


Immobilisation of Sucrase A from Bacillus subtilis on the Surface of Escherichia coli Mediated by the AIDA-I Autotransporter: Application on the Homolactic Fermentation

 , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Molecular Cloning of E. coli
2.2. Construction of the pAIDA-sacA
2.3. Detection of Saccharolytic Activity on Plate
2.4. Culture Media and Anaerobic Fermentation Conditions
2.5. Adaptive Evolution of E. coli Strains Under Glucose and Sucrose Conditions
2.6. Bioreactor Cultures
2.7. Statistical Analysis
3. Results
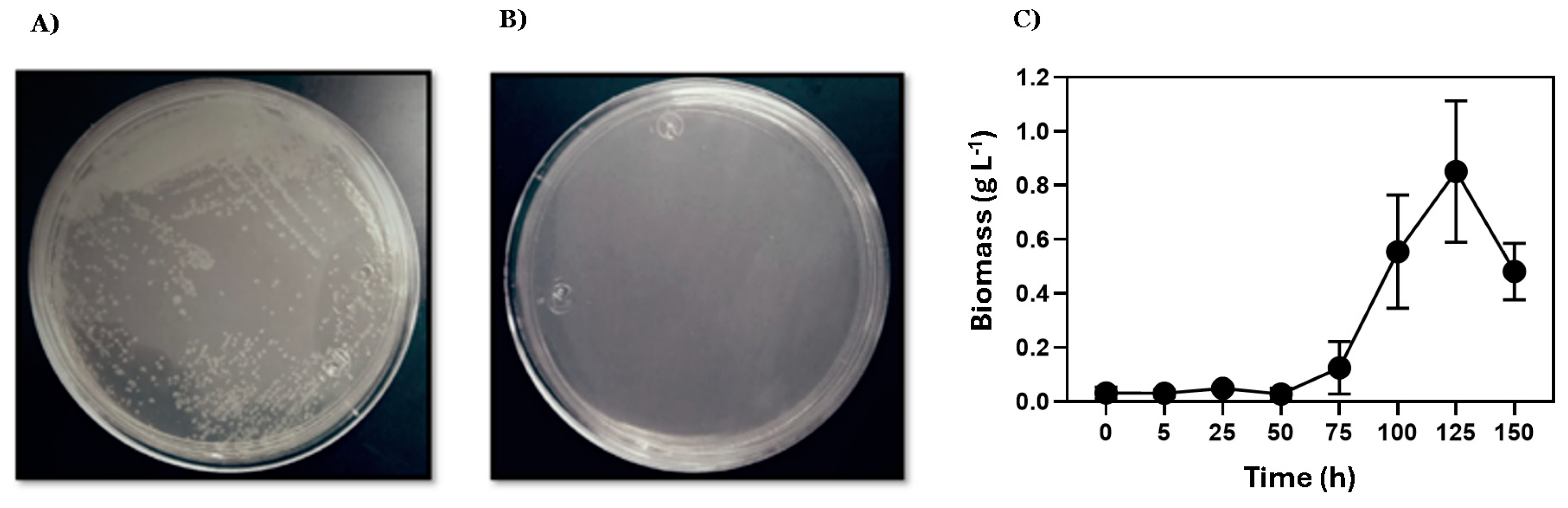
3.1. Qualitative Confirmation of Saccharolytic Activity in E. coli Expressing AIDA-SacA
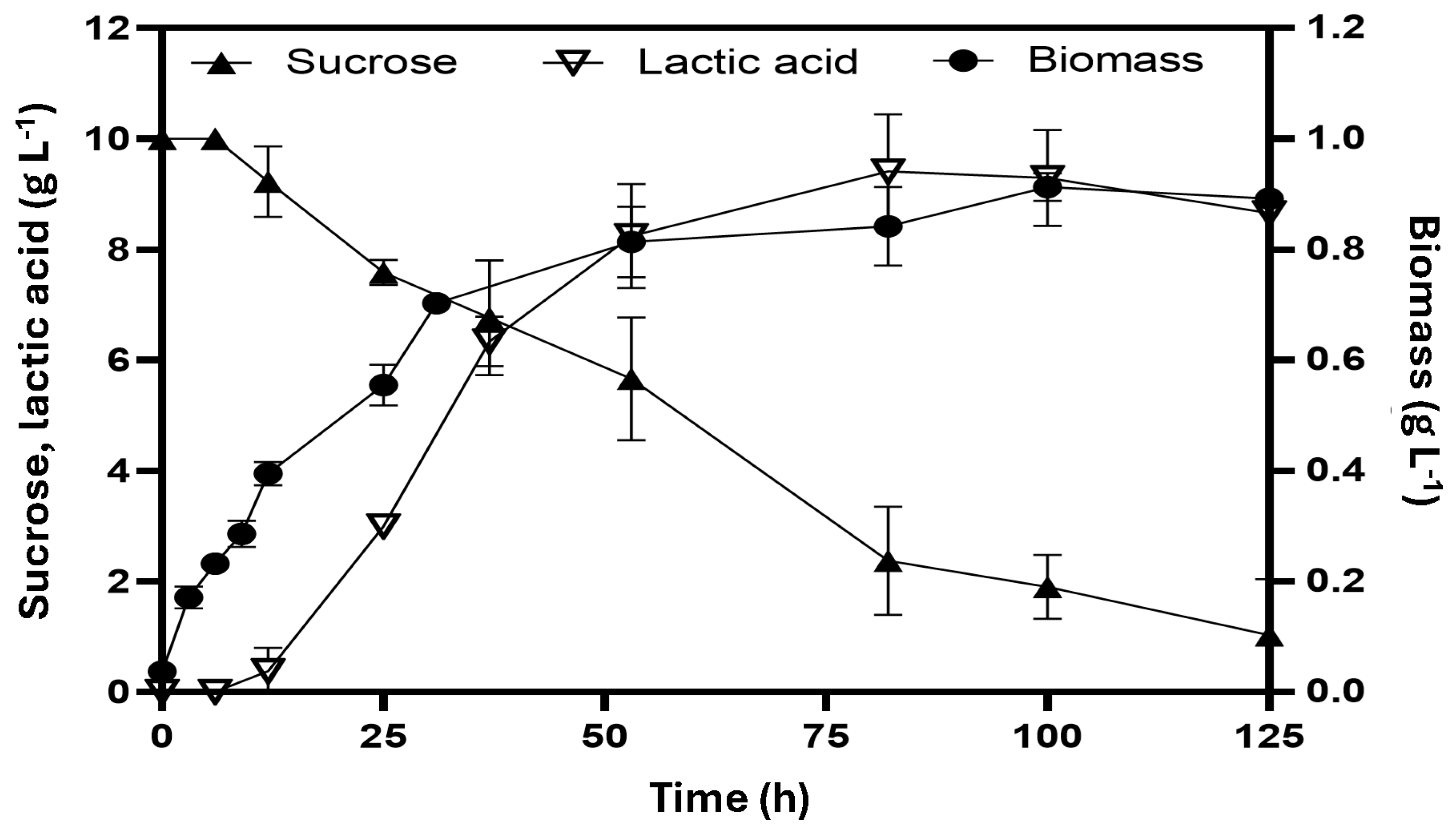
3.2. Lactic Acid Production Prevails in Sucrose Fermentation
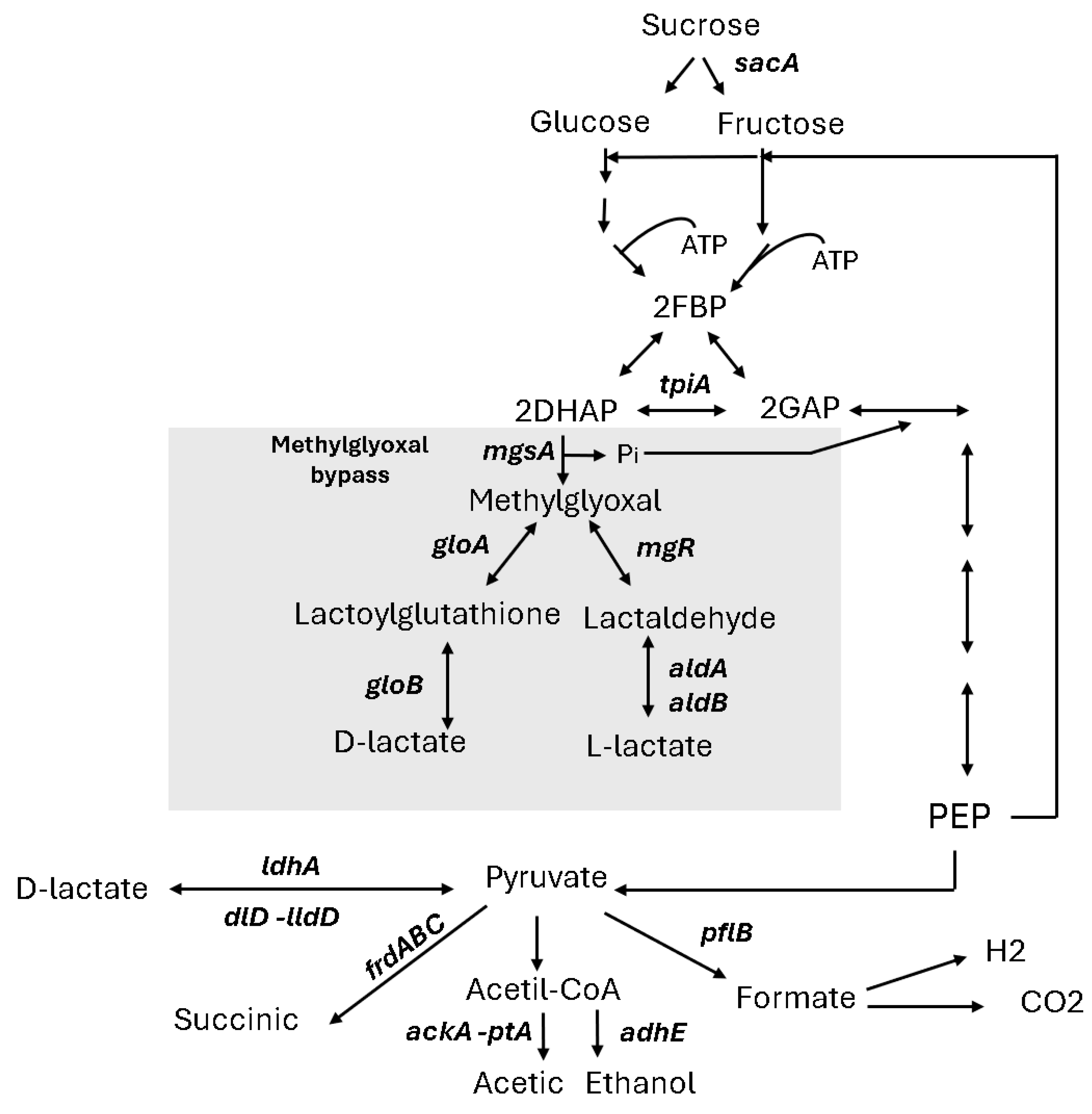
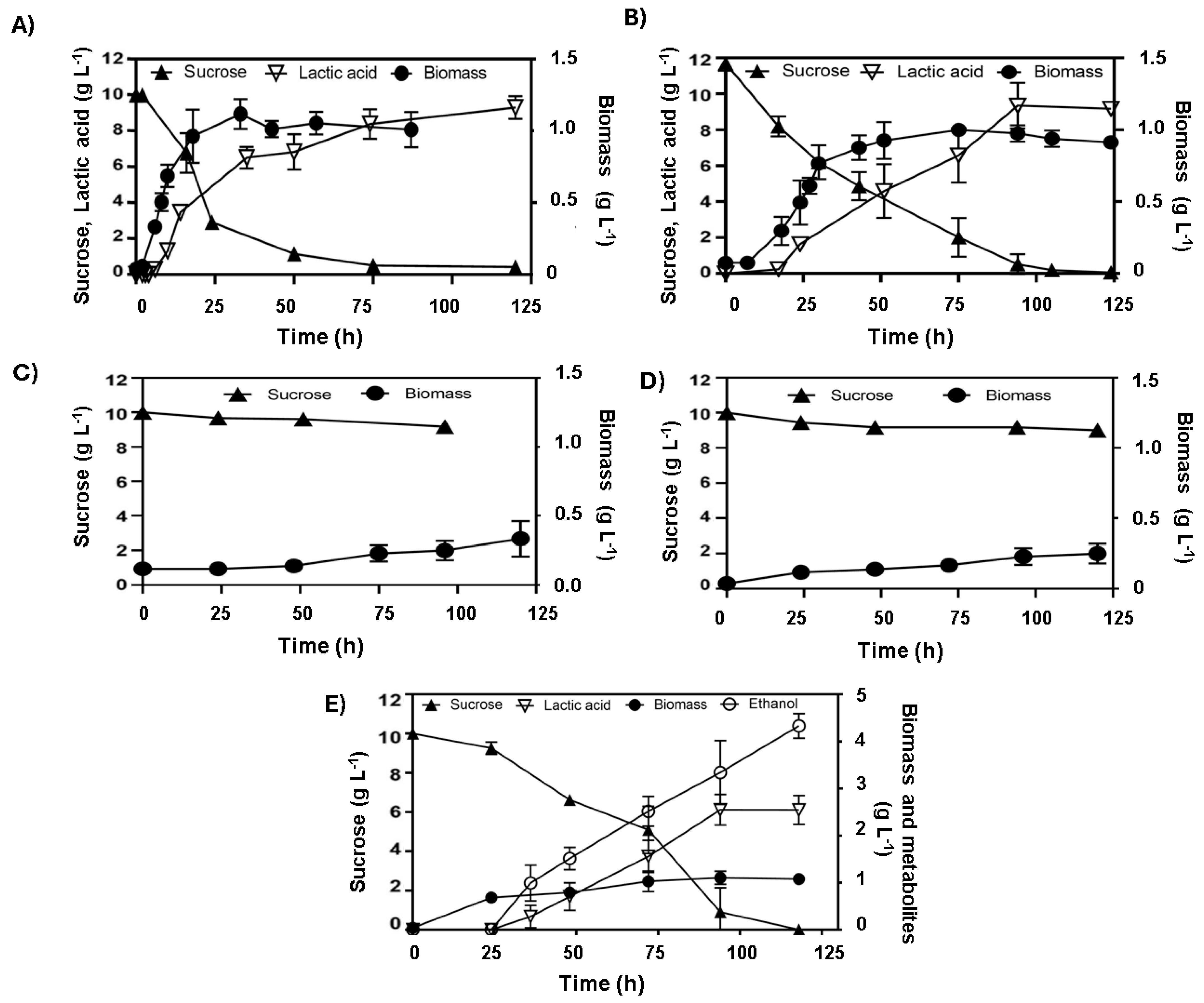
3.3. Homolactic Fermentation on Sucrose in E. coli Strains Lacking ldhA Suggests Alternative Metabolic Pathways
3.4. The mgsA Gene Deletion Underscores the Role of the Methylglyoxal Bypass in Sucrose Metabolism and Lactate Production in E. coli
3.5. Adaptive Evolution of Escherichia coli for Efficient Sucrose Fermentation Through the Methylglyoxal Bypass and the pAIDA-sacA System
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Li, K.; Huang, F.; Wang, J.; Zhao, J.; Zhao, X.; Garza, E.; Manow, R.; Grayburn, S.; Zhou, S. Engineering and adaptive evolution of Escherichia coli W for L-lactic acid fermentation from molasses and corn steep liquor without additional nutrients. Bioresour. Technol. 2013, 148, 394–400. [Google Scholar] [CrossRef]
- Koutinas, A.; Wang, R.; Webb, C. Evaluation of wheat as generic feedstock for chemical production. Ind. Crops Prod. 2004, 20, 75–88. [Google Scholar] [CrossRef]
