
Converting Sugars to Biofuels: Ethanol and Beyond

Abstract
:1. Introduction: Sources of Sugars for Biofuel Production
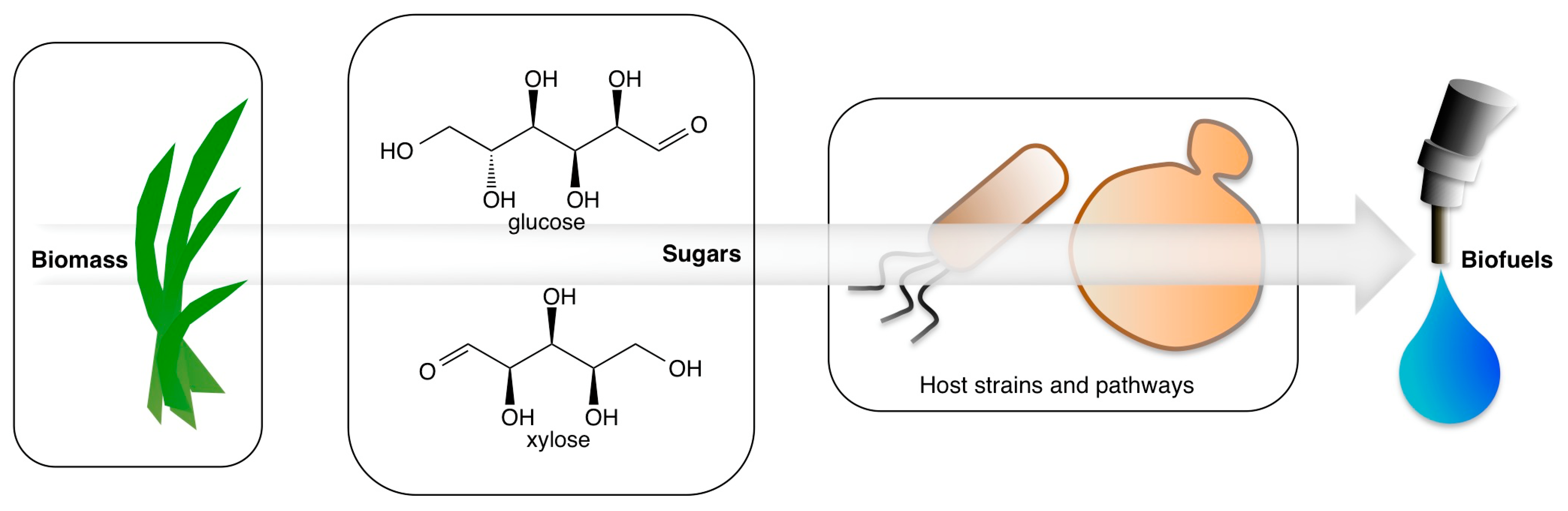
2. Fermentation Pathways and Hosts for Ethanol Production
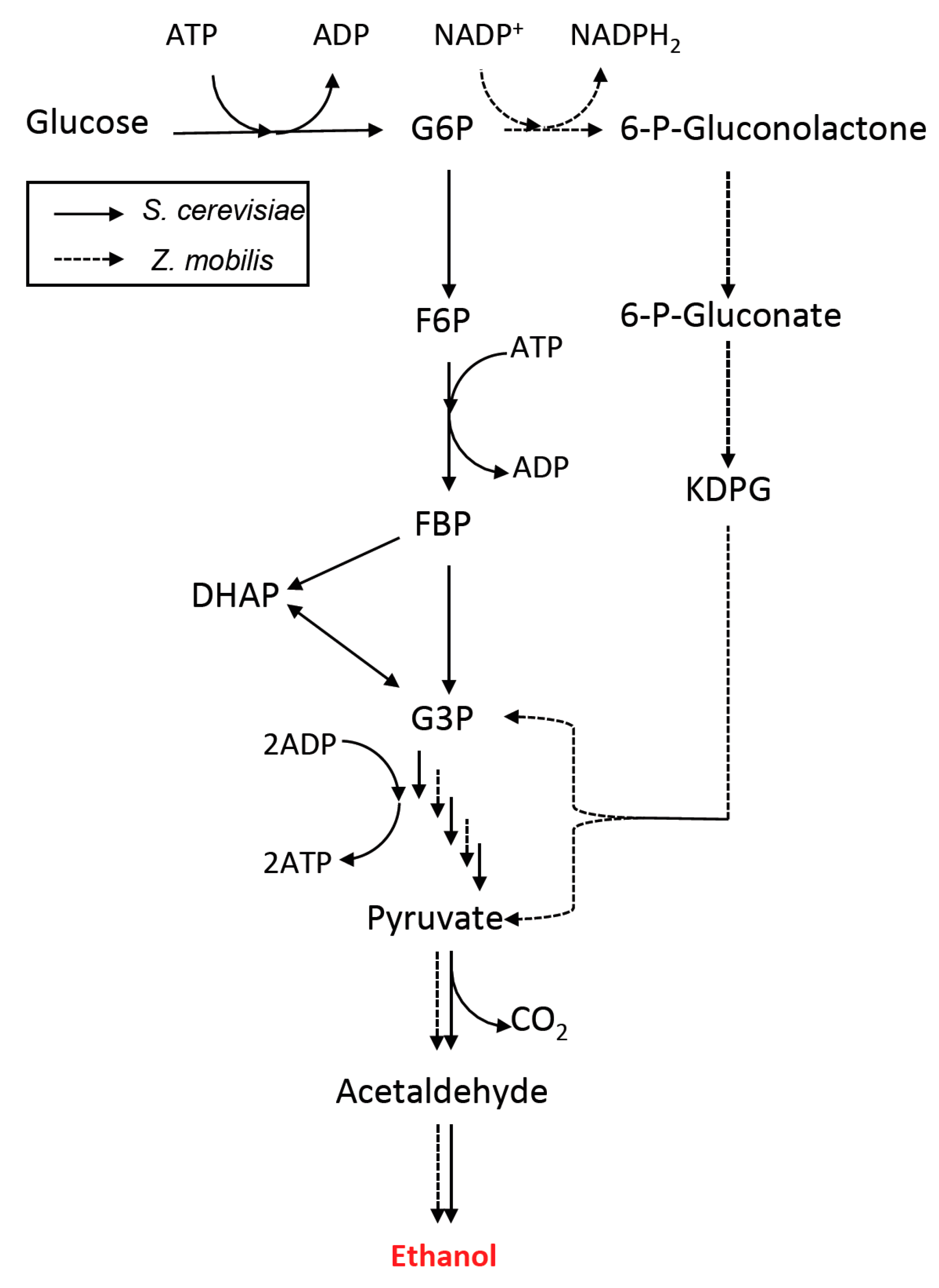
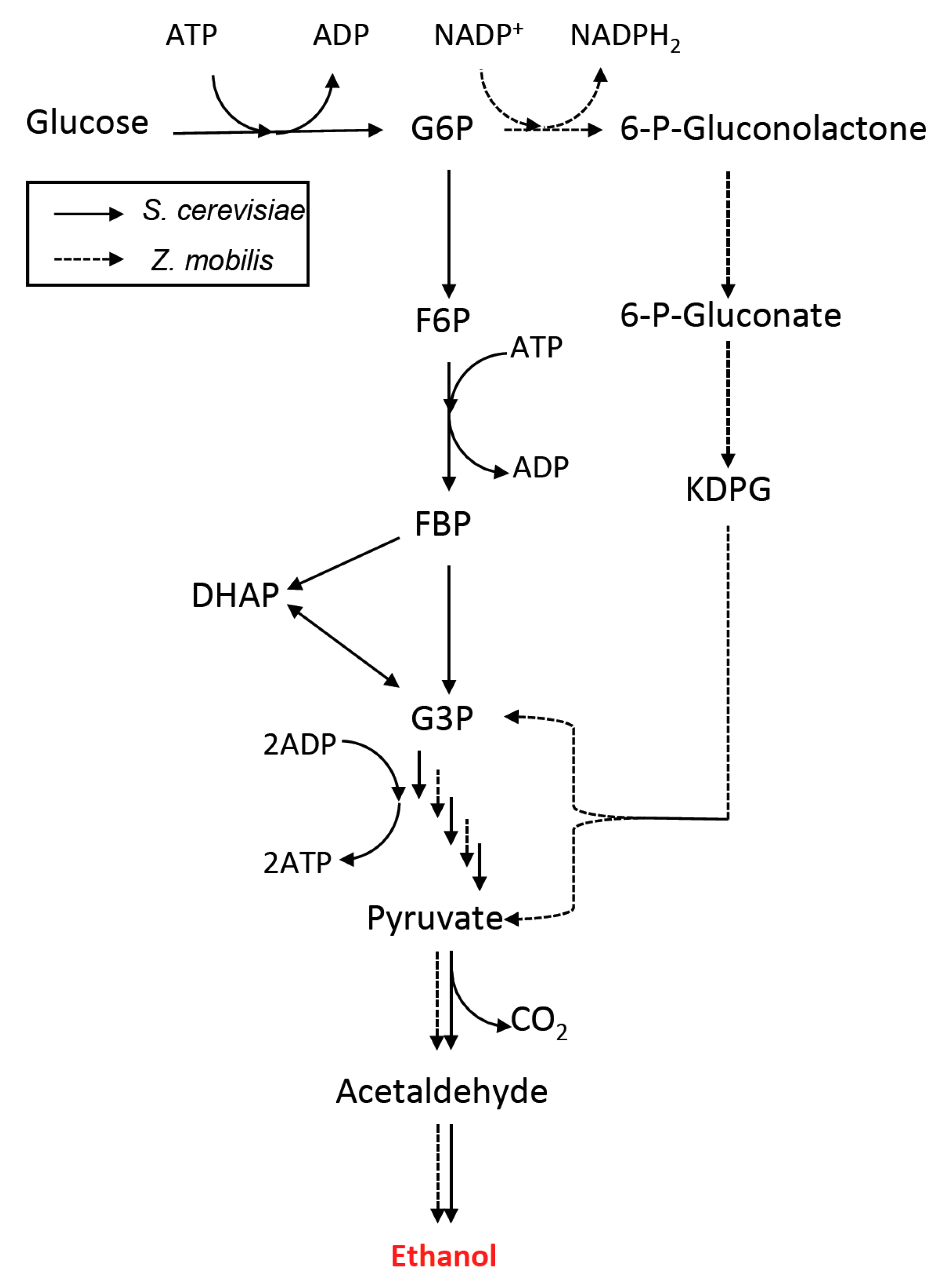
3. Metabolic Pathway and Host Engineering for Ethanol Production
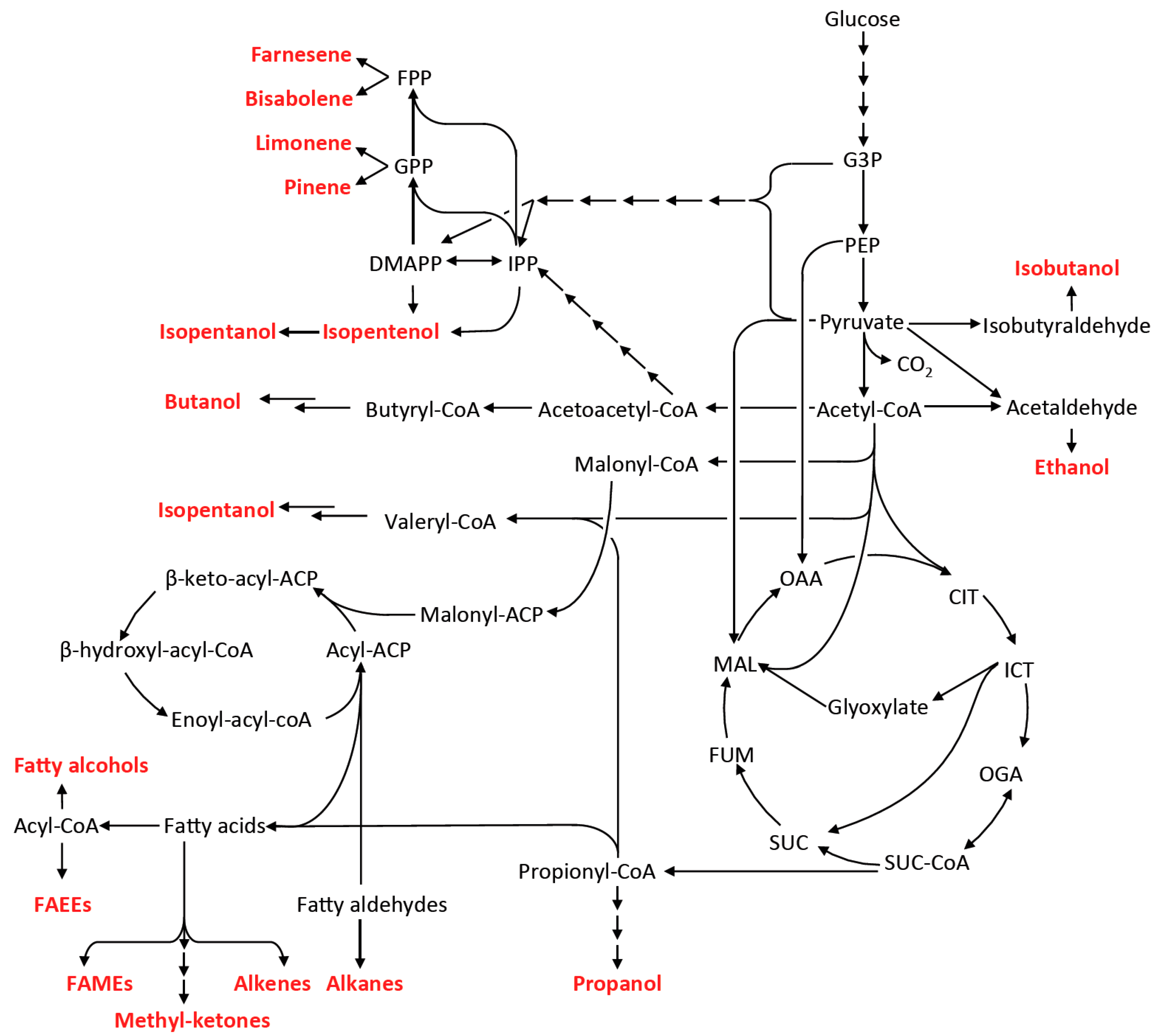
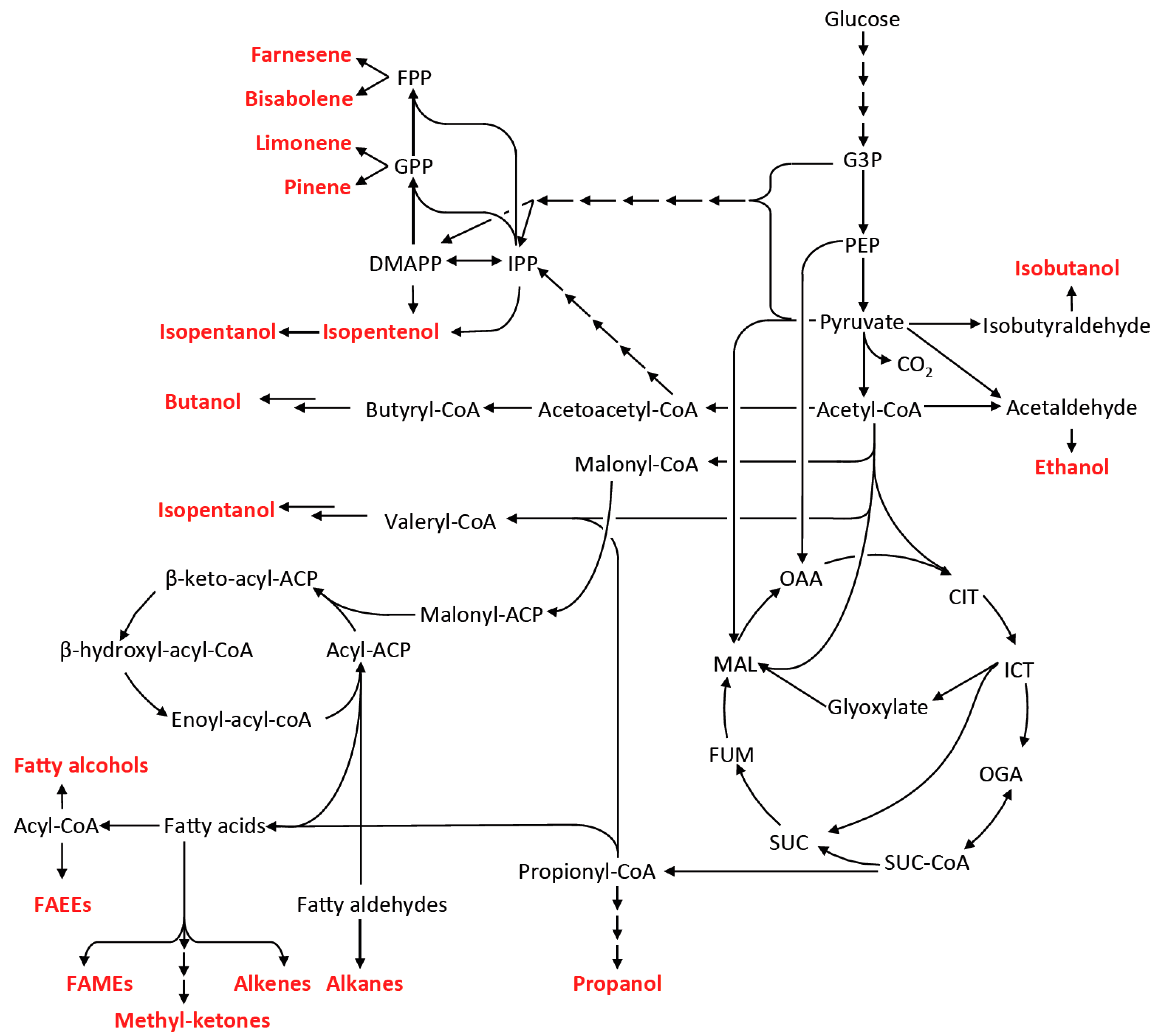
4. Metabolic Pathway and Host Engineering for Advanced Biofuels Production

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product | Pathway | % of Apparent Theoretical Yield (from Glucose) | Highest Titer Reported (g/L) | Host | Reference |
|---|---|---|---|---|---|
| Gasoline | |||||
| 1-Butanol | CoA-dependent | 41% a | 30 | E. coli | [20] |
| Isobutanol | 2-keto acids | 41% a | 50 | E. coli | [21] |
| 3-methyl-1-butanol | 2-keto acids | 33% a | 9.8 | E. coli | [22] |
| 3-methyl-3-butenol | MVA | 41% b | 2.2 | E. coli | [23] |
| 2-methyl-1-butanol | 2-keto acids | 39% b | 1.25 | E. coli | [24] |
| Diesel & Jet Fuel | |||||
| Farnesene | MVA | 25% a | NA | [25] | |
| Farnesene | MEP | 29% a | NA | [25] | |
| Bisabolene | MVA | 25% | 1.15 | E. coli | [26] |
| Limonene | MVA | 25% | 0.605 | E. coli | [26] |
| Pinene | MVA | 25% | 0.032 | E. coli | [27] |
| FAEE | Fatty acids | 35% a | 1.5 | E. coli | [28] |
| Methyl ketones | Fatty acids | 33% | 3.4 | E. coli | [29] |
4.1. Fermentative Pathways for 1-Butanol and Other Short Chain Alcohols
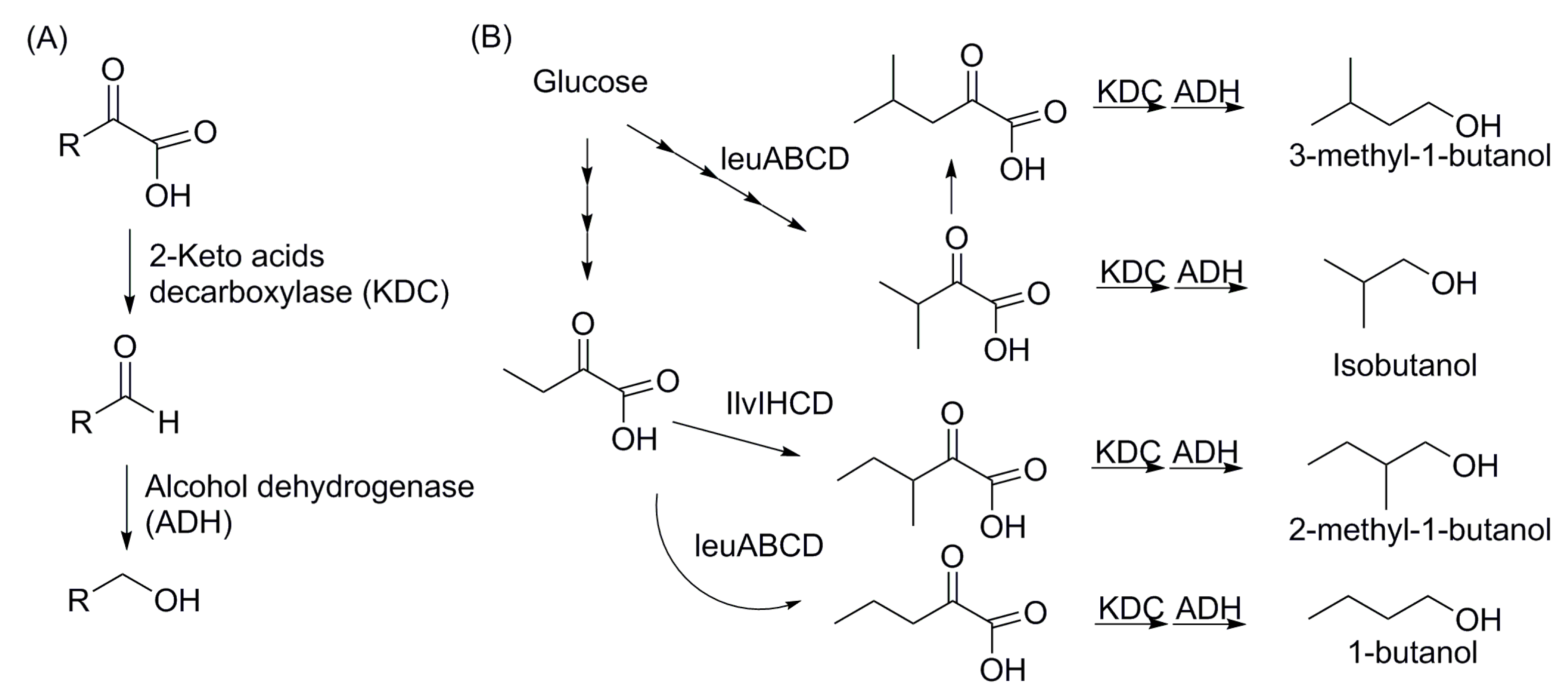
4.2. Non-Fermentative Pathways for Short Chain Alcohols: 2-Keto Acid Pathway
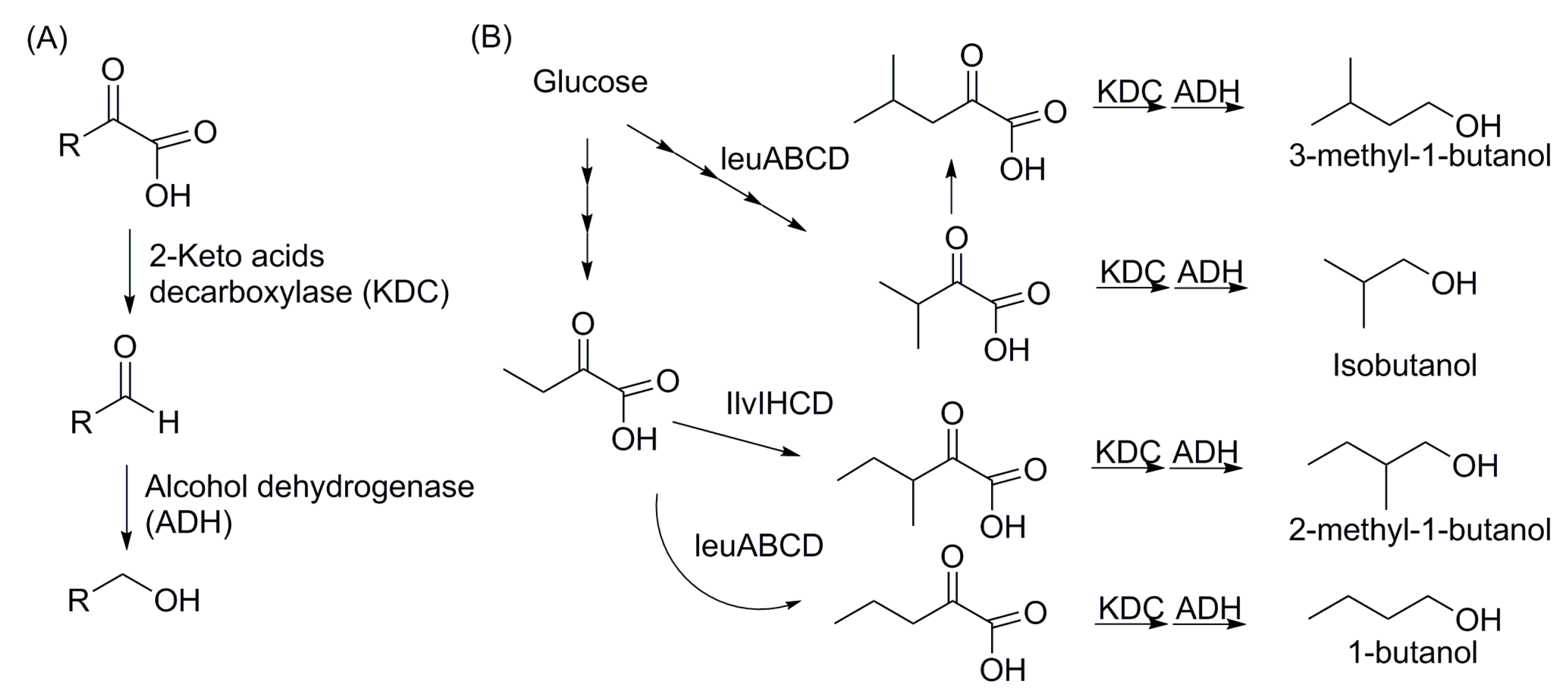
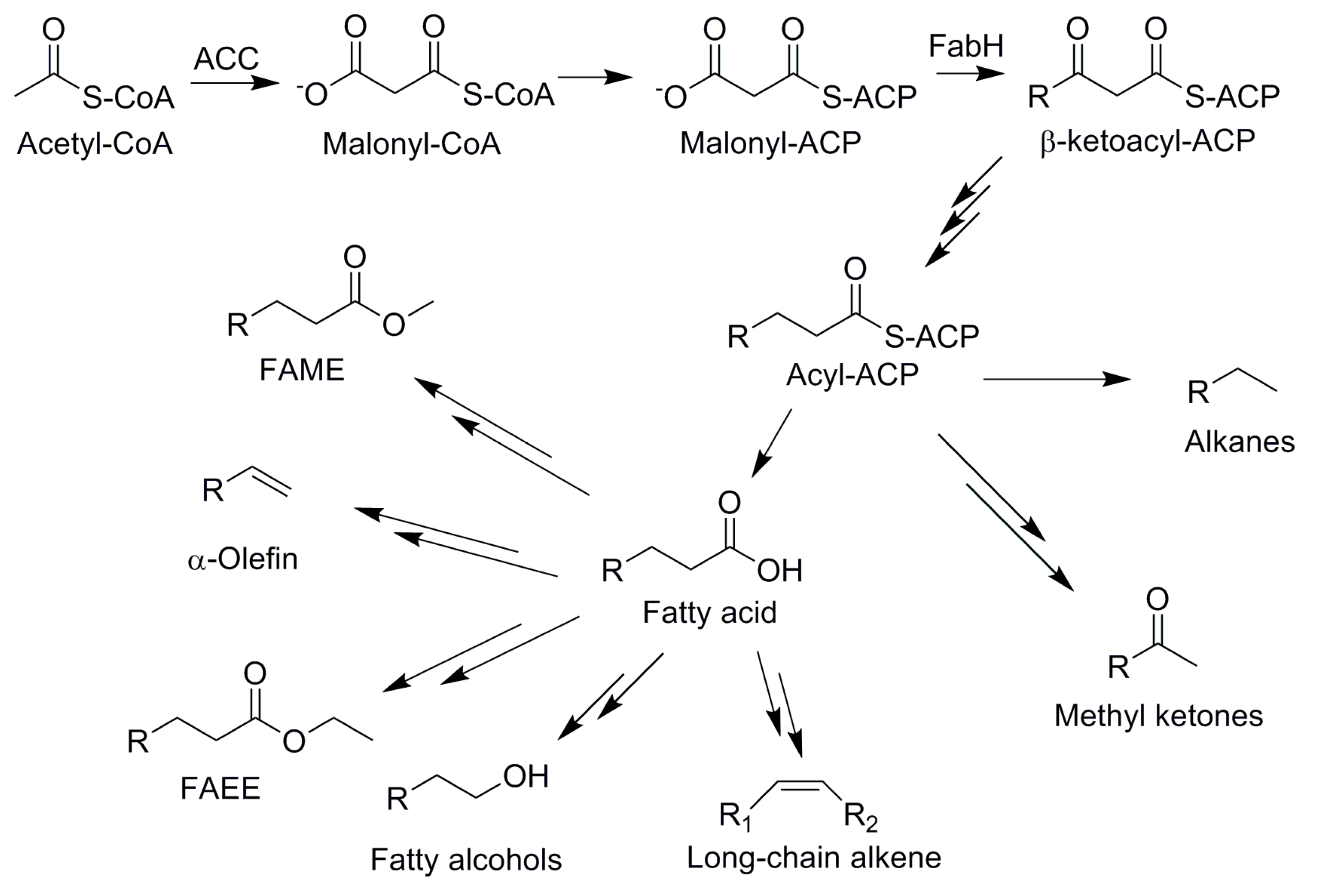
4.3. Fatty Acid-Based Biofuels
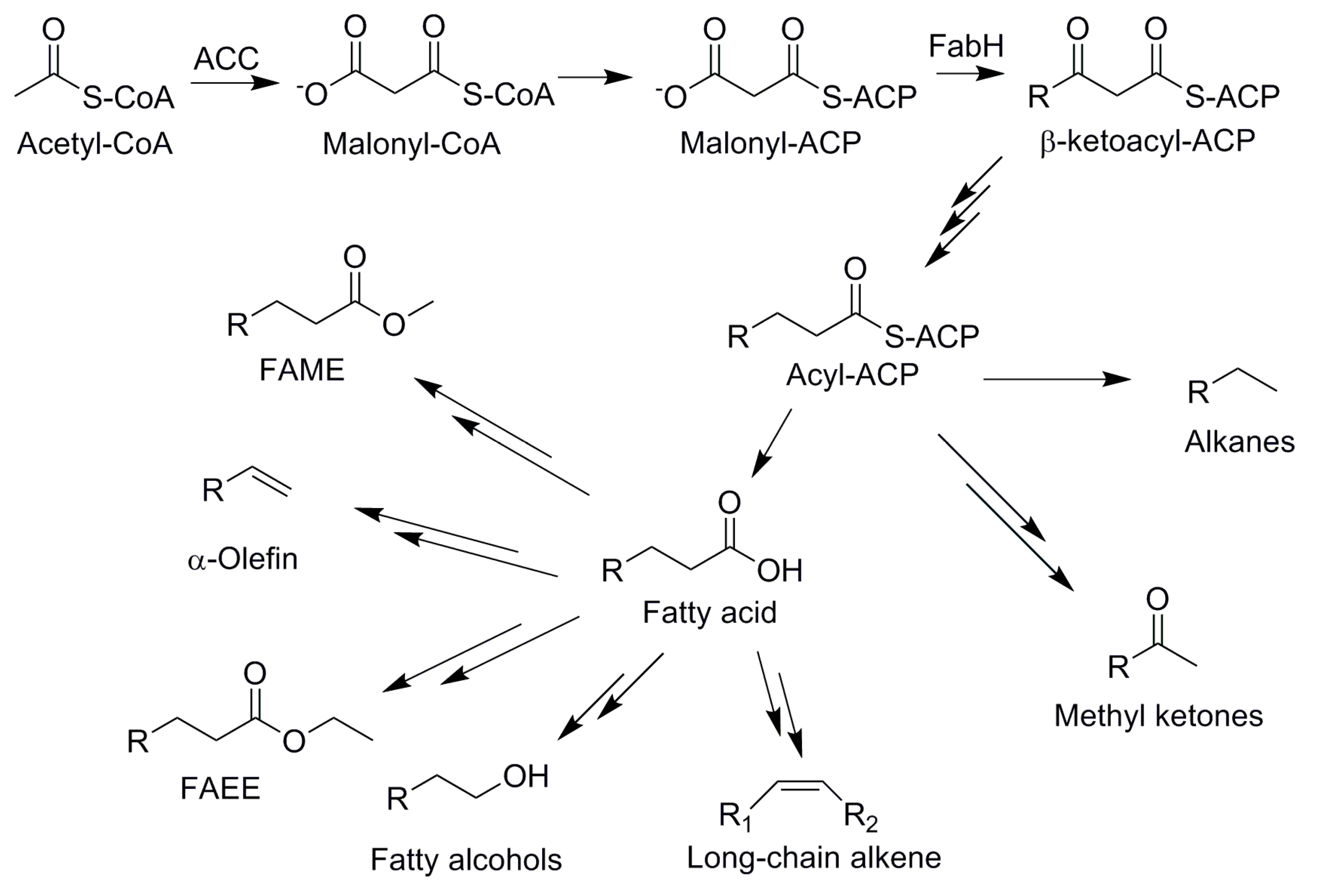
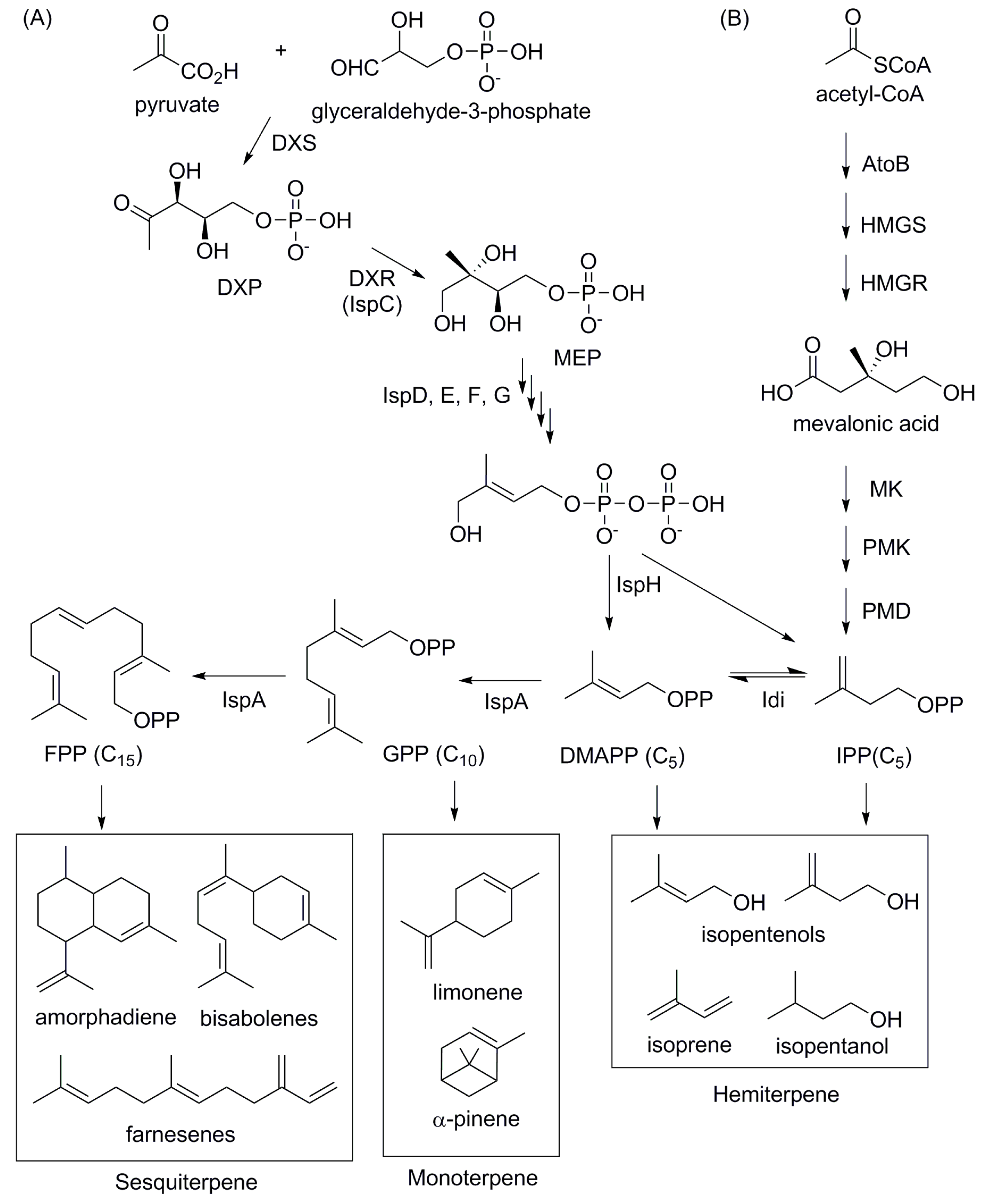
4.4. Isoprenoid-Based Biofuels
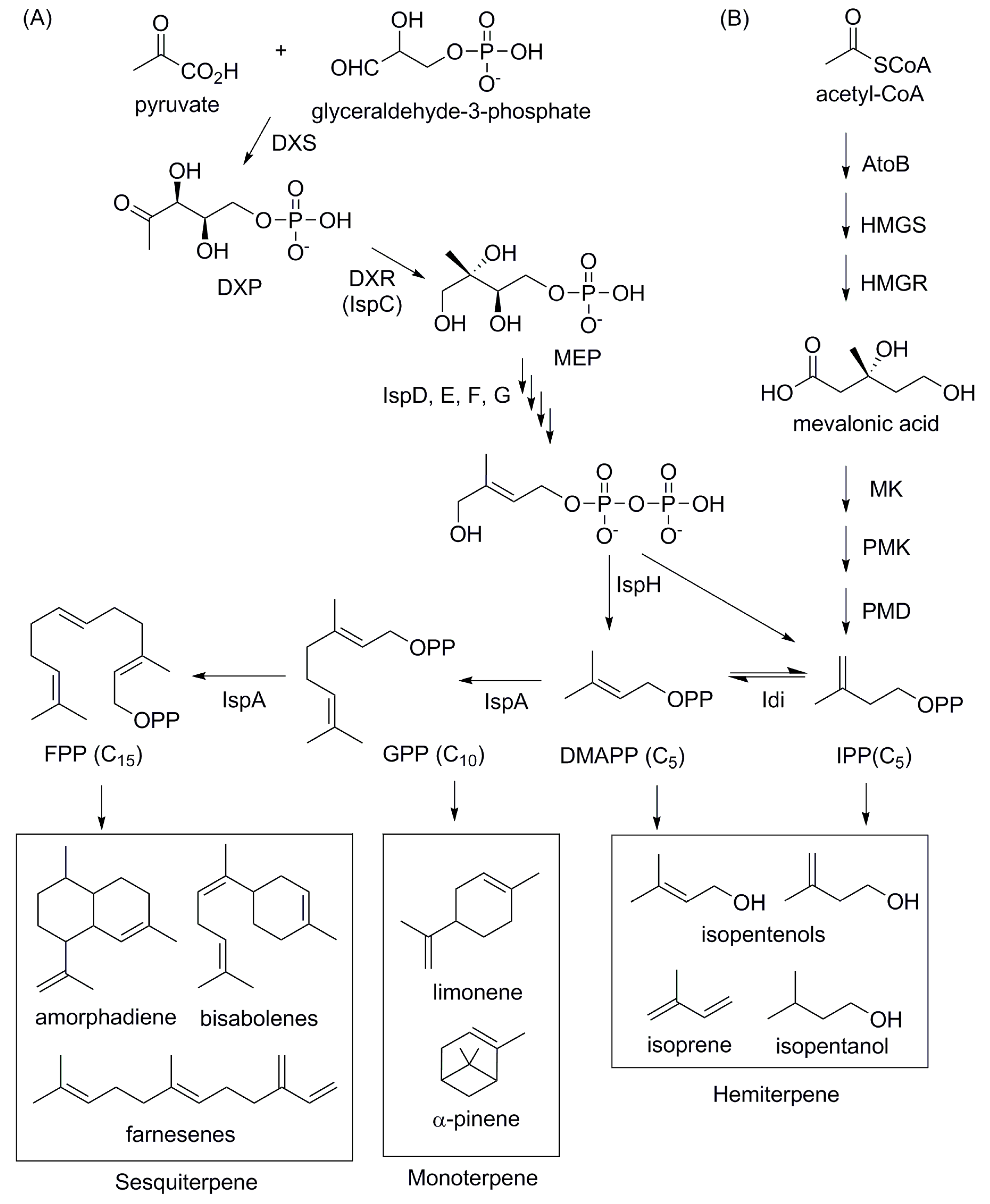
4.5. Advanced Biofuels Production from LCB-Derived Sugars or Hydrolysates
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Geddes, C.C.; Nieves, I.U.; Ingram, L.O. Advances in ethanol production. Curr. Opin. Biotechnol. 2011, 22, 312–319. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ingram, L.O.; Conway, T.; Clark, D.P.; Sewell, G.W.; Preston, J.F. Genetic engineering of ethanol production in Escherichia coli. Appl. Environ. Microbiol. 1987, 53, 2420–2425. [Google Scholar] [PubMed]
- Bai, F.W.; Anderson, W.A.; Moo-Young, M. Ethanol fermentation technologies from sugar and starch feedstocks. Biotechnol. Adv. 2008, 26, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, G. Carbohydrate metabolism in Zymomonas mobilis: A catabolic highway with some scenic routes. FEMS Microbiol. Lett. 1996, 145, 301–307. [Google Scholar] [CrossRef]
