


Hypomagnetic Fields and Their Multilevel Effects on Living Organisms


Abstract
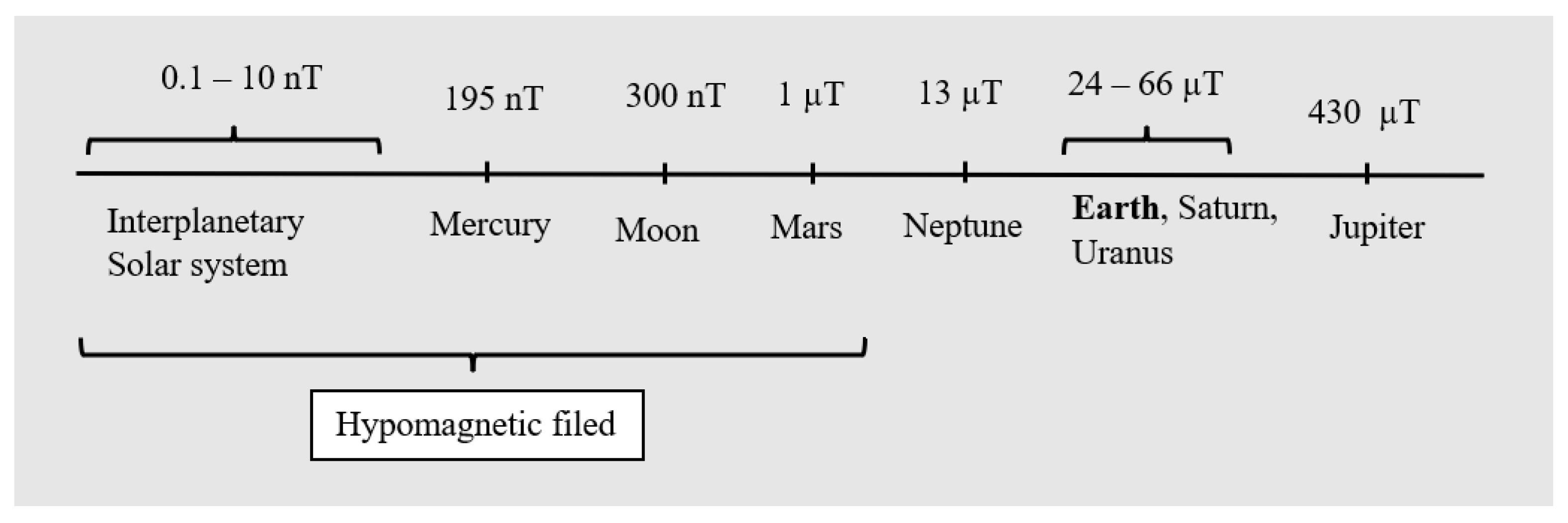
1. Introduction
2. Materials and Methods
3. Mechanisms of Magnetoreception of HMFs
- 1.
- Radical pair mechanism
- 2.
- Universal physical mechanism
- 3.
- Molecular gyroscope mechanism
4. Influences of HMFs on Organisms
4.1. Animals and Animal Cell Cultures
4.1.1. Cell Transport and Respiration
4.1.2. Animals
4.2. Plants
4.3. Prokaryotes
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Monteil, C.L.; Lefevre, C.T. Magnetoreception in Microorganisms. Trends Microbiol. 2019, 28, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Binhi, V.N.; Prato, F.S. Biological effects of the hypomagnetic field: An analytical review of experiments and theories. PLoS ONE 2017, 12, e0179340. [Google Scholar] [CrossRef] [PubMed]
- Mo, W.; Liu, Y.; He, R. Hypomagnetic field, an ignorable environmental factor in space? Sci. China Life Sci. 2014, 57, 726–728. [Google Scholar] [CrossRef] [PubMed]
- Kivelson, M.G.; Bagenal, F. Planetary magnetospheres. In Encyclopedia of the Solar System, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2014; Chapter 7; pp. 137–157. [Google Scholar]
- Zhang, Z.; Xue, Y.; Yang, J.; Shang, P.; Yuan, X. Biological Effects of Hypomagnetic Field: Ground-Based Data for Space Exploration. Bioelectromagnetics 2021, 42, 516–531. [Google Scholar] [CrossRef]
- Pavlík, M. Compare of shielding effectiveness for building materials. Prz. Elektrotechniczny 2019, 95, 137–140. [Google Scholar] [CrossRef]
- Guo, C.; Liu, D. Quantitative Analyses of Magnetic Field Distributions for Buildings of Steel Structure. In Proceedings of the 2012 Sixth International Conference on Electromagnetic Field Problems and Applications, Dalian, China, 19–21 June 2012. [Google Scholar]
- Erdmann, W.; Kmita, H.; Kosicki, J.Z.; Kaczmarek, Ł. How the Geomagnetic Field Influences Life on Earth—An Integrated Approach to Geomagnetobiology. Space Life Sci. 2021, 51, 231–257. [Google Scholar] [CrossRef]
- Wajnberg, E.; Acosta-Avalos, D.; Alves, O.C.; de Oliveira, J.F.; Srygley, R.B.; Esquivel, D.M.S. Magnetoreception in eusocial insects: An update. J. R. Soc. Interface 2010, 7, S207–S225. [Google Scholar] [CrossRef] [PubMed]
- Binhi, V.N.; Savin, A.V. Molecular gyroscopes and biological effects of weak extremely low-frequency magnetic fields. Phys. Rev. E 2002, 65, 051912. [Google Scholar] [CrossRef]
- Dröge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Gauron, C.; Rampon, C.; Bouzaffour, M.; Ipendey, E.; Teillon, J.; Volovitch, M.; Vriz, S. Sustained production of ROS triggers compensatory proliferation and is required for regeneration to proceed. Sci. Rep. 2013, 3, srep02084. [Google Scholar] [CrossRef]
- Van Huizen, A.V.; Morton, J.M.; Kinsey, L.J.; Von Kannon, D.G.; Saad, M.A.; Birkholz, T.R.; Czajka, J.M.; Cyrus, J.; Barnes, F.S.; Beane, W.S. Weak magnetic fields alter stem cell–mediated growth. Sci. Adv. 2019, 5, eaau7201. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.; Sinayskiy, I.; Petruccione, F. An open quantum system approach to the radical pair mechanism. Sci. Rep. 2018, 8, 15719. [Google Scholar] [CrossRef] [PubMed]
