
Production of Astaxanthin by Animal Cells via Introduction of an Entire Astaxanthin Biosynthetic Pathway

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Transfection
Assessment of Transfection Effectiveness
2.2. Synthetic Heterologous Genes Expression and Plasmid Construction
2.3. Ketocarotenoid Extraction
2.4. Ketocarotenoid Analysis
2.5. Statistical Analysis
2.6. Nucleotide Accession Number
3. Results and Discussion
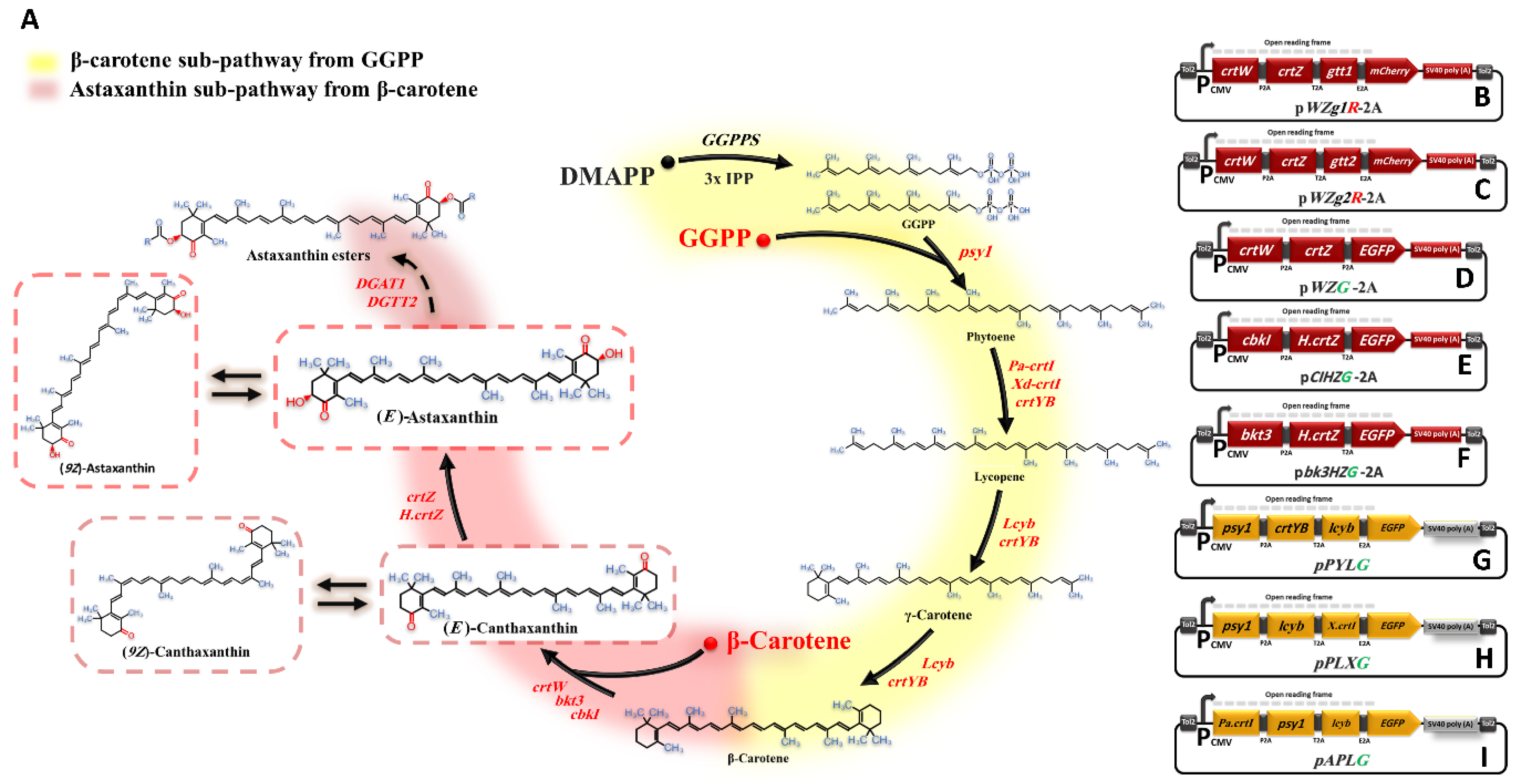
3.1. Genes/Enzymes Selection for Astaxanthin Biosynthesis in Animal Cells In Vitro
3.2. Constructing Ast subp in Animal Cells In Vitro
3.3. Introduction of DGTT1 and DGTT2 Genes for Optimization of Astaxanthin Production and Stability
3.4. Redirection of GGPP Flux towards Astaxanthin Accumulation
3.5. Optimization of the Astaxanthin Biosynthetic Pathway through Exogenous GGPP
3.6. Qualitative Optimization of Total Ketocarotenoids by Individual Expression of the crtZ Gene
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davinelli, S.; Nielsen, M.E.; Scapagnini, G. Astaxanthin in Skin Health, Repair, and Disease: A Comprehensive Review. Nutrients 2018, 10, 522. [Google Scholar] [CrossRef]
- Eggersdorfer, M.; Wyss, A. Carotenoids in Human Nutrition and Health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.A.; Jarvik, T. Lateral Transfer of Genes from Fungi Underlies Carotenoid Production in Aphids. Science 2010, 328, 624–627. [Google Scholar] [CrossRef]
- Pierce, E.C.; LaFayette, P.R.; Ortega, M.A.; Joyce, B.L.; Kopsell, D.A.; Parrott, W.A. Ketocarotenoid Production in Soybean Seeds through Metabolic Engineering. PLoS ONE 2015, 10, e0138196. [Google Scholar] [CrossRef]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. (Eds.) Carotenoids: Handbook; Springer: Bronx, NY, USA; Birkhäuser: Basel, Switzerland, 2004; ISBN 978-3-7643-6180-8. [Google Scholar]
- Ye, L.; Zhu, X.; Wu, T.; Wang, W.; Zhao, D.; Bi, C.; Zhang, X. Optimizing the Localization of Astaxanthin Enzymes for Improved Productivity. Biotechnol. Biofuels 2018, 11, 278. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.X.; Gantt, E. Genes and Enzymes of Carotenoid Biosynthesis in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 557–583. [Google Scholar] [CrossRef]
- Hussein, G.; Sankawa, U.; Goto, H.; Matsumoto, K.; Watanabe, H. Astaxanthin, a Carotenoid with Potential in Human Health and Nutrition. J. Nat. Prod. 2006, 69, 443–449. [Google Scholar] [CrossRef]