- Reid, S.J.; Abratt, V.R. Sucrose utilisation in bacteria: Genetic organisation and regulation. Appl. Microbiol. Biotechnol. 2005, 67, 312–321. [Google Scholar] [CrossRef]
- Bruschi, M.; Boyes, S.J.; Sugiarto, H.; Nielsen, L.K.; Vickers, C.E. A transferable sucrose utilization approach for non-sucrose-utilizing Escherichia coli strains. Biotechnol. Adv. 2012, 30, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-García, A.K.; Alvarez-Guzmán, C.L.; De Leon-Rodriguez, A. Autodisplay of alpha amylase from Bacillus megaterium in E. coli for the bioconversion of starch into hydrogen, ethanol and succinic acid. Enzym. Microb. Technol. 2020, 134, 109477. [Google Scholar] [CrossRef]
- Lopez-Hidalgo, A.M.; Hernandez, V.E.B.; De Leon-Rodriguez, A. Scale-up of hydrogen and ethanol co-production by an engineered Escherichia coli. Fuel 2021, 300, 121002. [Google Scholar] [CrossRef]
- Dien, B.S.; Nichols, N.N.; Bothast, R.J. Recombinant Escherichia coli engineered for production of L-lactic acid from hexose and pentose sugars. J. Ind. Microbiol. Biotechnol. 2001, 27, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Choi, S.; Park, J.H.; Vickers, C.E.; Nielsen, L.K.; Lee, S.Y. Development of sucrose-utilizing Escherichia coli K-12 strain by cloning β-fructofuranosidases and its application for L-threonine production. Appl. Microbiol. Biotechnol. 2010, 88, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Fouet, A.; Klier, A.; Rapoport, G. Cloning and expression in Escherichia coli of the sucrase gene from Bacillus subtilis. Mol. Gen. Genet. 1982, 186, 399–404. [Google Scholar] [CrossRef]
- Maurer, J.; Jose, J.; Meyer, T.F. Characterization of the essential transport function of the AIDA-I autotransporter and evidence supporting structural predictions. J. Bacteriol. 1999, 181, 7014–7020. [Google Scholar] [CrossRef] [PubMed]
