
Alginate Hydrogel Microtubes for Salivary Gland Cell Organization and Cavitation

 ,
,  ,
, 
Abstract
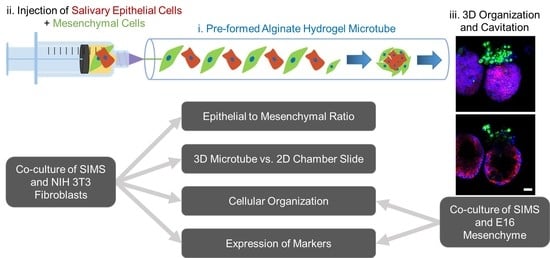
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Needle-in-Needle Microfluidic Device
2.3. Microfluidic Fabrication of Alginate Hydrogel Microtubes
2.4. Cell Culture
2.5. Isolation and Culture of Mouse Primary E16 Salivary Mesenchyme Cells
2.6. Cell Injection and Co-Culture in Pre-Formed Alginate Hydrogel Microtubes
2.7. Optical Imaging
2.8. Live and Dead Cell Assay
2.9. Immunocytochemistry Analysis
2.10. CellTracker™ Labeling of Mesenchymal and Epithelial Cells
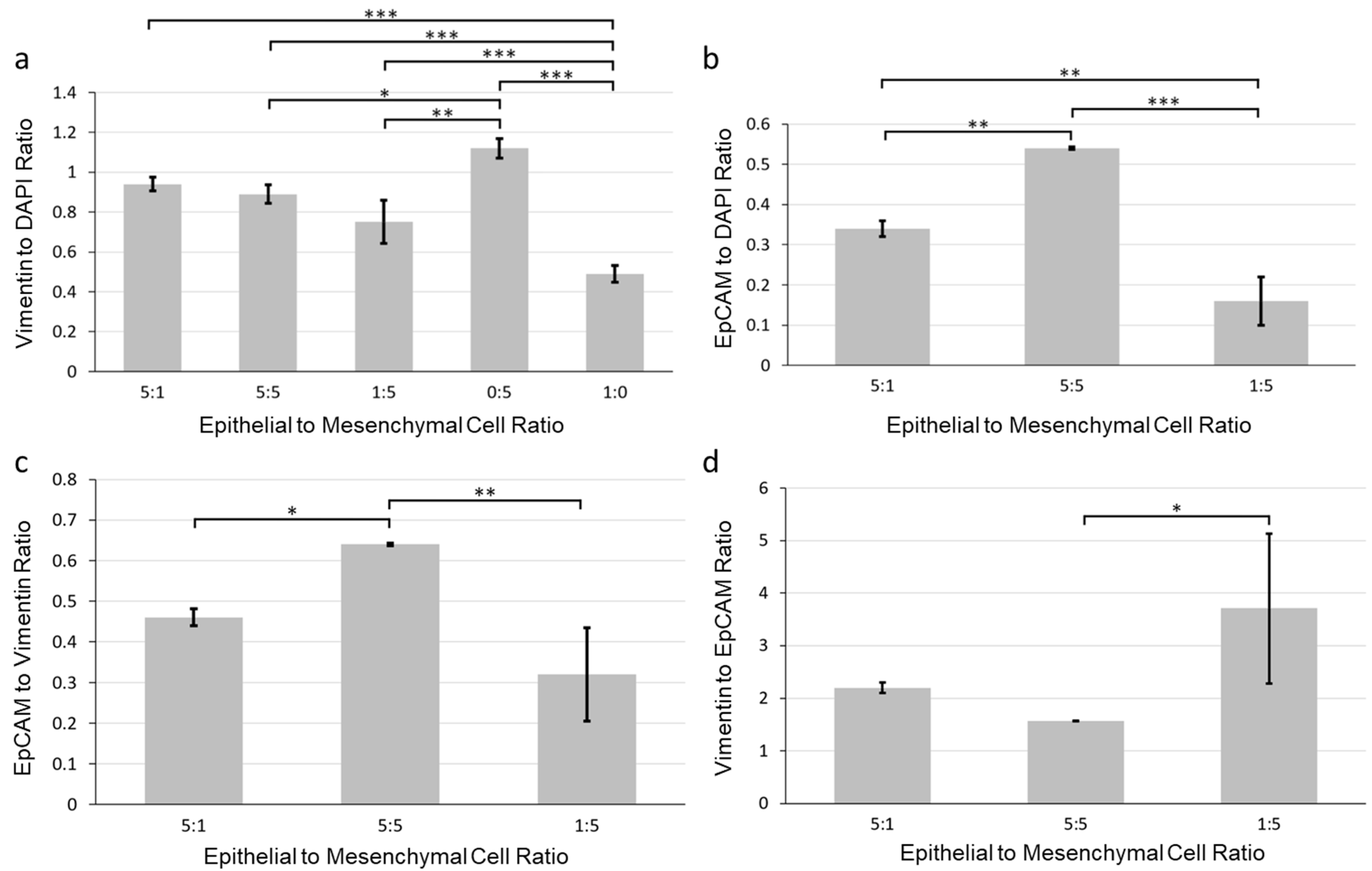
2.11. Image Quantification
2.12. Statistical Analysis
3. Results
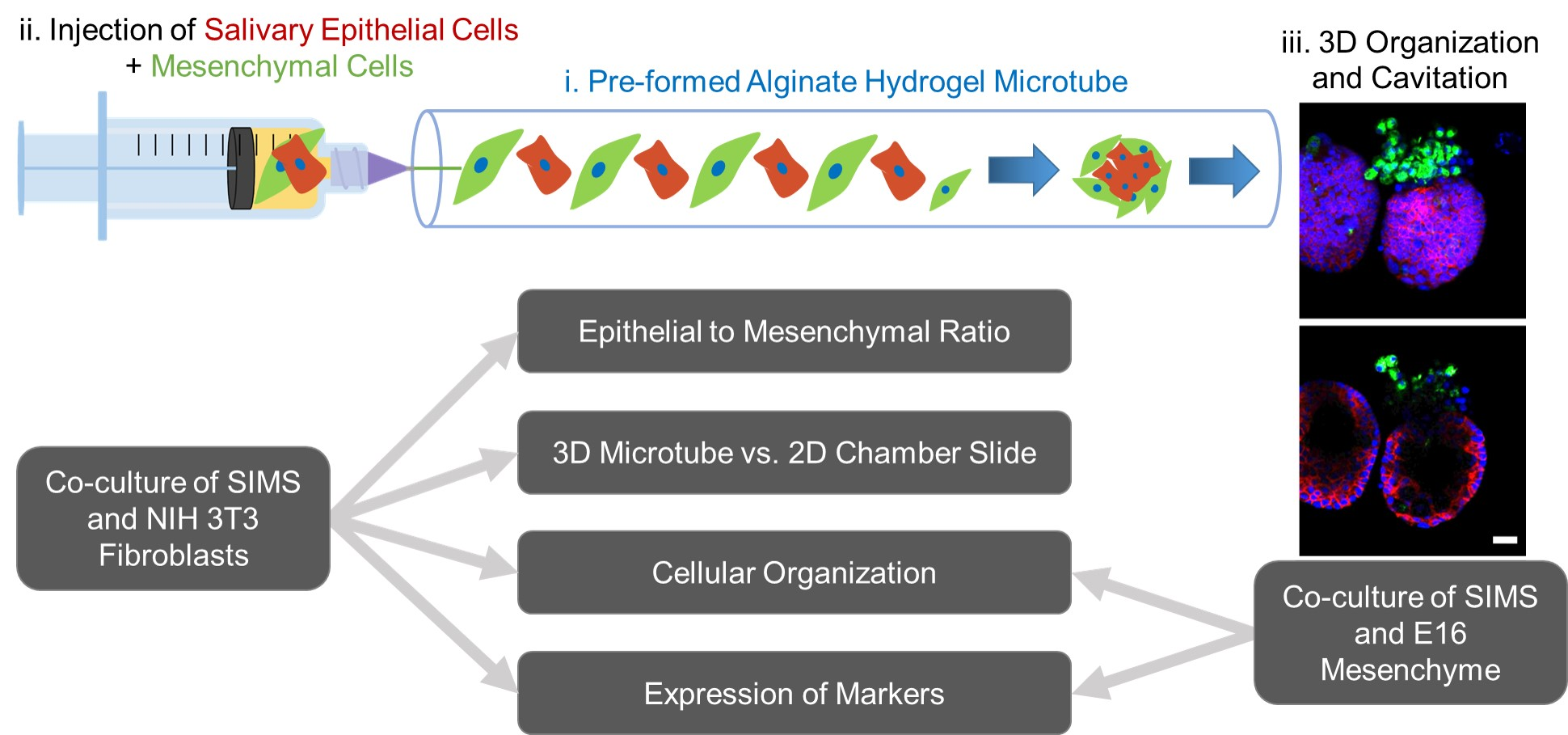
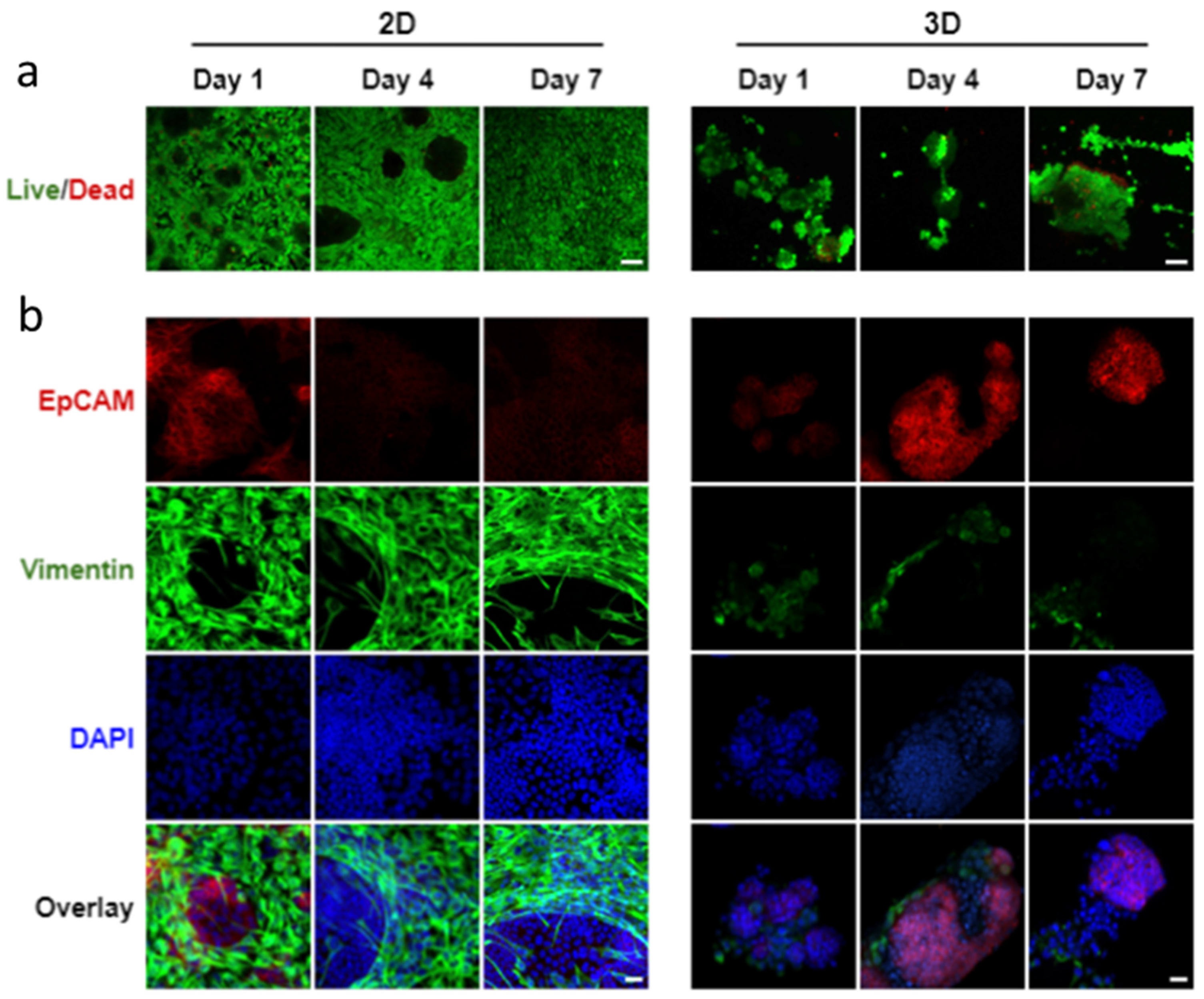
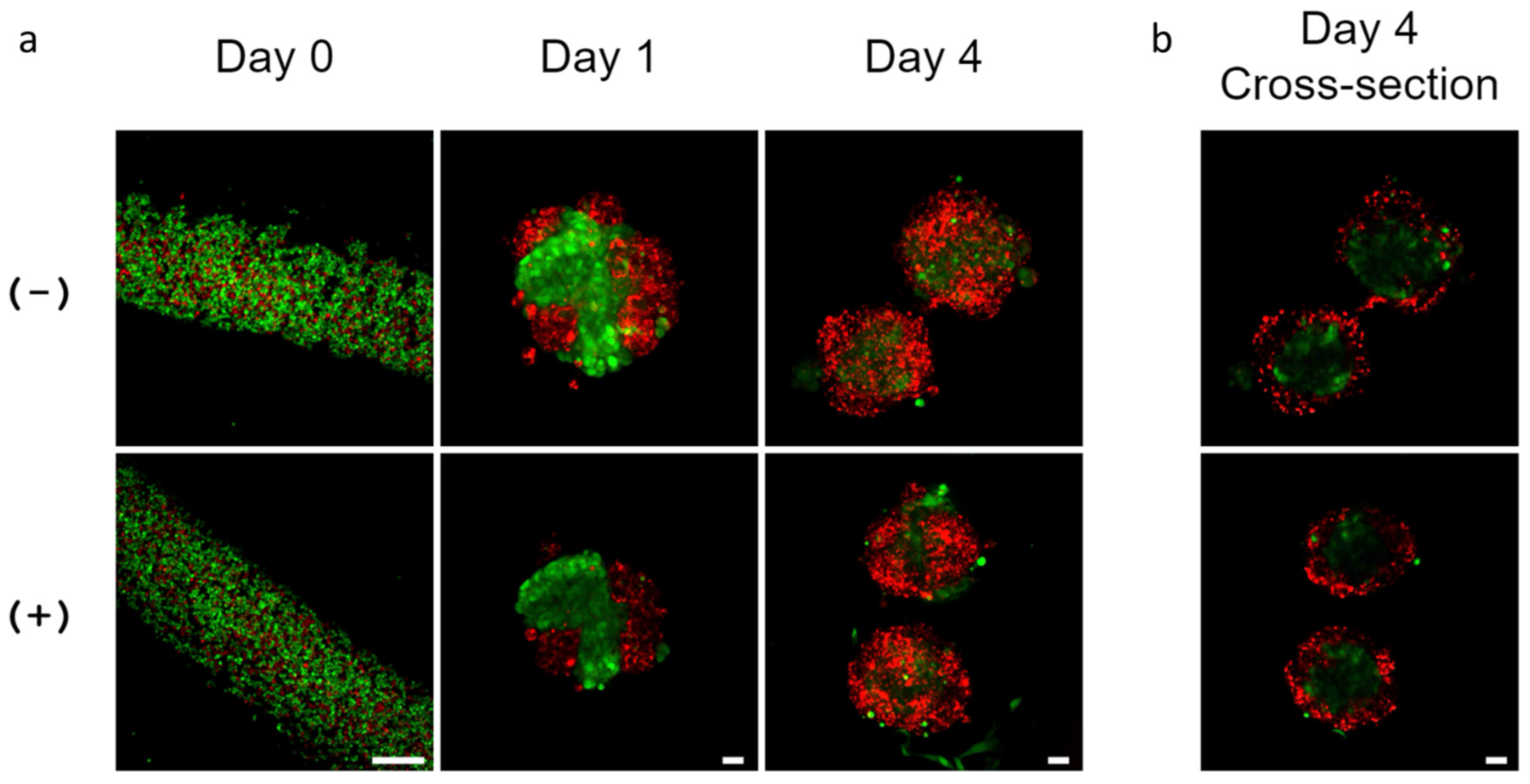
3.1. Reproducible Fabrication of Alginate Hydrogel Microtubes
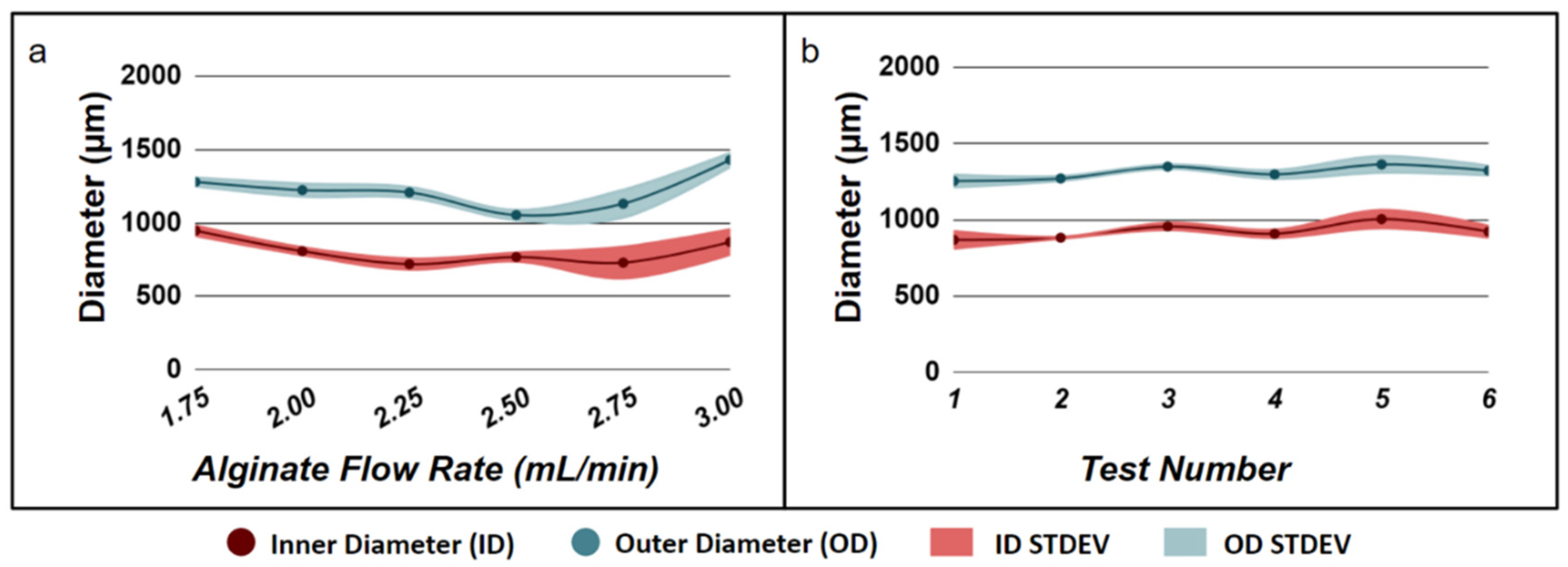
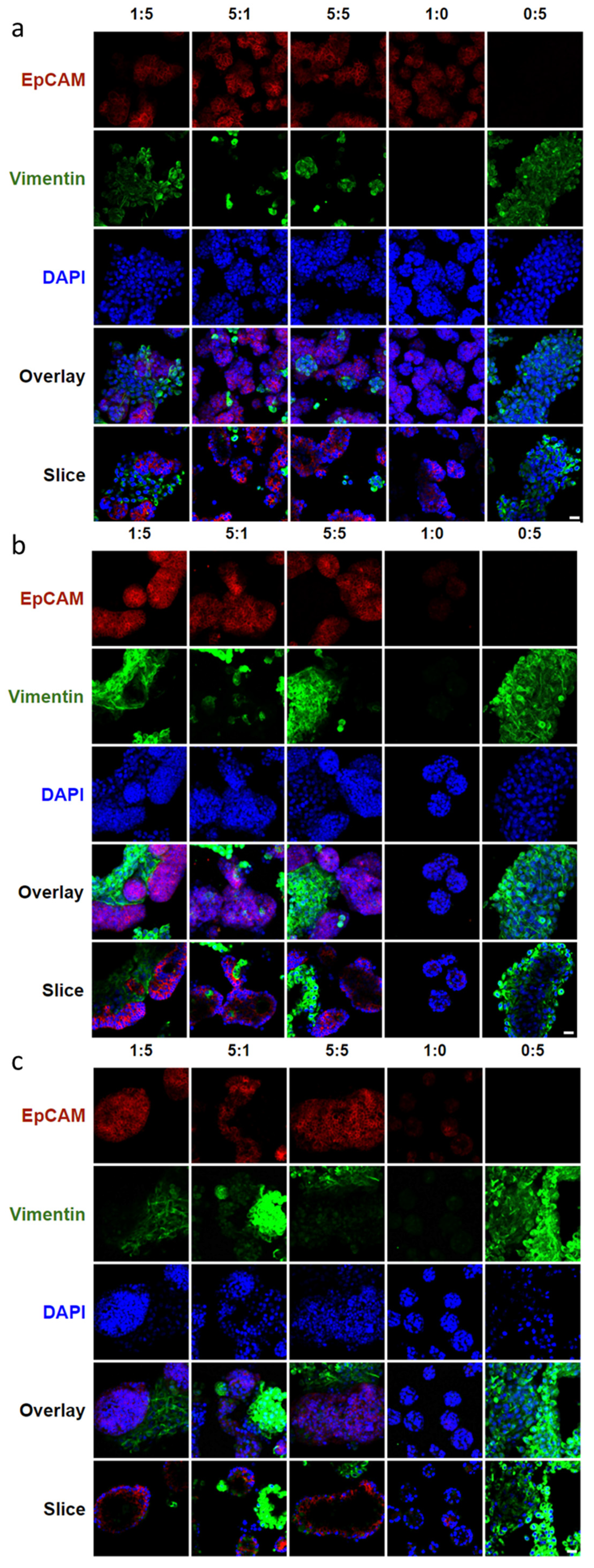
3.2. Effects of Epithelial to Mesenchymal Cell Ratio on Salivary Gland Cell Organization in Pre-Formed Alginate Hydrogel Microtubes
3.3. Facilitation of 3D Salivary Gland Cell Organization by Co-Culture of Epithelial SIMS Cells and NIH 3T3 Fibroblasts in Alginate Hydrogel Microtubes
3.4. Co-Culture of Salivary Gland Epithelial SIMS Cells and Primary E16 Mesenchyme Cells in Hydrogel Microtubes
4. Discussion
4.1. Pre-Formed Alginate Hydrogel Microtubes for Cell Injection and Cellular Organization
4.2. 3D Co-Culture of Salivary Gland Epithelial and Mesenchymal Cells for Cavitation in Alginate Hydrogel Microtubes
4.3. Future Strategies to Induce Branching Morphogenesis from Cavitated Structures Formed in Alginate Hydrogel Microtubes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Barrows, C.; Wu, D.; Farach-Carson, M.C.; Young, S. Building a functional salivary gland for cell-based therapy: More than secretory epithelial acini. Tissue Eng. Part A 2020, 26, 1332–1348. [Google Scholar] [CrossRef] [PubMed]
- Chansaenroj, A.; Yodmuang, S.; Ferreira, J.N. Trends in salivary gland tissue engineering: From stem cells to secretome and organoid bioprinting. Tissue Eng. Part B Rev. 2021, 27, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Jean-Gilles, R.; Soscia, D.; Sequeira, S.; Melfi, M.; Gadre, A.; Castracane, J.; Larsen, M. Novel modeling approach to generate a polymeric nanofiber scaffold for salivary gland cells. J. Nanotechnol. Eng. Med. 2010, 1, 31008. [Google Scholar] [CrossRef]
- Zhang, N.; Huang, G.; Han, Q.; Hu, X.; Yi, J.; Yao, L.; He, Y. Functional regeneration of irradiated salivary glands with human amniotic epithelial cells transplantation. Int. J. Clin. Exp. Pathol. 2013, 6, 2039–2047. [Google Scholar]
