
Recent Advances and Challenges towards Sustainable Polyhydroxyalkanoate (PHA) Production

 , , , and
, , , and 
Abstract
:1. Introduction
2. PHA Production by Pure Bacterial Cultures
2.1. Lignocellulose Degraders
2.1.1. Saccharophagus Degradans
2.1.2. Caldimonas Taiwanensis
2.2. Bioremediation Technologies Allowing PHA Production
2.3. Halophiles
2.4. Synthetic Biology of PHA Producers
3. PHA Production by Mixed Microbial Consortia (MMC)
3.1. Types of Enrichments
3.1.1. Anaerobic/Aerobic Enrichments (AN/AE)
3.1.2. Aerobic Dynamic Feeding (ADF)
3.1.3. Variations of the ADF Enrichments
3.1.4. ADF Enrichments in Halophilic Conditions
3.1.5. Mixed Photosynthetic Consortia
3.1.6. Aerobic–Anoxic Enrichment Coupled with Nitrification/Denitrification
3.1.7. Anoxic–Aerobic Strategy Coupled with Nitrification/Denitrification
3.1.8. Microaerophilic Conditions
3.1.9. PHA Accumulation without Previous Enrichment
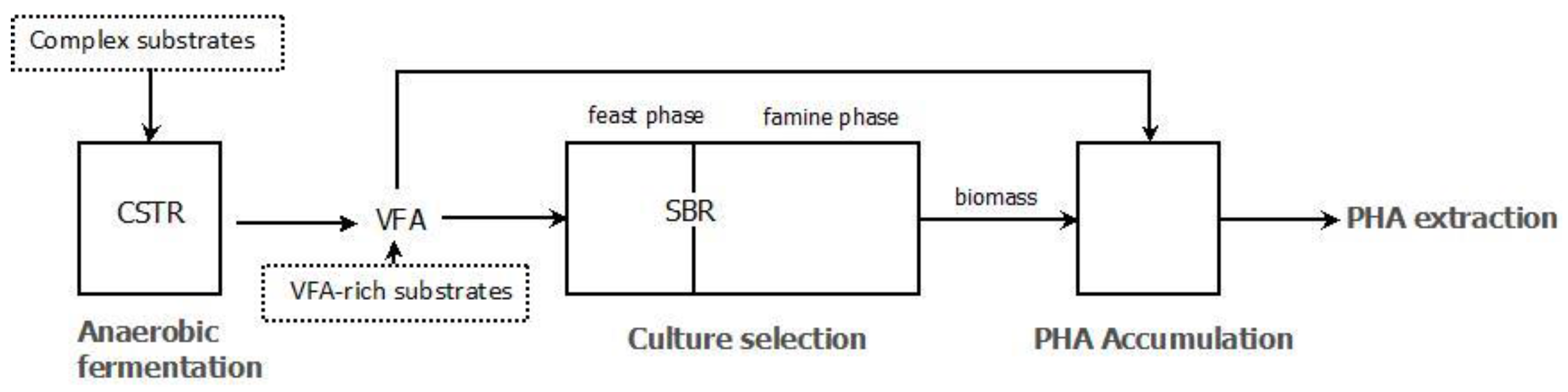
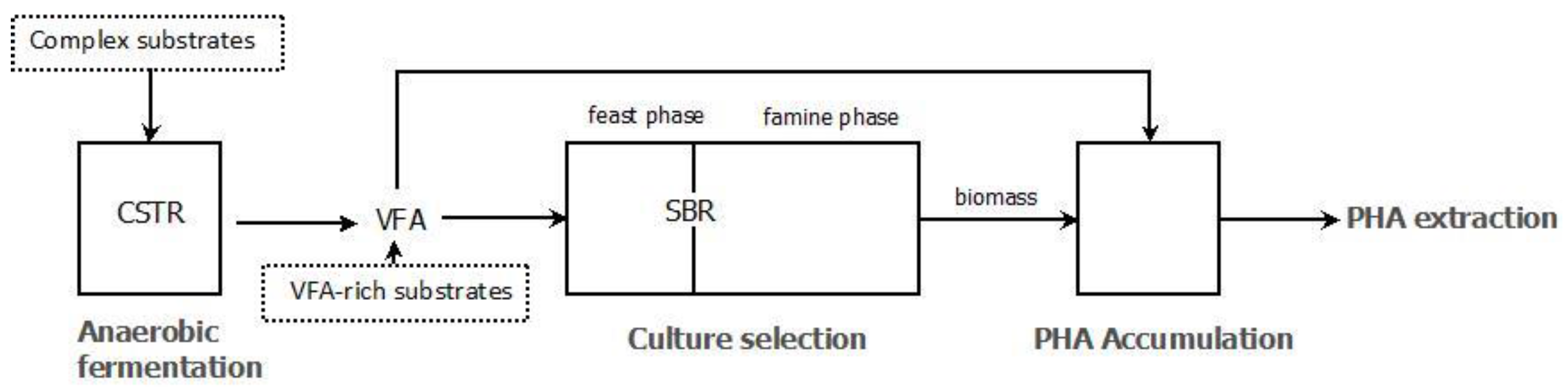
3.2. PHA Accumulation
3.3. Pilot Scale Experiences
3.4. Challenges and Perspectives Regarding PHA Production by Mixed Microbial Consortia
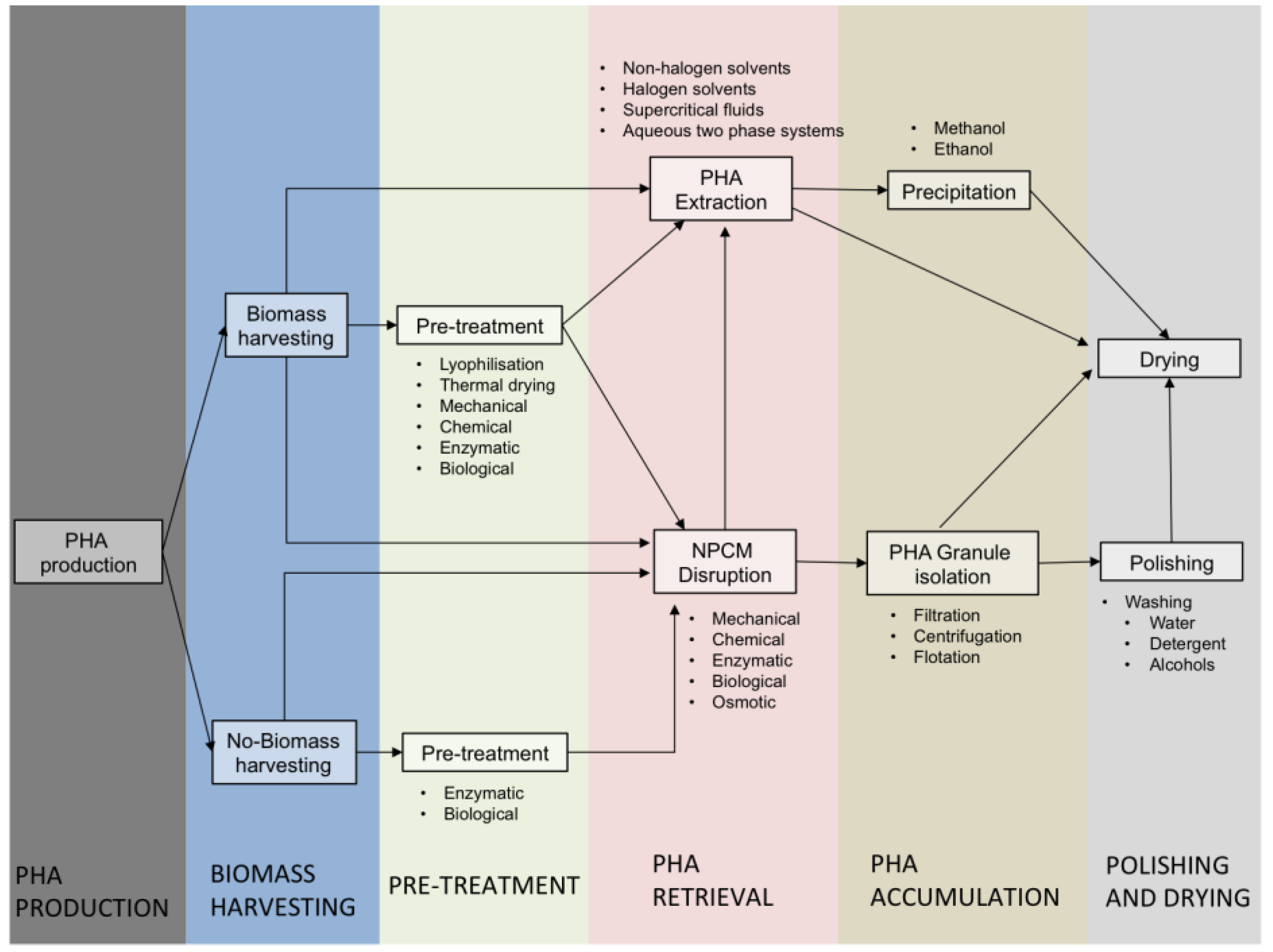
4. PHA Recovery
4.1. Pre-Treatments
4.2. Retrieval Techniques
4.2.1. Non-PHA Cell Mass (NPCM) Disruption
Chemical Disruption
Enzymatic NPCM Disruption
Biological NPCM Disruption
4.2.2. PHA Extraction
Non-Halogenated Solvents
Aqueous Two-Phase Extraction Systems (ATPS)
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Plastics Europe Plastics—The Facts 2016; Plastics Europe: Brussels, Belgium, 2016; pp. 1–38.
- Kourmentza, C.; Kornaros, M. Biotransformation of volatile fatty acids to polyhydroxyalkanoates by employing mixed microbial consortia: The effect of pH and carbon source. Bioresour. Technol. 2016, 222, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Singh Saharan, B.; Grewal, A.; Kumar, P. Biotechnological Production of Polyhydroxyalkanoates: A Review on Trends and Latest Developments. Chin. J. Biol. 2014, 2014, 1–18. [Google Scholar] [CrossRef]
- Chen, G.-Q. A microbial polyhydroxyalkanoates (PHA) based bio- and materials industry. Chem. Soc. Rev. 2009, 38, 2434–2446. [Google Scholar] [CrossRef] [PubMed]
- López, N.I.; Pettinari, M.J.; Nikel, P.I.; Méndez, B.S. Polyhydroxyalkanoates: Much More than Biodegradable Plastics. Adv. Appl. Microbiol. 2015, 93, 73–106. [Google Scholar] [PubMed]
- Kourmentza, C.; Koutra, E.; Venetsaneas, N.; Kornaros, M. Integrated Biorefinery Approach for the Valorization of Olive Mill Waste Streams Towards Sustainable Biofuels and Bio-Based Products. In Microbial Appications Vol. 1—Bioremediation and Bioenergy; Kalia, V.C., Kumar, P., Eds.; Springer: Berlin, Germany, 2017; Volume 1, pp. 211–238. [Google Scholar]
- Plastics Technology. Available online: http://www.ptonline.com/articles/prices-bottom-out-for-polyolefins-pet-ps-pvc-move-up (accessed on 26 May 2017).