- Hernández, V.E.B.; Salas-Montantes, C.J.; la Rosa, A.P.B.-D.; De Leon-Rodriguez, A. Autodisplay of an endo-1,4-β-xylanase from Clostridium cellulovorans in Escherichia coli for xylans degradation. Enzym. Microb. Technol. 2021, 149, 109834. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Chen, Y. Constructing Escherichia coli co-display systems for biodegradation of polyethylene terephthalate. Bioresour. Bioprocess. 2023, 10, 91. [Google Scholar] [CrossRef]
- Rosales-Colunga, L.M.; Razo-Flores, E.; Ordoñez, L.G.; Alatriste-Mondragón, F.; De León-Rodríguez, A. Hydrogen production by Escherichia coli ΔhycA ΔlacI using cheese whey as substrate. Int. J. Hydrogen Energy 2010, 35, 491–499. [Google Scholar] [CrossRef]
- Balderas-Hernandez, V.E.; Maldonado, K.P.L.; Sánchez, A.; Smoliński, A.; Rodriguez, A.D.L. Improvement of hydrogen production by metabolic engineering of Escherichia coli: Modification on both the PTS system and central carbon metabolism. Int. J. Hydrogen Energy 2020, 45, 5687–5696. [Google Scholar] [CrossRef]
- Thomason, L.C.; Costantino, N.; Court, D.L. E. coli genome manipulation by P1 transduction. Curr. Protoc. Mol. Biol. 2007, 79, 1.17.1–1.17.8. [Google Scholar] [CrossRef] [PubMed]
- Thouvenot, B.; Charpentier, B.; Branlant, C. The strong efficiency of the Escherichia coli gapA P1 promoter depends on a complex combination of functional determinants. Biochem. J. 2004, 383 Pt 2, 371–382. [Google Scholar] [CrossRef]
- Davila-Vazquez, G.; de León-Rodríguez, A.; Alatriste-Mondragón, F.; Razo-Flores, E. The buffer composition impacts the hydrogen production and the microbial community composition in non-axenic cultures. Biomass Bioenergy 2011, 35, 3174–3181. [Google Scholar] [CrossRef]
- Förster, A.H.; Gescher, J. Metabolic Engineering of Escherichia coli for Production of Mixed-Acid Fermentation End Products. Front. Bioeng. Biotechnol. 2014, 2, 16. [Google Scholar] [CrossRef]