- Rocchi, C.; Emmerson, E. Mouth-watering results: Clinical need, current approaches, and future directions for salivary gland regeneration. Trends Mol. Med. 2020, 26, 649–669. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Laguna, L.; Sarkar, A. Aging-related changes in quantity and quality of saliva: Where do we stand in our understanding? J. Texture Stud. 2019, 50, 27–35. [Google Scholar] [CrossRef]
- Lombaert, I.; Movahednia, M.; Adine, C.; Ferreira, J. Concise review: Salivary gland regeneration: Therapeutic approaches from stem cells to tissue organoids. Stem Cells 2017, 35, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Maimets, M.; Rocchi, C.; Bron, R.; Pringle, S.; Kuipers, J.; Giepmans, B.; Vries, R.; Clevers, H.; de Haan, G.; van Os, R.; et al. Long-term in vitro expansion of salivary gland stem cells driven by Wnt signals. Stem Cell Rep. 2016, 6, 150–162. [Google Scholar] [CrossRef]
- Hosseini, Z.; Nelson, D.; Moskwa, N.; Sfakis, L.; Castracane, J.; Larsen, M. FGF2-dependent mesenchyme and laminin-111 are niche factors in salivary gland organoids. J. Cell Sci. 2018, 131, jcs208728. [Google Scholar] [CrossRef]
- Morrison, S.; Spradling, A. Stem cells and niches: Mechanisms that promote stem cell maintenance throughout life. Cell 2008, 132, 598–611. [Google Scholar] [CrossRef]
- Park, Y.; Koh, J.; Gauna, A.; Chen, S.; Cha, S. Identification of regulatory factors for mesenchymal stem cell-derived salivary epithelial cells in a co-culture system. PLoS ONE 2014, 9, e112158. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Lee, B.; Liao, C.; Cheng, W.; Chang, F.; Chen, M. Transdifferentiation of bone marrow stem cells into acinar cells using a double chamber system. J. Formos. Med. Assoc. 2007, 106, 1–7. [Google Scholar] [CrossRef][Green Version]
- Lin, C.; Yang, J.; Teng, S.; Tsai, S.; Chen, M. Microarray analysis of gene expression of bone marrow stem cells cocultured with salivary acinar cells. J. Formos. Med. Assoc. 2013, 112, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Maria, O.; Tran, S. Human mesenchymal stem cells cultured with salivary gland biopsies adopt an epithelial phenotype. Stem Cell Dev. 2011, 20, 959–967. [Google Scholar] [CrossRef]
- Nogawa, H.; Takahashi, Y. Substitution for mesenchyme by basement-membrane-like substratum and epidermal growth factor in inducing branching morphogenesis of mouse salivary epithelium. Development 1991, 112, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Farahat, M.; Sathi, G.; Hara, E.; Taketa, H.; Kuboki, T.; Matsumoto, T. MSCs feeder layers induce SMG self-organization and branching morphogenesis. PLoS ONE 2017, 12, e0176453. [Google Scholar] [CrossRef]