- Eno, R.; Hill, J. Metabolix Bio-industrial Evolution. In Proceedings of the Jefferies 11th Global Clean Technology Conference, New York, NY, USA, 23–24 February 2011. [Google Scholar]
- Markets and Markets Polyhydroxyalkanoate (PHA) Market by Type (Monomers, Co-Polymers, Terpolymers), Manufacturing Technology (Bacterial Fermentation, Biosynthesis, Enzymatic Catalysis), Application (Packaging, Bio Medical, Food Services, Agriculture)-Global Forecast to 202. Available online: http://www.marketsandmarkets.com/Market-Reports/pha-market-395.html (accessed on 26 May 2017).
- Kourmentza, C.; Ntaikou, I.; Lyberatos, G.; Kornaros, M. Polyhydroxyalkanoates from Pseudomonas sp. using synthetic and olive mill wastewater under limiting conditions. Int. J. Biol. Macromol. 2015, 74, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Fraiberg, M.; Borovok, I.; Weiner, R.M.; Lamed, R. Discovery and characterization of cadherin domains in Saccharophagus degradans 2-40. J. Bacteriol. 2010, 192, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Weiner, R.M.; Taylor, L.E.; Henrissat, B.; Hauser, L.; Land, M.; Coutinho, P.M.; Rancurel, C.; Saunders, E.H.; Longmire, A.G.; Zhang, H.; et al. Complete genome sequence of the complex carbohydrate-degrading marine bacterium, Saccharophagus degradans strain 2-40T. PLoS Genet. 2008, 4, e1000087. [Google Scholar] [CrossRef] [PubMed]
- Ekborg, N.A.; Taylor, L.E.; Longmire, A.G.; Henrissat, B.; Weiner, R.M.; Steven, W.; Hutcheson, S.W. Genomic and Proteomic Analyses of the Agarolytic System Expressed by Saccharophagus degradans 2-40. Appl. Environ. Microbiol. 2006, 72, 3396–3405. [Google Scholar] [CrossRef] [PubMed]
- Andrykovitch, G.; Marx, I. Isolation of a new polysaccharide-digesting bacterium from a salt marsh. Appl. Environ. Microbiol. 1988, 54, 1061–1062. [Google Scholar] [PubMed]
- Taylor, L.E.; Henrissat, B.; Coutinho, P.M.; Ekborg, N.A.; Hutcheson, S.W.; Weiner, R.M. Complete cellulase system in the marine bacterium Saccharophagus degradans strain 2-40T. J. Bacteriol. 2006, 188, 3849–3861. [Google Scholar] [CrossRef] [PubMed]
- Howard, M.B.; Ekborg, N.A.; Taylor, L.E.; Weiner, R.M.; Hutcheson, S.W. Genomic Analysis and Initial Characterization of the Chitinolytic System of Microbulbifer degradans Strain 2-40. J. Bacteriol 2003, 185, 3352–3360. [Google Scholar] [CrossRef] [PubMed]
- Howard, M.B.; Ekborg, N.A.; Taylor, L.E.; Weiner, R.M.; Hutcheson, S.W. Chitinase B of “Microbulbifer degradans” 2-40 Contains Two Catalytic Domains with Different Chitinolytic Activities. J. Bacteriol. 2004, 186, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Suvorov, M.; Kumar, R.; Zhang, H.; Hutcheson, S. Novelties of the cellulolytic system of a marine bacterium applicable to cellulosic sugar production. Biofuels 2011, 2, 59–70. [Google Scholar] [CrossRef]
- Munoz, L.E.A.; Riley, M.R. Utilization of cellulosic waste from tequila bagasse and production of polyhydroxyalkanoate (pha) bioplastics by Saccharophagus degradans. Biotechnol. Bioeng. 2008, 100, 882–888. [Google Scholar] [CrossRef] [PubMed]
- González-García, Y.; Nungaray, J.; Córdova, J.; González-Reynoso, O.; Koller, M.; Atlic, A.; Braunegg, G. Biosynthesis and characterization of polyhydroxyalkanoates in the polysaccharide-degrading marine bacterium Saccharophagus degradans ATCC 43961. J. Ind. Microbiol. Biotechnol. 2008, 35, 629–633. [Google Scholar] [CrossRef] [PubMed]
- González-García, Y.; Rosales, M.A.; González-Reynoso, O.; Sanjuán-Dueñas, R.; Córdova, J. Polyhydroxybutyrate production by Saccharophagus degradans using raw starch as carbon source. Eng. Life Sci. 2011, 11, 59–64. [Google Scholar] [CrossRef]
- Sheu, D.S.; Chen, W.M.; Yang, J.Y.; Chang, R.C. Thermophilic bacterium Caldimonas taiwanensis produces poly(3-hydroxybutyrate-co-3-hydroxyvalerate) from starch and valerate as carbon sources. Enzyme Microb. Technol. 2009, 44, 289–294. [Google Scholar] [CrossRef]
- Sawant, S.S.; Tran, T.K.; Salunke, B.K.; Kim, B.S. Potential of Saccharophagus degradans for production of polyhydroxyalkanoates using cellulose. Process Biochem. 2017. [Google Scholar] [CrossRef]
- Sawant, S.S.; Salunke, B.K.; Taylor, L.E.; Kim, B.S. Enhanced agarose and xylan degradation for production of polyhydroxyalkanoates by co-culture of marine bacterium, Saccharophagus degradans and its contaminant, Bacillus cereus. Appl. Sci. 2017, 7, 225. [Google Scholar] [CrossRef]
- Sawant, S.S.; Salunke, B.K.; Kim, B.S. A Laboratory Case Study of Efficient Polyhydoxyalkonates Production by Bacillus cereus, a Contaminant in Saccharophagus degradans ATCC 43961 in Minimal Sea Salt Media. Curr. Microbiol. 2014, 69, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Salamanca-Cardona, L.; Ashe, C.S.; Stipanovic, A.J.; Nomura, C.T. Enhanced production of polyhydroxyalkanoates (PHAs) from beechwood xylan by recombinant Escherichia coli. Appl. Microbiol. Biotechnol. 2014, 98, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.M.; Chang, J.S.; Chiu, C.H.; Chang, S.C.; Chen, W.C.; Jiang, C.M. Caldimonas taiwanensis sp. nov., a amylase producing bacterium isolated from a hot spring. Syst. Appl. Microbiol. 2005, 28, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Elleuche, S.; Antranikian, G. Starch-Hydrolyzing Enzymes from Thermophiles. In Thermophilic Microbes in Environmental and Industrial Biotechnology: Biotechnology of Thermophiles; Satyanarayana, T., Littlechild, J., Kawarabayasi, Y., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 509–533. [Google Scholar]
- Ward, P.G.; De Roo, G.; O’Connor, K.E. Accumulation of polyhydroxyalkanoate from styrene and phenylacetic acid by Pseudomonas putida CA-3. Appl. Environ. Microbiol. 2005, 71, 2046–2052. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Moore, C.J.; Vom Saal, F.S.; Swan, S.H. Plastics, the environment and human health: Current consensus and future trends. Philos. Trans. R. Soc. B 2009, 364, 2153–2166. [Google Scholar] [CrossRef] [PubMed]
- Khandare, R.V.; Govindwar, S.P. Phytoremediation of textile dyes and effluents: Current scenario and future prospects. Biotechnol. Adv. 2015, 33, 1697–1714. [Google Scholar] [CrossRef] [PubMed]
- Goudarztalejerdi, A.; Tabatabaei, M.; Eskandari, M.H.; Mowla, D.; Iraji, A. Evaluation of bioremediation potential and biopolymer production of pseudomonads isolated from petroleum hydrocarbon-contaminated areas. Int. J. Environ. Sci. Technol. 2015, 12, 2801–2808. [Google Scholar] [CrossRef]
- Hori, K.; Abe, M.; Unno, H. Production of triacylglycerol and poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by the toluene-degrading bacterium Rhodococcus aetherivorans IAR1. J. Biosci. Bioeng. 2009, 108, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Kobayashi, A.; Ikeda, H.; Unno, H. Rhodococcus aetherivorans IAR1, a new bacterial strain synthesizing poly(3-hydroxybutyrate-co-3-hydroxyvalerate) from toluene. J. Biosci. Bioeng. 2009, 107, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Nikodinovic-Runic, J.; Casey, E.; Duane, G.F.; Mitic, D.; Hume, A.R.; Kenny, S.T.; O’Connor, K.E. Process analysis of the conversion of styrene to biomass and medium chain length polyhydroxyalkanoate in a two-phase bioreactor. Biotechnol. Bioeng. 2011, 108, 2447–2455. [Google Scholar] [CrossRef] [PubMed]
- Tan, G.-Y.A.; Chen, C.-L.; Ge, L.; Li, L.; Tan, S.N.; Wang, J.-Y. Bioconversion of styrene to poly(hydroxyalkanoate) (PHA) by the new bacterial strain Pseudomonas putida NBUS12. Microbes Environ. 2015, 30, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Goff, M.; Ward, P.G.; O’Connor, K.E. Improvement of the conversion of polystyrene to polyhydroxyalkanoate through the manipulation of the microbial aspect of the process: A nitrogen feeding strategy for bacterial cells in a stirred tank reactor. J. Biotechnol. 2007, 132, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Tamboli, D.P.; Kurade, M.B.; Waghmode, T.R.; Joshi, S.M.; Govindwar, S.P. Exploring the ability of Sphingobacterium sp. ATM to degrade textile dye Direct Blue GLL, mixture of dyes and textile effluent and production of polyhydroxyhexadecanoic acid using waste biomass generated after dye degradation. J. Hazard. Mater. 2010, 182, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Tamboli, D.P.; Gomare, S.S.; Kalme, S.S.; Jadhav, U.U.; Govindwar, S.P. Degradation of Orange 3R, mixture of dyes and textile effluent and production of polyhydroxyalkanoates from biomass obtained after degradation. Int. Biodeterior. Biodegrad. 2010, 64, 755–763. [Google Scholar] [CrossRef]
- Kahlon, R.S. Pseudomonas: Molecular and Applied Biology; Springer: Berlin, Germany, 2016. [Google Scholar]
- Nikodinovic, J.; Kenny, S.T.; Babu, R.P.; Woods, T.; Blau, W.J.; O’Connor, K.E. The conversion of BTEX compounds by single and defined mixed cultures to medium-chain-length polyhydroxyalkanoate. Appl. Microbiol. Biotechnol. 2008, 80, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.Y.; Kim, D.Y.; Chung, M.G.; Lee, S.H.; Park, H.Y.; Rhee, Y.H. Biosynthesis of medium-chain-length poly(3-hydroxyalkanoates) by volatile aromatic hydrocarbons-degrading Pseudomonas fulva TY16. Bioresour. Technol. 2010, 101, 8485–8488. [Google Scholar] [CrossRef] [PubMed]
