

Magnesium and Calcium Transport along the Male Rat Kidney: Effect of Diuretics


Abstract
1. Introduction
2. Materials and Methods
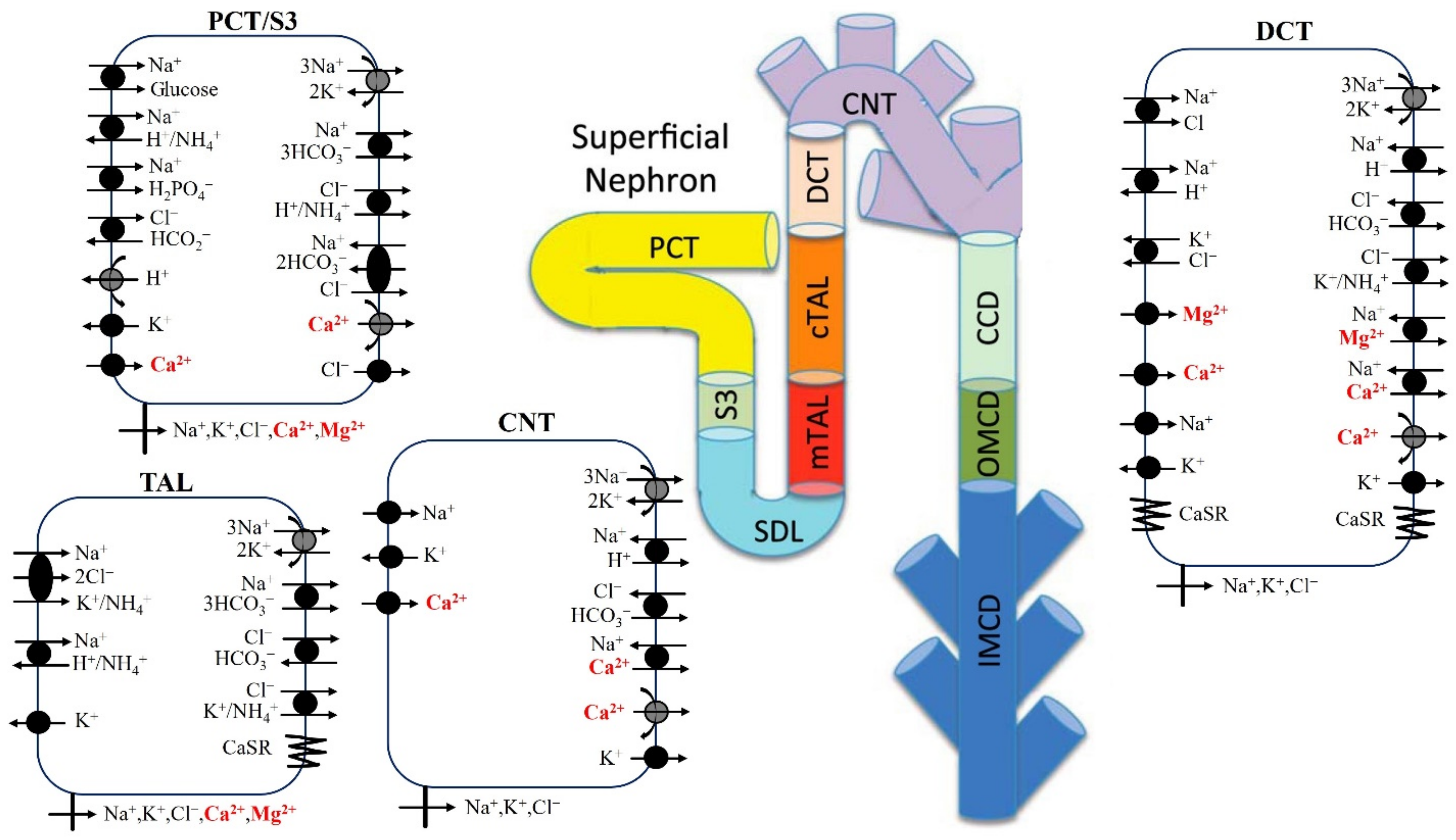
2.1. Mg2+ Transport along the Proximal Tubule
2.2. Mg2+ Transport along the Thick Ascending Limb
2.3. Mg2+ Transport along the Distal Convoluted Tubule
2.4. Calcium-Sensing Receptor

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| PT | |
| Tight junction permeability to Mg2+ at the lumen-LIS interface () | 1.1 × 10−5 cm/s [26] |
| Reflection coefficient of tight junction to Mg2+ | 0.89 (*) |
| cTAL | |
| Tight junction permeability at the lumen-LIS interface in the absence of Mg2+ () | 38 × 10−5 cm/s (*) |
| Maximum half concentration of Ca2+ () | 1.25 mM [24] |
| Maximum half concentration of Mg2+ () | 2.5 mM [24] |
| Hill function coefficient, n | 4 [24] |
| Inhibitory coefficient of Ca2+ on tight junction permeability () | −4/7 [27] |
| Inhibitory coefficient of Mg2+ on tight junction permeability () | −0.34 (*) |
| Inhibitory coefficient of Ca2+ on NKCC2 activity () | −0.4 [28] |
| Inhibitory coefficient of Mg2+ on NKCC2 activity () | −0.24 (*) |
| Inhibitory coefficient of Ca2+ on ROMK activity () | −0.8 [29,30] |
| Inhibitory coefficient of Mg2+ on ROMK activity () | −0.48 (*) |
| DCT | |
| TRPM6/7 channel density () | 26 × 104 cm-2 (*) |
| Single channel conductance of TRPM6/7 at pH 7.4 () | 56.6 pS [20] |
| Luminal pH for half-maximal conductance of TRPM6/7 () | 5.5 [20] |
| Maximum Mg2+ flux through Na+/Mg2+ exchanger () | 8.6 × 10−9 mmol/cm2/s (*) |
| Intracellular Mg2+ half-saturation constant () | 3.59 M [19,21] |
| Extracellular Mg2+ half-saturation constant () | 1.3 mM [19,21] |
| Intracellular Na+ half-saturation constant () | 12.29 mM [19,21] |
| Extracellular Na+ half-saturation constant () | 87.5 mM [19,21] |
| Excitatory coefficient of Ca2+ on NCC activity () | 0.5 [31] |
| Excitatory coefficient of Mg2+ on NCC activity () | 0.3 (*) |
| CD | |
| Ca2+ promoting coefficient for apical HATPase activity of type A OMCD cells () | 2 [27] |
| Mg2+ promoting coefficient for apical HATPase activity of type A OMCD cells () | 1.2 (*) |
| Ca2+ inhibitory coefficient for apical water permeability of IMCD cells () | −3/8 [27] |
