Betulin Stimulates Osteogenic Differentiation of Human Osteoblasts-Loaded Alginate–Gelatin Microbeads



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture Experiments
2.3. Fabrication of Cell-Loaded Microbeads
2.4. Cell Proliferation and Metabolic Activity
2.5. Live and Dead Assay
2.6. Concanavalin A–Alexafluor (ConA-AF488) and DAPI Staining
2.7. Osteopontin Production Assay
2.8. Dimethylmethylene Blue Assay (DMMB)
2.9. Alizarin Red S Staining
2.10. Alkaline Phosphatase Activity
2.11. Statistical Analyses
3. Results
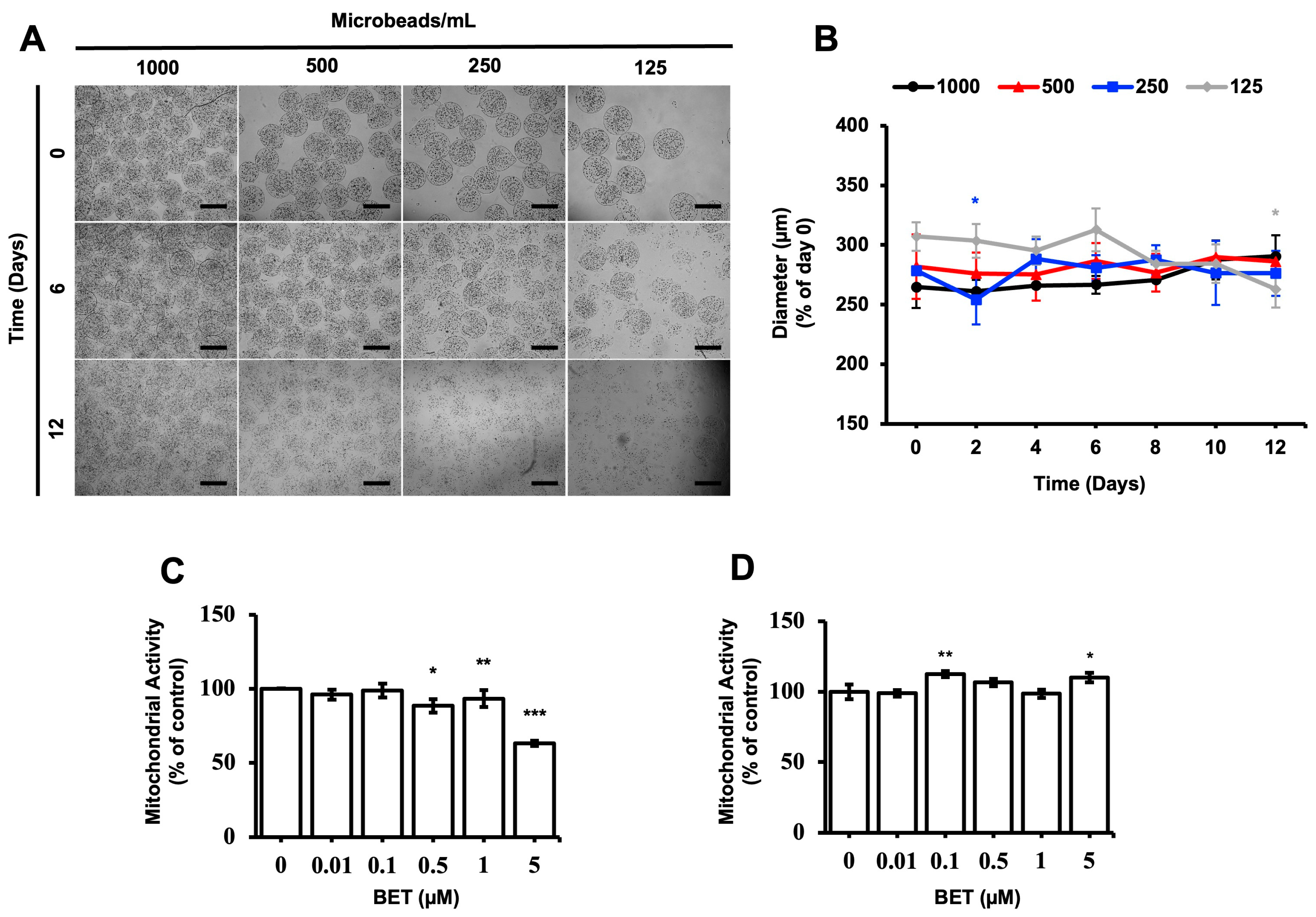
3.1. Morphological Evaluation of hFOB 1.19 Cell-Loaded Microbeads
3.2. Effect of BET on the Metabolic Activity of hFOB 1.19 Cells
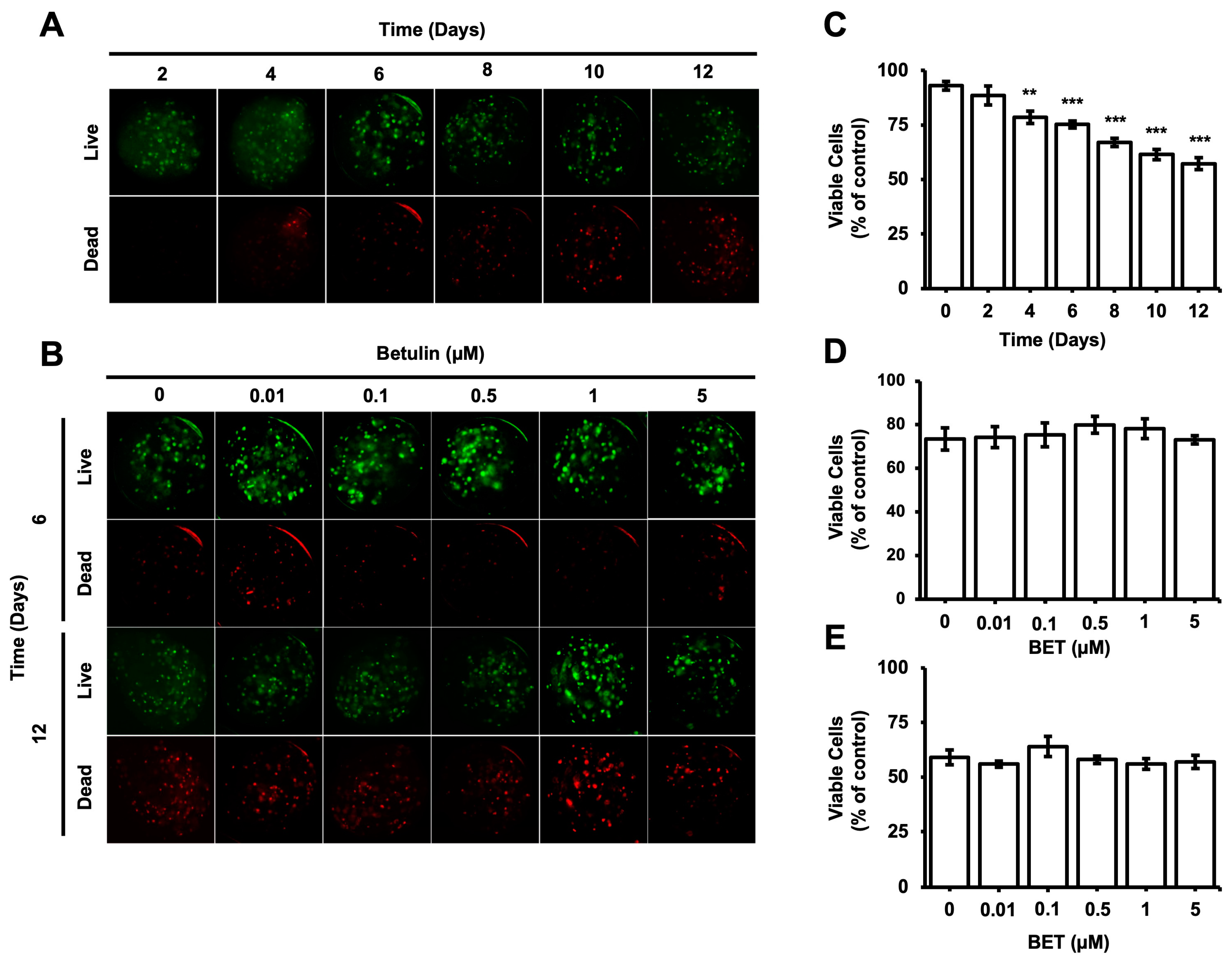
3.3. Effect of BET on Cell Viability of hFOB 1.19 Cell-Loaded AlGel Microbeads
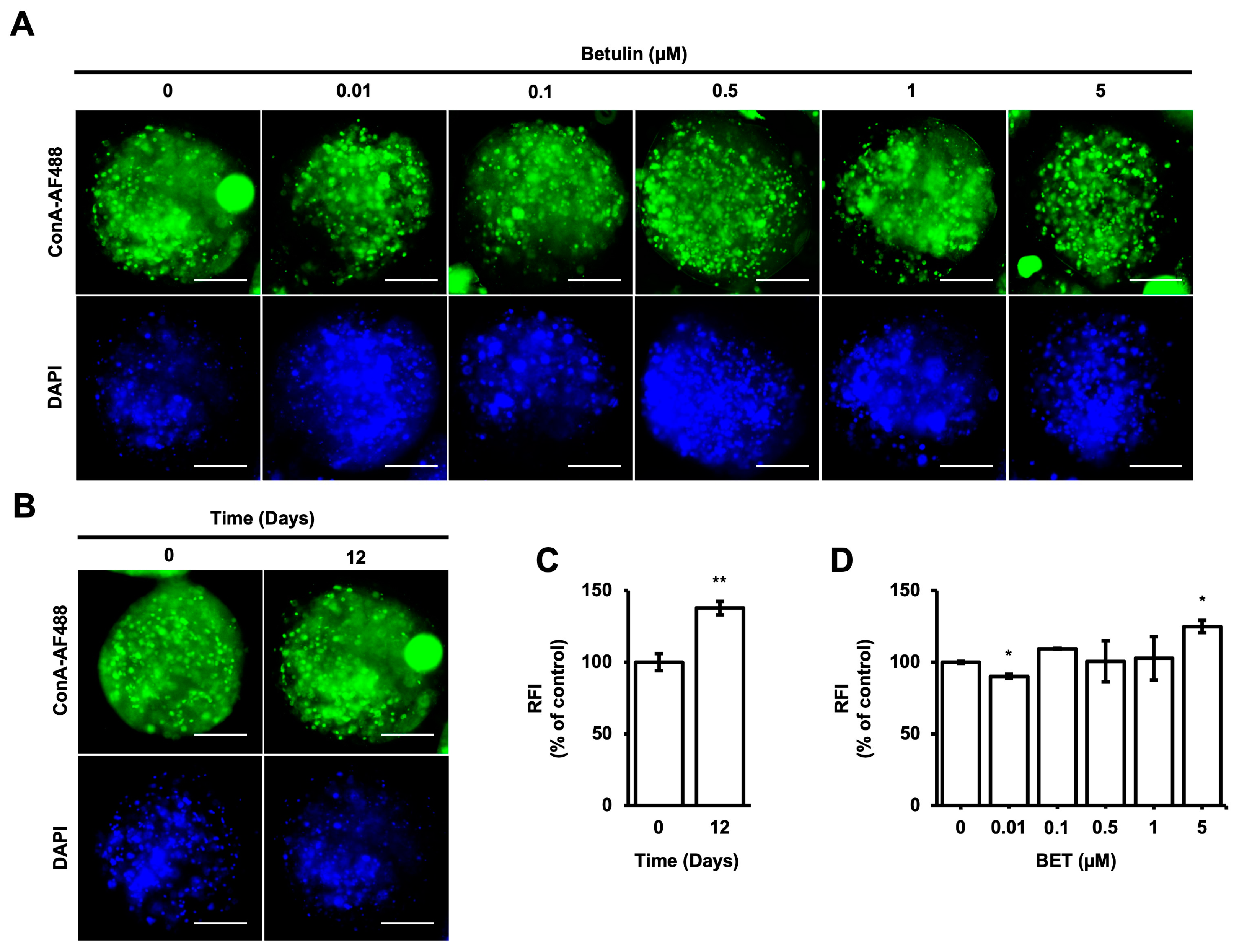
3.4. Alteration of High Mannose-Type Glycans (HM) Levels in Microbeads
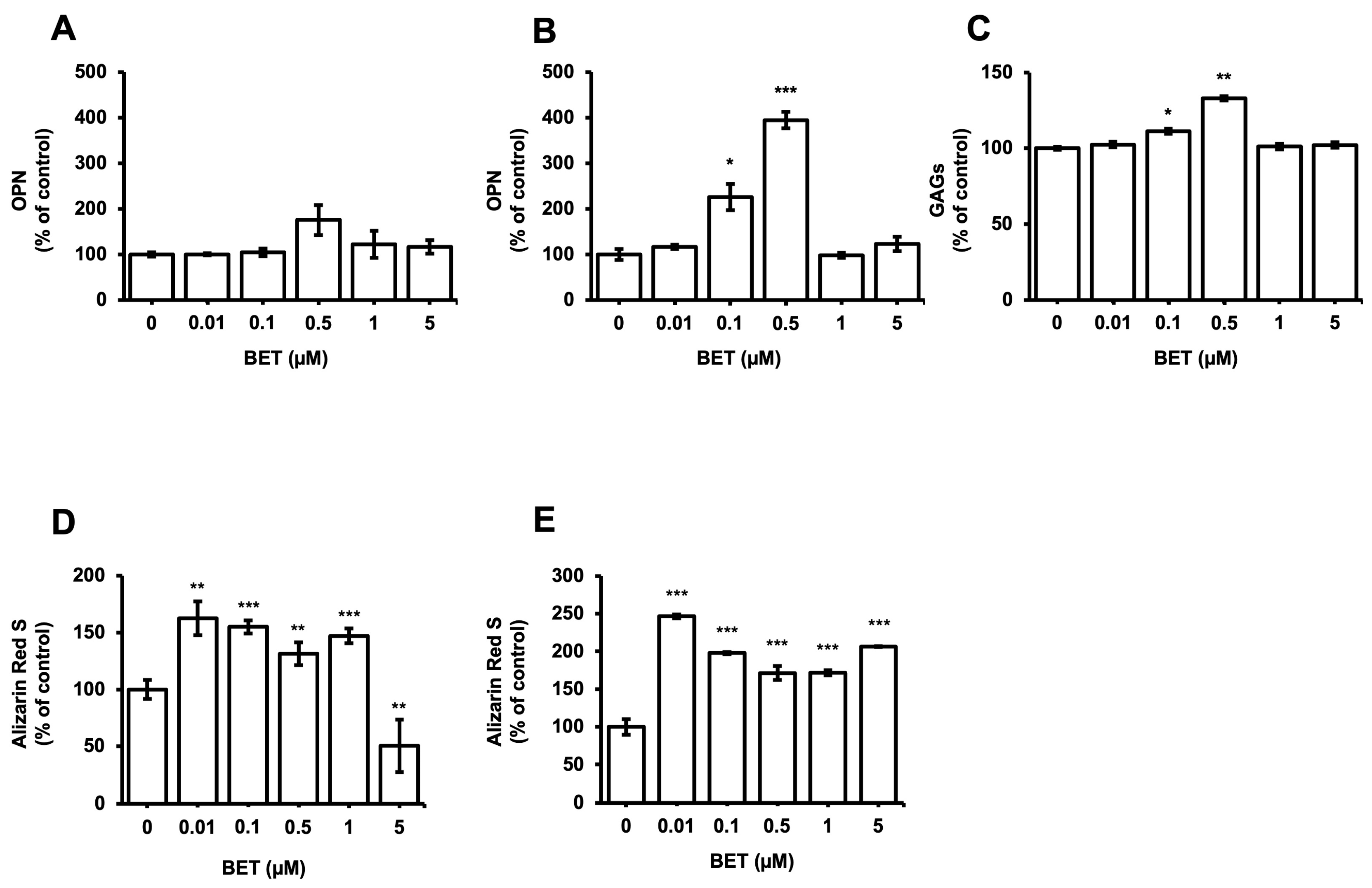
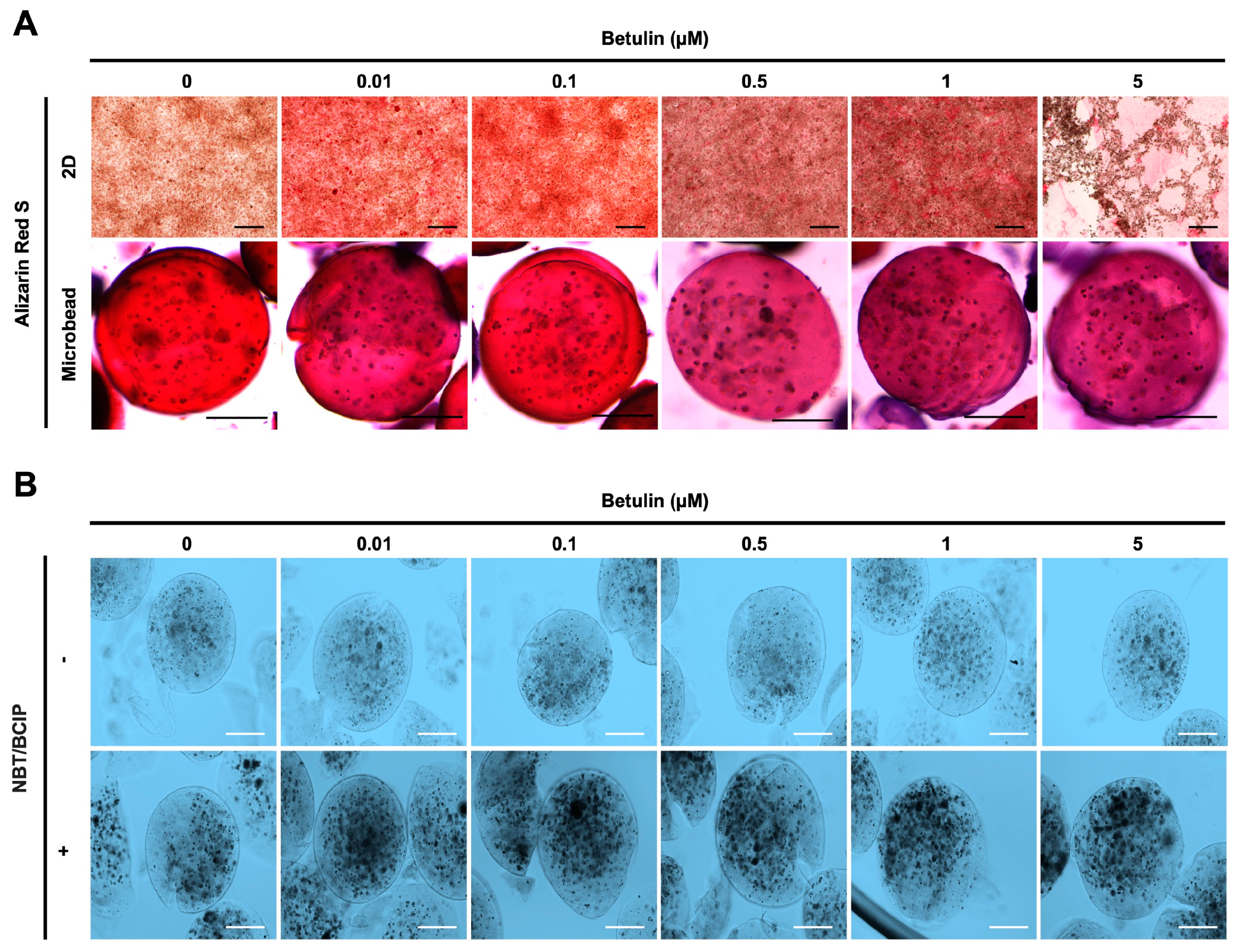
3.5. Influence of BET on the Expression of Osteogenic Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andric, T.; Sampson, A.C.; Freeman, J.W. Fabrication and Characterization of Electrospun Osteon Mimicking Scaffolds for Bone Tissue Engineering. Mater. Sci. Eng. C 2011, 31, 2–8. [Google Scholar] [CrossRef]
- Florencio-Silva, R.; Sasso, G.R.D.S.; Sasso-Cerri, E.; Simões, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. Biomed. Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef]
- Skjødt, M.K.; Frost, M.; Abrahamsen, B. Side Effects of Drugs for Osteoporosis and Metastatic Bone Disease. Br. J. Clin. Pharmacol. 2019, 85, 1063–1071. [Google Scholar] [CrossRef]
- Sen, M.K.; Miclau, T. Autologous Iliac Crest Bone Graft: Should It Still Be the Gold Standard for Treating Nonunions? Injury 2007, 38, S75–S80. [Google Scholar] [CrossRef]
- PerićKačarević, Ž.; Rider, P.; Alkildani, S.; Retnasingh, S.; Pejakić, M.; Schnettler, R.; Gosau, M.; Smeets, R.; Jung, O.; Barbeck, M. An Introduction to Bone Tissue Engineering. Int. J. Artif. Organs 2020, 43, 69–86. [Google Scholar] [CrossRef]
- Nabavinia, M.; Khoshfetrat, A.B.; Naderi-Meshkin, H. Nano-Hydroxyapatite-Alginate-Gelatin Microcapsule as a Potential Osteogenic Building Block for Modular Bone Tissue Engineering. Mater. Sci. Eng. C 2019, 97, 67–77. [Google Scholar] [CrossRef]