- Flamholz, A.; Noor, E.; Bar-Even, A.; Liebermeister, W.; Milo, R. Glycolytic strategy as a tradeoff between energy yield and protein cost. Proc. Natl. Acad. Sci. USA 2013, 110, 10039–10044. [Google Scholar] [CrossRef] [PubMed]
- Flamholz, A.; Noor, E.; Bar-Even, A.; Milo, R. eQuilibrator—The biochemical thermodynamics calculator. Nucleic Acids Res. 2012, 40, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.; Farwick, A.; Benisch, F.; Brat, D.; Dietz, H.; Subtil, T.; Boles, E. Trends and challenges in the microbial production of lignocellulosic bioalcohol fuels. Appl. Microbiol. Biotechnol. 2010, 87, 1303–1315. [Google Scholar] [CrossRef] [PubMed]
- Yanase, H.; Miyawaki, H.; Sakurai, M.; Kawakami, A.; Matsumoto, M.; Haga, K.; Kojima, M.; Okamoto, K. Ethanol production from wood hydrolysate using genetically engineered Zymomonas mobilis. Appl. Microbiol. Biotechnol. 2012, 94, 1667–1678. [Google Scholar] [CrossRef] [PubMed]
- Lütke-Eversloh, T.; Bahl, H. Metabolic engineering of Clostridium acetobutylicum: Recent advances to improve butanol production. Curr. Opin. Biotechnol. 2011, 22, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, W. Factors affecting utilization of carbohydrates by clostridia. FEMS Microbiol. Rev. 1995, 17, 317–329. [Google Scholar] [CrossRef]
- Papoutsakis, E.T. Engineering solventogenic clostridia. Curr. Opin. Biotechnol. 2008, 19, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Basso, T.O.; de Kok, S.; Dario, M.; do Espirito-Santo, J.C.; Müller, G.; Schlölg, P.S.; Silva, C.P.; Tonso, A.; Daran, J.-M.M.; Gombert, A.K.; et al. Engineering topology and kinetics of sucrose metabolism in Saccharomyces cerevisiae for improved ethanol yield. Metab. Eng. 2011, 13, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Larsson, C.; van Maris, A.; Pronk, J. Metabolic engineering of yeast for production of fuels and chemicals. Curr. Opin. Biotechnol. 2013, 24, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Young, E.M.; Comer, A.D.; Huang, H.; Alper, H.S. A molecular transporter engineering approach to improving xylose catabolism in Saccharomyces cerevisiae. Metab. Eng. 2012, 14, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Young, E.M.; Tong, A.; Bui, H.; Spofford, C.; Alper, H.S. Rewiring yeast sugar transporter preference through modifying a conserved protein motif. Proc. Natl. Acad. Sci. USA 2014, 111, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Farwick, A.; Bruder, S.; Schadeweg, V.; Oreb, M.; Boles, E. Engineering of yeast hexose transporters to transport D-xylose without inhibition by D-glucose. Proc. Natl. Acad. Sci. USA 2014, 111, 5159–5164. [Google Scholar] [CrossRef] [PubMed]