| Mg2+ inhibitory coefficient for apical water permeability of IMCD cells () | −0.225 (*) |
3. Results
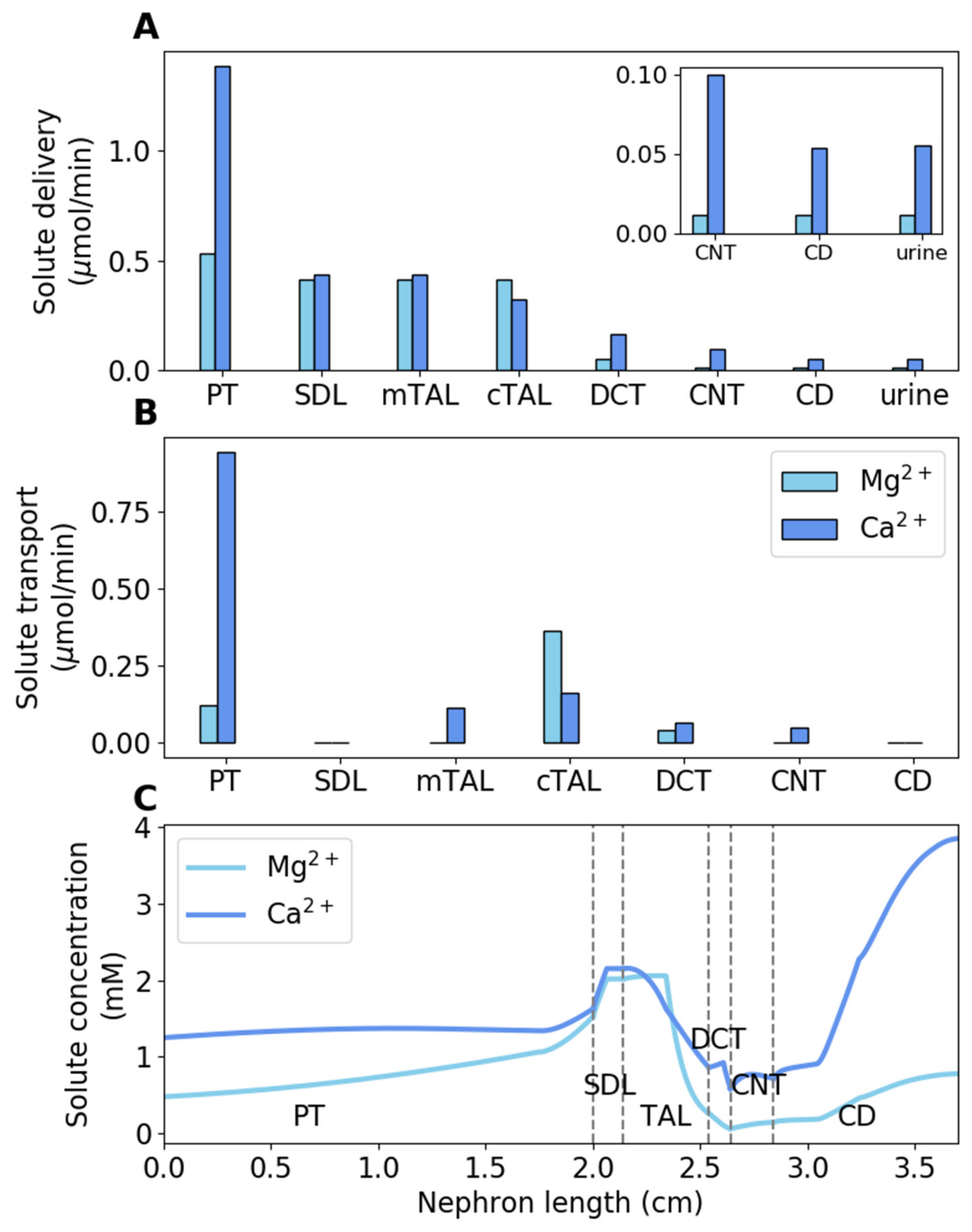
3.1. Baseline Results
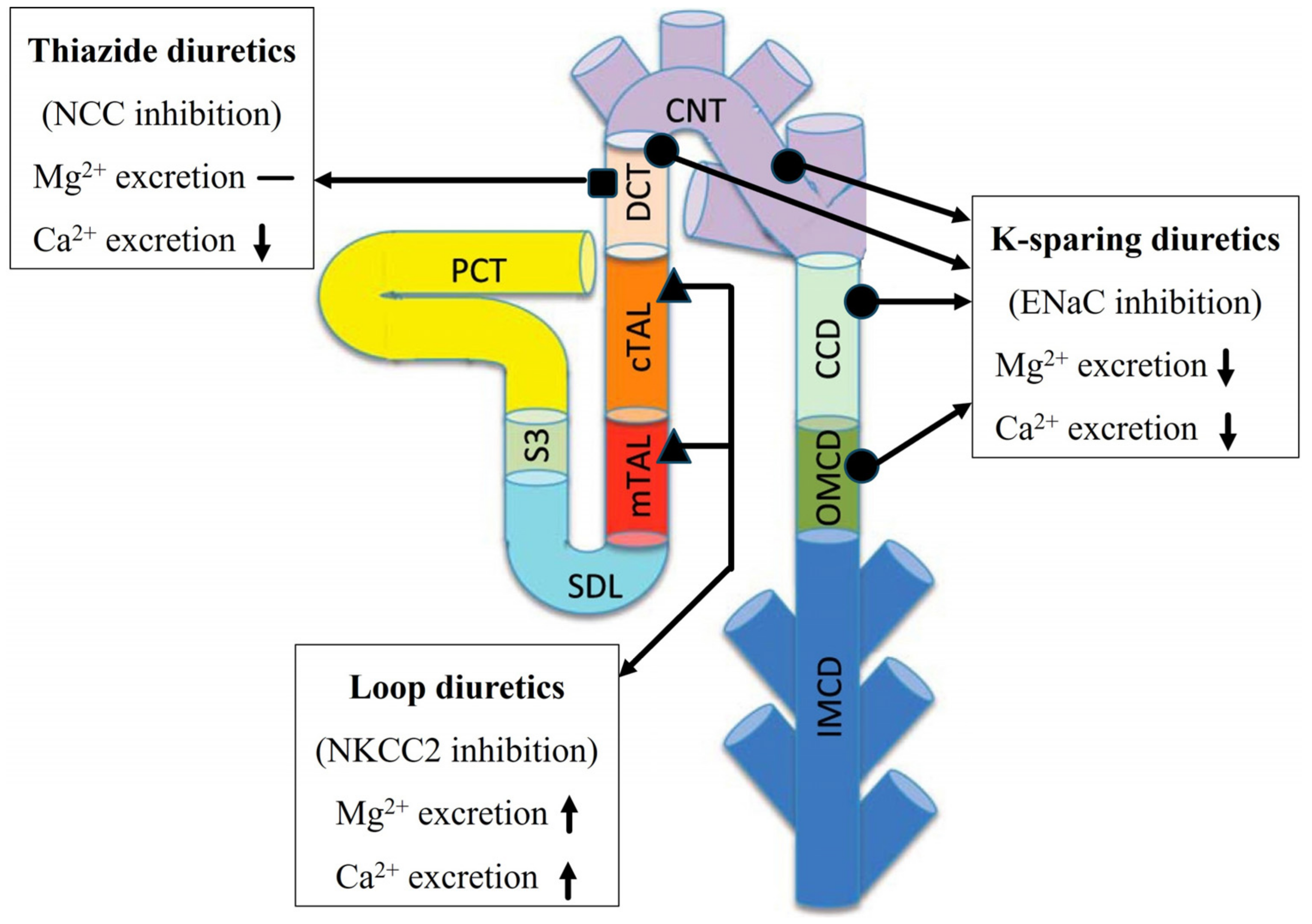
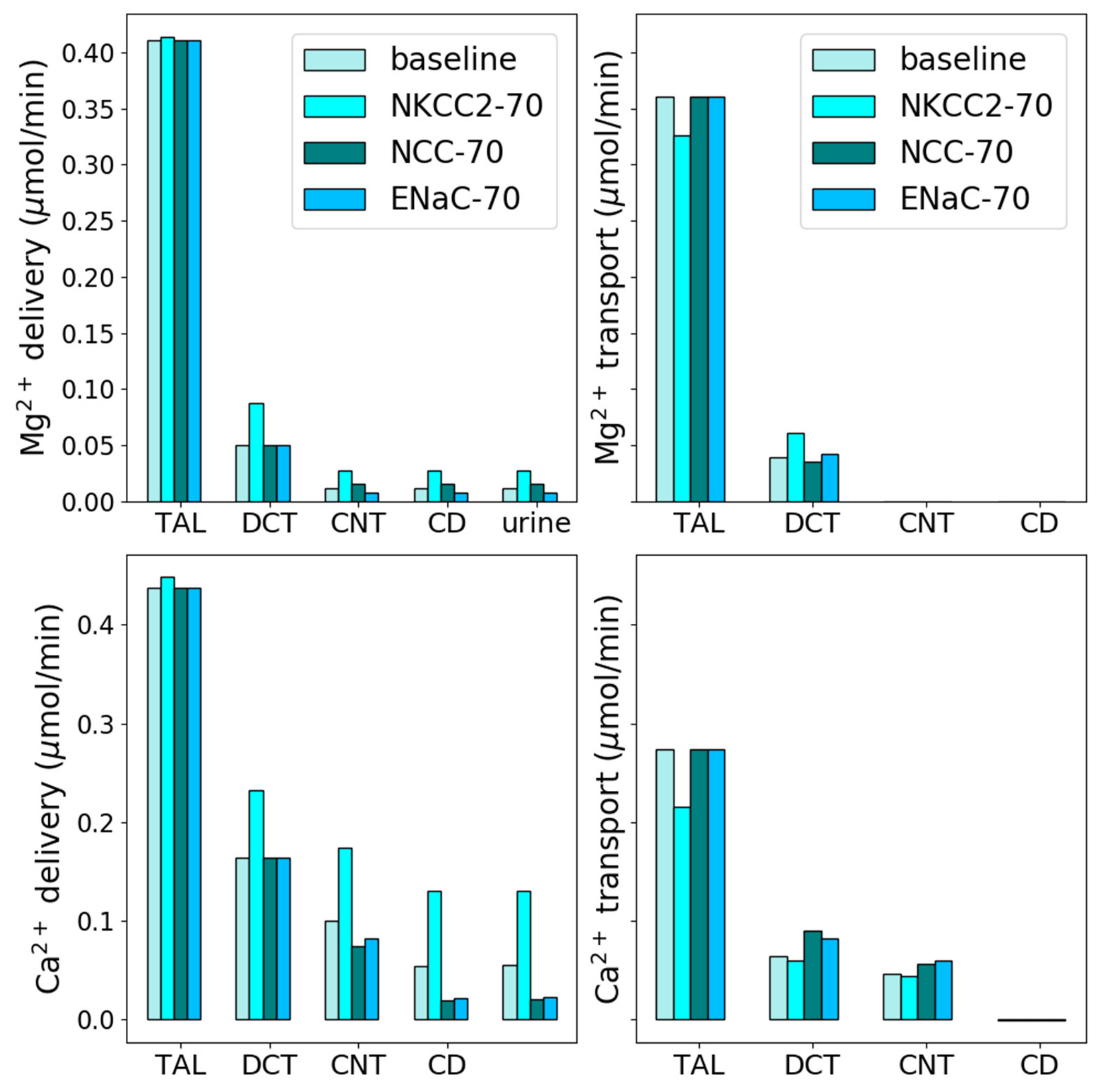
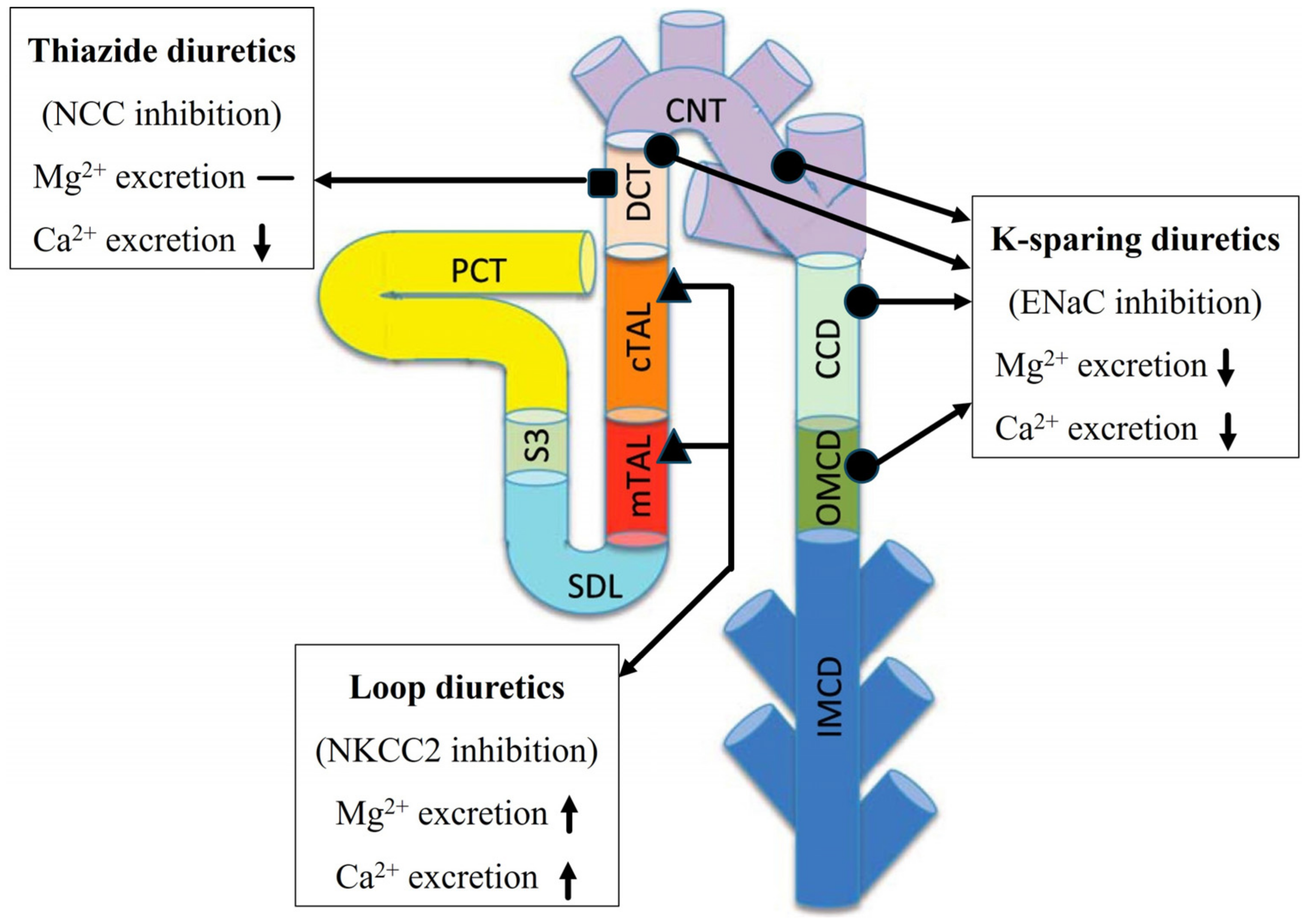
3.2. Effect of Loop Diuretics
3.3. Thiazide Diuretics
3.4. K-Sparing Diuretics
3.5. TRPM6/7 Inhibition
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pu, F.; Chen, N.; Xue, S. Calcium intake, calcium homeostasis and health. Food Sci. Hum. Wellness 2016, 5, 8–16. [Google Scholar] [CrossRef]
- Taal, M.W.; Chertow, G.M.; Marsden, P.A.; Skorecki, K.; Alan, S.; Brenner, B.M. Brenner and Rector’s the Kidney E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2011; ISBN 1-4557-2304-5. [Google Scholar]
- Alexander, R.T.; Dimke, H. Effect of diuretics on renal tubular transport of calcium and magnesium. Am. J. Physiol. Ren. Physiol. 2017, 312, F998–F1015. [Google Scholar] [CrossRef]
- Brunette, M.; Vigneault, N.; Carriere, S. Micropuncture study of magnesium transport along the nephron in the young rat. Am. J. Physiol. Leg. Content 1974, 227, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Le Grimellec, C.; Giocondi, M.C.; Philippe, P. Micropuncture study along the proximal convoluted tubule electrolyte reabsorption in first convolutions. Pflug. Arch. 1975, 354, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Curry, J.N.; Yu, A.S.L. Magnesium Handling in the Kidney. Adv. Chronic Kidney Dis. 2018, 25, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Layton, A.T.; Layton, H.E. A Computational Model of Epithelial Solute and Water Transport along a Human Nephron. PLoS Comput. Biol. 2018, 15, e1006108. [Google Scholar] [CrossRef] [PubMed]
- Layton, A.T.; Vallon, V. SGLT2 Inhibition in a Kidney with Reduced Nephron Number: Modeling and Analysis of Solute Transport and Metabolism. Am. J. Physiol. Ren. Physiol. 2018, 314, F969–F984. [Google Scholar] [CrossRef] [PubMed]
- Layton, A.T.; Laghmani, K.; Vallon, V.; Edwards, A. Solute transport and oxygen consumption along the nephrons: Effects of Na+ transport inhibitors. Am. J. Physiol. Ren. Physiol. 2016, 311, F1217–F1229. [Google Scholar] [CrossRef]
