The Hypothesis of “Embryonic Recall”: Mechanotransduction as Common Denominator Linking Normal Cardiogenesis to Recovery in Adult Failing Hearts


 ,
,
Abstract
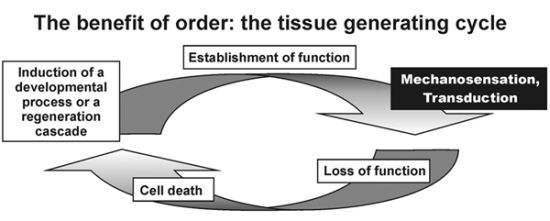
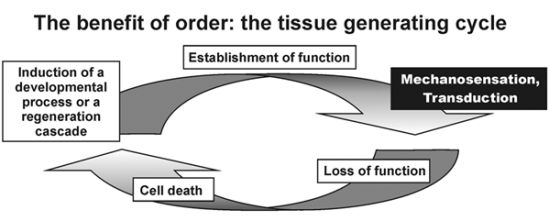
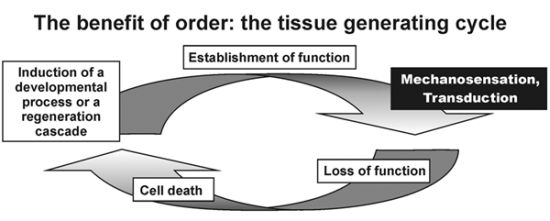
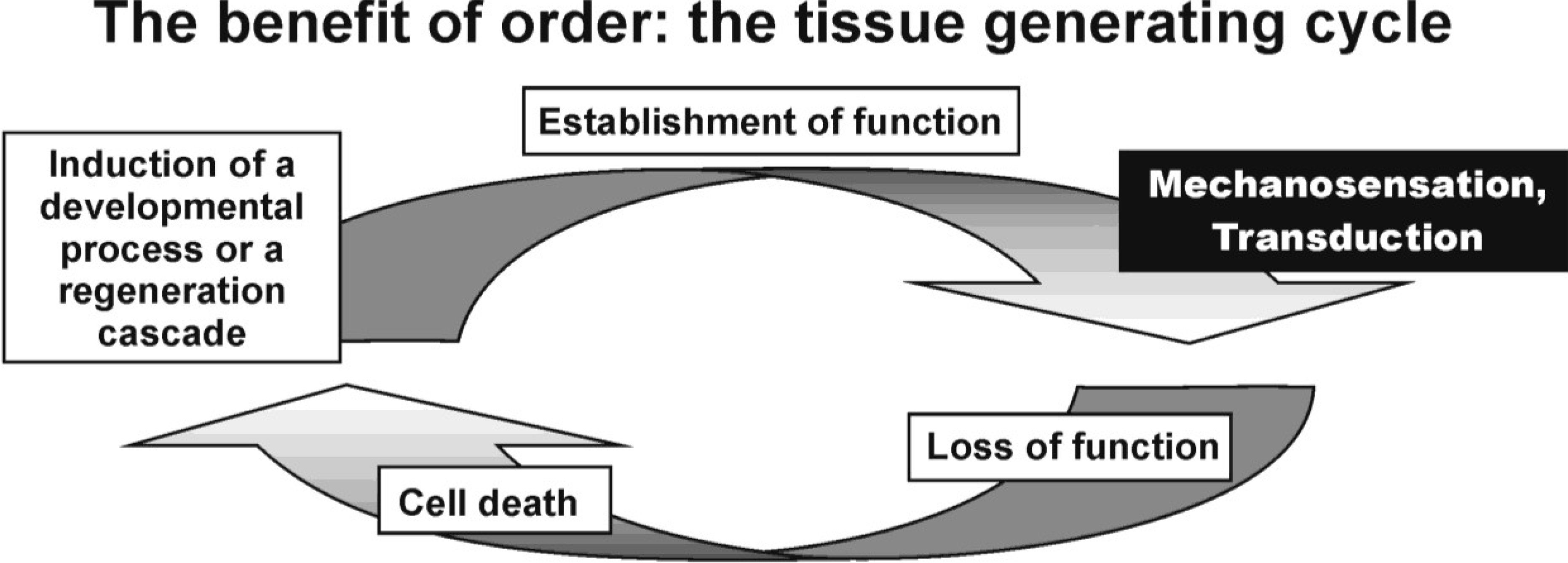
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
1.1. An Embryonic Developmental Process as Impulse for Regeneration
1.2. Hemodynamic Force Sculpturing the Developing Heart
1.3. The Term “Embryonic Recall”
1.4. Gain of Function Sensing Sculpturing Tissue Maturation
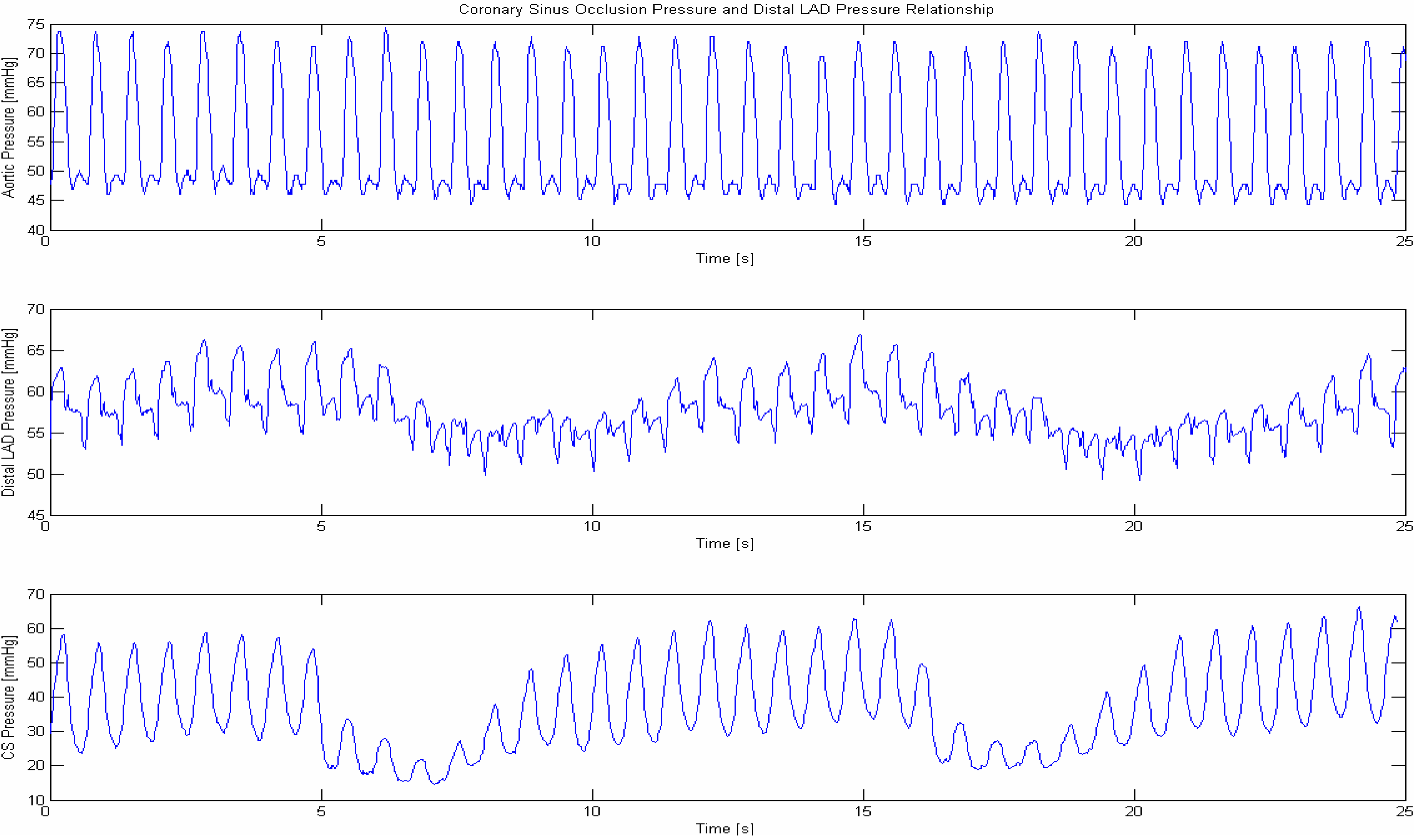
1.5. Pressure Controlled Coronary Sinus Occlusion (PICSO)
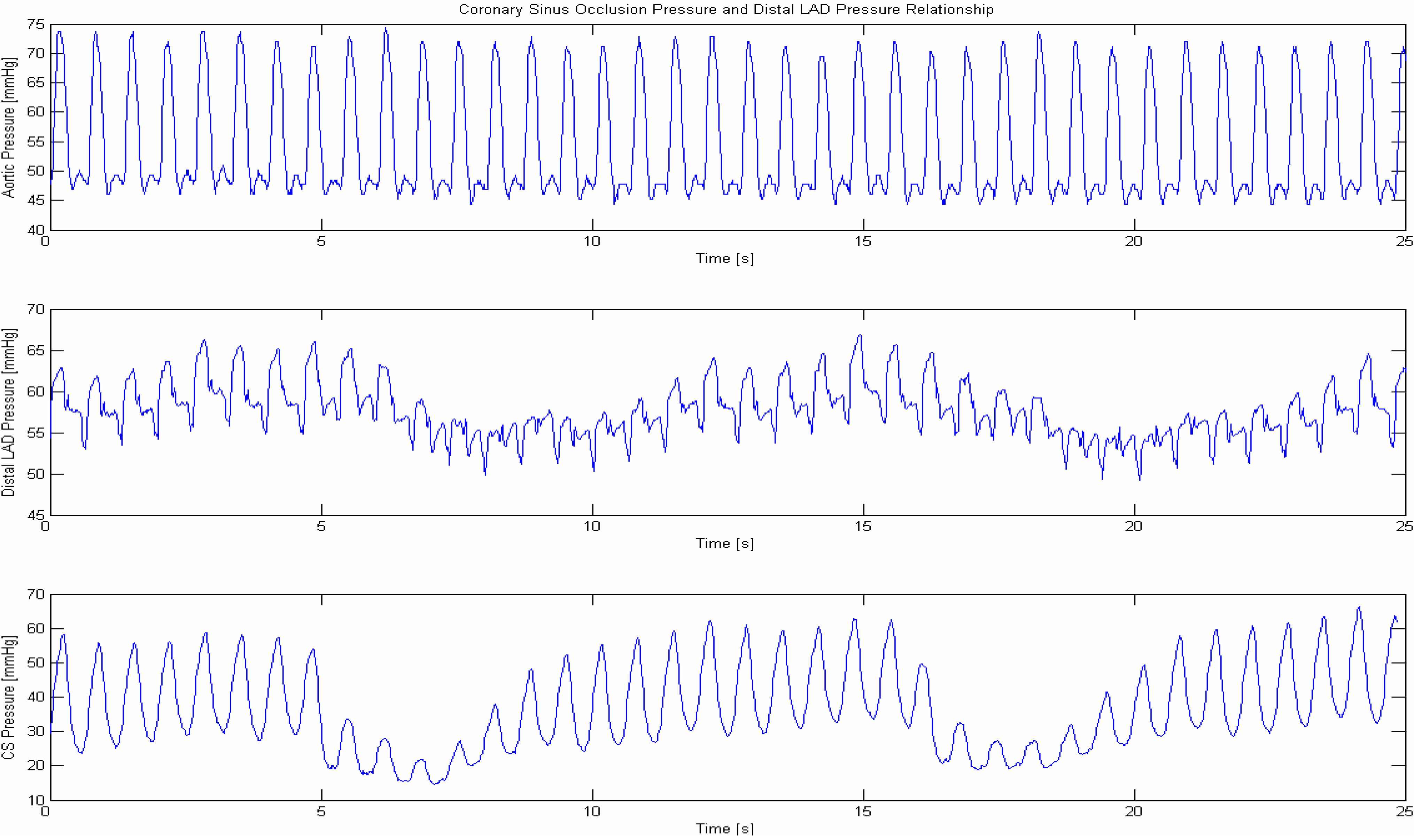
2. Periodic Elevation of Coronary Sinus Pressure
3. Results and Discussion
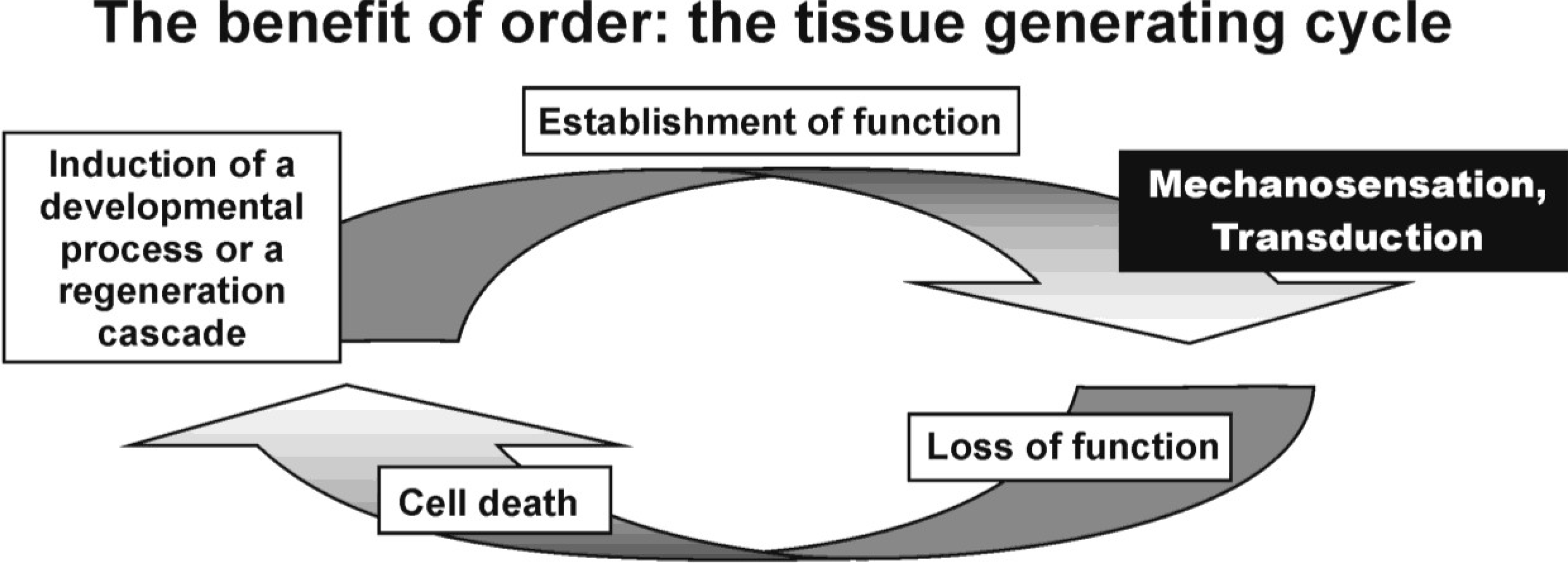
Structure Defines Function and Function Creates Structure
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Schneider, M.D. Development. A cardiac nonproliferation treaty. Science 2011, 332, 426–427. [Google Scholar] [CrossRef]
- Bergmann, O.; Bhardwaj, R.D.; Bernard, S.; Zdunek, S.; Barnabé-Heider, F.; Walsh, S.; Zupicich, J.; Alkass, K.; Buchholz, B.A.; Druid, H.; Jovinge, S.; Frisén, J. Evidence for cardiomyocyte renewal in humans. Science 2009, 324, 98–102. [Google Scholar] [CrossRef]
- Noseda, M.; Schneider, M.D. Fibroblasts inform the heart: Control of cardiomyocyte cycling and size by age-dependent paracrine signals. Dev. Cell. 2009, 16, 161–162. [Google Scholar] [CrossRef]