- El-Mansi, M. Control of central metabolism’s architecture in Escherichia coli: An overview. Microbiol. Res. 2023, 266, 127224. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Gu, Y.; Quan, Y.; Gao, W.; Dang, Y.; Cao, M.; Lu, X.; Wang, Y.; Song, C.; Wang, S. Construction of energy-conserving sucrose utilization pathways for improving poly-γ-glutamic acid production in Bacillus amyloliquefaciens. Microb. Cell Fact. 2017, 16, 98. [Google Scholar] [CrossRef]
- Olavarria, K.; Fina, A.; Velasco, M.I.; van Loosdrecht, M.C.M.; Wahl, S.A. Metabolism of sucrose in a non-fermentative Escherichia coli under oxygen limitation. Appl. Microbiol. Biotechnol. 2019, 103, 6245–6256. [Google Scholar] [CrossRef]
- Zhao, C.; Dong, H.; Zhang, Y.; Li, Y. Discovery of potential genes contributing to the biosynthesis of short-chain fatty acids and lactate in gut microbiota from systematic investigation in E. coli. Biofilms Microbiomes 2019, 5, 19. [Google Scholar] [CrossRef]
- Zhou, L.; Cui, W.; Liu, Z.; Zhou, Z. Metabolic engineering strategies for D-lactate over production in Escherichia coli. J. Chem. Technol. Biotechnol. 2016, 91, 576–584. [Google Scholar] [CrossRef]
- Cooper, R.A. Metabolism of methylglyoxal in microorganisms. Annu. Rev. Microbiol. 1984, 38, 49–68. [Google Scholar] [CrossRef]
- McCloskey, D.; Xu, S.; Sandberg, T.E.; Brunk, E.; Hefner, Y.; Szubin, R.; Feist, A.M.; Palsson, B.O. Adaptation to the coupling of glycolysis to toxic methylglyoxal production in tpiA deletion strains of Escherichia coli requires synchronized and counterintuitive genetic changes. Metab. Eng. 2018, 48, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Yomano, L.P.; York, S.W.; Shanmugam, K.T.; Ingram, L.O. Deletion of methylglyoxal synthase gene (mgsA) increased sugar co-metabolism in ethanol-producing Escherichia coli. Biotechnol. Lett. 2009, 31, 1389–1398. [Google Scholar] [CrossRef]