- Hakimi, S.; Dutta, P.; Layton, A.T. Coupling of renal sodium and calcium transport: A modeling analysis of transporter inhibition and sex differences. Am. J. Physiol. Ren. Physiol. 2023, 325, F536–F551. [Google Scholar] [CrossRef]
- Layton, A. A Complete Set of Equations for a Computational Model of Electrolyte and Water Transport along the Nephrons in a Mammalian Kidney. bioRxiv 2022, bioRxiv:509286. [Google Scholar] [CrossRef]
- Layton, A.T.; Vallon, V.; Edwards, A. A computational model for simulating solute transport and oxygen consumption along the nephrons. Am. J. Physiol. Ren. Physiol. 2016, 311, F1378–F1390. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.T.; Dimke, H. Molecular mechanisms underlying paracellular calcium and magnesium reabsorption in the proximal tubule and thick ascending limb. Ann. N. Y. Acad. Sci. 2022, 1518, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Topham, D.J.; Park, S.Y.; Hollenbaugh, J.; Treanor, J.; Mosmann, T.R.; Jin, X.; Ward, B.M.; Miao, H.; Holden-Wiltse, J. Simulation and prediction of the adaptive immune response to influenza A virus infection. J. Virol. 2009, 83, 7151–7165. [Google Scholar] [CrossRef] [PubMed]
- Wittner, M.; di Stefano, A.; Wangermann, P.; Nitschke, R.; Greger, R.; Bailly, C.; Amiel, C.; Roinel, N.; de Rouffignac, C. Differential effects of ADH on sodium, chloride, potassium, calcium and magnesium transport in cortical and medullary thick ascending limbs of mouse nephron. Pflug. Arch. 1988, 412, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Quamme, G.A. Control of magnesium transport in the thick ascending limb. Am. J. Physiol. Ren. Physiol. 1989, 256, F197–F210. [Google Scholar] [CrossRef]
- Di Stefano, A.; Wittner, M.; Nitschke, R.; Braitsch, R.; Greger, R.; Bailly, C.; Amiel, C.; Roinel, N.; de Rouffignac, C. Effects of parathyroid hormone and calcitonin on Na+, Cl−, K+, Mg2+ and Ca2+ transport in cortical and medullary thick ascending limbs of mouse kidney. Pflug. Arch. 1990, 417, 161–167. [Google Scholar] [CrossRef] [PubMed]
- De Baaij, J.H.F.; Hoenderop, J.G.J.; Bindels, R.J.M. Magnesium in Man: Implications for Health and Disease. Physiol. Rev. 2015, 95, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Franken, G.A.C.; Adella, A.; Bindels, R.J.M.; de Baaij, J.H.F. Mechanisms coupling sodium and magnesium reabsorption in the distal convoluted tubule of the kidney. Acta Physiol. 2021, 231, e13528. [Google Scholar] [CrossRef]