- Trinh, C.T.; Srienc, F. Metabolic engineering of Escherichia coli for efficient conversion of glycerol to ethanol. Appl. Environ. Microbiol. 2009, 75, 6696–6705. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Chi, W.-J.; Hong, S.-K.; Yang, J.-W.; Chang, Y.K. Bioethanol production by heterologous expression of Pdc and AdhII in Streptomyces lividans. Appl. Microbiol. Biotechnol. 2013, 97, 6089–6097. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.C.; Zhang, Q.; Lyne, P.W.L. Ethanol-diesel fuel blends—A review. Bioresour. Technol. 2005, 96, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.R.; Lan, E.I.; Dekishima, Y.; Baez, A.; Cho, K.M.; Liao, J.C. Driving forces enable high-titer anaerobic 1-butanol synthesis in Escherichia coli. Appl. Environ. Microbiol. 2011, 77, 2905–2915. [Google Scholar] [CrossRef] [PubMed]
- Baez, A.; Cho, K.M.; Liao, J.C. High-flux isobutanol production using engineered Escherichia coli: A bioreactor study with in situ product removal. Appl. Microbiol. Biotechnol. 2011, 90, 1681–1690. [Google Scholar] [CrossRef] [PubMed]
- Connor, M.R.; Cann, A.F.; Liao, J.C. 3-Methyl-1-butanol production in Escherichia coli: Random mutagenesis and two-phase fermentation. Appl. Microbiol. Biotechnol. 2010, 86, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- George, K.W.; Thompson, M.G.; Kang, A.; Baidoo, E.; Wang, G.; Chan, L.J.G.; Adams, P.D.; Petzold, C.J.; Keasling, J.D.; Lee, T.S. Metabolic engineering for the high-yield production of isoprenoid-based C5 alcohols in E. coli. Sci. Rep. 2015, 5, 11128. [Google Scholar] [CrossRef] [PubMed]
- Cann, A.F.; Liao, J.C. Production of 2-methyl-1-butanol in engineered Escherichia coli. Appl. Microbiol. Biotechnol. 2008, 81, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Rude, M.A.; Schirmer, A. New microbial fuels: A biotech perspective. Curr. Opin. Microbiol. 2009, 12, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Gutierrez, J.; Kim, E.-M.; Batth, T.S.; Cho, N.; Hu, Q.; Chan, L.J.G.; Petzold, C.J.; Hillson, N.J.; Adams, P.D.; Keasling, J.D.; et al. Principal component analysis of proteomics (PCAP) as a tool to direct metabolic engineering. Metab. Eng. 2015, 28, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Sarria, S.; Wong, B.; García Martín, H.; Keasling, J.D.; Peralta-Yahya, P.P. Microbial synthesis of pinene. ACS Synth. Biol. 2014, 3, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Carothers, J.M.; Keasling, J.D. Design of a dynamic sensor-regulator system for production of chemicals and fuels derived from fatty acids. Nat. Biotechnol. 2012, 30, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Goh, E.-B.; Baidoo, E.E.K.; Burd, H.; Lee, T.S.; Keasling, J.D.; Beller, H.R. Substantial improvements in methyl ketone production in E. coli and insights on the pathway from in vitro studies. Metab. Eng. 2014, 26, 67–76. [Google Scholar] [PubMed]
- Dugar, D.; Stephanopoulos, G. Relative potential of biosynthetic pathways for biofuels and bio-based products. Nat. Biotechnol. 2011, 29, 1074–1078. [Google Scholar] [CrossRef] [PubMed]