- Hore, P.J.; Mouritsen, H. The Radical-Pair Mechanism of Magnetoreception. Annu. Rev. Biophys. 2016, 45, 299–344. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Gómez, M.J.; Sendra-Portero, F.; Martínez-Morillo, M. Effect of 2.45 mT sinusoidal 50 Hz magnetic field on Saccharomyces cerevisiae strains deficient in DNA strand breaks repair. Int. J. Radiat. Biol. 2010, 86, 602–611. [Google Scholar] [CrossRef]
- Barnes, F.; Greenenbaum, B. Some Effects of Weak Magnetic Fields on Biological Systems: RF fields can change radical concentrations and cancer cell growth rates. IEEE Power Electron. Mag. 2016, 3, 60–68. [Google Scholar] [CrossRef]
- Brocklehurst, B.; Mclauchlan, K.A. Free radical mechanism for the effects of environmental electromagnetic fields on biological systems. Int. J. Radiat. Biol. 1996, 69, 3–24. [Google Scholar] [CrossRef]
- Binhi, V.N.; Prato, F.S. A physical mechanism of magnetoreception: Extension and analysis. Bioelectromagnetics 2016, 38, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, H.; Mitsui, H.; Miura, K.; Okano, K.; Imamoto, Y.; Okano, T. Rapid Oxidation Following Photoreduction in the Avian Cryptochrome4 Photocycle. Biochemistry 2020, 59, 3615–3625. [Google Scholar] [CrossRef] [PubMed]
- Novikov, V.V.; Yablokova, E.V.; Fesenko, E.E. The Effect of a “Zero” Magnetic Field on the Production of Reactive Oxygen Species in Neutrophils. Biophysics 2018, 63, 365–368. [Google Scholar] [CrossRef]
- Yan, M.-M.; Zhang, L.; Cheng, Y.-X.; Sappington, T.W.; Pan, W.-D.; Jiang, X.-F. Effect of a near-zero magnetic field on development and flight of oriental armyworm (Mythimna separata). J. Integr. Agric. 2021, 20, 1336–1345. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, L.; Zhan, A.; Wang, M.; Tian, L.; Guo, W.; Pan, Y. Long-term exposure to a hypomagnetic field attenuates adult hippocampal neurogenesis and cognition. Nat. Commun. 2021, 12, 1174. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Stait-Gardner, T.; Price, W.S. Is It Time to Forgo the Use of the Terms “Spin–Lattice” and “Spin–Spin” Relaxation in NMR and MRI? J. Phys. Chem. Lett. 2021, 12, 6305–6312. [Google Scholar] [CrossRef] [PubMed]
- Zangi, R.; Hagen, M.; Berne, B.J. Effect of Ions on the Hydrophobic Interaction between Two Plates. J. Am. Chem. Soc. 2007, 129, 4678–4686. [Google Scholar] [CrossRef] [PubMed]
- Zhao, V.; Jacobs, W.M.; Shakhnovich, E.I. Effect of Protein Structure on Evolution of Cotranslational Folding. Biophys. J. 2020, 119, 1123–1134. [Google Scholar] [CrossRef]
- Ogneva, I.V.; Usik, M.A.; Burtseva, M.V.; Biryukov, N.S.; Zhdankina, Y.S.; Sychev, V.N.; Orlov, O.I. Drosophila melanogaster Sperm under Simulated Microgravity and a Hypomagnetic Field: Motility and Cell Respiration. Int. J. Mol. Sci. 2020, 21, 5985. [Google Scholar] [CrossRef]