- Li, M.; Jiang, J.; Yue, L. Functional Characterization of Homo- and Heteromeric Channel Kinases TRPM6 and TRPM7. J. Gen. Physiol. 2006, 127, 525–537. [Google Scholar] [CrossRef]
- Günther, T.; Vormann, J.; Höllriegl, V. Characterization of Na+-dependent Mg2+ efflux from Mg2+-loaded rat erythrocytes. Biochim. Biophys. Acta BBA Biomembr. 1990, 1023, 455–461. [Google Scholar] [CrossRef]
- Ruat, M.; Snowman, A.M.; Hester, L.D.; Snyder, S.H. Cloned and Expressed Rat Ca2+-sensing Receptor: Differential Cooperative Responses to Calcium and Magnesium. J. Biol. Chem. 1996, 271, 5972–5975. [Google Scholar] [CrossRef] [PubMed]
- Bräuner-Osborne, H.; Jensen, A.A.; Sheppard, P.O.; O’Hara, P.; Krogsgaard-Larsen, P. The Agonist-Binding Domain of the Calcium-Sensing Receptor Is Located at the Amino-terminal Domain. J. Biol. Chem. 1999, 274, 18382–18386. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M.; Chen, C.J. Calcium, magnesium and the control of PTH secretion. Bone Miner. 1989, 5, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Riccardi, D.; Valenti, G. Localization and function of the renal calcium-sensing receptor. Nat. Rev. Nephrol. 2016, 12, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Murayama, Y.; Morel, F.; Le Grimellec, C. Phosphate, calcium and magnesium transfers in proximal tubules and loops of henle, as measured by single nephron microperfusion experiments in the rat. Pflug. Arch. 1972, 333, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A. Regulation of calcium reabsorption along the rat nephron: A modeling study. Am. J. Physiol. Ren. Physiol. 2015, 308, F553–F566. [Google Scholar] [CrossRef]
- Toka, H.R.; Al-Romaih, K.; Koshy, J.M.; DiBartolo, S.I.; Kos, C.H.; Quinn, S.J.; Curhan, G.C.; Mount, D.B.; Brown, E.M.; Pollak, M.R. Deficiency of the Calcium-Sensing Receptor in the Kidney Causes Parathyroid Hormone–Independent Hypocalciuria. J. Am. Soc. Nephrol. 2012, 23, 1879. [Google Scholar] [CrossRef]