- Farahat, M.; Kazi, G.; Taketa, H.; Hara, E.; Oshima, M.; Kuboki, T.; Matsumoto, T. Fibronectin-induced ductal formation in salivary gland self-organization model. Dev. Dyn. 2019, 248, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, Z.F.; Nelson, D.A.; Moskwa, N.; Larsen, M. Generating embryonic salivary gland organoids. Curr. Protoc. Cell Biol. 2019, 83, e76. [Google Scholar] [CrossRef] [PubMed]
- Sfakis, L.; Kamaldinov, T.; Khmaladze, A.; Hosseini, Z.; Nelson, D.; Larsen, M.; Castracane, J. Mesenchymal cells affect salivary epithelial cell morphology on PGS/PLGA core/shell nanofibers. Int. J. Mol. Sci. 2018, 19, 1031. [Google Scholar] [CrossRef]
- Sequeira, S.; Soscia, D.; Oztan, B.; Mosier, A.; Jean-Gilles, R.; Gadre, A.; Cady, N.; Yener, B.; Castracane, J.; Larsen, M. The regulation of focal adhesion complex formation and salivary gland epithelial cell organization by nanofibrous PLGA scaffolds. Biomaterials 2012, 33, 3175–3186. [Google Scholar] [CrossRef]
- Peters, S.; Naim, N.; Nelson, D.; Mosier, A.; Cady, N.; Larsen, M. Biocompatible tissue scaffold compliance promotes salivary gland morphogenesis and differentiation. Tissue Eng. Part A 2014, 20, 1632–1642. [Google Scholar] [CrossRef]
- Foraida, Z.; Kamaldinov, T.; Nelson, D.; Larsen, M.; Castracane, J. Elastin-PLGA hybrid electrospun nanofiber scaffolds for salivary epithelial cell self-organization and polarization. Acta Biomater. 2017, 62, 116–127. [Google Scholar] [CrossRef]
- Soscia, D.; Sequeira, S.; Schramm, R.; Jayarathanam, K.; Cantara, S.; Larsen, M.; Castracane, J. Salivary gland cell differentiation and organization on micropatterned PLGA nanofiber craters. Biomaterials 2013, 34, 6773–6784. [Google Scholar] [CrossRef] [PubMed]
- Augst, A.; Kong, H.; Mooney, D. Alginate hydrogels as biomaterials. Macromol. Biosci. 2006, 6, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Mooney, D. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Khademhosseini, A. Advances in engineering hydrogels. Science 2017, 356, eaaf3627. [Google Scholar] [CrossRef] [PubMed]
- Namgung, B.; Ravi, K.; Vikraman, P.P.; Sengupta, S.; Jang, H.L. Engineered cell-laden alginate microparticles for 3D culture. Biochem. Soc. Trans. 2021, 49, 761–773. [Google Scholar] [CrossRef]
- Xu, M.; Qin, M.; Cheng, Y.; Niu, X.; Kong, J.; Zhang, X.; Huang, D.; Wang, H. Alginate microgels as delivery vehicles for cell-based therapies in tissue engineering and regenerative medicine. Carbohydr. Polym. 2021, 266, 118128. [Google Scholar] [CrossRef]