- Patakova, P.; Linhova, M.; Rychtera, M.; Paulova, L.; Melzoch, K. Novel and neglected issues of acetone-butanol-ethanol (ABE) fermentation by clostridia: Clostridium metabolic diversity, tools for process mapping and continuous fermentation systems. Biotechnol. Adv. 2013, 31, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Ezeji, T.; Milne, C.; Price, N.D.; Blaschek, H.P. Achievements and perspectives to overcome the poor solvent resistance in acetone and butanol-producing microorganisms. Appl. Microbiol. Biotechnol. 2010, 85, 1697–1712. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Cann, A.F.; Connor, M.R.; Shen, C.R.; Smith, K.M.; Brynildsen, M.P.; Chou, K.J.Y.; Hanai, T.; Liao, J.C. Metabolic engineering of Escherichia coli for 1-butanol production. Metab. Eng. 2008, 10, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Bond-Watts, B.B.; Bellerose, R.J.; Chang, M.C.Y. Enzyme mechanism as a kinetic control element for designing synthetic biofuel pathways. Nat. Chem. Biol. 2011, 7, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Seo, S.W.; Kim, S.Y.; Jung, G.Y. Model-driven rebalancing of the intracellular redox state for optimization of a heterologous n-butanol pathway in Escherichia coli. Metab. Eng. 2013, 20, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.A.; Boyarskiy, S.; Yamada, M.R.; Kong, N.; Bauer, S.; Tullman-Ercek, D. Enhancing tolerance to short-chain alcohols by engineering the Escherichia coli AcrB efflux pump to secrete the non-native substrate n-butanol. ACS Synth. Biol. 2014, 3, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Inokuma, K.; Liao, J.C.; Okamoto, M.; Hanai, T. Improvement of isopropanol production by metabolically engineered Escherichia coli using gas stripping. J. Biosci. Bioeng. 2010, 110, 696–701. [Google Scholar] [CrossRef] [PubMed]
- Lamsen, E.N.; Atsumi, S. Recent progress in synthetic biology for microbial production of C3-C10 alcohols. Front. Microbiol. 2012, 3, 196. [Google Scholar] [CrossRef]
- Gronenberg, L.S.; Marcheschi, R.J.; Liao, J.C. Next generation biofuel engineering in prokaryotes. Curr. Opin. Chem. Biol. 2013, 17, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Hanai, T.; Liao, J.C. Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 2008, 451, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Marcheschi, R.J.; Li, H.; Zhang, K.; Noey, E.L.; Kim, S.; Chaubey, A.; Houk, K.N.; Liao, J.C. A synthetic recursive “+1” pathway for carbon chain elongation. ACS Chem. Biol. 2012, 7, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Avalos, J.L.; Fink, G.R.; Stephanopoulos, G. Compartmentalization of metabolic pathways in yeast mitochondria improves the production of branched-chain alcohols. Nat. Biotechnol. 2013, 31, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.-C.; Prather, K.L.J. Controlled biosynthesis of odd-chain fuels and chemicals via engineered modular metabolic pathways. Proc. Natl. Acad. Sci. USA 2012, 109, 17925–17930. [Google Scholar] [CrossRef] [PubMed]