- Wang, W.H.; Lu, M.; Hebert, S.C. Cytochrome P-450 metabolites mediate extracellular Ca2+-induced inhibition of apical K+ channels in the TAL. Am. J. Physiol. Cell Physiol. 1996, 271, C103–C111. [Google Scholar] [CrossRef]
- Wang, W.; Lu, M.; Balazy, M.; Hebert, S.C. Phospholipase A2 is involved in mediating the effect of extracellular Ca2+ on apical K+ channels in rat TAL. Am. J. Physiol. Ren. Physiol. 1997, 273, F421–F429. [Google Scholar] [CrossRef]
- Bazúa-Valenti, S.; Rojas-Vega, L.; Castañeda-Bueno, M.; Barrera-Chimal, J.; Bautista, R.; Cervantes-Pérez, L.G.; Vázquez, N.; Plata, C.; Murillo-de-Ozores, A.R.; González-Mariscal, L.; et al. The Calcium-Sensing Receptor Increases Activity of the Renal NCC through the WNK4-SPAK Pathway. J. Am. Soc. Nephrol. 2018, 29, 1838. [Google Scholar] [CrossRef]
- Hu, R.; McDonough, A.A.; Layton, A.T. Functional implications of the sex differences in transporter abundance along the rat nephron: Modeling and analysis. Am. J. Physiol. Ren. Physiol. 2019, 317, F1462–F1474. [Google Scholar] [CrossRef] [PubMed]
- Van Angelen, A.A.; van der Kemp, A.W.; Hoenderop, J.G.; Bindels, R.J. Increased expression of renal TRPM6 compensates for Mg2+ wasting during furosemide treatment. Nephrol. Dial. Transplant. Plus 2012, 5, 535–544. [Google Scholar] [CrossRef]
- Devane, J.; Ryan, M. Diuretics and magnesium excretion. Magnes. Bull. 1981, 3, 122–123. [Google Scholar]
- Ryan, M.P.; Devane, J.; Ryan, M.F.; Counihan, T.B. Effects of diuretics on the renal handling of magnesium. Drugs 1984, 28, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-T.; Shang, S.; Lai, L.-W.; Yong, K.-C.; Lien, Y.-H.H. Effect of thiazide on renal gene expression of apical calcium channels and calbindins. Am. J. Physiol. Ren. Physiol. 2004, 287, F1164–F1170. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, L.S. Comparison of calcium and sodium transport in early and late rat distal tubules: Effect of amiloride. Am. J. Physiol. Ren. Physiol. 1984, 246, F937–F945. [Google Scholar] [CrossRef]
- Devane, J.; Ryan, M.P. The effects of amiloride and triamterene on urinary magnesium excretion in conscious saline-loaded rats. Br. J. Pharmacol. 1981, 72, 285. [Google Scholar] [CrossRef]
- Devane, J.; Ryan, M.P. Dose-dependent reduction in renal magnesium clearance by amiloride during frusemide-induced diuresis in rats. Br. J. Pharmacol. 1983, 80, 421. [Google Scholar] [CrossRef]