- Yamada, M.; Utoh, R.; Ohashi, K.; Tatsumi, K.; Yamato, M.; Okano, T.; Seki, M. Controlled formation of heterotypic hepatic micro-organoids in anisotropic hydrogel microfibers for long-term preservation of liver-specific functions. Biomaterials 2012, 33, 8304–8315. [Google Scholar] [CrossRef]
- Onoe, H.; Okitsu, T.; Itou, A.; Kato-Negishi, M.; Gojo, R.; Kiriya, D.; Sato, K.; Miura, S.; Iwanaga, S.; Kuribayashi-Shigetomi, K.; et al. Metre-long cell-laden microfibres exhibit tissue morphologies and functions. Nat. Mater. 2013, 12, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Onoe, H.; Kato-Negishi, M.; Itou, A.; Takeuchi, S. Differentiation induction of mouse neural stem cells in hydrogel tubular microenvironments with controlled tube dimensions. Adv. Healthc. Mater. 2016, 5, 1104–1111. [Google Scholar] [CrossRef]
- Li, Q.; Lin, H.; Du, Q.; Liu, K.; Wang, O.; Evans, C.; Christian, H.; Zhang, C.; Lei, Y. Scalable and physiologically relevant microenvironments for human pluripotent stem cell expansion and differentiation. Biofabrication 2018, 10, 025006. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Du, Q.; Li, Q.; Wang, O.; Wang, Z.; Liu, K.; Elowsky, C.; Zhang, C.; Lei, Y. Hydrogel-based bioprocess for scalable manufacturing of human pluripotent stem cell-derived neural stem cells. ACS Appl. Mater. Interfaces 2018, 10, 29238–29250. [Google Scholar] [CrossRef]
- Jorgensen, M.; Gibbons, A.; Sui, K.; Carpenter, R.; Zhang, X.; Xie, Y. Predictable fabrication of pre-made alginate hydrogel microtubes for stem cell aggregation using needle-in-needle devices. Biofabrication 2021, 13, 035043. [Google Scholar] [CrossRef]
- Liu, R.; Kong, B.; Chen, Y.; Liu, X.; Mi, S. Formation of helical alginate microfibers using different G/M ratios of sodium alginate based on microfluidics. Sens. Actuators B Chem. 2020, 304, 127069. [Google Scholar] [CrossRef]
- Jainchill, J.L.; Aaronson, S.A.; Todaro, G.J. Murine sarcoma and leukemia viruses: Assay using clonal lines of contact-inhibited mouse cells. J. Virol. 1969, 4, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Laoide, B.; Courty, Y.; Gastinne, I.; Thibaut, C.; Kellermann, O.; Rougeon, F. Immortalised mouse submandibular epithelial cell lines retain polarised structural and functional properties. J. Cell Sci. 1996, 109 Pt 12, 2789–2800. [Google Scholar] [CrossRef]
- Koslow, M.; O’Keefe, K.J.; Hosseini, Z.F.; Nelson, D.A.; Larsen, M. ROCK inhibitor increases proacinar cells in adult salivary gland organoids. Stem Cell Res. 2019, 41, 101608. [Google Scholar] [CrossRef] [PubMed]