- Tötemeyer, S.; Booth, N.A.; Nichols, W.W.; Dunbar, B.; Booth, I.R. From famine to feast: The role of methylglyoxal production in Escherichia coli. Mol. Microbiol. 1998, 27, 553–562. [Google Scholar] [CrossRef]
- Ferguson, G.P.; Tötemeyer, S.; MacLean, M.J.; Booth, I.R. Methylglyoxal production in bacteria: Suicide or survival? Arch. Microbiol. 1998, 170, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Valdovinos, M.A.; Martínez-Antonio, A. Optimizing. Fermentation Strategies for Enhanced Tryptophan Production in Escherichia coli: Integrating Genetic and Environmental Controls for Industrial Applications. Processes 2024, 12, 2422. [Google Scholar] [CrossRef]
- Booth, I.R. Glycerol and Methylglyoxal Metabolism. EcoSal Plus 2005, 1, 10.1128/ecosalplus.3.4.3. [Google Scholar] [CrossRef] [PubMed]
- Iacometti, C.; Marx, K.; Hönick, M.; Biletskaia, V.; Schulz-Mirbach, H.; Dronsella, B.; Satanowski, A.; Delmas, V.A.; Berger, A.; Dubois, I.; et al. Activating Silent Glycolysis Bypasses in Escherichia coli. BioDes. Res. 2022, 2022, 9859643. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tian, T.; Zhao, J.; Wang, J.; Yan, T.; Xu, L.; Liu, Z.; Garza, E.; Iverson, A.; Manow, R.; et al. Homofermentative production of D-lactic acid from sucrose by a metabolically engineered Escherichia coli. Biotechnol. Lett. 2012, 34, 2069–2075. [Google Scholar] [CrossRef] [PubMed]
- da Silva, G.P.; de Araújo, E.F.; Silva, D.O.; Guimarães, W.V. Ethanolic fermentation of sucrose, sugarcane juice and molasses by Escherichia coli strain kO11 and klebsiella oxytoca strain p2. Braz. J. Microbiol. 2005, 36, 395–404. [Google Scholar] [CrossRef]
- Weber, J.; Kayser, A.; Rinas, U. Metabolic flux analysis of Escherichia coli in glucose-limited continuous culture. II. Dynamic response to famine and feast, activation of the methylglyoxal pathway and oscillatory behaviour. Microbiology 2005, 151 Pt 3, 707–716. [Google Scholar] [CrossRef]
