Abstract
Introduction: Pseudomonas aeruginosa is a common agent causing community acquired and nosocomial respiratory tract infections, with particularly life-threatening manifestations in patients who are immunocompromised of who have cystic fibrosis. This study investigated the occurrence of extended-spectrum β-lactamases (ESBLs) and metallo β-lactamase (MBL) in association with important putative virulence genes and genotypes variation among P. aeruginosa isolates from respiratory tract infection of Jordanian patients. Methods: Over a period of 8-month, a total of 284 respiratory tract samples were obtained from patients diagnosed with respiratory tract infection while attending the Pulmonary Clinic/Intensive Care Unit, Jordan University Hospital (JUH). At the time of sampling most were inpatients (86.9%). Samples were cultured specifically for P. aeruginosa. Results: A total of 61/284 (21.5%) P. aeruginosa isolates were recovered from respiratory samples of patients. The percentage of MDR P. aeruginosa isolates was 52.5%, and all isolates were susceptible to colistin with lower rates of susceptibility to other tested antibiotics. Positive genes of blaCTX-M, blaVEB, blaTEM, blaGES and blaSHV were detected in 68.9%, 18.9%, 18.9%, 15.6% and 12.5% of isolates, respectively. Genotyping revealed no significant genetic relationship among MDR P. aeruginosa isolates from hospitalized patients as judged by the constructed dendrogram and the presence of 14 genotypic groups. The percentages of the virulence genes algD, lasB, toxA, exoS, and exoU among P. aeruginosa isolates were 98%, 98%, 80%, 33% and 33%, respectively, and 87% of isolates produced pyocyanin. Conclusion: The present study demonstrates high occurrence of MDR P. aeruginosa isolates carrying blaCTX-M genes. No specific associations were found between antibiotic resistance, virulence genes and genotypes among MDR isolates.
Introduction
Pseudomonas aeruginosa respiratory infections are causing high morbidity and mortality due to the germ’s capacity to rapidly develop antibiotic resistance, especially during antibiotic treatment of patients. The organism can carry a variety of putative virulence factors, which are highly controlled by cell-to-cell signaling systems [1,2].
P. aeruginosa can secrete an exopolysaccharide called alginate (algD) and an elastase B (lasB) enzyme in response to environmental conditions. Both these virulence factors can influence pathogenesis by enhancing adhesion, colonization, and invasion of tissues, causing chronic pulmonary inflammation [3]. Additionally, elastase B is an important protease of P. aeruginosa. This enzyme has a tissue-damaging activity, it can degrade a number of plasma proteins and it contributes to the survival of P. aeruginosa in infected tissues [4].
Other potential virulence factors secreted by P. aeruginosa that are important in its pathogenicity include exotoxin A (toxA), which is the most toxic virulence factor detected in this organism. It inhibits protein biosynthesis, it has a necrotizing activity on tissues and contributes to the colonization process [5]. Exoenzyme S (exoS) is secreted into the cytosol of epithelial cells and it contributes to increased pneumonia severity [6].
Infections caused by multidrug-resistant (MDR) P. aeruginosa are difficult to treat and can be associated with high mortality especially in patients who are immunocompromised or who present chronic lung diseases. P. aeruginosa acquires resistance primarily through the production of plasmid mediated AmpC β-lactamase, different types of extended-spectrum β-lactamases (ESBLs) and metallo-β-lactamases (MBLs) [7,8]. The recent emergence of bla-KPC enzymes in P. aeruginosa strains, currently KPC-2 and KPC-5, is alarming since these strains show activity against most β-lactams including monobactams, carbapenems, and oxyimino-cephalosporins [8].
Recent studies report that ESBLs and MBLs are increasing among P. aeruginosa in the Middle East Arab countries, and this development could result in prolonged hospital stay, increased cost and fatal outcome among patients [9,10].
This study investigated the occurrence and relationship of MDR P. aeruginosa recovered from Jordanian patients with respiratory tract infection, in association with putative virulence genes and genotypes.
Methods
Patients
This prospective convenience sampling study included a total of 284 respiratory tract samples obtained from adult patients attending the Pulmonary Clinic, The Jordan University Hospital (JUH), over the period from November 2014 to June 2015. JUH includes 550 beds, and is a tertiary care teaching hospital with 24 of the beds distributed in 3 adult Intensive Care Units (ICUs).