- Moskwa, N.; Mahmood, A.; Nelson, D.A.; Altrieth, A.L.; Forni, P.; Larsen, M. PDFGRα+ stromal cells promote salivary gland proacinar differentiation through FGF2-dependent BMP7 signaling. bioRxiv 2021. [Google Scholar] [CrossRef]
- Rouwkema, J.; Koopman, B.; Blitterswijk, C.; Dhert, W.; Malda, J. Supply of nutrients to cells in engineered tissues. Biotechnol. Genet. Eng. Rev. 2010, 26, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.; Wei, C.; Yamada, K. Cell and fibronectin dynamics during branching morphogenesis. J. Cell Sci. 2006, 119 Pt 16, 3376–3384. [Google Scholar] [CrossRef]
- Hirayama, K.; Okitsu, T.; Teramae, H.; Kiriya, D.; Onoe, H.; Takeuchi, S. Cellular building unit integrated with microstrand-shaped bacterial cellulose. Biomaterials 2013, 34, 2421–2427. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; He, X.; Yang, Y.; Wei, D.; Sun, J.; Zhong, M.; Xie, R.; Fan, H.; Zhang, X. Microfluidic-based generation of functional microfibers for biomimetic complex tissue construction. Acta Biomater. 2016, 38, 153–162. [Google Scholar] [CrossRef]
- Fujimoto, K.; Higashi, K.; Onoe, H.; Miki, N. Development of a triple-coaxial flow device for fabricating a hydrogel microtube and its application to bioremediation. Micromachines 2018, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Liang, Z.; Ai, Y.; Zheng, W.; Xiong, J.; Xu, P.; Liu, Y.; Ding, M.; Gao, J.; Wang, J.; et al. Composable microfluidic spinning platforms for facile production of biomimetic perfusable hydrogel microtubes. Nat. Protoc. 2021, 16, 937–964. [Google Scholar] [CrossRef]
- Bandyopadhyay, B.C.; Swaim, W.D.; Sarkar, A.; Liu, X.; Ambudkar, I.S. Extracellular Ca(2+) sensing in salivary ductal cells. J. Biol. Chem. 2012, 287, 30305–30316. [Google Scholar] [CrossRef]
- Fox, R.I.; Fox, C.M. Therapy of oral and cutaneous dryness manifestations in Sjogren’s syndrome. In Sjögren’s Syndrome Diagnosis and Therapeutics; Casals, M.R., Stone, J.H., Moutsopouls, H.M., Eds.; Springer: New York, NY, USA, 2012; p. 522. [Google Scholar]
- Haug, A.; Smidsrod, O.A.; Larsen, B.; Gronowitz, S.; Hoffman, R.A.; Westerdahl, A. The effect of divalent metals on the properties of alginate solutions. II. Comparison of different metal ions. Acta Chem. Scand. 1965, 19, 341–351. [Google Scholar] [CrossRef]
- Thiery, J.P.; Sleeman, J.P. Complex networks orchestrate epithelial-mesenchymal transitions. Nat. Rev. Mol. Cell Biol. 2006, 7, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.; Yamada, K. Salivary gland branching morphogenesis--recent progress and future opportunities. Int. J. Oral Sci. 2010, 2, 117–126. [Google Scholar] [CrossRef]