- Rocca, D.G.D.; Willenberg, B.J.; Qi, Y.; Simmons, C.S.; Rubiano, A.; Ferreira, L.F.; Huo, T.; Petersen, J.W.; Ruchaya, P.J.; Wate, P.S.; et al. An Injectable Capillary-like Microstructured Alginate Hydrogel Improves Left Ventricular Function after Myocardial Infarction in Rats. Int. J. Cardiol. 2016, 220, 149–154. [Google Scholar] [CrossRef]
- Sukegawa, A.; Iwasaki, N.; Kasahara, Y.; Onodera, T.; Igarashi, T.; Minami, A. Repair of Rabbit Osteochondral Defects by an Acellular Technique with an Ultrapurified Alginate Gel Containing Stromal Cell-Derived Factor-1. Tissue Eng. Part A 2012, 18, 934–945. [Google Scholar] [CrossRef]
- Ding, S.L.; Zhang, M.Y.; Tang, S.J.; Yang, H.; Tan, W.Q. Effect of Calcium Alginate Microsphere Loaded with Vascular Endothelial Growth Factor on Adipose Tissue Transplantation. Ann. Plast. Surg. 2015, 75, 644–651. [Google Scholar] [CrossRef]
- Poels, J.; Abou-Ghannam, G.; Decamps, A.; Leyman, M.; Des Rieux, A.; Wyns, C. Transplantation of Testicular Tissue in Alginate Hydrogel Loaded with VEGF Nanoparticles Improves Spermatogonial Recovery. J. Control. Release 2016, 234, 79–89. [Google Scholar] [CrossRef]
- Capone, S.H.; Dufresne, M.; Rechel, M.; Fleury, M.J.; Salsac, A.V.; Paullier, P.; Daujat-Chavanieu, M.; Legallais, C. Impact of Alginate Composition: From Bead Mechanical Properties to Encapsulated HepG2/C3A Cell Activities for In Vivo Implantation. PLoS ONE 2013, 8, e62032. [Google Scholar] [CrossRef]
- Mhanna, R.; Kashyap, A.; Palazzolo, G.; Vallmajo-Martin, Q.; Becher, J.; Möller, S.; Schnabelrauch, M.; Zenobi-Wong, M. Chondrocyte Culture in Three Dimensional Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers. Tissue Eng. Part A 2014, 20, 1454–1464. [Google Scholar] [CrossRef]
- Bochenek, M.A.; Veiseh, O.; Vegas, A.J.; McGarrigle, J.J.; Qi, M.; Marchese, E.; Omami, M.; Doloff, J.C.; Mendoza-Elias, J.; Nourmohammadzadeh, M.; et al. Alginate Encapsulation as Long-Term Immune Protection of Allogeneic Pancreatic Islet Cells Transplanted into the Omental Bursa of Macaques. Nat. Biomed. Eng. 2018, 2, 810–821. [Google Scholar] [CrossRef]
- Sandvig, I.; Karstensen, K.; Rokstad, A.M.; Aachmann, F.L.; Formo, K.; Sandvig, A.; Skjåk-Bræk, G.; Strand, B.L. RGD-Peptide Modified Alginate by a Chemoenzymatic Strategy for Tissue Engineering Applications. J. Biomed. Mater. Res. A 2015, 103, 896–906. [Google Scholar] [CrossRef]