Only one sample was collected from each patient with signs and symptoms of respiratory tract infection. The majority of specimens (247; 86.9%) were obtained from hospitalized patients and only 37 (13.1%) were collected from outpatients. Bio-data of each patient was obtained and registered on a special sheet and included the following: patient’s identification number, gender, age, clinical diagnosis, specific admission conditions, type of specimen, type of current or previous antibiotic treatment within the past 4 weeks prior to specimen sampling.
The study was approved by the School of Medicine and the School of Graduate studies, The Jordan. A permission was also obtained from the Institutional Ethical Review Board (IERB) at the Jordan University Hospital (JUH, Permission No. 9/2015). Verbal consent was obtained from all patients after explaining the aim of the study. The identity of patients was protected by applying numbers instead of names for all collected specimens.
Culture, isolation and bacterial identification
All collected respiratory samples (sputum, bronchoalveolar lavage [BAL], pleural fluid) were inoculated directly using cotton swab onto Pseudomonas CN agar plates (Oxoid, Basingstoke, United Kingdom) and incubated for 24-48 hours at 37 °C. The culture plates were examined to detect the presence of straw-colored colonies with yellow or blue-green pigmentation, and colonies were confirmed later as being P. aeruginosa using the following preliminary characteristics: positive oxidase test, growth at 42 °C, negative lactose, and glucose fermentation in tube of Kligler iron agar. Five colonies of pure P. aeruginosa were subcultured into cysteine lactose electrolyte deficient agar plates (CLED, Oxoid), and all isolates were stored in cryotubes containing brain-heart infusion agar with 15% glycerol at -70 °C for further investigation.
Antimicrobial susceptibility
Antimicrobial susceptibility of P. aeruginosa isolates was assessed using disc diffusion test according to the recommendation of the Clinical Laboratory and Standards Institute (CLSI, 2015) [11]. P. aeruginosa isolates which showed resistance to three or more antibiotic classes (MDR) were tested for minimum inhibitory concentrations (MICs) by E-test strips for ceftazidime, ciprofloxacin, colistin, imipenem and meropenem (Oxoid). The results of susceptibility tests were interpreted according to the breakpoints of tested antibiotics as recommended in CLSI guidelines (2015). P. aeruginosa ATCC 27853 was used as control during all tests.
Plasmid DNA extraction and PCR procedures
After thawing, stored P. aeruginosa isolates were subcultured onto blood agar, and incubated at 37 °C for 24 hours. A few colonies were picked from the agar, and inoculated into 3 mL Luria Broth (LB) (Oxoid) and incubated for 24 hours at 37 °C. The bacterial plasmid was extracted using the Zyppy TM Plasmid Miniprep Kit, Zymo (QIAGEN, Germantown, MD, USA) according to manufacturer’s instructions. The quantity of DNA from each isolate extract was tested to ensure that the DNA concentration was more than 50 μg/mL-1 with a purity at A260/A280=1.6-1.8, using a spectrophotometer (Bio-Rad, Hercules, CA, USA).
Identification of P. aeruginosa by PCR
Specific identification of P. aeruginosa was performed by nested PCR using a first primer set for identification of the genus Pseudomonas, followed by a second primer set specific for P. aeruginosa. The 16 ribosomal DNA (rDNA) was detected as described by Spilker et al. as shown in Table 1 [12].

Table 1.
List of all primers, their product sizes and references used for detection of ESBLs, MBLs and virulence genes.
Detection of virulence genes by PCR
The following genes: algD, lasB, toxA, exoS and exoU were detected using specific primers for each gene as described in references listed in Table 1 [6,13,14,15,16,17,18,19,20]. P. aeruginosa (PAO-1 strain) was used as a positive control for lasB and toxA genes and P. aeruginosa ATCC 27853 was used as a positive control for algD gene, and nuclease free water was used as a negative control.
Detection of genes of ESBLs, MBLs and blaKPC
A total of 32 MDR P. aeruginosa isolated were investigated for genes of blaCTX-M, blaTEM, blaVEB-1, blaSHV, blaGES-1 and MBLs (IMP-1, VIM-2, blaKPC), as presented in Table 1. A positive control of E. coli (ATCC 51446) was used for detection of blaCTX-M genes, P. aeruginosa (CL 120 & CL 162, Université Libanaise, Tripoli, Lebanon) for IMP-1, VIM-2 genes, and K. pneumoniae (ATCC BAA-1705) for blaKPC genes. Nuclease free water was used as a negative control in all tests.