- Morrisey, E.E. Not too large and not too small—Just the right size: A hippo-sized heart. Circ. Res. 2011, 109, 614–615. [Google Scholar] [CrossRef]
- Leri, A.; Anversa, P.; Frishman, W.H. Cardiovascular Regeneration and Stem Cell Therapy. Blackwell Futura: Malden, MA, USA, 2007. [Google Scholar]
- Mohl, W.; Mina, S.; Milasinovic, D.; Kasahara, H.; Wei, S.; Maurer, G. Is activation of coronary venous cells the key to cardiac regeneration? Nat. Clin. Pract. Cardiovasc. Med. 2008, 5, 528–530. [Google Scholar] [CrossRef]
- Miyasaka, K.Y.; Kida, Y.S.; Banjo, T.; Ueki, Y.; Nagayama, K.; Matsumoto, T.; Sato, M.; Ogura, T. Heartbeat regulates cardiogenesis by suppressing retinoic acid signaling via expression of miR-143. Mech. Dev. 2011, 128, 18–28. [Google Scholar] [CrossRef]
- Granados-Riveron, J.T.; Brook, J.D. The impact of mechanical forces in heart morphogenesis. Circ. Cardiovasc. Genet. 2012, 5, 132–142. [Google Scholar] [CrossRef]
- Fahed, A.C.; Gelb, B.D.; Seidman, J.G.; Seidman, C.E. Genetics of congenital heart disease: The glass half empty. Circ. Res. 2013, 112, 707–720. [Google Scholar] [CrossRef]
- Sasai, Y. Cytosystems dynamics in self-organization of tissue architecture. Nature 2013, 493, 318–326. [Google Scholar] [CrossRef]
- Sperling, S.R. Systems biology approaches to heart development and congenital heart disease. Cardiovasc. Res. 2011, 91, 269–278. [Google Scholar] [CrossRef]
- Mohl, W. Embryonic recall: Myocardial regeneration beyond stem cell transplantation. Wien. Klin. Wochenschr. 2007, 119, 333–336. [Google Scholar] [CrossRef]