- Yang, F.; Yang, D.; Tu, J.; Zheng, Q.; Cai, L.; Wang, L. Strontium Enhances Osteogenic Differentiation of Mesenchymal Stem Cells and in Vivo Bone Formation by Activating Wnt/Catenin Signaling. Stem Cells 2011, 29, 981–991. [Google Scholar] [CrossRef]
- Hassani, A.; Avci, Ç.B.; Kerdar, S.N.; Amini, H.; Amini, M.; Ahmadi, M.; Sakai, S.; Bagca, B.G.; Ozates, N.P.; Rahbarghazi, R.; et al. Interaction of Alginate with Nano-Hydroxyapatite-Collagen Using Strontium Provides Suitable Osteogenic Platform. J. Nanobiotechnol. 2022, 20, 310. [Google Scholar] [CrossRef]
- Lee, C.S.D.; Moyer, H.R.; Gittens, I.R.A.; Williams, J.K.; Boskey, A.L.; Boyan, B.D.; Schwartz, Z. Regulating in Vivo Calcification of Alginate Microbeads. Biomaterials 2010, 31, 4926–4934. [Google Scholar] [CrossRef]
- Azhar, F.F.; Olad, A.; Salehi, R. Fabrication and Characterization of Chitosan-Gelatin/Nanohydroxyapatite- Polyaniline Composite with Potential Application in Tissue Engineering Scaffolds. Des. Monomers Polym. 2014, 17, 654–667. [Google Scholar] [CrossRef]
- Anamizu, M.; Tabata, Y. Design of Injectable Hydrogels of Gelatin and Alginate with Ferric Ions for Cell Transplantation. Acta Biomater. 2019, 100, 184–190. [Google Scholar] [CrossRef]
- Boonyagul, S.; Pukasamsombut, D.; Pengpanich, S.; Toobunterng, T.; Pasanaphong, K.; Sathirapongsasuti, N.; Tawonsawatruk, T.; Wangtueai, S.; Tanadchangsaeng, N. Bioink Hydrogel from Fish Scale Gelatin Blended with Alginate for 3D-Bioprinting Application. J. Food Process Preserv. 2021, 46, e15864. [Google Scholar] [CrossRef]
- Amini, H.; Hashemzadeh, S.; Heidarzadeh, M.; Mamipour, M.; Yousefi, M.; Saberianpour, S.; Rahbarghazi, R.; Nouri, M.; Sokullu, E. Cytoprotective and Cytofunctional Effect of Polyanionic Polysaccharide Alginate and Gelatin Microspheres on Rat Cardiac Cells. Int. J. Biol. Macromol. 2020, 161, 969–976. [Google Scholar] [CrossRef]
- Saberianpour, S.; Karimi, A.; Nemati, S.; Amini, H.; Alizadeh Sardroud, H.; Khaksar, M.; Mamipour, M.; Nouri, M.; Rahbarghazi, R. Encapsulation of Rat Cardiomyoblasts with Alginate-Gelatin Microspheres Preserves Stemness Feature in Vitro. Biomed. Pharmacother. 2019, 109, 402–407. [Google Scholar] [CrossRef]
- Lewińska, D.; Rosiński, S.; Weryński, A. Influence of Process Conditions during Impulsed Electrostatic Droplet Formation on Size Distribution of Hydrogel Beads. Artif. Cells Blood Substit. Biotechnol. 2004, 32, 41–53. [Google Scholar] [CrossRef]