- Narancic, T.; Kenny, S.T.; Djokic, L.; Vasiljevic, B.; O’Connor, K.E.; Nikodinovic-Runic, J. Medium-chain-length polyhydroxyalkanoate production by newly isolated Pseudomonas sp. TN301 from a wide range of polyaromatic and monoaromatic hydrocarbons. J. Appl. Microbiol. 2012, 113, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.G.; Goff, M.; Donner, M. A Two Step Chemo—Biotechnological Conversion of Polystyrene to a Biodegradable Thermoplastic. Environ. Sci. Technol. 2006, 40, 2433–2437. [Google Scholar]
- Kenny, S.T.; Runic, J.N.; Kaminsky, W.; Woods, T.; Babu, R.P.; Keely, C.M.; Blau, W.; O’Connor, K.E. Up-cycling of PET (Polyethylene Terephthalate) to the biodegradable plastic PHA (Polyhydroxyalkanoate). Environ. Sci. Technol. 2008, 42, 7696–7701. [Google Scholar] [CrossRef] [PubMed]
- Guzik, M.W.; Kenny, S.T.; Duane, G.F.; Casey, E.; Woods, T.; Babu, R.P.; Nikodinovic-Runic, J.; Murray, M.; O’Connor, K.E. Conversion of post consumer polyethylene to the biodegradable polymer polyhydroxyalkanoate. Appl. Microbiol. Biotechnol. 2014, 98, 4223–4232. [Google Scholar] [CrossRef] [PubMed]
- Tamboli, D.P.; Kagalkar, A.N.; Jadhav, M.U.; Jadhav, J.P.; Govindwar, S.P. Production of polyhydroxyhexadecanoic acid by using waste biomass of Sphingobacterium sp. ATM generated after degradation of textile dye Direct Red 5B. Bioresour. Technol. 2010, 101, 2421–2427. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, D.K.; Saraf, M. Halophiles: Biodiversity and Sustainable Exploitation; Springer: Berlin, Germany, 2015; Volume 6. [Google Scholar]
- Konstantinidis, G. Elsevier’S Dictionary of Medicine and Biology Greek German Italian Latin; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Setati, M. Diversity and industrial potential of hydrolaseproducing halophilic/halotolerant eubacteria. Afr. J. Biotechnol. 2010, 9, 1555–1560. [Google Scholar]
- Yin, J.; Chen, J.C.; Wu, Q.; Chen, G.Q. Halophiles, coming stars for industrial biotechnology. Biotechnol. Adv. 2015, 33, 1433–1442. [Google Scholar] [CrossRef] [PubMed]
- Quillaguamán, J.; Guzmán, H.; Van-Thuoc, D.; Hatti-Kaul, R. Synthesis and production of polyhydroxyalkanoates by halophiles: Current potential and future prospects. Appl. Microbiol. Biotechnol. 2010, 85, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Kirk, R.G.; Ginzburg, M. Ultrastructure of two species of halobacterium. J. Ultrastruct. Res. 1972, 41, 80–94. [Google Scholar] [CrossRef]
- Rodriguez-Valera, F.; Ruiz-Berraquero, F.; Ramos-Cormenzana, A. Isolation of Extremely Halophilic Bacteria Able to Grow in Defined Inorganic Media with Single Carbon Sources. Microbiology 1980, 119, 535–538. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Pramanik, A.; Maji, S.K.; Haldar, S.; Mukhopadhyay, U.K.; Mukherjee, J. Utilization of vinasse for production of poly-3-(hydroxybutyrate-co-hydroxyvalerate) by Haloferax mediterranei. AMB Express 2012, 2, 34. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.; Saha, J.; Haldar, S.; Bhowmic, A.; Mukhopadhyay, U.K.; Mukherjee, J. Production of poly-3-(hydroxybutyrate-co-hydroxyvalerate) by Haloferax mediterranei using rice-based ethanol stillage with simultaneous recovery and re-use of medium salts. Extremophiles 2014, 18, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Pais, J.; Serafim, L.S.; Freitas, F.; Reis, M.A.M. Conversion of cheese whey into poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by Haloferax mediterranei. New Biotechnol. 2016, 33, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Alsafadi, D.; Al-Mashaqbeh, O. A one-stage cultivation process for the production of poly-3-(hydroxybutyrate-co-hydroxyvalerate) from olive mill wastewater by Haloferax mediterranei. New Biotechnol. 2017, 34, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.; Xue, Y.-S.; Aibaidula, G.; Chen, G.-Q. Unsterile and continuous production of polyhydroxybutyrate by Halomonas TD01. Bioresour. Technol. 2011, 102, 8130–8136. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Ling, C.; Yang, T.; Chen, X.; Chen, Y.; Deng, H.; Wu, Q.; Chen, J.; Chen, G.-Q. A seawater-based open and continuous process for polyhydroxyalkanoates production by recombinant Halomonas campaniensis LS21 grown in mixed substrates. Biotechnol. Biofuels 2014, 7, 108. [Google Scholar] [CrossRef]
- Salgaonkar, B.B.; Mani, K.; Braganca, J.M. Characterization of polyhydroxyalkanoates accumulated by a moderately halophilic salt pan isolate Bacillus megaterium strain H16. J. Appl. Microbiol. 2013, 114, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Moorkoth, D.; Nampoothiri, K.M. Production and characterization of poly(3-hydroxy butyrate-co-3 hydroxyvalerate) (PHBV) by a novel halotolerant mangrove isolate. Bioresour. Technol. 2016, 201, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Contreras, A.; Koller, M.; Braunegg, G.; Marqués-Calvo, M.S. Poly[(R)-3-hydroxybutyrate] production under different salinity conditions by a novel Bacillus megaterium strain. New Biotechnol. 2016, 33, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-Q. New challenges and opportunities for industrial biotechnology. Microb. Cell Fact. 2012, 11, 111. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yin, J.; Chen, G.Q. Polyhydroxyalkanoates, challenges and opportunities. Curr. Opin. Biotechnol. 2014, 30, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Ye, J.; Shen, R.; Zong, Y.; Zhao, X.; Lou, C.; Chen, G.-Q. Semirational Approach for Ultrahigh Poly(3-hydroxybutyrate) Accumulation in Escherichia coli by Combining One-Step Library Construction and High-Throughput Screening. ACS Synth. Biol. 2016, 5, 1308–1317. [Google Scholar] [PubMed]
- Fu, X.-Z.; Tan, D.; Aibaidula, G.; Wu, Q.; Chen, J.-C.; Chen, G.-Q. Development of Halomonas TD01 as a host for open production of chemicals. Metab. Eng. 2014, 23, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, H.; Jiang, X.; Chen, G.-Q. Engineering Escherichia coli for enhanced production of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) in larger cellular space. Metab. Eng. 2014, 25, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.-R.; Wang, H.; Shen, R.; Chen, G.-Q. Engineering the bacterial shapes for enhanced inclusion bodies accumulation. Metab. Eng. 2015, 29, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.-R.; Chen, G.-Q. Morphology engineering of bacteria for bio-production. Biotechnol. Adv. 2016, 34, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, D.; Jendrossek, D. Localization of poly(3-Hydroxybutyrate) (PHB) granule-associated proteins during PHB granule formation and identification of two new phasins, phap6 and phap7, in Ralstonia eutropha H16. J. Bacteriol. 2012, 194, 5909–5921. [Google Scholar] [CrossRef] [PubMed]
- Martínez, V.; García, P.; García, J.L.; Prieto, M.A. Controlled autolysis facilitates the polyhydroxyalkanoate recovery in Pseudomonas putida KT2440. Microb. Biotechnol. 2011, 4, 533–547. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, L.; Wu, L.-P.; Chen, J.; Chen, G.-Q. Synthesis of Diblock copolymer poly-3-hydroxybutyrate -block-poly-3-hydroxyhexanoate [PHB-b-PHHx] by a β-oxidation weakened Pseudomonas putida KT2442. Microb. Cell Fact. 2012, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, L.; Wu, L.-P.; Dechuan, M.; Chen, J.; Wu, Q.; Chen, G.-Q. Pseudomonas putida KT2442 as a platform for the biosynthesis of polyhydroxyalkanoates with adjustable monomer contents and compositions. Bioresour. Technol. 2013, 142, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Cai, L.; Wu, L.; Zeng, G.; Chen, J.; Wu, Q.; Chen, G.-Q. Microbial Synthesis of Functional Homo-, Random, and Block Polyhydroxyalkanoates by β-Oxidation Deleted Pseudomonas entomophila. Biomacromolecules 2014, 15, 2310–2319. [Google Scholar] [CrossRef] [PubMed]