- Mohl, W.; Mayr, W.R.; Hauser, G.; Reuer, E.; Wimmer, M.; Herbich, J. Mechanisms common to the development of malformation in congenital and sporadic forms of atrial septal defect (type II) (in German). Wien. Klin. Wochenschr. 1979, 91, 307–314. [Google Scholar]
- Schluter, K.D.; Piper, H.M. Regulation of growth in the adult cardiomyocytes. FASEB J. 1999, 13, S17–S22. [Google Scholar]
- Wu, B.; Zhang, Z.; Lui, W.; Chen, X.; Wang, Y.; Chamberlain, A.A.; Moreno-Rodriguez, R.A.; Markwald, R.R.; O’Rourke, B.P.; Sharp, D.J.; Zheng, D.; Lenz, J.; Baldwin, H.S.; Chang, C.P.; Zhou, B. Endocardial Cells Form the Coronary Arteries by Angiogenesis through Myocardial-Endocardial VEGF Signaling. Cell 2012, 151, 1083–1096. [Google Scholar]
- del Monte, G.; Harvey, R.P. An Endothelial Contribution to Coronary Vessels. Cell 2012, 151, 932–934. [Google Scholar] [CrossRef]
- Zheng, Z.M.; Wang, X. Regulation of Cellular miRNA Expression by Human Papillomaviruses. Biochim. Biophys. Acta 2011, 1809, 668–677. [Google Scholar] [CrossRef]
- Eulalio, A.; Mano, M.; Dal Ferro, M.; Zentilin, L.; Sinagra, G.; Zacchigna, S.; Giacca, M. Functional screening identifies miRNAs inducing cardiac regeneration. Nature 2012, 492, 376–381. [Google Scholar] [CrossRef]
- Bauersachs, J.; Thum, T. Biogenesis and Regulation of Cardiovascular microRNAs. Circ. Res. 2011, 109, 334–347. [Google Scholar] [CrossRef]
- Jayawardena, T.M.; Egemnazarov, B.; Finch, E.A.; Zhang, L.; Payne, J.A.; Pandya, K.; Zhang, Z.; Rosenberg, P.; Mirotsou, M.; Dzau, V.J. MicroRNA-mediated in vitro and in vivo direct reprogramming of cardiac fibroblasts to cardiomyocytes. Circ. Res. 2012, 110, 1465–1473. [Google Scholar] [CrossRef]