- Friedman, P.A.; Gesek, F.A. Stimulation of calcium transport by amiloride in mouse distal convoluted tubule cells. Kidney Int. 1995, 48, 1427–1434. [Google Scholar] [CrossRef]
- Chubanov, V.; Ferioli, S.; Wisnowsky, A.; Simmons, D.G.; Leitzinger, C.; Einer, C.; Jonas, W.; Shymkiv, Y.; Bartsch, H.; Braun, A.; et al. Epithelial magnesium transport by TRPM6 is essential for prenatal development and adult survival. eLife 2016, 5, e20914. [Google Scholar] [CrossRef]
- Mittermeier, L.; Demirkhanyan, L.; Stadlbauer, B.; Breit, A.; Recordati, C.; Hilgendorff, A.; Matsushita, M.; Braun, A.; Simmons, D.G.; Zakharian, E.; et al. TRPM7 is the central gatekeeper of intestinal mineral absorption essential for postnatal survival. Proc. Natl. Acad. Sci. USA 2019, 116, 4706–4715. [Google Scholar] [CrossRef]
- Verschuren, E.H.; Hoenderop, J.G.; Peters, D.J.; Arjona, F.J.; Bindels, R.J. Tubular flow activates magnesium transport in the distal convoluted tubule. FASEB J. 2019, 33, 5034–5044. [Google Scholar] [CrossRef]
- Bailly, C.; Roinel, N.; Amiel, C. PTH-like glucagon stimulation of Ca and Mg reabsorption in Henle’s loop of the rat. Am. J. Physiol. Ren. Physiol. 1984, 246, F205–F212. [Google Scholar] [CrossRef]
- Quamme, G.A.; de Rouffignac, C. Renal magnesium handling. In The Kidney: Physiology and Pathophysiology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2000; p. 375. [Google Scholar]
- O’Grady, S.M.; Palfrey, H.C.; Field, M. Characteristics and functions of Na-K-Cl cotransport in epithelial tissues. Am. J. Physiol. Cell Physiol. 1987, 253, C177–C192. [Google Scholar] [CrossRef]
- Burg, M.B. Tubular chloride transport and the mode of action of some diuretics. Kidney Int. 1976, 9, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, A.; Roinel, N.; de Rouffignac, C.; Wittner, M. Transepithelial Ca2+ and Mg2+ Transport in the Cortical Thick Ascending Limb of Henle’s Loop of the Mouse Is a Voltage-Dependent Process. Ren. Physiol. Biochem. 2008, 16, 157–166. [Google Scholar] [CrossRef]