- Ambati, R.R.; Moi, P.S.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, Extraction, Stability, Biological Activities and Its Commercial Applications—A Review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef] [PubMed]
- Holtin, K.; Kuehnle, M.; Rehbein, J.; Schuler, P.; Nicholson, G.; Albert, K. Determination of Astaxanthin and Astaxanthin Esters in the Microalgae Haematococcus pluvialis by LC-(APCI)MS and Characterization of Predominant Carotenoid Isomers by NMR Spectroscopy. Anal. Bioanal. Chem. 2009, 395, 1613–1622. [Google Scholar] [CrossRef]
- Schmidt, I.; Schewe, H.; Gassel, S.; Jin, C.; Buckingham, J.; Hümbelin, M.; Sandmann, G.; Schrader, J. Biotechnological Production of Astaxanthin with Phaffia rhodozyma/Xanthophyllomyces dendrorhous. Appl. Microbiol. Biotechnol. 2011, 89, 555–571. [Google Scholar] [CrossRef]
- Hartz, P.; Milhim, M.; Trenkamp, S.; Bernhardt, R.; Hannemann, F. Characterization and Engineering of a Carotenoid Biosynthesis Operon from Bacillus Megaterium. Metab. Eng. 2018, 49, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, M.; Enfissi, E.M.A.; Welsch, R.; Beyer, P.; Zurbriggen, M.D.; Fraser, P.D. Construction of a Fusion Enzyme for Astaxanthin Formation and Its Characterisation in Microbial and Plant Hosts: A New Tool for Engineering Ketocarotenoids. Metab. Eng. 2019, 52, 243–252. [Google Scholar] [CrossRef]
- Zhu, Q.; Zeng, D.; Yu, S.; Cui, C.; Li, J.; Li, H.; Chen, J.; Zhang, R.; Zhao, X.; Chen, L.; et al. From Golden Rice to ASTARice: Bioengineering Astaxanthin Biosynthesis in Rice Endosperm. Mol. Plant 2018, 11, 1440–1448. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.H.; Kim, J.K.; Jeong, Y.S.; You, M.K.; Lim, S.H.; Kim, J.K. Stepwise Pathway Engineering to the Biosynthesis of Zeaxanthin, Astaxanthin and Capsanthin in Rice Endosperm. Metab. Eng. 2019, 52, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Alok, A.; Shivani; Kumar, P.; Kaur, N.; Awasthi, P.; Chaturvedi, S.; Pandey, P.; Pandey, A.; Pandey, A.K.; et al. CRISPR/Cas9 Directed Editing of Lycopene Epsilon-Cyclase Modulates Metabolic Flux for β-Carotene Biosynthesis in Banana Fruit. Metab. Eng. 2020, 59, 76–86. [Google Scholar] [CrossRef]
- Becker, J.; Wittmann, C. Bio-Based Production of Chemicals, Materials and Fuels—Corynebacterium Glutamicum as Versatile Cell Factory. Curr. Opin. Biotechnol. 2012, 23, 631–640. [Google Scholar] [CrossRef]
- Du, J.; Shao, Z.; Zhao, H. Engineering Microbial Factories for Synthesis of Value-Added Products. J. Ind. Microbiol. Biotechnol. 2011, 38, 873–890. [Google Scholar] [CrossRef]