- Mohl, W.; Kajgana, I.; Bergmeister, H.; Rattay, F. Intermittent pressure elevation of the coronary venous system as a method to protect ischemic myocardium. Interact. Cardiovasc. Thorac. Surg. 2005, 4, 66–69. [Google Scholar] [CrossRef]
- Campàs, O.; Mammoto, T.; Hasso, S.; Sperling, R.A.; O’Connell, D.; Bischof, A.G.; Maas, R.; Weitz, D.A.; Mahadevan, L.; Ingber, D.E. Quantifying cell-generated mechanical forces within living embryonic tissues. Nat. Meth. 2014, 11, 183–189. [Google Scholar]
- Mammoto, A.; Mammoto, T.; Ingber, D.E. Mechanosensitive mechanisms in transcriptional regulation. J. Cell Sci. 2012, 125, 3061–3073. [Google Scholar] [CrossRef]
- Poelmann, R.E.; Gittenberger-de Groot, A.C.; Mentink, M.M.; Bökenkamp, R.; Hogers, B. Development of the cardiac coronary vascular endothelium, studied with antiendothelial antibodies, in chicken-quail chimeras. Circ. Res. 1993, 73, 559–568. [Google Scholar] [CrossRef]
- Poelmann, R.E.; van der Heiden, K.; Groot, A.G.; Hierck, B.P. Deciphering the Endothelial Shear Stress Sensor. Circulation 2008, 117, 1124–1126. [Google Scholar] [CrossRef]
- Groenendijk, B.C.; Hierck, B.P.; Vrolijk, J.; Baiker, M.; Pourquie, M.J.; Gittenberger-de Groot, A.C.; Poelmann, R.E. Changes in Shear Stress-Related Gene Expression after Experimentally Altered Venous Return in the Chicken Embryo. Circ. Res. 2005, 96, 1291–1298. [Google Scholar] [CrossRef]
- Mohl, W. The momentum of coronary sinus interventions clinically. Circulation 1988, 77, 6–12. [Google Scholar] [CrossRef]