- Quamme, G.A. Effect of furosemide on calcium and magnesium transport in the rat nephron. Am. J. Physiol. Ren. Physiol. 1981, 241, F340–F347. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.G. Effects of ethacrynic acid and furosemide on urinary calcium, phosphate and magnesium. Metabolism 1968, 17, 867–876. [Google Scholar] [CrossRef]
- Leary, W.P.; Reyes, A.J.; Wynne, R.D.; van der Byl, K. Renal Excretory Actions of Furosemide, of Hydrochlorothiazide and of the Vasodilator Flosequinan in Healthy Subjects. J. Int. Med. Res. 1990, 18, 120–141. [Google Scholar] [CrossRef] [PubMed]
- Brickman, A.S.; Massry, S.G.; Coburn, J.W. Changes in Serum and Urinary Calcium during Treatment with Hydrochlorothiazide: Studies on Mechanisms. J. Clin. Investig. 1972, 51, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Kroenke, K.; Wood, D.R.; Hanley, J.F. The Value of Serum Magnesium Determination in Hypertensive Patients Receiving Diuretics. Arch. Intern. Med. 1987, 147, 1553–1556. [Google Scholar] [CrossRef]
- Stadt, M.M.; Layton, A.T. Adaptive changes in single-nephron GFR, tubular morphology, and transport in a pregnant rat nephron: Modeling and analysis. Am. J. Physiol. Ren. Physiol. 2022, 322, F121–F137. [Google Scholar] [CrossRef]
- Stadt, M.M.; West, C.A.; Layton, A.T. Effect of pregnancy and hypertension on kidney function in female rats: Modeling and functional implications. PLoS ONE 2023, 18, e0279785. [Google Scholar] [CrossRef]
- Layton, A.T.; Vallon, V.; Edwards, A. Predicted consequences of diabetes and SGLT inhibition on transport and oxygen consumption along a rat nephron. Am. J. Physiol. Ren. Physiol. 2016, 310, F1269–F1283. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Layton, A. A computational model of kidney function in a patient with diabetes. Int. J. Mol. Sci. 2021, 22, 5819. [Google Scholar] [CrossRef]
- Swapnasrita, S.; Carlier, A.; Layton, A.T. Sex-specific computational models of kidney function in patients with diabetes. Front. Physiol. 2022, 13, 741121. [Google Scholar] [CrossRef]
- Layton, A.T.; Edwards, A.; Vallon, V. Renal potassium handling in rats with subtotal nephrectomy: Modeling and Analysis. Am. J. Physiol. Ren. Physiol. 2017, 314, F643–F657. [Google Scholar] [CrossRef]
- Stadt, M.M.; Layton, A.T. Mathematical modeling of calcium homeostasis in female rats: An analysis of sex differences and maternal adaptations. J. Theor. Biol. 2023, 572, 111583. [Google Scholar] [CrossRef] [PubMed]