- Kalscheuer, R.; Steinbüchel, A. A novel bifunctional wax ester synthase/acyl-CoA:diacylglycerol acyltransferase mediates wax ester and triacylglycerol biosynthesis in Acinetobacter calcoaceticus ADP1. J. Biol. Chem. 2003, 278, 8075–8082. [Google Scholar] [CrossRef] [PubMed]
- Kalscheuer, R.; Stölting, T.; Steinbüchel, A. Microdiesel: Escherichia coli engineered for fuel production. Microbiology 2006, 152, 2529–2536. [Google Scholar] [CrossRef] [PubMed]
- Steen, E.J.; Kang, Y.; Bokinsky, G.; Hu, Z.; Schirmer, A.; McClure, A.; del Cardayre, S.B.; Keasling, J.D. Microbial production of fatty-acid-derived fuels and chemicals from plant biomass. Nature 2010, 463, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Runguphan, W.; Keasling, J.D. Metabolic engineering of Saccharomyces cerevisiae for production of fatty acid-derived biofuels and chemicals. Metab. Eng. 2014, 21, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Tai, M.; Stephanopoulos, G. Engineering the push and pull of lipid biosynthesis in oleaginous yeast Yarrowia lipolytica for biofuel production. Metab. Eng. 2013, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lennen, R.M.; Pfleger, B.F. Microbial production of fatty acid-derived fuels and chemicals. Curr. Opin. Biotechnol. 2013, 24, 1044–1053. [Google Scholar] [CrossRef] [PubMed]
- Beller, H.R.; Lee, T.S.; Katz, L. Natural products as biofuels and bio-based chemicals: Fatty acids and isoprenoids. Nat. Prod. Rep. 2015, 32, 1508–1526. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Chang, W.-C.; Warui, D.M.; Booker, S.J.; Krebs, C.; Bollinger, J.M. Evidence for only oxygenative cleavage of aldehydes to alk(a/e)nes and formate by cyanobacterial aldehyde decarbonylases. Biochemistry 2012, 51, 7908–7916. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, A.; Rude, M.A.; Li, X.; Popova, E.; del Cardayre, S.B. Microbial biosynthesis of alkanes. Science 2010, 329, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Perez, D.; Begemann, M.B.; Pfleger, B.F. Modular synthase-encoding gene involved in α-olefin biosynthesis in Synechococcus sp. strain PCC 7002. Appl. Environ. Microbiol. 2011, 77, 4264–4267. [Google Scholar] [CrossRef] [PubMed]
- Rude, M.A.; Baron, T.S.; Brubaker, S.; Alibhai, M.; Del Cardayre, S.B.; Schirmer, A. Terminal olefin (1-alkene) biosynthesis by a novel p450 fatty acid decarboxylase from Jeotgalicoccus species. Appl. Environ. Microbiol. 2011, 77, 1718–1727. [Google Scholar] [CrossRef] [PubMed]
- Willis, R.M.; Wahlen, B.D.; Seefeldt, L.C.; Barney, B.M. Characterization of a fatty acyl-CoA reductase from Marinobacter aquaeolei VT8: A bacterial enzyme catalyzing the reduction of fatty acyl-CoA to fatty alcohol. Biochemistry 2011, 50, 10550–10558. [Google Scholar] [CrossRef] [PubMed]