- Janebodin, K.; Buranaphatthana, W.; Ieronimakis, N.; Hays, A.L.; Reyes, M. An in vitro culture system for long-term expansion of epithelial and mesenchymal salivary gland cells: Role of TGF-β1 in salivary gland epithelial and mesenchymal differentiation. BioMed Res. Int. 2013, 2013, 815895. [Google Scholar] [CrossRef]
- Kawakami, M.; Ishikawa, H.; Tachibana, T.; Tanaka, A.; Mataga, I. Functional transplantation of salivary gland cells differentiated from mouse early ES cells in vitro. Hum. Cell 2013, 26, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Maria, O.M.; Zeitouni, A.; Gologan, O.; Tran, S.D. Matrigel improves functional properties of primary human salivary gland cells. Tissue Eng. Part A 2011, 17, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Schnell, U.; Cirulli, V.; Giepmans, B. EpCAM: Structure and function in health and disease. Biochim. Biophys. Acta Biomembr. 2013, 1828, 1989–2001. [Google Scholar] [CrossRef] [PubMed]
- Hai, B.; Yang, Z.; Millar, S.E.; Choi, Y.S.; Taketo, M.M.; Nagy, A.; Liu, F. Wnt/β-catenin signaling regulates postnatal development and regeneration of the salivary gland. Stem Cells Dev. 2010, 19, 1793–1801. [Google Scholar] [CrossRef]
- Wend, P.; Fang, L.; Zhu, Q.; Schipper, J.H.; Loddenkemper, C.; Kosel, F.; Brinkmann, V.; Eckert, K.; Hindersin, S.; Holland, J.D.; et al. Wnt/beta-catenin signaling induces MLL to create epigenetic changes in salivary gland tumours. EMBO J. 2013, 32, 1977–1989. [Google Scholar] [CrossRef] [PubMed]
- Ono, H.; Obana, A.; Usami, Y.; Sakai, M.; Nohara, K.; Egusa, H.; Sakai, T. Regenerating salivary glands in the microenvironment of induced pluripotent stem cells. BioMed Res. Int. 2015, 2015, 293570. [Google Scholar] [CrossRef]
- Lim, J.-Y.; Ra, J.C.; Shin, S.; Jang, Y.H.; An, H.-Y.; Choi, J.-S.; Kim, W.C.; Kim, Y.-M. Systemic transplantation of human adipose tissue-derived mesenchymal stem cells for the regeneration of irradiation-induced salivary gland damage. PLoS ONE 2013, 8, e71167. [Google Scholar] [CrossRef]
- Andrew, D.J.; Ewald, A.J. Morphogenesis of epithelial tubes: Insights into tube formation, elongation, and elaboration. Dev. Biol. 2010, 341, 34–55. [Google Scholar] [CrossRef]
- Borghese, E. The development in vitro of the submandibular and sublingual glands of Mus musculus. J. Anat. 1950, 84, 287–302. [Google Scholar]
- Pradhan-Bhatt, S.; Harrington, D.A.; Duncan, R.L.; Jia, X.; Witt, R.L.; Farach-Carson, M.C. Implantable three-dimensional salivary spheroid assemblies demonstrate fluid and protein secretory responses to neurotransmitters. Tissue Eng. Part A 2013, 19, 1610–1620. [Google Scholar] [CrossRef]