- Ferguson, G.P.; Chacko, A.D.; Lee, C.H.; Booth, I.R. The Activity of the High-Affinity K Uptake System Kdp Sensitizes Cells of Escherichia coli to Methylglyoxal. J. Bacteriol. 1996, 178, 3957–3961. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, H.; Cocaign-Bousquet, M.; Lindley, N.D. Simultaneous consumption of glucose and fructose from sugar mixtures during batch growth of Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 1997, 47, 600–603. [Google Scholar] [CrossRef]
- Zwaig, N.; Kistler, W.S.; Lin, E.C.C. Glycerol kinase, the pacemaker for the dissimilation of glycerol in Escherichia coli. J. Bacteriol. 1970, 102, 753–759. [Google Scholar] [CrossRef]
- Ackerman, R.S.; Cozzarelli, N.R.; Epstein, W. Accumulation of toxic concentrations of methylglyoxal by wild-type Escherichia coli K-12. J. Bacteriol. 1974, 119, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jiang, X.; Zheng, Y.; Li, K.; Zhang, R.; Xu, J.; Wang, Z.; Zhang, Y.; Yin, H.; Li, J. Expression, Characterization, and Immobilization of a Novel D-Lactate Dehydrogenase from Salinispirillum sp. LH 10-3-1. Processes 2024, 12, 1349. [Google Scholar] [CrossRef]
- Ali, S.; Isha; Chang, Y.-C. Ecotoxicological Impact of Bioplastics Biodegradation: A Comprehensive Review. Processes 2023, 11, 3445. [Google Scholar] [CrossRef]
- Hirokawa, Y.; Goto, R.; Umetani, Y.; Hanai, T. Construction of a novel d-lactate producing pathway from dihydroxyacetone phosphate of the Calvin cycle in cyanobacterium, Synechococcus elongatus PCC 7942. J. Biosci. Bioeng. 2017, 124, 54–61. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Genotype Description | Reference |
|---|---|---|
| E. coli DH5α | Cloning host | [13] |