Gel electrophoresis
In all tests, 6 µL of each amplified PCR product, controls and 100 bp ladder (molecular size marker) were analyzed in 1.5% agarose gel (Promega, Madison, WI, USA) in 1X TBE buffer containing ethidium bromide, and run for 50 minutes at 120 volts using horizontal apparatus. After electrophoresis, the DNA bands in gel were visualized under ultraviolet light (UVP system, Upland, CA, USA).
RAPD-PCR to detect genotypic variations among P. aeruginosa isolates
Two short (10 mers) single primers of 272 (5-AGCGGGCCAA-3) and 208 (5-ACGGCCGACC-3) were used in two separated PCR reactions to create a repetitive electrophoresis patterns and to detect the genotypic variation according to the number and weight of detected band differences among the isolates [21]. The results were analyzed by computer with PyElph software (USA).
Statistical analysis
Data generated from the study were tabulated as Microsoft Excel sheet and uploaded to Statistical Package for Social Sciences version 20 (IBM Corp, Armonk, NY, USA). Frequencies and percentages were calculated for categorical data and Pearson’s chi-squared test or Fisher’s exact test were applied to determine potential factors associated with P. aeruginosa and to determine whether there are any statistical differences between groups. The level of significance was set at a p value of 0.05 to test the hypothesis of no association. Fisher’s exact test replaces chi-squared test when the minimum expected count is less than five.
Results
Patients
A total of 61/284 (21.5%) P. aeruginosa isolates were recovered from respiratory samples of patients. These included 247 (86.9%) hospitalized patients and 37 (13.1%) non-hospitalized patients. Recovery of P. aeruginosa was significantly more frequent (p=0.036) in hospitalized (73.8%) versus non-hospitalized patients (26.2%). Table 2 presents the demographic and clinical characteristics of the 61 investigated patients. Table 3 shows demographic characteristics and clinical features of all patients infected with P. aeruginosa and hospitalized in wards (60%) and ICUs (40%). All clinical features were almost similar in both groups. Only one female patient hospitalized in the ICU died during the study period.
Table 2.
Demographic characteristics of the 61 investigated patients and their clinical features in association with positive P. aeruginosa.
Table 3.
Demographic characteristics of the 45 hospitalized patients in clinical wards and intensive care units and their clinical features in association with positive P. aeruginosa.
Occurrence of P. aeruginosa and antimicrobial susceptibility profiles
The identity of 61 P. aeruginosa isolates was confirmed by biochemical testing and PCR. A total of 53 (87%) isolates showed clear blue-green pigment production (pyocyanin) on Pseudomonas CN agar. The antimicrobial susceptibility patterns and MICs of P. aeruginosa isolates for important used antibiotics are shown in Table 4. A total of 32 (52.5%) isolates were completely resistant to three or more antibiotic classes (MDR) and all were recovered from hospitalized patients (Table 4).
Table 4.
Antimicrobial susceptibility pattern of 61 P. aeruginosa isolates*.
MDR and putative virulence genes
All 32 MDR P. aeruginosa isolates carried one to four of the investigated ESBL genes and all were positive for genes of algD, lasB, toxA and pyocyanin, as shown in Table 5. Most MDR isolates (22/32; 68.9%) harbored blaCTX-M genes, whereas other ESBL genes were much less common among the isolates, and none of the MDR P. aeruginosa isolates was positive for the blaIMP-1 or blaKPC genes (Table 5).
Table 5.
Distribution of ESBL and MBL genes in association with potential virulence genes among 32 MDR P. aeruginosa isolates.
The most common virulence genes detected among the all 61 isolates were algD and lasB (98%), followed by toxA (80%), exoS and exoU (33), and 87% were positive for pyocyanin (Table 6).
Table 6.
Distribution of potential virulence genes among 61 P. aeruginosa isolates.