- Shen, R.; Cai, L.W.; Meng, D.C.; Wu, L.P.; Guo, K.; Dong, G.X.; Liu, L.; Chen, J.C.; Wu, Q.; Chen, G.Q. Benzene containing polyhydroxyalkanoates homo- and copolymers synthesized by genome edited Pseudomonas entomophila. Sci. China Life Sci. 2014, 57, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.C.; Shen, R.; Yao, H.; Chen, J.C.; Wu, Q.; Chen, G.Q. Engineering the diversity of polyesters. Curr. Opin. Biotechnol. 2014, 29, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, L.; Wu, L.-P.; Meng, D.; Chen, J.; Chen, G.-Q. Biosynthesis and Characterization of Diblock Copolymer of P(3-Hydroxypropionate)-block-P(4-hydroxybutyrate) from Recombinant Escherichia coli. Biomacromolecules 2013, 14, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Q.; Wang, Q.; Liang, Q.; Qi, Q. Synthesis of polyhydroxyalkanoates from glucose that contain medium-chain-length monomers via the reversed fatty acid β-oxidation cycle in Escherichia coli. Metab. Eng. 2014, 24, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Luan, Y.; Cheng, X.; Zhuang, Q.; Qi, Q. Engineering of Escherichia coli for the biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from glucose. Appl. Microbiol. Biotechnol. 2015, 99, 2593–2602. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.-C.; Wang, Y.; Wu, L.-P.; Shen, R.; Chen, J.-C.; Wu, Q.; Chen, G.-Q. Production of poly(3-hydroxypropionate) and poly(3-hydroxybutyrate-co-3-hydroxypropionate) from glucose by engineering Escherichia coli. Metab. Eng. 2015, 29, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; Hajnal, I.; Wu, H.; Lv, L.; Ye, J. Engineering Biosynthesis Mechanisms for Diversifying Polyhydroxyalkanoates. Trends Biotechnol. 2015, 33, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Koller, M.; Maršálek, L.; de Sousa Dias, M.M.; Braunegg, G. Producing microbial polyhydroxyalkanoate (PHA) biopolyesters in a sustainable manner. New Biotechnol. 2017, 37, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Możejko-Ciesielska, J.; Kiewisz, R. Bacterial polyhydroxyalkanoates: Still fabulous? Microbiol. Res. 2016, 192, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Bugnicourt, E.; Cinelli, P.; Lazzeri, A.; Alvarez, V. Polyhydroxyalkanoate (PHA): Review of synthesis, characteristics, processing and potential applications in packaging. Express Polym. Lett. 2014, 8, 791–808. [Google Scholar] [CrossRef]
- Oliveira, C.S.S.; Silva, C.E.; Carvalho, G.; Reis, M.A. Strategies for efficiently selecting PHA producing mixed microbial cultures using complex feedstocks: Feast and famine regime and uncoupled carbon and nitrogen availabilities. New Biotechnol. 2016, 37, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, R.; van Loosdrecht, M.C. Mixed culture biotechnology for bioenergy production. Curr. Opin. Biotechnol. 2007, 18, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Serafim, L.S.; Lemos, P.C.; Albuquerque, M.G.E.; Reis, M.A.M. Strategies for PHA production by mixed cultures and renewable waste materials. Appl. Microbiol. Biotechnol. 2008, 81, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Jiang, Y.; Kleerebezem, R.; Muyzer, G.; Van Loosdrecht, M.C.M. Enrichment of a mixed bacterial culture with a high polyhydroxyalkanoate storage capacity. Biomacromolecules 2009, 10, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Van Loosdrecht, M.C.M.; Pot, M.A.; Heijnen, J.J. Importance of bacterial storage polymers in bioprocesses. Water Sci. Technol. 1997, 35, 41–47. [Google Scholar] [CrossRef]
- Dircks, K.; Beun, J.J.; Van Loosdrecht, M.; Heijnen, J.J.; Henze, M. Glycogen metabolism in aerobic mixed cultures. Biotechnol. Bioeng. 2001, 73, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Moralejo-Gárate, H.; Mar’Atusalihat, E.; Kleerebezem, R.; Van Loosdrecht, M.C.M. Microbial community engineering for biopolymer production from glycerol. Appl. Microbiol. Biotechnol. 2011, 92, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Dionisi, D.; Carucci, G.; Petrangeli Papini, M.; Riccardi, C.; Majone, M.; Carrasco, F. Olive oil mill effluents as a feedstock for production of biodegradable polymers. Water Res. 2005, 39, 2076–2084. [Google Scholar] [CrossRef] [PubMed]
- Ntaikou, I.; Valencia Peroni, C.; Kourmentza, C.; Ilieva, V.I.; Morelli, A.; Chiellini, E.; Lyberatos, G. Microbial bio-based plastics from olive-mill wastewater: Generation and properties of polyhydroxyalkanoates from mixed cultures in a two-stage pilot scale system. J. Biotechnol. 2014, 188C, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Duque, A.F.; Oliveira, C.S.S.; Carmo, I.T.D.; Gouveia, A.R.; Pardelha, F.; Ramos, A.M.; Reis, M.A.M. Response of a three-stage process for PHA production by mixed microbial cultures to feedstock shift: Impact on polymer composition. New Biotechnol. 2014, 31, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Amulya, K.; Jukuri, S.; Venkata Mohan, S. Sustainable multistage process for enhanced productivity of bioplastics from waste remediation through aerobic dynamic feeding strategy: Process integration for up-scaling. Bioresour. Technol. 2015, 188, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Valentino, F.; Morgan-Sagastume, F.; Campanari, S.; Villano, M.; Werker, A.; Majone, M. Carbon recovery from wastewater through bioconversion into biodegradable polymers. New Biotechnol. 2016, 37, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Kourmentza, C.; Mitova, E.; Stoyanova, N.; Ntaikou, I.; Kornaros, M. Investigation of PHAs production from acidified olive oil mill wastewater (OOMW) by pure cultures of Pseudomonas spp. strains. New Biotechnol. 2009, 25, S269. [Google Scholar] [CrossRef]
- Kourmentza, C.; Ntaikou, I.; Kornaros, M.; Lyberatos, G. Production of PHAs from mixed and pure cultures of Pseudomonas sp. using short-chain fatty acids as carbon source under nitrogen limitation. Desalination 2009, 248, 723–732. [Google Scholar] [CrossRef]
- Marang, L.; Jiang, Y.; van Loosdrecht, M.C.M.; Kleerebezem, R. Butyrate as preferred substrate for polyhydroxybutyrate production. Bioresour. Technol. 2013, 142, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Hu, H.; Ji, H.; Cai, J.; He, N.; Li, Q.; Wang, Y. Production of poly(hydroxybutyrate-hydroxyvalerate) from waste organics by the two-stage process: Focus on the intermediate volatile fatty acids. Bioresour. Technol. 2014, 166, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.M.L.; Lemos, P.C.; Serafim, L.S.; Oliveira, C.; Eiroa, M.; Albuquerque, M.G.E.; Ramos, A.M.; Oliveira, R.; Reis, M.A.M. Recent advances in polyhydroxyalkanoate production by mixed aerobic cultures: From the substrate to the final product. Macromol. Biosci. 2006, 6, 885–906. [Google Scholar] [CrossRef] [PubMed]
- Salehizadeh, H.; Van Loosdrecht, M.C.M. Production of polyhydroxyalkanoates by mixed culture: Recent trends and biotechnological importance. Biotechnol. Adv. 2004, 22, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Reis, M.A.M.; Serafim, L.S.; Lemos, P.C.; Ramos, A.M.; Aguiar, F.R.; Van Loosdrecht, M.C.M. Production of polyhydroxyalkanoates by mixed microbial cultures. Bioprocess Biosyst. Eng. 2003, 25, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, M.G.E.; Eiroa, M.; Torres, C.; Nunes, B.R.; Reis, M.A.M. Strategies for the development of a side stream process for polyhydroxyalkanoate (PHA) production from sugar cane molasses. J. Biotechnol. 2007, 130, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; De Roo, G.; Ruth, K.; Witholt, B.; Zinn, M. Simultaneous Accumulation and Degradation of Polyhydroxyalkanoates: Futile Cycle or Clever Regulation? Biomacromolecules 2009, 10, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Frigon, D.; Muyzer, G.; Van Loosdrecht, M.; Raskin, L. rRNA and poly-β-hydroxybutyrate dynamics in bioreactors subjected to feast and famine cycles. Appl. Environ. Microbiol. 2006, 72, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- Prieto, A.; Escapa, I.F.; Martínez, V.; Dinjaski, N.; Herencias, C.; de la Peña, F.; Tarazona, N.; Revelles, O. A holistic view of polyhydroxyalkanoate metabolism in Pseudomonas putida. Environ. Microbiol. 2016, 18, 341–357. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Marang, L.; Kleerebezem, R.; Muyzer, G.; van Loosdrecht, M.C.M. Polyhydroxybutyrate production from lactate using a mixed microbial culture. Biotechnol. Bioeng. 2011, 108, 2022–2035. [Google Scholar] [CrossRef] [PubMed]
- Pardelha, F.A. Constraint-Based Modelling of Mixed Microbial Populations: Application to Polyhydroxyalkanoates Production; Faculdade de Ciências e Tecnologia: Caparica, Portugal, 2013. [Google Scholar]
- Albuquerque, M.G.E.; Concas, S.; Bengtsson, S.; Reis, M.A.M. Mixed culture polyhydroxyalkanoates production from sugar molasses: The use of a 2-stage CSTR system for culture selection. Bioresour. Technol. 2010, 101, 7112–7122. [Google Scholar] [CrossRef] [PubMed]
- Marang, L.; Jiang, Y.; Van Loosdrecht, M.C.M.; Kleerebezem, R. Impact of non-storing biomass on PHA production: An enrichment culture on acetate and methanol. Int. J. Biol. Macromol. 2014, 71, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Korkakaki, E.; van Loosdrecht, M.C.M.; Kleerebezem, R. Survival of the fastest: Selective removal of the side population for enhanced PHA production in a mixed substrate enrichment. Bioresour. Technol. 2016, 216, 1022–1029. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Guo, Z.; Wen, Q.; Huang, L.; Bakke, R.; Du, M. A new method for polyhydroxyalkanoate (PHA) accumulating bacteria selection under physical selective pressure. Int. J. Biol. Macromol. 2015, 72, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Campanari, S.; Matteo, S.; Valentino, F.; Majone, M.; Villano, M. Impact of nitrogen feeding regulation on polyhydroxyalkanoates production by mixed microbial cultures. New Biotechnol. 2017, 37, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Burniol-Figols, A.; Varrone, C.; Daugaard, A.E.; Skiadas, I.V.; Gavala, H.N. Polyhydroxyalkanoates (PHA) production from fermented crude glycerol by mixed microbial cultures. In Proceedings of the Sustain ATV Conference, Book of Abstracts, Kobenhavn, Denmark, 30 November 2016. [Google Scholar]