- Goh, E.-B.; Baidoo, E.E.K.; Keasling, J.D.; Beller, H.R. Engineering of bacterial methyl ketone synthesis for biofuels. Appl. Environ. Microbiol. 2012, 78, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Dellomonaco, C.; Clomburg, J.M.; Miller, E.N.; Gonzalez, R. Engineered reversal of the β-oxidation cycle for the synthesis of fuels and chemicals. Nature 2011, 476, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.K.; Turner, N.J.; Jones, P.R. Carboxylic acid reductase is a versatile enzyme for the conversion of fatty acids into fuels and chemical commodities. Proc. Natl. Acad. Sci. USA 2013, 110, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Howard, T.P.; Middelhaufe, S.; Moore, K.; Edner, C.; Kolak, D.M.; Taylor, G.N.; Parker, D.A.; Lee, R.; Smirnoff, N.; Aves, S.J.; et al. Synthesis of customized petroleum-replica fuel molecules by targeted modification of free fatty acid pools in Escherichia coli. Proc. Natl. Acad. Sci. USA 2013, 110, 7636–7641. [Google Scholar] [CrossRef] [PubMed]
- Haushalter, R.W.; Kim, W.; Chavkin, T.A.; The, L.; Garber, M.E.; Nhan, M.; Adams, P.D.; Petzold, C.J.; Katz, L.; Keasling, J.D. Production of anteiso-branched fatty acids in Escherichia coli; next generation biofuels with improved cold-flow properties. Metab. Eng. 2014, 26, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Yahya, P.P.; Zhang, F.; del Cardayre, S.B.; Keasling, J.D. Microbial engineering for the production of advanced biofuels. Nature 2012, 488, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-concepción, M. Plant Isoprenoids. Methods Mol. Biol. 2014, 1153, 1–5. [Google Scholar] [PubMed]
- Wang, K.; Ohnuma, S. Chain-length determination mechanism of isoprenyl diphosphate synthases and implications for molecular evolution. Trends Biochem. Sci. 1999, 24, 445–451. [Google Scholar] [CrossRef]
- Degenhardt, J.; Köllner, T.G.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef] [PubMed]
- Renninger, N.; McPhee, D. Fuel compositions comprising farnesane and method of making and using same. U.S. Patent 20080098645 A1, 1 May 2008. Available online: https://www.google.com/patents/US20080098645 (accessed on 22 October 2015). [Google Scholar]
- Özaydın, B.; Burd, H.; Lee, T.S.; Keasling, J.D. Carotenoid-based phenotypic screen of the yeast deletion collection reveals new genes with roles in isoprenoid production. Metab. Eng. 2013, 15, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Yahya, P.P.; Ouellet, M.; Chan, R.; Mukhopadhyay, A.; Keasling, J.D.; Lee, T.S. Identification and microbial production of a terpene-based advanced biofuel. Nat. Commun. 2011, 2, 483. [Google Scholar] [CrossRef] [PubMed]