Genotyping
The RAPD profile showed high diversity among the isolates. Each isolate generated 2-6 fragments with band sizes ranging from 180 to 1200 bp. The dendrogram of the isolates is presented in Figure 1; by applying 70% similarity index of dendrogram, the isolates were clustered into 14 different groups. The major group was presented by 5 isolates and all other groups were presented with less than 4 isolates. There was no significant association between genotype groups and antimicrobial resistance phenotypes using RAPD cluster groups.
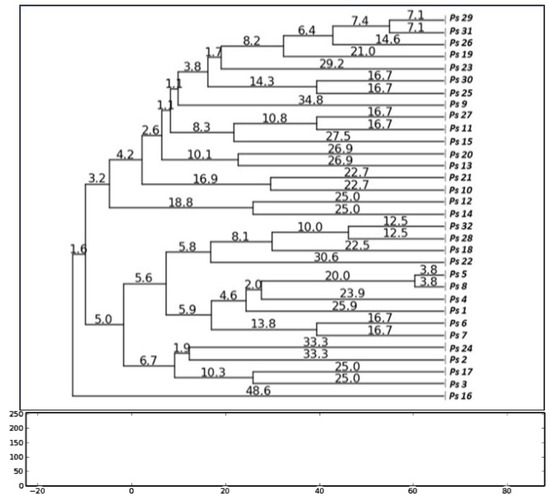
Figure 1.
Dendrogram based on RAPD results showing relatedness between 32 MDR P. aeruginosa isolates; the dendrogram was generated by the neighbor joining method with PyElph software (USA).
Discussion
The general rate of P. aeruginosa isolates recovery in this study (21.5%) is relatively in the middle of other recently reported studies from neighboring countries such as Egypt, Saudi Arabia, United Arab Emirates, where the isolation rates of P. aeruginosa from sputum samples of patients with lower respiratory tract infection ranged between 16.7% and 50% [9,21,22,23]. A study from the United Arab Emirates reported that the most common pathogen isolated from the sputum of patients with hospital-acquired pneumonia was P. aeruginosa (50%), and that it caused a mortality rate of 24% [23]. A prevalence study conducted in ICU patients for one day in 72 countries around the world has also demonstrated that P. aeruginosa was one of the most common organisms, and that it accounted for 19.9% of all organisms isolated from respiratory tract cultures [24].
Numerous studies reported that nosocomially-acquired P. aeruginosa isolates tend to be often more MDR and commonly identified ICU patients [9,24,25,26]. The rate of MDR P. aeruginosa isolates in our study was 52.2%, and this result is similar to recently reported studies from Jordan and neighboring Arab countries (Egypt, Saudi Arabia, Lebanon) [23,25,26,27].
The present study showed that hospitalized patients were significantly (p=0.036) more frequently infected with P. aeruginosa than non-hospitalized patients in association with lower respiratory tract infection. This study also showed that almost all 61 patients with positive P. aeruginosa isolates had purulent sputum, cough and were treated with antibiotics (96.7%), whereas other significant clinical respiratory infection features such as chest pain, dyspnea and hemoptysis were less frequently observed in our patients. There was only one death in patients with lower respiratory infection during the study period.
The present study demonstrates that MDR P. aeruginosa isolates (68.9%) commonly are ESBL-producers and blaCTX-M was the most frequent phenotype detected in these isolates, whereas the other types of TEM, VEB, GES, and SHV genes were less often detected (Table 5). The spread of ESBL-producing Gram-negative bacteria is considered an important driver for the use of carbapenems, which can increase the selection of carbapenemase-producing organisms causing infections, particularly in ICU patients [28].
Most recent studies reported that hospital outbreaks caused by MDR P. aeruginosa clones with MBLs such as VIM and IMP types can be developed by either chromosomal mutations or horizontal gene transfer [29]. VIMs are the most frequent MBL type found among members of the Enterobacteriaceae family, as well as among Pseudomonas spp. and Acinetobacter spp. [29]. The present study indicated that MBLs were only detected as type VIM-2 in a few of our MDR P. aeruginosa isolates (9.4%), while the other types, blaIMP and blaKPC, were absent (Table 5). In Jordan, a recent study has also reported that all Acinetobacter baumannii isolates from the hospital, environment, and patients’ respiratory tract were negative for blaVIM and blaIMP-1 genes [30].