- Cui, Y.-W.; Zhang, H.-Y.; Lu, P.-F.; Peng, Y.-Z. Effects of carbon sources on the enrichment of halophilic polyhydroxyalkanoate-storing mixed microbial culture in an aerobic dynamic feeding process. Sci. Rep. 2016, 6, 30766. [Google Scholar] [CrossRef] [PubMed]
- Palmeiro-Sánchez, T.; Fra-Vázquez, A.; Rey-Martínez, N.; Campos, J.L.; Mosquera-Corral, A. Transient concentrations of NaCl affect the PHA accumulation in mixed microbial culture. J. Hazard. Mater. 2016, 306, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Fradinho, J.C.; Domingos, J.M.B.; Carvalho, G.; Oehmen, A.; Reis, M.A.M. Polyhydroxyalkanoates production by a mixed photosynthetic consortium of bacteria and algae. Bioresour. Technol. 2013, 132, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Fradinho, J.C.; Reis, M.A.M.; Oehmen, A. Beyond feast and famine: Selecting a PHA accumulating photosynthetic mixed culture in a permanent feast regime. Water Res. 2016, 105, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Fradinho, J.C.; Oehmen, A.; Reis, M.A.M. Photosynthetic mixed culture polyhydroxyalkanoate (PHA) production from individual and mixed volatile fatty acids (VFAs): Substrate preferences and co-substrate uptake. J. Biotechnol. 2014, 185, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Fradinho, J.C.; Oehmen, A.; Reis, M.A.M. Effect of dark/light periods on the polyhydroxyalkanoate production of a photosynthetic mixed culture. Bioresour. Technol. 2013, 148, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Basset, N.; Katsou, E.; Frison, N.; Malamis, S.; Dosta, J.; Fatone, F. Integrating the selection of PHA storing biomass and nitrogen removal via nitrite in the main wastewater treatment line. Bioresour. Technol. 2016, 200, 820–829. [Google Scholar] [CrossRef] [PubMed]
- Morgan-Sagastume, F.; Karlsson, A.; Johansson, P.; Pratt, S.; Boon, N.; Lant, P.; Werker, A. Production of polyhydroxyalkanoates in open, mixed cultures from a waste sludge stream containing high levels of soluble organics, nitrogen and phosphorus. Water Res. 2010, 44, 5196–5211. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Xiong, H.; Wang, H.; Shi, H.; Sheng, X.; Sun, R.; Chen, G. Production of polyhydroxyalkanoates (PHA) by bacterial consortium from excess sludge fermentation liquid at laboratory and pilot scales. Bioresour. Technol. 2014, 171, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Anterrieu, S.; Quadri, L.; Geurkink, B.; Dinkla, I.; Bengtsson, S.; Arcos-Hernandez, M.; Alexandersson, T.; Morgan-Sagastume, F.; Karlsson, A.; Hjort, M.; et al. Integration of biopolymer production with process water treatment at a sugar factory. New Biotechnol. 2014, 31, 308–323. [Google Scholar] [CrossRef] [PubMed]
- Satoh, H.; Iwamoto, Y.; Mino, T.; Matsuo, T. Activated sludge as a possible source of biodegradable plastic. Water Sci. Technol. 1998, 38, 103–109. [Google Scholar] [CrossRef]
- Din, M.F.; Mohanadoss, P.; Ujang, Z.; van Loosdrecht, M.; Yunus, S.M.; Chelliapan, S.; Zambare, V.; Olsson, G. Development of Bio-PORec® system for polyhydroxyalkanoates (PHA) production and its storage in mixed cultures of palm oil mill effluent (POME). Bioresour. Technol. 2012, 124, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Pratt, S.; Werker, A.; Morgan-Sagastume, F.; Lant, P. Microaerophilic conditions support elevated mixed culture polyhydroxyalkanoate (PHA) yields, but result in decreased PHA production rates. Water Sci. Technol. 2012, 65, 243–246. [Google Scholar] [CrossRef]
- Wang, X.; Oehmen, A.; Freitas, E.B.; Carvalho, G.; Reis, M.A.M. The link of feast-phase dissolved oxygen (DO) with substrate competition and microbial selection in PHA production. Water Res. 2017, 112, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Gurieff, N.; Lant, P. Comparative life cycle assessment and financial analysis of mixed culture polyhydroxyalkanoate production. Bioresour. Technol. 2007, 98, 3393–3403. [Google Scholar] [CrossRef] [PubMed]
- Takabatake, H.; Satoh, H.; Mino, T.; Matsuo, T. PHA (polyhydroxyalkanoate) production potential of activated sludge treating wastewater. Water Sci. Technol. 2002, 45, 119–126. [Google Scholar] [PubMed]
- Mengmeng, C.; Hong, C.; Qingliang, Z.; Shirley, S.N.; Jie, R. Optimal production of polyhydroxyalkanoates (PHA) in activated sludge fed by volatile fatty acids (VFAs) generated from alkaline excess sludge fermentation. Bioresour. Technol. 2009, 100, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Cavaillé, L.; Grousseau, E.; Pocquet, M.; Lepeuple, A.S.; Uribelarrea, J.L.; Hernandez-Raquet, G.; Paul, E. Polyhydroxybutyrate production by direct use of waste activated sludge in phosphorus-limited fed-batch culture. Bioresour. Technol. 2013, 149, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Cavaillé, L.; Albuquerque, M.; Grousseau, E.; Lepeuple, A.S.; Uribelarrea, J.L.; Hernandez-Raquet, G.; Paul, E. Understanding of polyhydroxybutyrate production under carbon and phosphorus-limited growth conditions in non-axenic continuous culture. Bioresour. Technol. 2016, 201, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; van Geest, J.; Kleerebezem, R.; van Loosdrecht, M.C.M. Short- and long-term temperature effects on aerobic polyhydroxybutyrate producing mixed cultures. Water Res. 2010, 44, 1689–1700. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, M.G.E.; Martino, V.; Pollet, E.; Avérous, L.; Reis, M.A.M. Mixed culture polyhydroxyalkanoate (PHA) production from volatile fatty acid (VFA)-rich streams: Effect of substrate composition and feeding regime on PHA productivity, composition and properties. J. Biotechnol. 2011, 151, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Villano, M.; Beccari, M.; Dionisi, D.; Lampis, S.; Miccheli, A.; Vallini, G.; Majone, M. Effect of pH on the production of bacterial polyhydroxyalkanoates by mixed cultures enriched under periodic feeding. Process Biochem. 2010, 45, 714–723. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, L.; Wen, Q.; Guo, Z. Efficient polyhydroxyalkanoate (PHA) accumulation by a new continuous feeding mode in three-stage mixed microbial culture (MMC) PHA production process. J. Biotechnol. 2015, 209, 68–75. [Google Scholar] [CrossRef]
- Serafim, L.S.; Lemos, P.C.; Oliveira, R.; Reis, M.A.M. Optimization of polyhydroxybutyrate production by mixed cultures submitted to aerobic dynamic feeding conditions. Biotechnol. Bioeng. 2004, 87, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Moita, R.; Freches, A.; Lemos, P.C. Crude glycerol as feedstock for polyhydroxyalkanoates production by mixed microbial cultures. Water Res. 2014, 58, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Pardelha, F.; Albuquerque, M.G.E.; Reis, M.A.M.; Dias, J.M.L.; Oliveira, R. Flux balance analysis of mixed microbial cultures: Application to the production of polyhydroxyalkanoates from complex mixtures of volatile fatty acids. J. Biotechnol. 2012, 162, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Meng, H.; Nie, Z.; Zhang, M. Polyhydroxyalkanoate production from fermented volatile fatty acids: Effect of pH and feeding regimes. Bioresour. Technol. 2013, 128, 533–538. [Google Scholar] [CrossRef]
- Jiang, Y.; Marang, L.; Tamis, J.; van Loosdrecht, M.C.M.; Dijkman, H.; Kleerebezem, R. Waste to resource: Converting paper mill wastewater to bioplastic. Water Res. 2012, 46, 5517–5530. [Google Scholar] [CrossRef] [PubMed]
- Valentino, F.; Karabegovic, L.; Majone, M.; Morgan-Sagastume, F.; Werker, A. Polyhydroxyalkanoate (PHA) storage within a mixed-culture biomass with simultaneous growth as a function of accumulation substrate nitrogen and phosphorus levels. Water Res. 2015, 77, 49–63. [Google Scholar] [CrossRef]
- Johnson, K.; Kleerebezem, R.; van Loosdrecht, M.C.M. Influence of ammonium on the accumulation of polyhydroxybutyrate (PHB) in aerobic open mixed cultures. J. Biotechnol. 2010, 147, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Venkateswar Reddy, M.; Venkata Mohan, S. Effect of substrate load and nutrients concentration on the polyhydroxyalkanoates (PHA) production using mixed consortia through wastewater treatment. Bioresour. Technol. 2012, 114, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Moralejo-Garate, H.; Palmeiro-Sanchez, T.; Kleerebezem, R.; Mosquera-Corral, A.; Campos, J.L.; van Loosdrecht, M.C.M. Influence of the cycle lenght on the production of PHA from Glycerol by Bacterial Enrichments in Sequencing Batch Reactors. Biotechnol. Bioeng. 2013, 110, 3148–3155. [Google Scholar] [CrossRef] [PubMed]
- Dionisi, D.; Majone, M.; Vallini, G.; Di Gregorio, S.; Beccari, M. Effect of the applied organic load rate on biodegradable polymer production by mixed microbial cultures in a sequencing batch reactor. Biotechnol. Bioeng. 2006, 93, 76–88. [Google Scholar] [CrossRef]