- Bahadur, B.; Rajam, M.V.; Sahijram, L.; Krishnamurthy, K.V. Metabolic Engineering in Plants. Plant Biol. Biotechnol. Vol. II Plant Genom. Biotechnol. 2015, II, 1–768. [Google Scholar] [CrossRef]
- Guo, W.; Sheng, J.; Feng, X. Mini-Review: In Vitro Metabolic Engineering for Biomanufacturing of High-Value Products. Comput. Struct. Biotechnol. J. 2017, 15, 161–167. [Google Scholar] [CrossRef]
- O’Connor, S.E. Engineering of Secondary Metabolism. Annu. Rev. Genet. 2015, 49, 71–94. [Google Scholar] [CrossRef]
- Gòdia, F.; Cairó, J.J. Metabolic Engineering of Animal Cells. Bioprocess Biosyst. Eng. 2002, 24, 289–298. [Google Scholar] [CrossRef]
- Templeton, N.; Young, J.D. Biochemical and Metabolic Engineering Approaches to Enhance Production of Therapeutic Proteins in Animal Cell Cultures. Biochem. Eng. J. 2018, 136, 40–50. [Google Scholar] [CrossRef]
- Pang, S.C.; Wang, H.P.; Li, K.Y.; Zhu, Z.Y.; Kang, J.X.; Sun, Y.H. Double Transgenesis of Humanized Fat1 and Fat2 Genes Promotes Omega-3 Polyunsaturated Fatty Acids Synthesis in a Zebrafish Model. Mar. Biotechnol. 2014, 16, 580–593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Pang, S.; Liu, C.; Wang, H.; Ye, D.; Zhu, Z.; Sun, Y. A Novel Dietary Source of EPA and DHA: Metabolic Engineering of an Important Freshwater Species—Common Carp by Fat1-Transgenesis. Mar. Biotechnol. 2019, 21, 171–185. [Google Scholar] [CrossRef]
- Mohammed, Y.; Ye, D.; He, M.; Wang, H.; Li, Y.; Han, D.; Zhu, Z.; Sun, Y. Induction of Biosynthesis of Ketocarotenoid from β-Carotene in Fish Embryos. Aquaculture 2021, 542, 736863. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, O.; Wall, J.B.J.; Zheng, M.; Zhou, Y.; Wang, L.; Ruth Vaseghi, H.; Qian, L.; Liu, J. Systematic Comparison of 2A Peptides for Cloning Multi-Genes in a Polycistronic Vector. Sci. Rep. 2017, 7, 2193. [Google Scholar] [CrossRef] [PubMed]
- Malm, M.; Saghaleyni, R.; Lundqvist, M.; Giudici, M.; Chotteau, V.; Field, R.; Varley, P.G.; Hatton, D.; Grassi, L.; Svensson, T.; et al. Evolution from Adherent to Suspension—Systems Biology of HEK293 Cell Line Development. Sci. Rep. 2020, 10, 18996. [Google Scholar] [CrossRef]
- Iuchi, K.; Oya, K.; Hosoya, K.; Sasaki, K.; Sakurada, Y. Different Morphologies of Human Embryonic Kidney 293T Cells in Various Types of Culture Dishes. Cytotechnology 2019, 72, 131–140. [Google Scholar] [CrossRef]
- Lin, Y.; Boone, M.; Meuris, L.; Lemmens, I.; Van Roy, N.; Soete, A.; Drmanac, R.; Chen, J.; Reumers, J.; Moisse, M.; et al. Genome Dynamics of the Human Embryonic Kidney 293 Lineage in Response to Cell Biology Manipulations. Nat. Commun. 2014, 5, 4767. [Google Scholar] [CrossRef]