- Nemati, S.; Rezabakhsh, A.; Khoshfetrat, A.B.; Nourazarian, A.; BirayAvci, Ç.; GokerBagca, B.; Alizadeh Sardroud, H.; Khaksar, M.; Ahmadi, M.; Delkhosh, A.; et al. Alginate-Gelatin Encapsulation of Human Endothelial Cells Promoted Angiogenesis in in Vivo and in Vitro Milieu. Biotechnol. Bioeng. 2017, 114, 2920–2930. [Google Scholar] [CrossRef]
- Nemati, S.; Alizadeh Sardroud, H.; Baradar Khoshfetrat, A.; Khaksar, M.; Ahmadi, M.; Amini, H.; Saberianpour, S.; Delkhosh, A.; Akbar Movassaghpour, A.; Rahbarghazi, R. The Effect of Alginate–Gelatin Encapsulation on the Maturation of Human Myelomonocytic Cell Line U937. J. Tissue Eng. Regen. Med. 2019, 13, 25–35. [Google Scholar] [CrossRef]
- Alizadeh Sardroud, H.; Nemati, S.; Baradar Khoshfetrat, A.; Nabavinia, M.; BeygiKhosrowshahi, Y. Barium-Cross-Linked Alginate-Gelatine Microcapsule as a Potential Platform for Stem Cell Production and Modular Tissue Formation. J. Microencapsul. 2017, 34, 488–497. [Google Scholar] [CrossRef]
- Ling, S.D.; Liu, Z.; Ma, W.; Chen, Z.; Du, Y.; Xu, J. A Novel Step-T-Junction Microchannel for the Cell Encapsulation in Monodisperse Alginate-Gelatin Microspheres of Varying Mechanical Properties at High Throughput. Biosensors 2022, 12, 659. [Google Scholar] [CrossRef]
- Firouzi, N.; Baradar Khoshfetrat, A.; Kazemi, D. Enzymatically Gellable Gelatin Improves Nano-Hydroxyapatite-Alginate Microcapsule Characteristics for Modular Bone Tissue Formation. J. Biomed. Mater. Res. A 2020, 108, 340–350. [Google Scholar] [CrossRef]
- Gleeson, J.P.; Plunkett, N.A.; O’Brien, F.J. Addition of Hydroxyapatite Improves Stiffness, Interconnectivity and Osteogenic Potential of a Highly Porous Collagen-Based Scaffold for Bone Tissue Regeneration. Eur. Cells Mater. 2010, 20, 218–230. [Google Scholar] [CrossRef]
- Wang, P.; Song, Y.; Weir, M.D.; Sun, J.; Zhao, L.; Simon, C.G.; Xu, H.H.K. A Self-Setting IPSMSC-Alginate-Calcium Phosphate Paste for Bone Tissue Engineering. Dent. Mater. 2016, 32, 252–263. [Google Scholar] [CrossRef]
- Shih, Y.R.V.; Hwang, Y.; Phadke, A.; Kang, H.; Hwang, N.S.; Caro, E.J.; Nguyen, S.; Siu, M.; Theodorakis, E.A.; Gianneschi, N.C.; et al. Calcium Phosphate-Bearing Matrices Induce Osteogenic Differentiation of Stem Cells through Adenosine Signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 990–995. [Google Scholar] [CrossRef]
- Rezaei, S.; Shakibaie, M.; Kabir-Salmani, M.; Moghaddam, M.S.; Rezvani, M.; Shahali, M.; Naseri, M. Improving the Growth Rate of Human Adipose-Derived Mesenchymal Stem Cells in Alginate/Gelatin versus Alginate Hydrogels. Iran. J. Biotechnol. 2016, 14, 1–8. [Google Scholar] [CrossRef]