- Stadt, M.M.; Leete, J.; Devinyak, S.; Layton, A.T. A mathematical model of potassium homeostasis: Effect of feedforward and feedback controls. PLoS Comput. Biol. 2022, 18, e1010607. [Google Scholar] [CrossRef] [PubMed]
- Leete, J.; Layton, A.T. Sex-specific long-term blood pressure regulation: Modeling and analysis. Comput. Biol. Med. 2019, 104, 139–148. [Google Scholar] [CrossRef]
- Ahmed, S.; Layton, A.T. Sex-specific computational models for blood pressure regulation in the rat. Am. J. Physiol. Ren. Physiol. 2020, 318, F888–F900. [Google Scholar] [CrossRef] [PubMed]
- Funato, Y.; Furutani, K.; Kurachi, Y.; Miki, H. CrossTalk proposal: CNNM proteins are Na+/Mg2+ exchangers playing a central role in transepithelial Mg2+ (re)absorption. J. Physiol. 2018, 596, 743. [Google Scholar] [CrossRef] [PubMed]
- Arjona, F.J.; de Baaij, J.H. CrossTalk opposing view: CNNM proteins are not Na+/Mg2+ exchangers but Mg2+ transport regulators playing a central role in transepithelial Mg2+ (re)absorption. J. Physiol. 2018, 596, 747. [Google Scholar] [CrossRef] [PubMed]
- Sponder, G.; Mastrototaro, L.; Kurth, K.; Merolle, L.; Zhang, Z.; Abdulhanan, N.; Smorodchenko, A.; Wolf, K.; Fleig, A.; Penner, R.; et al. Human CNNM2 is not a Mg2+ transporter per se. Pflug. Arch. Eur. J. Physiol. 2016, 468, 1223–1240. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dutta, P.; Layton, A.T. Magnesium and Calcium Transport along the Male Rat Kidney: Effect of Diuretics. Math. Comput. Appl. 2024, 29, 13. https://doi.org/10.3390/mca29010013
Dutta P, Layton AT. Magnesium and Calcium Transport along the Male Rat Kidney: Effect of Diuretics. Mathematical and Computational Applications. 2024; 29(1):13. https://doi.org/10.3390/mca29010013
Chicago/Turabian StyleDutta, Pritha, and Anita T. Layton. 2024. "Magnesium and Calcium Transport along the Male Rat Kidney: Effect of Diuretics" Mathematical and Computational Applications 29, no. 1: 13. https://doi.org/10.3390/mca29010013
APA StyleDutta, P., & Layton, A. T. (2024). Magnesium and Calcium Transport along the Male Rat Kidney: Effect of Diuretics. Mathematical and Computational Applications, 29(1), 13. https://doi.org/10.3390/mca29010013