- Chakravarty, P.; Mhaisalkar, V.; Chakrabarti, T. Study on poly-hydroxyalkanoate (PHA) production in pilot scale continuous mode wastewater treatment system. Bioresour. Technol. 2010, 101, 2896–2899. [Google Scholar] [CrossRef] [PubMed]
- Morgan-Sagastume, F.; Hjort, M.; Cirne, D.; Gérardin, F.; Lacroix, S.; Gaval, G.; Karabegovic, L.; Alexandersson, T.; Johansson, P.; Karlsson, A.; et al. Integrated production of polyhydroxyalkanoates (PHAs) with municipal wastewater and sludge treatment at pilot scale. Bioresour. Technol. 2015, 181, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, S.; Karlsson, A.; Alexandersson, T.; Quadri, L.; Hjort, M.; Johansson, P.; Morgan-Sagastume, F.; Anterrieu, S.; Arcos-Hernandez, M.; Karabegovic, L.; et al. A process for polyhydroxyalkanoate (PHA) production from municipal wastewater treatment with biological carbon and nitrogen removal demonstrated at pilot-scale. New Biotechnol. 2017, 35, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Tamis, J.; Lužkov, K.; Jiang, Y.; van Loosdrecht, M.C.M.; Kleerebezem, R. Enrichment of Plasticicumulans acidivorans at pilot-scale for PHA production on industrial wastewater. J. Biotechnol. 2014, 192, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y. Bacterial polyhydroxyalkanoates. Biotechnol. Bioeng. 1996, 49, 1–14. [Google Scholar] [CrossRef]
- Obruca, S.; Benesova, P.; Marsalek, L.; Marova, I. Use of Lignocellulosic Materials for PHA Production. Chem. Biochem. Eng. Q. 2015, 29, 135–144. [Google Scholar] [CrossRef]
- Ienczak, J.L.; Schmidell, W.; De Aragão, G.M.F. High-cell-density culture strategies for polyhydroxyalkanoate production: A review. J. Ind. Microbiol. Biotechnol. 2013, 40, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Ahn, W.S.; Park, S.J.; Lee, S.Y. Production of poly (3-hydroxybutyrate) from whey by cell recycle fed-batch culture of recombinant Escherichia coli. Biotechnol. Lett. 2001, 23, 235–240. [Google Scholar] [CrossRef]
- Shaffer, D.L.; Werber, J.R.; Jaramillo, H.; Lin, S.; Elimelech, M. Forward osmosis: Where are we now? Desalination 2015, 356, 271–284. [Google Scholar] [CrossRef]
- Jung, K.; Choi, J.D.R.; Lee, D.; Seo, C.; Lee, J.; Lee, S.Y.; Chang, H.N.; Kim, Y.C. Permeation characteristics of volatile fatty acids solution by forward osmosis. Process Biochem. 2015, 50, 669–677. [Google Scholar] [CrossRef]
- Kalafatakis, S.; Braekevelt, S.; Vilhelmsen Carlsen, N.S.; Lange, L.; Skiadas, I.V.; Gavala, H.N. On a novel strategy for water recovery and recirculation in biorefineries through application of forward osmosis membranes. Chem. Eng. J. 2017, 311, 209–216. [Google Scholar] [CrossRef]
- Koller, M.; Niebelschütz, H.; Braunegg, G. Strategies for recovery and purification of poly[(R)-3-hydroxyalkanoates] (PHA) biopolyesters from surrounding biomass. Eng. Life Sci. 2013, 13, 549–562. [Google Scholar] [CrossRef]
- Madkour, M.H.; Heinrich, D.; Alghamdi, M.A.; Shabbaj, I.I.; Steinbüchel, A. PHA recovery from biomass. Biomacromolecules 2013, 14, 2963–2972. [Google Scholar] [CrossRef] [PubMed]
- Anis, S.N.S.; Md Iqbal, N.; Kumar, S.; Amirul, A.A. Effect of different recovery strategies of P(3HB-co-3HHx) copolymer from Cupriavidus necator recombinant harboring the PHA synthase of Chromobacterium sp. USM2. Sep. Purif. Technol. 2013, 102, 111–117. [Google Scholar] [CrossRef]
- Leong, Y.K.; Lan, J.C.-W.; Loh, H.-S.; Ling, T.C.; Ooi, C.W.; Show, P.L. Cloud-point extraction of green-polymers from Cupriavidus necator lysate using thermoseparating-based aqueous two-phase extraction. J. Biosci. Bioeng. 2017, 123, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Aramvash, A.; Gholami-Banadkuki, N.; Seyedkarimi, M.-S. An efficient method for the application of PHA-poor solvents to extract polyhydroxybutyrate from Cupriavidus necator. Biotechnol. Prog. 2016, 32, 1480–1486. [Google Scholar] [CrossRef] [PubMed]
- Samorì, C.; Abbondanzi, F.; Galletti, P.; Giorgini, L.; Mazzocchetti, L.; Torri, C.; Tagliavini, E. Extraction of polyhydroxyalkanoates from mixed microbial cultures: Impact on polymer quality and recovery. Bioresour. Technol. 2015, 189, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.H.; Jeon, J.M.; Yi, D.H.; Kim, J.H.; Seo, H.M.; Rha, C.K.; Sinskey, A.J.; Brigham, C.J. Application of a non-halogenated solvent, methyl ethyl ketone (MEK) for recovery of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) [P(HB-co-HV)] from bacterial cells. Biotechnol. Bioprocess Eng. 2015, 20, 291–297. [Google Scholar] [CrossRef]
- Neves, A.; Müller, J. Use of enzymes in extraction of polyhydroxyalkanoates produced by Cupriavidus necator. Biotechnol. Prog. 2012, 28, 1575–1580. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Mikova, G.; Kleerebezem, R.; van der Wielen, L.A.; Cuellar, M.C. Feasibility study of an alkaline-based chemical treatment for the purification of polyhydroxybutyrate produced by a mixed enriched culture. AMB Express 2015, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- López-Abelairas, M.; García-Torreiro, M.; Lú-Chau, T.; Lema, J.M.; Steinbüchel, A. Comparison of several methods for the separation of poly(3-hydroxybutyrate) from Cupriavidus necator H16 cultures. Biochem. Eng. J. 2015, 93, 250–259. [Google Scholar] [CrossRef]
- Villano, M.; Valentino, F.; Barbetta, A.; Martino, L.; Scandola, M.; Majone, M. Polyhydroxyalkanoates production with mixed microbial cultures: From culture selection to polymer recovery in a high-rate continuous process. New Biotechnol. 2014, 31, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, D.; Madkour, M.H.; Al-Ghamdi, M.A.; Shabbaj, I.I.; Steinbüchel, A. Large scale extraction of poly(3-hydroxybutyrate) from Ralstonia eutropha H16 using sodium hypochlorite. AMB Express 2012, 2, 59. [Google Scholar] [CrossRef] [PubMed]
- Rathi, D.-N.; Amir, H.G.; Abed, R.M.M.; Kosugi, A.; Arai, T.; Sulaiman, O.; Hashim, R.; Sudesh, K. Polyhydroxyalkanoate biosynthesis and simplified polymer recovery by a novel moderately halophilic bacterium isolated from hypersaline microbial mats. J. Appl. Microbiol. 2013, 114, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Hassan, M.A.; Phang, L.Y.; Shirai, Y.; Man, H.C.; Ariffin, H. Intracellular polyhydroxyalkanoates recovery by cleaner halogen-free methods towards zero emission in the palm oil mill. J. Clean. Prod. 2012, 37, 353–360. [Google Scholar] [CrossRef]
- Kachrimanidou, V.; Kopsahelis, N.; Vlysidis, A.; Papanikolaou, S.; Kookos, I.K.; Monje Martínez, B.; Escrig Rondán, M.C.; Koutinas, A.A. Downstream separation of poly(hydroxyalkanoates) using crude enzyme consortia produced via solid state fermentation integrated in a biorefinery concept. Food Bioprod. Process. 2016, 100, 323–334. [Google Scholar] [CrossRef]
- Gutt, B.; Kehl, K.; Ren, Q.; Boesel, L.F. Using ANOVA Models to Compare and Optimize Extraction Protocols of P3HBHV from Cupriavidus necator. Ind. Eng. Chem. Res. 2016, 55, 10355–10365. [Google Scholar] [CrossRef]
- Martino, L.; Cruz, M.V.; Scoma, A.; Freitas, F.; Bertin, L.; Scandola, M.; Reis, M.A.M. Recovery of amorphous polyhydroxybutyrate granules from Cupriavidus necator cells grown on used cooking oil. Int. J. Biol. Macromol. 2014, 71, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Murugan, P.; Han, L.; Gan, C.Y.; Maurer, F.H.J.; Sudesh, K. A new biological recovery approach for PHA using mealworm, Tenebrio molitor. J. Biotechnol. 2016, 239, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Kunasundari, B.; Arza, C.R.; Maurer, F.H.J.; Murugaiyah, V.; Kaur, G.; Sudesh, K. Biological recovery and properties of poly(3-hydroxybutyrate) from Cupriavidus necator H16. Sep. Purif. Technol. 2017, 172, 1–6. [Google Scholar] [CrossRef]
- Martínez, V.; Herencias, C.; Jurkevitch, E.; Prieto, M.A. Engineering a predatory bacterium as a proficient killer agent for intracellular bio-products recovery: The case of the polyhydroxyalkanoates. Sci. Rep. 2016, 6, 24381. [Google Scholar] [CrossRef] [PubMed]
- Martínez, V.; Jurkevitch, E.; García, J.L.; Prieto, M.A. Reward for Bdellovibrio bacteriovorus for preying on a polyhydroxyalkanoate producer. Environ. Microbiol. 2013, 15, 1204–1215. [Google Scholar] [CrossRef] [PubMed]
- Kunasundari, B.; Murugaiyah, V.; Kaur, G.; Maurer, F.H.J.; Sudesh, K. Revisiting the Single Cell Protein Application of Cupriavidus necator H16 and Recovering Bioplastic Granules Simultaneously. PLoS ONE 2013, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Anis, S.N.S.; Iqbal, N.M.; Kumar, S.; Al-Ashraf, A. Increased recovery and improved purity of PHA from recombinant Cupriavidus necator. Bioengineered 2013, 4, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Koller, M.; Bona, R.; Chiellini, E.; Braunegg, G. Extraction of short-chain-length poly-acetone under elevated temperature and pressure. Biotechnol. Lett. 2013, 35, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Aramvash, A.; Gholami-Banadkuki, N.; Moazzeni-Zavareh, F.; Hajizadeh-Turchi, S. An Environmentally Friendly and Efficient Method for Extraction of. J. Microbiol. Biotechnol. 2015, 25, 1936–1943. [Google Scholar] [CrossRef] [PubMed]