- Mohl, W. The development and rationale of pressure-controlled intermittent coronary sinus occlusion—A new approach to protect ischemic myocardium. Wien. Klin. Wochenschr. 1984, 96, 20–25. [Google Scholar]
- Mohl, W. Coronary sinus interventions: from concept to clinics. J. Card. Surg. 1987, 2, 467–493. [Google Scholar] [CrossRef]
- Syeda, B.; Schukro, C.; Heinze, G.; Modaressi, K.; Glogar, D.; Maurer, G.; Mohl, W. The salvage potential of coronary sinus interventions: Meta-analysis and pathophysiologic consequences. J. Thorac. Cardiovasc. Surg. 2004, 127, 1703–1712. [Google Scholar] [CrossRef]
- Weigel, G.; Kajgana, I.; Bergmeister, H.; Riedl, G.; Glogar, H.D.; Gyöngyösi, M.; Blasnig, S.; Heinze, G.; Mohl, W. Beck and back: A paradigm change in coronary sinus interventions—Pulsatile stretch on intact coronary venous endothelium. J. Thorac. Cardiovasc. Surg. 2007, 133, 1581–1587. [Google Scholar] [CrossRef]
- de Jonge, M.; Gittenberger-de Groot, A.C.; Wisse, L.J.; Mina, S.; Kasahara, H.; Mohl, W. Mechanical activation of coronary venous endothelium causes a rapid angiogenic impulse after induced myocardial infarction. Eur. Heart J. 2008, 29, 686. [Google Scholar] [CrossRef]
- Khatami, N.; Wadowski, P.; Wagh, V.; Hescheler, J.; Sachinidis, A.; Mohl, W. Pressure-controlled intermittent coronary sinus occlusion (PICSO) study on mechanical control of cardiac tissue morphogenesis. Cardiovasc. Res. 2012, 93 (Suppl. 1), S56. [Google Scholar]
- Mohl, W.; Komamura, K.; Kasahara, H.; Heinze, G.; Glogar, D.; Hirayama, A.; Kodama, K. Myocardial protection via the coronary sinus. Circ. J. 2008, 72, 526–533. [Google Scholar]