- Srinivasan, P.P.; Patel, V.N.; Liu, S.; Harrington, D.A.; Hoffman, M.P.; Jia, X.; Witt, R.L.; Farach-Carson, M.C.; Pradhan-Bhatt, S. Primary salivary human stem/progenitor cells undergo microenvironment-driven acinar-like differentiation in hyaluronate hydrogel culture. Stem Cells Trans. Med. 2017, 6, 110–120. [Google Scholar] [CrossRef]
- Tanaka, J.; Ogawa, M.; Hojo, H.; Kawashima, Y.; Mabuchi, Y.; Hata, K.; Nakamura, S.; Yasuhara, R.; Takamatsu, K.; Irié, T.; et al. Generation of orthotopically functional salivary gland from embryonic stem cells. Nat. Commun. 2018, 9, 4216. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Larsen, M.; Hoffman, M.; Yamada, K. Self-organization and branching morphogenesis of primary salivary epithelial cells. Tissue Eng. 2007, 13, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Joraku, A.; Sullivan, C.A.; Yoo, J.; Atala, A. In-vitro reconstitution of three-dimensional human salivary gland tissue structures. Differentiation 2007, 75, 318–324. [Google Scholar] [CrossRef]
- Sui, Y.; Zhang, S.; Li, Y.; Zhang, X.; Hu, W.; Feng, Y.; Xiong, J.; Zhang, Y.; Wei, S. Generation of functional salivary gland tissue from human submandibular gland stem/progenitor cells. Stem Cell Res. Ther. 2020, 11, 127. [Google Scholar] [CrossRef]
- Feng, J.; van der Zwaag, M.; Stokman, M.A.; van Os, R.; Coppes, R.P. Isolation and characterization of human salivary gland cells for stem cell transplantation to reduce radiation-induced hyposalivation. Radiother. Oncol. 2009, 92, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Nanduri, L.S.; Baanstra, M.; Faber, H.; Rocchi, C.; Zwart, E.; de Haan, G.; van Os, R.; Coppes, R.P. Purification and ex vivo expansion of fully functional salivary gland stem cells. Stem Cell Rep. 2014, 3, 957–964. [Google Scholar] [CrossRef]
- Pringle, S.; Maimets, M.; van der Zwaag, M.; Stokman, M.; van Gosliga, D.; Zwart, E.; Witjes, M.; de Haan, G.; van Os, R.; Coppes, R. Human salivary gland stem cells functionally restore radiation damaged salivary glands. Stem Cells 2016, 34, 640–652. [Google Scholar] [CrossRef]
- Wells, K.; Gaete, M.; Matalova, E.; Deutsch, D.; Rice, D.; Tucker, A. Dynamic relationship of the epithelium and mesenchyme during salivary gland initiation: The role of Fgf10. Biol. Open 2014, 3, 677. [Google Scholar] [CrossRef]
- Häärä, O.; Fujimori, S.; Schmidt-Ullrich, R.; Hartmann, C.; Thesleff, I.; Mikkola, M. Ectodysplasin and Wnt pathways are required for salivary gland branching morphogenesis. Development 2011, 138, 2681–2691. [Google Scholar] [CrossRef]