- Gao, C.; Peng, S.; Feng, P.; Shuai, C. Bone Biomaterials and Interactions with Stem Cells. Bone Res. 2017, 5, 17059–17166. [Google Scholar] [CrossRef]
- Shie, M.Y.; Ding, S.J. Integrin Binding and MAPK Signal Pathways in Primary Cell Responses to Surface Chemistry of Calcium Silicate Cements. Biomaterials 2013, 34, 6589–6606. [Google Scholar] [CrossRef]
- Aiyelabegan, H.T.; Sadroddiny, E. Fundamentals of Protein and Cell Interactions in Biomaterials. Biomed. Pharmacother. 2017, 88, 956–970. [Google Scholar] [CrossRef]
- Rastogi, S.; Pandey, M.M.; Rawat, A.K.S. Medicinal Plants of the Genus Betula—Traditional Uses and a Phytochemical-Pharmacological Review. J. Ethnopharmacol. 2015, 159, 62–83. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, H.J.; Kim, K.R.; Lee, S.K.; Lee, C.K.; Park, K.K.; Chung, W.Y. Betulinic Acid, a Bioactive Pentacyclic Triterpenoid, Inhibits Skeletal-Related Events Induced by Breast Cancer Bone Metastases and Treatment. Toxicol. Appl. Pharmacol. 2014, 275, 152–162. [Google Scholar] [CrossRef]
- Yogeeswari, P.; Sriram, D. Betulinic Acid and Its Derivatives: A Review on Their Biological Properties. Curr. Med. Chem. 2010, 12, 657–666. [Google Scholar] [CrossRef]
- Fulda, S. Betulinic Acid for Cancer Treatment and Prevention. Int. J. Mol. Sci. 2008, 9, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.C.; Chang, Y.H.; Wei, B.L.; Huang, Y.L.; Chiou, W.F. Betulinic Acid Stimulates the Differentiation and Mineralization of Osteoblastic MC3T3-E1 Cells: Involvement of BMP/Runx2 and β-Catenin Signals. J. Agric. Food Chem. 2010, 58, 6643–6649. [Google Scholar] [CrossRef] [PubMed]
- Mizerska-Kowalska, M.; Sławinska-Brych, A.; Kaławaj, K.; Zurek, A.; Pawinska, B.; Rzeski, W.; Zdzisinska, B. Betulin Promotes Differentiation of Human Osteoblasts in Vitro and Exerts an Osteoinductive Effect on the HfOB 1.19 Cell Line through Activation of JNK, ERK1/2, and MTOR Kinases. Molecules 2019, 24, 2637–2653. [Google Scholar] [CrossRef]
- Leslie, S.K.; Kinney, R.C.; Schwartz, Z.; Boyan, B.D. Microencapsulation of Stem Cells for Therapy. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2017; Volume 1479, pp. 251–259. [Google Scholar]