- Mohl, W.; Mina, S.; Milasinovic, D.; Kasahara, H.; Wei, S. The legacy of coronary sinus interventions: Endogenous cardioprotection and regeneration beyond stem cell research. J. Thorac. Cardiovasc. Surg. 2008, 136, 1131–1135. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mohl, W.; Milasinovic, D.; Aschacher, T.; Jusic, A.; Maimaitiaili, A.; Rattay, F. The Hypothesis of “Embryonic Recall”: Mechanotransduction as Common Denominator Linking Normal Cardiogenesis to Recovery in Adult Failing Hearts. J. Cardiovasc. Dev. Dis. 2014, 1, 73-82. https://doi.org/10.3390/jcdd1010073
Mohl W, Milasinovic D, Aschacher T, Jusic A, Maimaitiaili A, Rattay F. The Hypothesis of “Embryonic Recall”: Mechanotransduction as Common Denominator Linking Normal Cardiogenesis to Recovery in Adult Failing Hearts. Journal of Cardiovascular Development and Disease. 2014; 1(1):73-82. https://doi.org/10.3390/jcdd1010073
Chicago/Turabian StyleMohl, Werner, Dejan Milasinovic, Thomas Aschacher, Alem Jusic, Abudunaibi Maimaitiaili, and Frank Rattay. 2014. "The Hypothesis of “Embryonic Recall”: Mechanotransduction as Common Denominator Linking Normal Cardiogenesis to Recovery in Adult Failing Hearts" Journal of Cardiovascular Development and Disease 1, no. 1: 73-82. https://doi.org/10.3390/jcdd1010073
APA StyleMohl, W., Milasinovic, D., Aschacher, T., Jusic, A., Maimaitiaili, A., & Rattay, F. (2014). The Hypothesis of “Embryonic Recall”: Mechanotransduction as Common Denominator Linking Normal Cardiogenesis to Recovery in Adult Failing Hearts. Journal of Cardiovascular Development and Disease, 1(1), 73-82. https://doi.org/10.3390/jcdd1010073