The pathogenic potential of P. aeruginosa is complex and multifactorial, and virulence genes, especially algD, lasB, toxA and exoS can induce biofilm formation, infection and cytotoxicity in the human body, especially in the respiratory tract [6,31]. Additionally, P. aeruginosa strains carrying exoU genes can cause a more virulent form of pneumonia than other strains [32]. The prevalences of the virulence genes algD, lasB, toxA and secretion of pyocyanin in this study were very high among P. aeruginosa isolates (98%-80%), and all MDR isolates carried those 4 virulence factors, while both exoS and exoU genes were found less frequently and only in 33% of the isolates (Table 6). A recent study indicated that pyocyanin production was higher in earlier P. aeruginosa isolates from the respiratory tract compared to the isolates obtained later in the course of infection [33]. It has been also reported that pyocyanin induces various antagonistic effects on the host, especially pro-inflammatory and free radical effects which cause cellular damage and death [33].
The relationship between antimicrobial resistance markers and distribution of common virulence factors among P. aeruginosa isolates in this study was not significant, since all MDR isolates and most of the susceptible isolates carried 3 potential virulence genes (algD, lasB and toxA) and secreted pyocyanin (Table 5). A French study detected the virulence genes algD, lasB and toxA in all P. aeruginosa isolates from the sputa of patients with cystic fibrosis and, to a lesser rate, from other non-cystic fibrosis isolates [3]. A Bulgarian study reported that P. aeruginosa isolates from all clinical samples including the respiratory tract carried 100% lasB genes and a high rate of algD genes (95.3%) in cystic fibrosis cases [23]. A study from Iran demonstrated also that all P. aeruginosa isolates from the pulmonary tract harbored significantly higher rates of toxA genes than wound and burn isolates (p<0.004), and other virulence genes such as algD and lasB were less common in respiratory tract specimens [4].
It can be concluded from our study and other studies that virulence genes lasB, algD, and toxA are mostly associated with P. aeruginosa isolates from the pulmonary tract. However, none of the mentioned studies has previously reported on the association between antimicrobial resistance and virulence potential of their P. aeruginosa isolates as it has been done in this study.
RAPD fingerprinting of 32 MDR P. aeruginosa isolates from hospitalized patients and the construction of the dendrogram of the isolates demonstrated high diversity among P. aeruginosa isolates, with 14 genotype groups, by applying 70% similarity index (Figure 1). Association between the antimicrobial resistance phenotypes and genotype profiles of these 32 isolates revealed a limited similarity of DNA bands patterns between phenotypes and genotypes. Based on these results, it can be suggested that most isolates originated from various clones and there was no single genotype prevalent among more than 5 isolates.
Conclusions
In conclusion, this study has found no significant genetic relationship among MDR P. aeruginosa isolates from the respiratory tract of hospitalized patients or their carriage of virulence genes.
Author Contributions
HYAD performed all laboratory tests, NO supervised all clinical issues of patients, LFAQ helped in genotyping of the isolates, AAS supervised all laboratory tests and writing of the manuscript. All authors read and approved the final version of the manuscript.
Funding
This work was supported by a grant from The Jordan University, Dean of Research (No. 74/2014-2015).
Acknowledgments
The authors are thankful to the Jordan University Hospital for allowing them to collect clinical specimens and data from the admitted patients. Also, the authors thank Mrs. Ilham Abu-Khader for performing the statistical analysis of the study.
Conflicts of Interest
All authors – none to disclose.