| E. coli W3110 | Wild type, production host | [13] |
| WDH | ΔhycA | [13] |
| WDHA | ΔhycA ΔldhA | [14] |
| WDHFA | ΔhycA ΔldhA ΔfrdDABC | [14] |
| WDHFAK | ΔhycA ΔldhA ΔfrdDABC ΔackA | [14] |
| WDHFAP | ΔhycA ΔldhA ΔfrdDABC Δpta | [14] |
| WDHFAKM | ΔhycA ΔldhA ΔfrdDABC ΔackA ΔmgsA | This work |
| WDHFAPM | ΔhycA ΔldhA ΔfrdDABC Δpta ΔmgsA | This work |
| WDHFAPMEV WDHFAKEV | WDHFAPM (adaptively evolved mutant) WDHFAK (adaptively evolved mutant) | This work This work |
| Plasmid | ||
| pAIDA-sacA | Fusion aida-sacA genes | This work |
| pAIDA | aida gene | This work |
| Primers | Sequence (5′to 3′) | |
| ldhA-FCK | TCGCCATCGGTCTACGGGC | [14] |
| ldhA-RCK | CATAACACCATTAGCGAAAT | [14] |
| frdD-FCK | TCTGGTTTCCATACAA | [14] |
| frdD-RCK | TTAGATTGTAACGACACCAATC | [14] |
| mgsA-FCK | ATCGACTTTATGACCATTCATGCC | This work |
| mgsA-RCK | GTCGAGGTTGAACGCTATAACAAATC | This work |
| sacA FW | GGCGCGCCTACAGCACATGACCAGGAG | This work |
| sacA Rv | CTCGAGCGCATAAGTGTCCAAATTCC | This work |
| Carbon Source 10 (g L−1) | Biomass (g L−1) | Lactic Acid (g L−1) | Lactic Acid Yield (g g−1) | Succinic Acid (g L−1) | Succinic Acid Yield (g g−1) | Acetic Acid (g L−1) | Acetic Acid Yield (g g−1) | Ethanol (g L−1) | Ethanol Yield (g g−1) |
|---|---|---|---|---|---|---|---|---|---|
| Sucrose | 0.91 ± 0.02 | 9.25 ± 0.88 | 0.84 ± 0.09 | - | - | - | - | - | - |
| Glucose | 0.62 ± 0.05 | 1.88 ± 0.03 | 0.17 ± 0.03 | 2.36 ± 0.26 | 0.21 ± 0.30 | 1.01 ± 0.33 | 0.09 ± 0.02 | 1.90 ± 0.38 | 0.17 ± 0.03 |
| Fructose | 0.61 ± 0.09 | 1.39 ± 0.08 | 0.16 ± 0.08 | 1.78 ± 0.06 | 0.16 ± 0.01 | 0.93 ± 0.09 | 0.08 ± 0.01 | 1.50 ± 0.21 | 0.13 ± 0.09 |
| E. coli Strain | µ (h−1) | Maximum Cell (g L−1) | Lactic Acid (g L−1) | Yield Lactic Acid (g g−1) | Ethanol (g L−1) | Ethanol Yield (g g−1) |
|---|---|---|---|---|---|---|
| WDHFAP/pAIDA-sacA | 0.35 ± 0.03 | 1.33 ± 0.10 | 9.85 ± 0.28 | 0.89 ± 0.02 | 0.25 ± 0.05 | 0.02 ± 0.05 |