- Leslie, S.K.; Cohen, D.J.; Boyan, B.D.; Schwartz, Z. Production of Osteogenic and Angiogenic Factors by Microencapsulated Adipose Stem Cells Varies with Culture Conditions. J. Biomed. Mater. Res. B Appl. Biomater. 2020, 108, 1857–1867. [Google Scholar] [CrossRef] [PubMed]
- Chacon, E.; Acosta, D.; Lemasters, J.J. Primary Cultures of Cardiac Myocytes as In Vitro Models for Pharmacological and Toxicological Assessments. In In Vitro Methods in Pharmaceutical Research; Academic Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Koyama, R.; Hakamata, W.; Hirano, T.; Nishio, T. Identification of Small-Molecule Inhibitors of Human Golgi Mannosidase via a Drug Repositioning Screen. Chem. Pharm. Bull. 2018, 66, 678–681. [Google Scholar] [CrossRef] [PubMed]
- Jacobs-Tulleneers-Thevissen, D.; Chintinne, M.; Ling, Z.; Gillard, P.; Schoonjans, L.; Delvaux, G.; Strand, B.L.; Gorus, F.; Keymeulen, B.; Pipeleers, D. Sustained Function of Alginate-Encapsulated Human Islet Cell Implants in the Peritoneal Cavity of Mice Leading to a Pilot Study in a Type 1 Diabetic Patient. Diabetologia 2013, 56, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.Y.; Lin, F.H.; Sadhasivam, S.; Savitha, S. Antioxidant Effects of Betulin on Porcine Chondrocyte Behavior in Gelatin/C6S/C4S/HA Modified Tricopolymer Scaffold. Mater. Sci. Eng. C 2010, 30, 597–604. [Google Scholar] [CrossRef]
- Shoichet, M.S.; Li, R.H.; White, M.L.; Winn, S.R. Stability of Hydrogels Used in Cell Encapsulation: An in Vitro Comparison of Alginate and Agarose. Biotechnol. Bioeng. 1996, 50, 374–381. [Google Scholar] [CrossRef]
- Leslie, S.K.; Cohen, D.J.; Sedlaczek, J.; Pinsker, E.J.; Boyan, B.D.; Schwartz, Z. Controlled Release of Rat Adipose-Derived Stem Cells from Alginate Microbeads. Biomaterials 2013, 34, 8172–8184. [Google Scholar] [CrossRef]
- Modi, P.K.; Prabhu, A.; Bhandary, Y.P.; Sudheer Shenoy, P.; Hegde, A.; Sindhu Priya, E.S.; Johnson, R.P.; Das, S.P.; Vazirally, S.; Rekha, P.D. Effect of Calcium Glucoheptonate on Proliferation and Osteogenesis of Osteoblast-like Cells in Vitro. PLoS ONE 2019, 14, e0222240. [Google Scholar] [CrossRef]
- Li, C.; Qi, Y.; Zhou, Q.; Huang, X.; Deng, X.; Yu, Y.; Shi, L.E. Betulinic Acid Promotes the Osteogenic Differentiation of Human Periodontal Ligament Stem Cells by Upregulating EGR1. Acta Biochim. Sin. 2021, 53, 1266–1276. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, X.; Zhou, D.; Vacek, I.; Sun, A.M. Normalization of Diabetes in Spontaneously Diabetic Cynomologus Monkeys by Xenografts of Microencapsulated Porcine Islets without Immunosuppression. J. Clin. Investig. 1996, 98, 1417–1422. [Google Scholar] [CrossRef]