References
- Nathwani, D.; Raman, G.; Sulham, K.; Gavaghan, M.; Menon, V. Clinical and economic consequences of hospital-acquired resistant and multidrug-resistant Pseudomonas aeruginosa infections: a systematic review and meta-analysis. Antimicrob Resist Infect Control 2014, 3, 32. [Google Scholar] [CrossRef]
- Mitov, I.; Strateva, T.; Markova, B. Prevalence of virulence genes among Bulgarian nosocomial and cystic fibrosis isolates of Pseudomonas aeruginosa. Braz J Microbiol 2010, 41, 588–595. [Google Scholar] [CrossRef]
- Lanotte, P.; Watt, S.; Mereghetti, L.; et al. Genetic features of Pseudomonas aeruginosa isolates from cystic fibrosis patients compared with those of isolates from other origins. J Med Microbiol 2004, 53, 73–81. [Google Scholar] [CrossRef]
- Nikbin, V.S.; Aslani, M.M.; Sharafi, Z.; Hashemipour, M.; Shahcheraghi, F.; Ebrahimipour, G.H. Molecular identification and detection of virulence genes among Pseudomonas aeruginosa isolated from different infectious origins. Iran J Microbiol 2012, 4, 118–123. [Google Scholar]
- Michalska, M.; Wolf, P. Pseudomonas exotoxin A: optimized by evolution for effective killing. Front Microbiol 2015, 6, 963. [Google Scholar] [CrossRef]
- Shaver, C.M.; Hauser, A.R. Relative contributions of Pseudomonas aeruginosa ExoU, ExoS, and ExoT to virulence in the lung. Infect Immun 2004, 72, 6969–6977. [Google Scholar] [CrossRef] [PubMed]
- Farshadzadeh, Z.; Khosravi, A.D.; Alavi, S.M.; Parhizgari, N.; Hoveizavi, H. Spread of extended-spectrum β-lactamase genes of blaOXA-10, blaPER-1 and blaCTX-M in Pseudomonas aeruginosa strains isolated from burn patients. Burns 2014, 40, 1575–1580. [Google Scholar] [CrossRef]
- Poirel, L.; Nordmann, P.; Lagrutta, E.; Cleary, T.; Munoz-Price, L.S. Emergence of KPC-producing Pseudomonas aeruginosa in the United States. Antimicrob Agents Chemother 2010, 54, 3072. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.A.; Eldaly, O.; Jiman-Fatani, A.; Mohamed, M.L.; Ibrahim, E.M. Epidemiological characterization of P. aeruginosa isolates of intensive care units in Egypt and Saudi Arabia. East Mediterr Health J 2013, 19, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Hamze, M.; Mallat, H.; Dabboussi, F.; Achkar, M. Antibiotic susceptibility and serotyping of clinical Pseudomonas aeruginosa isolates in northern Lebanon. Int Arabic J Antimicrob Agents 2012, 2, 1–6. [Google Scholar]
- Clinical Laboratory and Standards Institute (CLSI). Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically; approved standard – tenth edition. CLSI document M07-A10 Villanova, PA, USA: CLSI. 2015. [Google Scholar]
- Spilker, T.; Coenye, T.; Vandamme, P.; LiPuma, J.J. PCR-based assay for differentiation of Pseudomonas aeruginosa from other Pseudomonas species recovered from cystic fibrosis patients. J Clin Microbiol 2004, 42, 2074–2079. [Google Scholar] [CrossRef]
- Bokaeian, M.; Shahraki Zahedani, S.; Soltanian Bajgiran, M.; Ansari Moghaddam, A. Frequency of PER, VEB, SHV, TEM and CTX-M genes in resistant strains of Pseudomonas aeruginosa producing extended spectrum β-lactamases. Jundishapur J Microbiol 2014, 8, e13783. [Google Scholar] [CrossRef]
- Pitout, J.D.D.; Gregson, D.B.; Poirel, L.; McClure, J.A.; Le, P.; Church, D.L. Detection of Pseudomonas aeruginosa producing metallo-β-lactamases in a large centralized laboratory. J Clin Microbiol 2005, 43, 3129–3135. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, Z.; Li, M.; Zhou, D.; Ruan, F.; Lu, Y. Detection of extended-spectrum β-lactamases in clinical isolates of Pseudomonas aeruginosa. Antimicrob Agents Chemother 2006, 50, 2990–2995. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, N.; Momtaz, H. Virulence gene profiles of multidrug resistant Pseudomonas aeruginosa isolated from Iranian hospital infections. Iran Red Crescent Med J 2014, 16, e15722. [Google Scholar] [CrossRef]