- Barredo, J.-L. Engineering Escherichia Coli for Canthaxanthin and Astaxanthin Biosynthesis. Ser. Methods Mol. Biol. 2012, 892, 355. [Google Scholar] [CrossRef]
- Chen, G.; Wang, B.; Han, D.; Sommerfeld, M.; Lu, Y.; Chen, F.; Hu, Q. Molecular Mechanisms of the Coordination between Astaxanthin and Fatty Acid Biosynthesis in Haematococcus pluvialis (Chlorophyceae). Plant J. 2015, 81, 95–107. [Google Scholar] [CrossRef]
- Ma, H.; Wu, X.; Wei, Z.; Zhao, L.; Li, Z.; Liang, Q.; Zheng, J.; Wang, Y.; Li, Y.; Huang, L.; et al. Functional Divergence of Diacylglycerol Acyltransferases in the Unicellular Green Alga Haematococcus pluvialis. J. Exp. Bot. 2021, 72, 510–524. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.W.; Liu, J.X.; Hu, B.; Xiao, W. Zebrafish Foxo3b Negatively Regulates Canonical Wnt Signaling to Affect Early Embryogenesis. PLoS ONE 2011, 6, e24469. [Google Scholar] [CrossRef]
- Ye, D.; Wang, X.; Wei, C.; He, M.; Wang, H.; Wang, Y.; Zhu, Z.; Sun, Y. Marcksb Plays a Key Role in the Secretory Pathway of Zebrafish Bmp2b. PLoS Genet. 2019, 15, e1008306. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, F.; Wang, R.; Zhao, P.; Xia, Q. 2A Self-Cleaving Peptide-Based Multi-Gene Expression System in the Silkworm Bombyx Mori. Sci. Rep. 2015, 5, 16273. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, S.R.; Li, L.H.; Park, H.J.; Park, J.H.; Lee, K.Y.; Kim, M.K.; Shin, B.A.; Choi, S.Y. High Cleavage Efficiency of a 2A Peptide Derived from Porcine Teschovirus-1 in Human Cell Lines, Zebrafish and Mice. PLoS ONE 2011, 6, e18556. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012; Volume 33, ISBN 978-1-936113-41-5. [Google Scholar]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic Assembly of DNA Molecules up to Several Hundred Kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Yuan, J.P.; Chen, F.; Liu, X.; Li, X.Z. Carotenoid Composition in the Green Microalga Chlorococcum. Food Chem. 2002, 76, 319–325. [Google Scholar] [CrossRef]
- Chekanov, K. Diversity and Distribution of Carotenogenic Algae in Europe: A Review. Mar. Drugs 2023, 21, 108. [Google Scholar] [CrossRef]
- Lane, K.T.; Beese, L.S. Structural Biology of Protein Farnesyltransferase and Geranylgeranyltransferase Type I. J. Lipid Res. 2006, 47, 681–699. [Google Scholar] [CrossRef] [PubMed]