- Kadoya, Y.; Kadoya, K.; Durbeej, M.; Holmvall, K.; Sorokin, L.; Ekblom, P. Antibodies against domain E3 of laminin-1 and integrin alpha 6 subunit perturb branching epithelial morphogenesis of submandibular gland, but by different modes. J. Cell Biol. 1995, 129, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Cantara, S.; Soscia, D.; Sequeira, S.; Jean-Gilles, R.; Castracane, J.; Larsen, M. Selective functionalization of nanofiber scaffolds to regulate salivary gland epithelial cell proliferation and polarity. Biomaterials 2012, 33, 8372–8382. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Sakai, T.; Larsen, M. Fibronectin requirement in branching morphogenesis. Nature 2003, 423, 876–881. [Google Scholar] [CrossRef]
- Onodera, T.; Sakai, T.; Hsu, J.C.; Matsumoto, K.; Chiorini, J.A.; Yamada, K.M. Btbd7 regulates epithelial cell dynamics and branching morphogenesis. Science 2010, 329, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Rebustini, I.T.; Hoffman, M.P. ECM and FGF-dependent assay of embryonic SMG epithelial morphogenesis: Investigating growth factor/matrix regulation of gene expression during submandibular gland development. Methods Mol. Biol. 2009, 522, 319–330. [Google Scholar] [CrossRef]
- Samuel, R.; Lei, P.; Nam, K.; Baker, O.; Andreadis, S. Engineering the mode of morphogenetic signal presentation to promote branching from salivary gland spheroids in 3D hydrogels. Acta Biomater. 2020, 105, 121–130. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jorgensen, M.; Ramesh, P.; Toro, M.; Evans, E.; Moskwa, N.; Zhang, X.; Sharfstein, S.T.; Larsen, M.; Xie, Y. Alginate Hydrogel Microtubes for Salivary Gland Cell Organization and Cavitation. Bioengineering 2022, 9, 38. https://doi.org/10.3390/bioengineering9010038
Jorgensen M, Ramesh P, Toro M, Evans E, Moskwa N, Zhang X, Sharfstein ST, Larsen M, Xie Y. Alginate Hydrogel Microtubes for Salivary Gland Cell Organization and Cavitation. Bioengineering. 2022; 9(1):38. https://doi.org/10.3390/bioengineering9010038
Chicago/Turabian StyleJorgensen, Matthew, Pujhitha Ramesh, Miriam Toro, Emily Evans, Nicholas Moskwa, Xulang Zhang, Susan T. Sharfstein, Melinda Larsen, and Yubing Xie. 2022. "Alginate Hydrogel Microtubes for Salivary Gland Cell Organization and Cavitation" Bioengineering 9, no. 1: 38. https://doi.org/10.3390/bioengineering9010038
APA StyleJorgensen, M., Ramesh, P., Toro, M., Evans, E., Moskwa, N., Zhang, X., Sharfstein, S. T., Larsen, M., & Xie, Y. (2022). Alginate Hydrogel Microtubes for Salivary Gland Cell Organization and Cavitation. Bioengineering, 9(1), 38. https://doi.org/10.3390/bioengineering9010038