| WDHFAPM/pAIDA-sacA | 0.02 ± 0.01 | 0.85 ± 0.12 | 1.98 ± 0.18 | 0.18± 0.16 | 4.29 ± 0.79 | 0.39 ± 0.07 |
| WDHFAPMEV | 0.03 ± 0.04 | 0.88 ± 0.07 | 1.95 ± 0.17 | 0.17 ± 0.23 | 4.04 ± 0.49 | 0.36 ± 0.04 |
| WDHFAK/pAIDA-sacA | 0.13 ± 0.01 | 0.96 ± 0.03 | 2.83 ± 0.71 | 0.25 ± 0.23 | 2.16 ± 0.09 | 0.19 ± 0.08 |
| WDHFAKM/pAIDA-sacA | 0.01 ± 0.06 | 1.01 ± 0.11 | 0.97 ± 0.72 | 0.08 ± 0.86 | 4.00 ± 0.33 | 0.36 ± 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Andrade, J.; Balderas-Hernández, V.E.; Barba de la Rosa, A.P.; De Leon-Rodriguez, A. Immobilisation of Sucrase A from Bacillus subtilis on the Surface of Escherichia coli Mediated by the AIDA-I Autotransporter: Application on the Homolactic Fermentation. Processes 2025, 13, 470. https://doi.org/10.3390/pr13020470
Sánchez-Andrade J, Balderas-Hernández VE, Barba de la Rosa AP, De Leon-Rodriguez A. Immobilisation of Sucrase A from Bacillus subtilis on the Surface of Escherichia coli Mediated by the AIDA-I Autotransporter: Application on the Homolactic Fermentation. Processes. 2025; 13(2):470. https://doi.org/10.3390/pr13020470
Chicago/Turabian StyleSánchez-Andrade, Jorge, Victor E. Balderas-Hernández, Ana P. Barba de la Rosa, and Antonio De Leon-Rodriguez. 2025. "Immobilisation of Sucrase A from Bacillus subtilis on the Surface of Escherichia coli Mediated by the AIDA-I Autotransporter: Application on the Homolactic Fermentation" Processes 13, no. 2: 470. https://doi.org/10.3390/pr13020470
APA StyleSánchez-Andrade, J., Balderas-Hernández, V. E., Barba de la Rosa, A. P., & De Leon-Rodriguez, A. (2025). Immobilisation of Sucrase A from Bacillus subtilis on the Surface of Escherichia coli Mediated by the AIDA-I Autotransporter: Application on the Homolactic Fermentation. Processes, 13(2), 470. https://doi.org/10.3390/pr13020470