- Bokinsky, G.; Peralta-Yahya, P.P.; George, A.; Holmes, B.M.; Steen, E.J.; Dietrich, J.; Lee, T.S.; Tullman-Ercek, D.; Voigt, C.; Simmons, B.; et al. Synthesis of three advanced biofuels from ionic liquid-pretreated switchgrass using engineered Escherichia coli. Proc. Natl. Acad. Sci. USA 2011, 108, 19949–19954. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Gutierrez, J.; Chan, R.; Batth, T.S.; Adams, P.D.; Keasling, J.D.; Petzold, C.J.; Lee, T.S. Metabolic engineering of Escherichia coli for limonene and perillyl alcohol production. Metab. Eng. 2013, 19, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Redding-Johanson, A.M.; Batth, T.S.; Chan, R.; Krupa, R.; Szmidt, H.L.; Adams, P.D.; Keasling, J.D.; Lee, T.S.; Mukhopadhyay, A.; Petzold, C.J. Targeted proteomics for metabolic pathway optimization: Application to terpene production. Metab. Eng. 2011, 13, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.M.; Garcia, D.E.; Redding-Johanson, A.M.; Friedland, G.D.; Chan, R.; Batth, T.S.; Haliburton, J.R.; Chivian, D.; Keasling, J.D.; Petzold, C.J.; et al. Optimization of a heterologous mevalonate pathway through the use of variant HMG-CoA reductases. Metab. Eng. 2011, 13, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Renninger, N.; McPhee, D. Petroleum Component, Fuel Additive and Microorganism Produced Isoprenoids; Conversion of Simple Sugars, Polysaccharides and/or Nonfermentable Carbon Sources; Diesel Fuel, Jet Fuel, Kerosene or Gasoline Replacements. US20080098645, 2008. Available online: https://www.google.com.ar/patents/US20080098645 (accessed on 26 October 2015). [Google Scholar]
- Wang, C.; Yoon, S.-H.; Jang, H.-J.; Chung, Y.-R.; Kim, J.-Y.; Choi, E.-S.; Kim, S.-W. Metabolic engineering of Escherichia coli for α-farnesene production. Metab. Eng. 2011, 13, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Pray, T. Biomass R&D Technical Advisory Committee: Drop-in Fuels Panel; Amyris: Emeryville, CA, USA, 2010. [Google Scholar]
- Ro, D.K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 2006, 440, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Tracy, N.I.; Chen, D.; Crunkleton, D.W.; Price, G.L. Hydrogenated monoterpenes as diesel fuel additives. Fuel 2009, 88, 2238–2240. [Google Scholar] [CrossRef]
- Carter, O.A.; Peters, R.J.; Croteau, R. Monoterpene biosynthesis pathway construction in Escherichia coli. Phytochemistry 2003, 64, 425–433. [Google Scholar] [PubMed]
- Yang, J.; Nie, Q.; Ren, M.; Feng, H.; Jiang, X.; Zheng, Y.; Liu, M.; Zhang, H.; Xian, M. Metabolic engineering of Escherichia coli for the biosynthesis of alpha-pinene. Biotechnol. Biofuels 2013, 6, 60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, Q.; Cao, Y.; Feng, X.; Zheng, Y.; Zou, H.; Liu, H.; Yang, J.; Xian, M. Microbial production of sabinene—A new terpene-based precursor of advanced biofuel. Microb. Cell Fact. 2014, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.H.; Keasling, J.D. Synthetic pathway for production of five-carbon alcohols from isopentenyl diphosphate. Appl. Environ. Microbiol. 2012, 78, 7849–7855. [Google Scholar] [CrossRef] [PubMed]