- Aguado, B.A.; Mulyasasmita, W.; Su, J.; Lampe, K.J.; Heilshorn, S.C. Improving Viability of Stem Cells during Syringe Needle Flow through the Design of Hydrogel Cell Carriers. Tissue Eng. Part A 2012, 18, 806–815. [Google Scholar] [CrossRef] [PubMed]
- Alipour, M.; Firouzi, N.; Aghazadeh, Z.; Samiei, M.; Montazersaheb, S.; Khoshfetrat, A.B.; Aghazadeh, M. The Osteogenic Differentiation of Human Dental Pulp Stem Cells in Alginate-Gelatin/Nano-Hydroxyapatite Microcapsules. BMC Biotechnol. 2021, 21, 6. [Google Scholar] [CrossRef] [PubMed]
- Salinas, C.N.; Anseth, K.S. The Enhancement of Chondrogenic Differentiation of Human Mesenchymal Stem Cells by Enzymatically Regulated RGD Functionalities. Biomaterials 2008, 29, 2370–2377. [Google Scholar] [CrossRef] [PubMed]
- Koyama, R.; Kano, Y.; Kikushima, K.; Mizutani, A.; Soeda, Y.; Miura, K.; Hirano, T.; Nishio, T.; Hakamata, W. A Novel Golgi Mannosidase Inhibitor: Molecular Design, Synthesis, Enzyme Inhibition, and Inhibition of Spheroid Formation. Bioorg. Med. Chem. 2020, 28, 115492. [Google Scholar] [CrossRef] [PubMed]
- Sarker, B.; Rompf, J.; Silva, R.; Lang, N.; Detsch, R.; Kaschta, J.; Fabry, B.; Boccaccini, A.R. Alginate-Based Hydrogels with Improved Adhesive Properties for Cell Encapsulation. Int. J. Biol. Macromol. 2015, 78, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Senamontree, S.; Lakthan, T.; Charoenpanich, P.; Chanchao, C.; Charoenpanich, A. Betulinic Acid Decreases Lipid Accumulation in Adipogenesis-Induced Human Mesenchymal Stem Cells with Upregulation of Pgc-1a and Ucp-1 and Post-Transcriptional Downregulation of Adiponectin and Leptin Secretion. PeerJ 2021, 9, e12321. [Google Scholar] [CrossRef] [PubMed]
- Poher, A.L.; Veyrat-Durebex, C.; Altirriba, J.; Montet, X.; Colin, D.J.; Caillon, A.; Lyautey, J.; Rohner-Jeanrenaud, F. Ectopic UCP1 Overexpression in White Adipose Tissue Improves Insulin Sensitivity in Lou/C Rats, a Model of Obesity Resistance. Diabetes 2015, 64, 3700–3712. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Shayan, P.; Busch, F.; Aldinger, C.; Buhrmann, C.; Lueders, C.; Mobasheri, A. Resveratrol Mediated Modulation of Sirt-1/Runx2 Promotes Osteogenic Differentiation of Mesenchymal Stem Cells: Potential Role of Runx2 Deacetylation. PLoS ONE 2012, 7, e35712. [Google Scholar] [CrossRef] [PubMed]