- Monteiro, J.; Widen, R.H.; Pignatari, A.C.; Kubasek, C.; Silbert, S. Rapid detection of carbapenemase genes by multiplex real-time PCR. J Antimicrob Chemother 2012, 67, 906–909. [Google Scholar] [CrossRef]
- Akpaka, P.E.; Swanston, W.H.; Ihemere, H.N.; et al. Emergence of KPC-producing Pseudomonas aeruginosa in Trinidad and Tobago. J Clin Microbiol 2009, 47, 2670–2671. [Google Scholar] [CrossRef] [PubMed]
- Wolska, K.; Szweda, P. Genetic features of clinical Pseudomonas aeruginosa strains. Pol J Microbiol 2009, 58, 255–260. [Google Scholar] [PubMed]
- Mitov, I.; Strateva, T.; Markova, B. Prevalence of virulence genes among Bulgarian nosocomial and cystic fibrosis isolates of Pseudomonas aeruginosa. Braz J Microbiol 2010, 41, 588–595. [Google Scholar] [CrossRef]
- Nanvazadeh, F.; Khosravi, A.D.; Zolfaghari, M.R.; Parhizgari, N. Genotyping of Pseudomonas aeruginosa strains isolated from burn patients by RAPD-PCR. Burns 2013, 39, 1409–1413. [Google Scholar] [CrossRef]
- Fatima, A.; Naqvi, S.B.; Khaliq, S.A.; Perveen, S.; Jabeen, S. Antimicrobial susceptibility pattern of clinical isolates of Pseudomonas aeruginosa isolated from patients of lower respiratory tract infections. Springerplus 2012, 1, 70. [Google Scholar] [CrossRef]
- Al-Muhairi, S.; Zoubeidi, T.; Ellis, M.; Nicholls, M.G.; Safa, W.; Joseph, J. Demographics and microbiological profile of pneumonia in United Arab Emirates. Monaldi Arch Chest Dis 2006, 65, 13–18. [Google Scholar] [CrossRef]
- Vincent, J.L.; Rello, J.; Marshall, J.; et al. International study of the prevalence and outcomes of infection in intensive care units. JAMA 2009, 302, 2323–2329. [Google Scholar] [CrossRef] [PubMed]
- Al-Hajje, A.; Ezedine, M.; Hammoud, H.; et al. [Current status of nosocomial infections in the Lebanese Hospital Center, Beirut]. East Mediterr Health J 2012, 18, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Wassef, M.; El Mahallawy, H.; Zafer, M.M.; Ghaith, D.; Abdel Hamid, R. Lab based surveillance of multidrug resistant Pseudomonas aeruginosa in Cairo University Hospitals, Egypt. J Microbiol Exp 2015, 2, 00039. [Google Scholar]
- Masaadeh, H.A.; Jaran, A.S. Incident of Pseudomonas aeruginosa in post-operative wound infection. Am J Infect Dis 2009, 5, 1–6. [Google Scholar] [CrossRef]
- Voor In ‘t Holt, A.F.; Severin, J.A.; Lesaffre, E.M.; Vos, M.C. A systematic review and meta-analyses show that carbapenem use and medical devices are the leading risk factors for carbapenem-resistant Pseudomonas aeruginosa. Antimicrob Agents Chemother 2014, 58, 2626–2637. [Google Scholar] [CrossRef]
- Zhao, W.H.; Hu, Z.Q. Acquired metallo-β-lactamases and their genetic association with class 1 integrons and ISCR elements in Gram-negative bacteria. Future Microbiol 2015, 10, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Obeidat, N.; Jawdat, F.; Al-Bakri, G.A.; Shehabi, A.A. Major biologic characteristics of Acinetobacter baumannii isolates from hospital environmental and patients’ respiratory tract sources. Am J Infect Control 2014, 42, 401–404. [Google Scholar] [CrossRef]
- Moreau-Marquis, S.; Stanton, B.A.; O’Toole, G.A. Pseudomonas aeruginosa biofilm formation in the cystic fibrosis airway. Pulm Pharmacol Ther 2008, 21, 595–599. [Google Scholar] [CrossRef]
- Schulert, G.S.; Feltman, H.; Rabin, S.D.; Martin, C.G.; Battle, S.E.; Rello, J.; Hauser, A.R. Secretion of the toxin ExoU is a marker for highly virulent Pseudomonas aeruginosa isolates obtained from patients with hospital-acquired pneumonia. J Infect Dis 2003, 188, 1695–1706. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, C.C.; Chen, Y.; Goetzmann, H.S.; et al. Pseudomonas aeruginosa exotoxin pyocyanin causes cystic fibrosis airway pathogenesis. Am J Pathol 2009, 175, 2473–2488. [Google Scholar] [CrossRef] [PubMed]







© GERMS 2017.