- Fei, T.; Cazeneuve, S.; Wen, Z.; Wu, L.; Wang, T. Effective Recovery of Poly-β-Hydroxybutyrate (PHB) Biopolymer from Cupriavidus necator Using a Novel and Environmentally Friendly Solvent System. Biotechnol. Prog. 2016, 32, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Furrer, P.; Panke, S.; Zinn, M. Efficient recovery of low endotoxin medium-chain-length poly ([R]-3-hydroxyalkanoate) from bacterial biomass. J. Microbiol. Methods 2007, 69, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Ali Hassan, M.; Phang, L.-Y.; Ariffin, H.; Shirai, Y.; Ando, Y. Recovery and purification of intracellular polyhydroxyalkanoates from recombinant Cupriavidus necator using water and ethanol. Biotechnol. Lett. 2012, 34, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Leong, Y.K.; Koroh, F.E.; Show, P.L.; Lan, C.-W.J.; Loh, H.-S. Optimisation of Extractive Bioconversion for Green Polymer via Aqueous Two-Phase System Optimisation of Extractive Bioconversion for Green Polymer. Chem. Eng. Trans. 2015, 45, 1495–1500. [Google Scholar]
- Iqbal, M.; Tao, Y.; Xie, S.; Zhu, Y.; Chen, D.; Wang, X.; Huang, L.; Peng, D.; Sattar, A.; Shabbir, M.A.B.; et al. Aqueous two-phase system (ATPS): An overview and advances in its applications. Biol. Proced. Online 2016, 18, 18. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Name of Company | Product (Trademark) | Substrate | Biocatalyst | Production Capacity |
|---|---|---|---|---|
| Biomatera, Canada | PHA resins (Biomatera) | Renewable raw materials | Non-pathogenic, non-transgenic bacteria isolated from soil | |
| Biomer, Germany | PHB pellets (Biomer®) | Sugar (sucrose) | ||
| Bio-On Srl., Italy | PHB, PHBV spheres (minerv®-PHA) | Sugar beets | Cupriavidus necator | 10,000 t/a |
| BluePHA, China | Customized PHBVHHx, PHV, P3HP3HB, P3HP4HB, P3HP, P4HB synthesis | Development of microbial strains via synthetic biology | ||
| Danimer Scientific, USA | mcl-PHA (Nodax® PHA) | Cold pressed canola oil | ||
| Kaneka Corporation, Japan | PHB-PHHx (AONILEX®) | Plant oils | 3500 t/a | |
| Newlight Technologies LLC, USA | PHA resins (AirCarbonTM) | Oxygen from air and carbon from captured methane emissions | Newlight’s 9X biocatalyst | |
| PHB Industrial S.A., Brazil | PHB, PHBV (BIOCYCLE®) | Saccharose | Alcaligenes sp. | 3000 t/a |
| PolyFerm, Canada | mcl-PHA (VersaMerTM PHA) | Sugars, vegetable oils | Naturally selected microorganisms | |
| Shenzhen Ecomann Biotechnology Co. Ltd., China | PHA pellets, resins, microbeads (AmBio®) | Sugar or glucose | 5000 t/a | |
| SIRIM Bioplastics Pilot Plant, Malaysia | Various types of PHA | Palm oil mill effluent (POME), crude palm kernel oil | 2000 t/a | |
| TianAn Biologic Materials Co. Ltd., China | PHB, PHBV (ENMATTM) | Dextrose deriving from corn of cassava grown in China | Ralstonia eutropha | 10,000 t/a, 50,000 t/a by 2020 |
| Tianjin GreenBio Material Co., China | P (3, 4HB) films, pellets/foam pellets (Sogreen®) | Sugar | 10,000 t/a |
| Strain | Carbon Source | PHA | Cultivation Mode | DCW (g L−1) | PHA (g L−1) | PHA (%) | YP/S | Ref. |
|---|---|---|---|---|---|---|---|---|
| Lignocellulose Degraders | ||||||||
| S. degradans | Glucose | PHB | 17.2 | [20] | ||||
| Glucose | PHB | Fed-batch | 12.7 | 2.7 | 21.4 | 0.17 | [21] | |
| Starch | Fed-batch | 11.7 | 2.0 | 17.5 | 0.14 | |||
| Glucose | PHB | Flask | 2.1 | 0.46 | 22.4 | [23] | ||
| Cellobiose | Flask | 2.0 | 0.42 | 20.8 | ||||
| α-Cellulose | Flask | 1.2 | 0.14 | 11.8 | ||||
| Avicel | Flask | 1.0 | 0.15 | 14.6 | ||||
| Sigmacell | Flask | 1.0 | 0.14 | 13.7 | ||||
| CMC | Flask | 1.1 | 0.14 | 12.7 | ||||
| Glucose | Batch | 1.6 | 0.40 | 25.3 | ||||
| Glucose | Fed-batch | 4.2 | 2.20 | 52.8 | ||||
| Avicel | Fed-batch | 2.1 | 0.40 | 19.2 | ||||
| Agarose | PHB | One-step batch | 0.24 | 18.1 | [24] | |||
| Xylan | One-step batch | 0.20 | 22.7 | |||||
| Agarose | Two-step batch | 0.31 | 18.4 | |||||
| Xylan | Two-step batch | 0.24 | 15.3 | |||||
| Co-culture of S. degradans and B. cerues | Agarose | PHB | One-step batch | 0.29 | 19.7 | [24] | ||
| Xylan | One-step batch | 0.27 | 34.5 | |||||
| Agarose | Two-step batch | 0.23 | 15.3 | |||||
| Xylan | Two-step batch | 0.33 | 30.2 | |||||
| C. taiwanensis | Propionate + Glc a | PHBV (88–12) f | Flask | 2.0 | 1.04 | 52 | [22] | |
| Valerate + Glc a | PHBV (49–51) | 1.0 | 0.51 | 51 | ||||
| Hexanoate + Glc a | PHBHHx c | 2.7 | 1.67 | 62 | ||||
| Hexanoate + Glc a + AA b | PHBHHx d | 1.2 | 0.56 | 47 | ||||
| Heptanoate + Glc a | PHBV (65–35) | 1.7 | 0.56 | 33 | ||||
| Heptanoate + Glc a + AA | PHBV (15–85) | 0.3 | 0.05 | 17 | ||||
| Octanoate + Glc a | PHB | 0.4 | 0.05 | 13 | ||||
| Cassava starch + Val e | PHBV (87–13) | 2.8 | 1.88 | 67 | ||||
| Corn starch + Val e | PHBV (80–10) | 3.3 | 2.14 | 65 | ||||
| Potato + Val e | PHBV (80–10) | 2.6 | 1.43 | 55 | ||||
| Sweet potato + Val e | PHBV (80–10) | 1.6 | 0.83 | 52 | ||||
| Wheat starch + Val e | PHBV (80–10) | 4.1 | 1.72 | 42 | ||||
| Polyhydroxyalkanoates and Bioremediation | ||||||||
| P. putida F1 | Benzene | mcl-PHA | Flask | 0.34 | 0.05 | 14 | [41] | |
| Toluene | 0.72 | 0.16 | 22 | |||||
| Ethylbenzene | 0.67 | 0.10 | 15 | |||||
| P. putida mt-2 | Toluene | mcl-PHA | Flask | 0.37 | 0.08 | 22 | [41] | |
| p-Xylene | 0.53 | 0.14 | 26 | |||||
| P. putida CA-3 | Styrene | mcl-PHA | Flask | 0.79 | 0.26 | 33 | [41] | |
| P. fluva TY16 | Benzene | mcl-PHA | Continuous feeding | 2.54 | 19 | 0.03 | [42] | |
| Toluene | 3.87 | 59 | 0.11 | |||||
| Ethylbenzene | 2.80 | 29 | 0.04 | |||||
| P. putida CA-3 | Styrene pyrolysis oil | mcl-PHA | Flask | 2.80 | 1.60 | 57 | 0.10 | [44] |
| Sphingobacterium sp. ATM | Orange 3R dye | PHA | Flask | 3.48 | 65 | [38] | ||
| B. odysseyi SUK3 | 2.10 | 61 | ||||||
| P. desmolyticim NCIM 2112 | 1.12 | 52 | ||||||
| Halophiles | ||||||||
| H. mediterranei DSM 1411 | 25% pre-treated vinasse | PHBV (86–14) | Flask | 19.7 | 70 | 0.87 | [55] | |
| Stillage | PHBV (85–15) | 16.4 | 71 | 0.35 | [56] | |||
| Hydrolyzed cheese whey | PHBV (98.5–1.5) | Batch | 7.54 | 54 | 0.78 | [57] | ||
| 15% v/v olive mill wastewater | PHBV (94-6) | Flask | 0.2 | 43 | [58] | |||
| Halomonas TD01 | Glucose salt medium | PHA | Continuous two-fermentor | 65 | 0.51 | [59] | ||
| Halomonas campaniensis LS21 | Mixed substrates (mostly comprised of kitchen waste) | PHB | Continuous pH-stat | 26 | [60] | |||
| B. megaterium H16 | Glucose salt medium | PHB | Flask | 39 | [61] | |||
| B. megaterium uyuni S29 | Glucose salt medium | PHB | Flask | 5.42 | 2.22 | 41 | 0.13 | [63] |
| Anaerobic-Aerobic Enrichment (AN/AE) (Section 3.1.1) | |||
| Feast phase | Famine phase | ||
| Aeration | No | Yes | |
| e− acceptor | -- (PHA) | Oxygen | |
| Energy source | Glycogen/polyphosphate | Oxidation of PHA | |
| Carbon source | External substrate | PHA | |
| Driving force for PHA accumulation | • Lack of electron acceptor * | ||
| • Transient presence of substrate *** | |||
| Aerobic Dynamic Feeding (ADF) (Section 3.1.2 and Section 3.1.3) | |||
| Classical Aerobic Dynamic Feeding (Section 3.1.2) | |||
| Feast phase | Famine phase | ||
| Aeration | Yes | Yes | |
| e− acceptor | Oxygen | Oxygen | |
| Energy source | Oxidation of substrate | Oxidation of PHA | |
| Carbon source | External substrate | PHA | |
| Nitrogen availability | Yes ** | Yes ** | |
| Driving force for PHA accumulation | • Transient presence of substrate *** | ||
| Aerobic Dynamic Feeding (ADF) with Intermediate Settling Phase (Section 3.1.3) | |||
| Feast phase | Famine phase | ||
| Aeration | Yes | Yes | |
| e− acceptor | Oxygen | Oxygen | |
| Energy source | Oxidation of substrate | Oxidation of PHA | |
| Carbon source | External substrate | PHA | |
| Nitrogen availability | Yes | Yes | |
| Driving force for PHA accumulation | • Transient presence of substrate *** | ||
| • Higher settling capacity of PHA rich cells | |||
| • Elimination of residual COD after feast phase prevents growth of non-PHA accumulating bacteria | |||
| Aerobic Dynamic Feeding (ADF) with Nitrogen Limitation in the Feast-Phase (Section 3.1.3) | |||
| Feast phase | Famine phase | ||
| Aeration | Yes | Yes | |
| e− acceptor | Oxygen | Oxygen | |
| Energy source | Oxidation of substrate | Oxidation of PHA | |
| Carbon source | External substrate | PHA | |
| Nitrogen availability | No | Yes | |
| Driving force for PHA accumulation | • Transient presence of substrate *** | ||
| • Nitrogen limitation during the feast phase | |||
| Photosynthetic Enrichment (Section 3.1.5) | |||
| Photosynthetic Enrichments—Illuminated SBR | |||
| Feast phase | Famine phase | ||
| Aeration | No | No | |
| e− acceptor | -- (PHA) | Oxygen produced by algae | |
| Energy source | Light | Oxidation of PHA + Light | |
| Carbon source | External substrate | PHA | |
| Driving force for PHA accumulation | • Lack of external electron acceptor with presence of light | ||
| • Transient presence of substrate *** | |||
| Photosynthetic Enrichment—Dark Feast Phase | |||
| Feast phase | Famine phase | ||
| Aeration | No | No | |
| e− acceptor | -- (PHA) | Oxygen produced by algae | |