- Zhang, B.; Tian, L. Reactive Oxygen Species: Potential Regulatory Molecules in Response to Hypomagnetic Field Exposure. Bioelectromagnetics 2020, 41, 573–580. [Google Scholar] [CrossRef]
- Erdmann, W.; Idzikowski, B.; Kowalski, W.; Kosicki, J.Z.; Kaczmarek, Ł. Tolerance of two anhydrobiotic tardigrades Echiniscus testudo and Milnesium inceptum to hypomagnetic conditions. PeerJ 2021, 9, e10630. [Google Scholar] [CrossRef]
- Jia, W.; Fan, Z.; Du, A.; Shi, L. Molecular mechanism of Mare Nectaris and magnetic field on the formation of ethyl carbamate during 19 years aging of Feng-flavor Baijiu. Food Chem. 2022, 382, 132357. [Google Scholar] [CrossRef]
- Tombarkiewicz, B. Effect of long-term geomagnetic field deprivation on the concentration of some elements in the hair of laboratory rats. Environ. Toxicol. Pharmacol. 2008, 26, 75–79. [Google Scholar] [CrossRef]
- Kantserova, N.P.; Krylov, V.V.; Lysenko, L.A.; Ushakova, N.V.; Nemova, N.N. Effects of Hypomagnetic Conditions and Reversed Geomagnetic Field on Calcium-Dependent Proteases of Invertebrates and Fish. Izv. Atmos. Ocean. Phys. 2017, 53, 719–723. [Google Scholar] [CrossRef]
- Fu, J.-P.; Mo, W.-C.; Liu, Y.; Bartlett, P.F.; He, R.-Q. Elimination of the geomagnetic field stimulates the proliferation of mouse neural progenitor and stem cells. Protein Cell 2016, 7, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.D.; Mo, W.C.; Fu, J.P.; Liu, Y.; He, R.Q. Long-term Hypogeomagnetic field exposure reduces muscular mitochondrial function and exercise capacity in adult male mice. Prog. Biochem. Biophys. 2020, 47, 426–438. [Google Scholar]
- Lednev, V.V. Bioeffects of weak static and alternating magnetic fields. Biofizika 1996, 41, 224–232. [Google Scholar] [PubMed]
- Krebs, J. Structure, Function and Regulation of the Plasma Membrane Calcium Pump in Health and Disease. Int. J. Mol. Sci. 2022, 23, 1027. [Google Scholar] [CrossRef] [PubMed]
- Poniedziałek, B.; Rzymski, P.; Karczewski, J.; Jaroszyk, F.; Wiktorowicz, K. Reactive oxygen species (ROS) production in human peripheral blood neutrophils exposed in vitro to static magnetic field. Electromagn. Biol. Med. 2013, 32, 560–568. [Google Scholar]
- Vergallo, C.; Ahmadi, M.; Mobasheri, H.; Dini, L. Impact of Inhomogeneous Static Magnetic Field (31.7–232.0 mT) Exposure on Human Neuroblastoma SH-SY5Y Cells during Cisplatin Administration. PLoS ONE 2014, 9, e113530. [Google Scholar] [CrossRef]
- Tang, R.; Xu, Y.; Ma, F.; Ren, J.; Shen, S.; Du, Y.; Hou, Y.; Wang, T. Extremely low frequency magnetic fields regulate differentiation of regulatory T cells: Potential role for ROS-mediated inhibition on AKT. Bioelectromagnetics 2016, 37, 89–98. [Google Scholar] [CrossRef]
- Angelova, P.R.; Dinkova-Kostova, A.T.; Abramov, A.Y. Assessment of ROS Production in the Mitochondria of Live Cells. In Reactive Oxygen Species; Humana: New York, NY, USA, 2021; pp. 33–42. [Google Scholar] [CrossRef]
- Belyavskaya, N. Biological effects due to weak magnetic field on plants. Adv. Space Res. 2004, 34, 1566–1574. [Google Scholar] [CrossRef]
