized light microscopy. Accuracy, limitations and contribution to the study of the fiber architecture of the ventricles during fetal and neonatal life☆. Eur. J. Cardiothorac. Surg. 2007, 31, 915–921. [Google Scholar] [CrossRef]
- Mourad, A. Description topologique de l’architecture fibreuse et modelisation mecanique du myocarde. Univ. Grenoble-Alpe Inst. Natl. Polytech. Grenoble 2003, 204. [Google Scholar]
- Mourad, A.; Biard, L.; Caillerie, D.; Jouk, P.-S.; Raoult, A.; Szafran, N.; Usson, Y. Geometrical Modelling of the Fibre Organization in the Human Left Ventricle. In Functional Imaging and Modeling of the Heart; Katila, T., Nenonen, J., Magnin, I.E., Clarysse, P., Montagnat, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; Volume 2230, pp. 32–38, (Lecture Notes in Computer Science); Available online: http://link.springer.com/10.1007/3-540-45572-8_5 (accessed on 6 December 2021).
- George, E.P. Box Science and statistics. J. Am. Stat. Assoc. 1976, 71, 791–799. [Google Scholar]
- Jean, R. Croissance Végétale Et Morphogénèse; Presses de l’Université du Québec: Paris, France, 1983. [Google Scholar]
- Douady, S.; Couder, Y. Phyllotaxis as a physical self-organized growth process. Phys. Rev. Lett. 1992, 68, 2098–2101. [Google Scholar] [CrossRef] [PubMed]
- Feneis, H. Das gefüge des herzmuskels bei systole und diastole. Morphol. Jahrb. 1943, 89, 371–406. [Google Scholar]
- Bouligand, Y. Sur une architecture torsadée répandue dans de nombreuses cellules d’arthropodes. CR Acad. Sci. 1965, 46, 3665–3701. [Google Scholar]
- Gray, G.W. Molecular Structure and the Properties of Liquid Crystals; Academic Press: London, UK, 1962; p. 338. [Google Scholar]
- Bouligand, Y. Recherches sur les textures des états mésomorphes-2.—Les champs polygonaux dans les cholestériques. J. Phys. 1972, 33, 715–736. [Google Scholar] [CrossRef]
- Bouligand, Y. Liquid crystals and biological morphogenesis: Ancient and new questions. Comptes Rendus Chim. 2008, 11, 281–296. [Google Scholar] [CrossRef]
- Saw, T.B.; Xi, W.; Ladoux, B.; Lim, C.T. Biological Tissues as Active Nematic Liquid Crystals. Adv. Mater. 2018, 30, 1802579. [Google Scholar] [CrossRef]
- O’Keeffe, K.P.; Hong, H.; Strogatz, S.H. Oscillators that sync and swarm. Nat. Commun. 2017, 8, 1504. [Google Scholar] [CrossRef] [PubMed]
- Duclos, G.; Adkins, R.; Banerjee, D.; Peterson, M.S.E.; Varghese, M.; Kolvin, I.; Baskaran, A.; Pelcovits, R.A.; Powers, T.R.; Baskaran, A.; et al. Topological structure and dynamics of three-dimensional active nematics. Science 2020, 367, 1120–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandle, R.J. The Shape of Things To Come: The Formation of Modulated Nematic Mesophases at Various Length Scales. Chem.-Eur. J. 2017, 23, 8771–8779. [Google Scholar] [CrossRef] [Green Version]
- Charvolin, J.; Sadoc, J.-F. About collagen, a tribute to Yves Bouligand. Interface Focus 2012, 2, 567–574. [Google Scholar] [CrossRef]
- Dixon, A.W.; Taberner, A.J.; Nash, M.P.; Nielsen, P.M.F. Quantifying optical anisotropy in soft tissue membranes using Mueller matrix imaging. J. Biomed. Opt. 2021, 26, 18. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jouk, P.-S.; Usson, Y. The Myosin Myocardial Mesh Interpreted as a Biological Analogous of Nematic Chiral Liquid Crystals. J. Cardiovasc. Dev. Dis. 2021, 8, 179. https://doi.org/10.3390/jcdd8120179
Jouk P-S, Usson Y. The Myosin Myocardial Mesh Interpreted as a Biological Analogous of Nematic Chiral Liquid Crystals. Journal of Cardiovascular Development and Disease. 2021; 8(12):179. https://doi.org/10.3390/jcdd8120179
Chicago/Turabian StyleJouk, Pierre-Simon, and Yves Usson. 2021. "The Myosin Myocardial Mesh Interpreted as a Biological Analogous of Nematic Chiral Liquid Crystals" Journal of Cardiovascular Development and Disease 8, no. 12: 179. https://doi.org/10.3390/jcdd8120179
APA StyleJouk, P.-S., & Usson, Y. (2021). The Myosin Myocardial Mesh Interpreted as a Biological Analogous of Nematic Chiral Liquid Crystals. Journal of Cardiovascular Development and Disease, 8(12), 179. https://doi.org/10.3390/jcdd8120179