| Energy source | Glycogen | Oxidation of PHA + Light | |
| Carbon source | External substrate | PHA | |
| Driving force for PHA accumulation | • Lack of external electron acceptor with presence of light | ||
| • Transient presence of substrate *** | |||
| Photosynthetic Enrichment—Permanent Feast Phase | |||
| Feast phase | Famine phase | ||
| Aeration | No | No famine phase | |
| e− acceptor | -- (PHA) | ||
| Energy source | Light | ||
| Carbon source | External substrate | ||
| Driving force for PHA accumulation | • Lack of external electron acceptor with presence of light | ||
| Aerobic-Anoxic Enrichment (Section 3.1.6) | |||
| Feast phase | Famine phase | ||
| Aeration | Yes | No | |
| e− acceptor | Oxygen | NO3 /NO2 | |
| Energy source | Oxidation of substrate | Oxidation of PHA | |
| Carbon source | External substrate | PHA | |
| Driving force for PHA accumulation | • Transient presence of substrate *** | ||
| Anoxic-Aerobic Enrichment (Section 3.1.7) | |||
| Feast phase | Famine phase | ||
| Aeration | No | Yes | |
| e− acceptor | NO3/NO2 | Oxygen | |
| Energy source | Oxidation of substrate | Oxidation of PHA | |
| Carbon source | External substrate | PHA | |
| Driving force for PHA accumulation | • Transient presence of substrate *** | ||
| Microaerophilic Enrichment (Section 3.1.8) | |||
| Feast phase | Famine phase | ||
| Aeration | Yes | Yes | |
| e− acceptor | Oxygen | Oxygen | |
| Energy source | Oxidation of substrate | Oxidation of PHA | |
| Carbon source | External substrate | PHA | |
| Driving force for PHA accumulation | • Transient presence of substrate *** | ||
| • Limitation of electron acceptor | |||
| Pilot Plant, Location | Feedstock | Origin of MMC and Enrichment Strategy | YP/S (g/g) * | PHA % (%mol HB: %mol HV) | mg PHA/g X/h | Ref. |
|---|---|---|---|---|---|---|
| Nagpur, India | Pre-fermented milk and ice cream processing wastewater | Activated sludge | 0.425 * | 39–43 | [150] | |
| Lucun WWTP in Wuxi, China | Hydrolyzed and acidified raw excess sludge | Activated sludge/synthetic mixture of VFA, ADF feast famine with carbon limitation and inhibitor of nitrification | 0.044–0.29 * | 2.06–39.31 | [125] | |
| Eslöv, Sweden | Beet process water, 38% in VFA | PHA producing MMC from pre-fermented effluent of Procordia Foods | 60 (85:15 HB:HV) | [126] | ||
| Brussels North WWTP (Aquiris, Belgium) | Pre-hydrolyzed and fermented WWTP sludge | Sludge fed with municipal WW under aerobic feast famine | 0.25–0.38 | 27–38 (66–74:26–34 HB:HV) | 100–140 | [151] |
| Leeuwarden WWTP, Friesland, Netherlands | Fermented residuals from green-house tomato production | Sludge fed with municipal WW under anoxic feast/aerobic famine | 0.30–0.39 | 34–42 (51–58:42:49 HB:HV) | 28–35 | [152] |
| Mars company, Veghel, Netherlands | Fermented wastewater from a candy bar factory | Activated sludge from a WWTP fed with the fermented wastewater under aerobic feast/famine with inhibitor of nitrification | 0.30 | 70–76 (84:16 HB:HV) | [153] |
| Pre-Treatment | Further PHA Retrieval Treatment | Pre-Treatment Conditions | Purity (%) | Recovery (%) | Ref. |
|---|---|---|---|---|---|
| Sodium chloride (NaCl) | NaOH digestion | NaCl (8 g/L), 30 °C, 3 h | 97.7 | 97.5 | [163] |
| Ultrasonication | Aqueous two-phase extraction | Ultrasonication at 30 kHz per cycle 15 min | - | [164] | |
| Sodium hypochlorite (NaClO) | Non-halogenated solvent extraction | NaClO (10%), 37 °C, 1 h | - | [165] | |
| Thermal pre-treatment | Enzymatic digestion and chloroform extraction | Autoclave, 15 min 121 °C | 94.1 | [168] | |
| Thermal pre-treatment 1 | Non-halogenated solvent extraction | 150 °C, 24 h | 50 | [166] | |
| Ultrasonication and glass beds 1 | Non-halogenated solvent extraction | Glass beads (0.5 mm) and Ultrasonication (10 pulses of 2 min) | 50 | [166] | |
| Sodium hypochlorite 1 | Non-halogenated solvent extraction | NaClO (5%), 100 °C, 15 min | 93 | 82 | [166] |
| Hot acetone 1 | Non-halogenated solvent extraction | Acetone, 100 °C, 30 min | - | [167] |
| NPCM Digestion Type | NPCM Disruption Method | Pre-Treatment | PHA Accumulation Method | Disruption Conditions | Microbial Strain | PHA Content in Biomass (%) | Purity (%) | Recovery (%) | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Chemical | NaOH | Chemical Pre-treatment | Centrifugation | NaOH (0.1 M), 30 °C, 1 h, 350 rpm | C. necator | 68 | 90.8 | 95.3 | [163] |
| Chemical | NaOH | Centrifugation | NaOH (0.1 M), 30 °C, 1 h, 350 rpm | C. necator | 68 | 82.7 | 94.4 | [163] | |
| Chemical | NaOH | Lyophilization | Centrifugation | NaOH (0.1 M), 30 °C, 1 h, | C. necator | 68 | 80–90 | 80–90 | [183] |
| Chemical | NaOH | Lyophilization | Centrifugation | NaOH 0.05 M, 3 h, 0 rpm, 4 °C | C. necator | 30 | 98.6 | 96.9 | [174] |
| Chemical | NaOH | Lyophilization and milling | Centrifugation | NaOH (0.5 N), 4 h, 37 °C, 500 rpm | C. necator | 65 | 93 | 80 | [170] |
| Chemical | NaOH | Centrifugation | NaOH (0.2 M), 200 rpm, 30 °C, 1 h | Mixed Culture | 62–72 | 87 | 97 | [169] | |
| Chemical | NaClO | Centrifugation | NaClO (5%) 24 h | Mixed Culture | 46 | 90 | ~100 | [171] | |
| Chemical | NaClO | Mechanical pre-treatment | Precipitation | NaClO 13% (v/v), room temperature, 1 h. | Ralstonia eutropha | 65.2 | 95.6 | 91.3 | [172] |
| Chemical | NaClO | Lyophilization and Milling | Centrifugation | NaClO (13%), 37 °C, 500 rpm,4 h. | C. necator | 65 | 97 | 82 | [170] |
| Chemical | NaOH and SDS | Centrifugation | NaOH (0.2 M) and SDS (0.2 %), 200 rpm, 30°C, 1 h | Mixed Culture | 62–72 | 99 | 91 | [169] | |
| Chemical | SDS | NaClO and Centrifugation | SDS (0.1%), 24 h | H. mediterranei | 70 | ~100 | 97 | [55] | |
| Chemical | SDS | Centrifugation | SDS (0.1%), 24 h | H. mediterranei | 71.2 | ~100 | 97 | [56] | |
| Chemical | SDS | Centrifugation | SDS (0.1%), 60 °C, 2 h | Halomonas sp. SK5 | 48 | 94 | 98 | [173] | |
| Chemical | SDS | Centrifugation | SDS (0.2 %), 200 rpm, 30 °C, 1 h | Mixed Culture | 62–72 | 79 | 63.5 | [169] | |
| Chemical | H2SO4 | Lyophilization and Milling | Chemical treatment and Centrifugation | H2SO4 (0.64 M), 6 h, 80 °C | C. necator | 65 | 98 | 79 | [170] |
| Chemical | Water | Lyophilization | Centrifugation | dH2O, 30 °C, 1 h, | Comamonassp. | 30 | 80.6 | 96 | [174] |
| Chemical | Water | Lyophilization | Centrifugation | dH2O, 30 °C, 18 h | Halomonas sp. | 48 | 94 | 98 | [173] |
| Enzymatic | Alcalase, SDS and EDTA | Centrifugation | Alcalase (0.3 U g−1), SDS (0.3 g g−1), EDTA (0.01 g g−1). Na2HPO4 buffer, 150 rpm, 55 °C, 1 h | C. necator | 37 | 94 | [177] | ||
| Enzymatic | Crude extract | Heat treatment and lyophilization | Centrifugation | Aspergillus oryzae crude extract, Na2HPO4-citric acid buffer and 47 °C | C. necator | 78.9 | 98 | 97 | [175] |
| Enzymatic | Lysozyme | Centrifugation | Lysozyme solution (2 mg/mL). 1 h, 3 °C | C. necator | 41 | 41 | 75 | [176] | |
| Biological | Mealworm (Tenebrio molitor) | Lyophilization | Chemical treatment, centrifugation | 50 g of mealworms fed 5% of their body weight per day for 16 days. | C. necator | 37 | 89% | [178] | |
| Biological | Sprague Dawley rats | Lyophilization and grinding | Chemical treatment, centrifugation | 150–200 g rats were feed 15 g/day/animal, 28 days 25 °C | C. necator | 37 | 89.3 | 100 | [179] |
| Biological | Sprague Dawley rats | Lyophilization | Water | 150–200 g rats were feed 15 g/day/animal, 28 days 25 °C | C. necator | 54 | 82–97 | 40–47 | [182] |
| Biological | Bdellovibrio bacteriovorus HD100 | Centrifugation | P. putida was inoculated with B. bacteriovorus strains 48 h. 30 °C | P. putida | 55 | 60 | [181] | ||
| Biological | Bdellovibrio bacteriovorus HD100 and Bd3709 | Centrifugation | P. putida and E. coli cultures were inoculated with B. bacteriovorus strains 48 h. 30 °C | P. putida | 55 | 80 | [180] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kourmentza, C.; Plácido, J.; Venetsaneas, N.; Burniol-Figols, A.; Varrone, C.; Gavala, H.N.; Reis, M.A.M. Recent Advances and Challenges towards Sustainable Polyhydroxyalkanoate (PHA) Production. Bioengineering 2017, 4, 55. https://doi.org/10.3390/bioengineering4020055
Kourmentza C, Plácido J, Venetsaneas N, Burniol-Figols A, Varrone C, Gavala HN, Reis MAM. Recent Advances and Challenges towards Sustainable Polyhydroxyalkanoate (PHA) Production. Bioengineering. 2017; 4(2):55. https://doi.org/10.3390/bioengineering4020055
Chicago/Turabian StyleKourmentza, Constantina, Jersson Plácido, Nikolaos Venetsaneas, Anna Burniol-Figols, Cristiano Varrone, Hariklia N. Gavala, and Maria A. M. Reis. 2017. "Recent Advances and Challenges towards Sustainable Polyhydroxyalkanoate (PHA) Production" Bioengineering 4, no. 2: 55. https://doi.org/10.3390/bioengineering4020055
APA StyleKourmentza, C., Plácido, J., Venetsaneas, N., Burniol-Figols, A., Varrone, C., Gavala, H. N., & Reis, M. A. M. (2017). Recent Advances and Challenges towards Sustainable Polyhydroxyalkanoate (PHA) Production. Bioengineering, 4(2), 55. https://doi.org/10.3390/bioengineering4020055