- George, K.W.; Chen, A.; Jain, A.; Batth, T.S.; Baidoo, E.E.K.; Wang, G.; Adams, P.D.; Petzold, C.J.; Keasling, J.D.; Lee, T.S. Correlation analysis of targeted proteins and metabolites to assess and engineer microbial isopentenol production. Biotechnol. Bioeng. 2014, 111, 1648–1658. [Google Scholar] [PubMed]
- Foo, J.L.; Jensen, H.M.; Dahl, R.H.; George, K.W.; Keasling, J.D.; Lee, T.S.; Leong, S.; Mukhopadhyay, A. Improving Microbial Biogasoline Production in Escherichia coli Using Tolerance Engineering. MBio 2014, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, C.; Yoon, S.H.; Jang, H.J.; Choi, E.S.; Kim, S.W. Engineering Escherichia coli for selective geraniol production with minimized endogenous dehydrogenation. J. Biotechnol. 2014, 169, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Balan, V. Current challenges in commercially producing biofuels from lignocellulosic biomass. ISRN Biotechnol. 2014, 2014, 31. [Google Scholar]
- Shi, J.; George, K.W.; Sun, N.; He, W.; Li, C.; Stavila, V.; Keasling, J.D.; Simmons, B.A.; Lee, T.S.; Singh, S. Impact of pretreatment technologies on saccharification and isopentenol fermentation of mixed lignocellulosic feedstocks. BioEnergy Res. 2015, 8, 1004–1013. [Google Scholar] [CrossRef]
- Akita, H.; Nakashima, N.; Hoshino, T. Bacterial production of isobutanol without expensive reagents. Appl. Microbiol. Biotechnol. 2015, 99, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Jiang, J.; Lu, Q.; Zhao, Z.; Xie, T.; Zhao, H.; Wang, M. Engineering Corynebacterium crenatum to produce higher alcohols for biofuel using hydrolysates of duckweed (Landoltia punctata) as feedstock. Microb. Cell Fact. 2015, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.H.; Rabinovitch-Deere, C.A.; Fan, Z.; Atsumi, S. Isobutanol production from cellobionic acid in Escherichia coli. Microb. Cell Fact. 2015, 14, 52. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, A.; Lee, T.S. Converting Sugars to Biofuels: Ethanol and Beyond. Bioengineering 2015, 2, 184-203. https://doi.org/10.3390/bioengineering2040184
Kang A, Lee TS. Converting Sugars to Biofuels: Ethanol and Beyond. Bioengineering. 2015; 2(4):184-203. https://doi.org/10.3390/bioengineering2040184
Chicago/Turabian StyleKang, Aram, and Taek Soon Lee. 2015. "Converting Sugars to Biofuels: Ethanol and Beyond" Bioengineering 2, no. 4: 184-203. https://doi.org/10.3390/bioengineering2040184
APA StyleKang, A., & Lee, T. S. (2015). Converting Sugars to Biofuels: Ethanol and Beyond. Bioengineering, 2(4), 184-203. https://doi.org/10.3390/bioengineering2040184