- Brusotti, G.; Montanari, R.; Capelli, D.; Cattaneo, G.; Laghezza, A.; Tortorella, P.; Loiodice, F.; Peiretti, F.; Bonardo, B.; Paiardini, A.; et al. Betulinic Acid Is a PPARλ Antagonist That Improves Glucose Uptake, Promotes Osteogenesis and Inhibits Adipogenesis. Sci. Rep. 2017, 7, 5777. [Google Scholar] [CrossRef]
- Qiu, G.; Huang, M.; Liu, J.; Ma, T.; Schneider, A.; Oates, T.W.; Lynch, C.D.; Weir, M.D.; Zhang, K.; Zhao, L.; et al. Human Periodontal Ligament Stem Cell Encapsulation in Alginate-Fibrin-Platelet Lysate Microbeads for Dental and Craniofacial Regeneration. J. Dent. 2022, 124, 104219. [Google Scholar] [CrossRef]
- Xu, M.; Liu, T.; Qin, M.; Cheng, Y.; Lan, W.; Niu, X.; Wei, Y.; Hu, Y.; Lian, X.; Zhao, L.; et al. Bone-like Hydroxyapatite Anchored on Alginate Microspheres for Bone Regeneration. Carbohydr. Polym. 2022, 287, 119330. [Google Scholar] [CrossRef] [PubMed]
- Deligianni, D.D.; Katsala, N.D.; Koutsoukos, P.G.; Missirlis, Y.F. Effect of Surface Roughness of Hydroxyapatite on Human Bone Marrow Cell Adhesion, Proliferation, Differentiation and Detachment Strength. Biomaterials 2000, 22, 87–96. [Google Scholar] [CrossRef] [PubMed]
- García, A.J.; Boettiger, D. Integrin-Fibronectin Interactions at the Cell-Material Interface: Initial Integrin Binding and Signaling. Biomaterials 1999, 20, 2427–2433. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karaca, M.A.; Kancagi, D.D.; Ozbek, U.; Ovali, E.; Gok, O. Betulin Stimulates Osteogenic Differentiation of Human Osteoblasts-Loaded Alginate–Gelatin Microbeads. Bioengineering 2024, 11, 553. https://doi.org/10.3390/bioengineering11060553
Karaca MA, Kancagi DD, Ozbek U, Ovali E, Gok O. Betulin Stimulates Osteogenic Differentiation of Human Osteoblasts-Loaded Alginate–Gelatin Microbeads. Bioengineering. 2024; 11(6):553. https://doi.org/10.3390/bioengineering11060553
Chicago/Turabian StyleKaraca, Mehmet Ali, Derya Dilek Kancagi, Ugur Ozbek, Ercument Ovali, and Ozgul Gok. 2024. "Betulin Stimulates Osteogenic Differentiation of Human Osteoblasts-Loaded Alginate–Gelatin Microbeads" Bioengineering 11, no. 6: 553. https://doi.org/10.3390/bioengineering11060553
APA StyleKaraca, M. A., Kancagi, D. D., Ozbek, U., Ovali, E., & Gok, O. (2024). Betulin Stimulates Osteogenic Differentiation of Human Osteoblasts-Loaded Alginate–Gelatin Microbeads. Bioengineering, 11(6), 553. https://doi.org/10.3390/bioengineering11060553