- Wiemer, A.J.; Wiemer, D.F.; Hohl, R.J. Geranylgeranyl Diphosphate Synthase: An Emerging Therapeutic Target. Clin. Pharmacol. Ther. 2009, 90, 804–812. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, W.J.; Olivier, L.M.; Krisans, S.K. Central Role of Peroxisomes in Isoprenoid Biosynthesis. Prog. Lipid Res. 2002, 41, 369–391. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Ast ± SD | Ast % | Can ± SD | Can % | Total (Ast + Can) |
|---|---|---|---|---|---|
| pbK3HZ-2A + β-carotene | 3.1 ± 5.4 | 7.8 | 37.1 ± 2.4 | 92.2 | 40.2 ± 7.6 |
| pC1HZG-2A + β-carotene | 17.8 ± 1.5 ** | 32.0 | 37.8 ± 8.0 * | 67.9 | 55.6 ± 7.3 |
| pWZG-2A + β-carotene | 31.6 ± 1.3 ** | 42.6 | 42.6 ± 1.5 *** | 57.4 | 74.2 ± 2.2 |
| pWZg2R-2A + β-carotene | 32.5 ± 1.3 | 35.9 | 57.9 ± 0.6 | 64.0 | 90.4 ± 1.8 ** |
| pWZg1R-2A + β-carotene | 34.1 ± 8.5 | 24.3 | 106.1 ± 12.0 *** | 75.7 | 140.2 ± 5.5 *** |
| pPLXG + pWZg1R | 21.7 ± 5.1 | 49.9 | 21.8 ± 1.3 | 50.0 | 43.5 ± 5.9 |
| pAPLG + pWZg1R | 31.4 ± 2.0 * | 60.7 | 20.3 ± 2.5 | 39.3 | 51.7 ± 1.7 |
| pPYLG + pWZg1R | 41.9 ± 1.3 *** | 66.2 | 21.4 ± 2.1 | 33.8 | 63.2 ± 3.1 ** |
| pPLXG + pWZg1R + GGPP | 19.5 ± 1.1 *** | 25.7 | 55.4 ± 3.5 | 73.9 | 74.9 ± 4.2 ** |
| pAPLG + pWZg1R + GGPP | 43.2 ± 8.7 *** | 34.7 | 81.2 ± 5.1 * | 65.3 | 124.4 ± 5 *** |
| pPYLG+pWZg1R+GGPP | 54.1 ± 5.0 * | 26.1 | 153.1 ± 0.5 *** | 73.9 | 207.2 ± 5.1 *** |
| pPLXG + pWZg1R + pZR + GGPP | 146.4 ± 7.8 *** | 94.7 | 8.2 ± 1.2 *** | 5.3 | 154.6 ± 7.3 *** |
| pAPLG + pWZg1R + pZR + GGPP | 136.7 ± 2.1 *** | 69.8 | 59.2 ± 8.0 ** | 30.2 | 196.0 ± 7.7 *** |
| pPYLG + pWZg1R + pZR + GGPP | 232.8 ± 6.0 *** | 88.8 | 29.3 ± 2.0 ** | 11.2 | 262.1 ± 4.3 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammed, Y.; Ye, D.; He, M.; Wang, H.; Zhu, Z.; Sun, Y. Production of Astaxanthin by Animal Cells via Introduction of an Entire Astaxanthin Biosynthetic Pathway. Bioengineering 2023, 10, 1073. https://doi.org/10.3390/bioengineering10091073
Mohammed Y, Ye D, He M, Wang H, Zhu Z, Sun Y. Production of Astaxanthin by Animal Cells via Introduction of an Entire Astaxanthin Biosynthetic Pathway. Bioengineering. 2023; 10(9):1073. https://doi.org/10.3390/bioengineering10091073
Chicago/Turabian StyleMohammed, Yousef, Ding Ye, Mudan He, Houpeng Wang, Zuoyan Zhu, and Yonghua Sun. 2023. "Production of Astaxanthin by Animal Cells via Introduction of an Entire Astaxanthin Biosynthetic Pathway" Bioengineering 10, no. 9: 1073. https://doi.org/10.3390/bioengineering10091073
APA StyleMohammed, Y., Ye, D., He, M., Wang, H., Zhu, Z., & Sun, Y. (2023). Production of Astaxanthin by Animal Cells via Introduction of an Entire Astaxanthin Biosynthetic Pathway. Bioengineering, 10(9), 1073. https://doi.org/10.3390/bioengineering10091073