- Fu, J.-P.; Mo, W.-C.; Liu, Y.; He, R.-Q. Decline of cell viability and mitochondrial activity in mouse skeletal muscle cell in a hypomagnetic field. Bioelectromagnetics 2016, 37, 212–222. [Google Scholar] [CrossRef]
- Montoya, R.D. Magnetic fields, radicals and cellular activity. Electromagn. Biol. Med. 2017, 36, 102–113. [Google Scholar] [CrossRef]
- Zhang, H.-T.; Zhang, Z.-J.; Mo, W.-C.; Hu, P.-D.; Ding, H.-M.; Liu, Y.; Hua, Q.; He, R.-Q. Shielding of the geomagnetic field reduces hydrogen peroxide production in human neuroblastoma cell and inhibits the activity of CuZn superoxide dismutase. Protein Cell 2017, 8, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Mo, W.; Liu, Y.; Bartlett, P.F.; He, R. Transcriptome profile of human neuroblastoma cells in the hypomagnetic field. Sci. China Life Sci. 2014, 57, 448–461. [Google Scholar] [CrossRef] [PubMed]
- Mo, W.C.; Zhang, Z.J.; Wang, D.L.; Liu, Y.; Bartlett, P.F.; He, R.Q. Shielding of the geomagnetic field alters actin assembly and inhibits cell motility in human neuroblastoma cells. Sci. Rep. 2016, 6, 22624. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Lu, H.; Xi, W.; Zhou, X.; Xu, S.; Zhang, K.; Jiang, J.; Li, Y.; Guo, A. Exposure to hypomagnetic field space for multiple generations causes amnesia in Drosophila melanogaster. Neurosci. Lett. 2004, 371, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Sarimov, R.M.; Binhi, V.N.; Milyaev, V.A. The influence of geomagnetic field compensation on human cognitive processes. Biophysics 2008, 53, 433–441. [Google Scholar] [CrossRef]
- Wang, G.-M.; Fu, J.-P.; Mo, W.-C.; Zhang, H.-T.; Liu, Y.; He, R.-Q. Shielded geomagnetic field accelerates glucose consumption in human neuroblastoma cells by promoting anaerobic glycolysis. Biochem. Biophys. Res. Commun. 2022, 601, 101–108. [Google Scholar] [CrossRef]
- Mo, W.-C.; Zhang, Z.-J.; Liu, Y.; Bartlett, P.F.; He, R.-Q. Magnetic Shielding Accelerates the Proliferation of Human Neuroblastoma Cell by Promoting G1-Phase Progression. PLoS ONE 2013, 8, e54775. [Google Scholar] [CrossRef]
- Gurfinkel, Y.; At’Kov, O.; Vasin, A.; Breus, T.; Sasonko, M.; Pishchalnikov, R. Effect of zero magnetic field on cardiovascular system and microcirculation. Life Sci. Space Res. 2016, 8, 1–7. [Google Scholar] [CrossRef]
- IuI, G.; Vasin, A.L.; Matveeva, T.A.; Sasonko, M.L. Evaluation of the hypomagnetic environment effects on capillary blood circulation, blood pressure and heart rate. Aviakosmicheskaia I Ekol. Meditsina= Aerosp. Environ. Med. 2014, 48, 24–30. [Google Scholar]
- Ciorba, D.; Morariu, V.V. Life in zero magnetic field. III. Activity of aspartate aminotransferase and alanine aminotransferase during in vitro aging of human blood. Electro- Magn. 2001, 20, 313–321. [Google Scholar] [CrossRef]
- Katiukhin, L.N. Rheological Properties of the Erythrocytes in Weakened Static Magnetic Field of the Earth In vitro Study. J. Sci. Res. Rep. 2019, 22, 1–12. [Google Scholar] [CrossRef]
- Krylov, V.V.; Bolotovskaya, I.V.; Osipova, E.A. The response of European Daphnia magna Straus and Australian Daphnia carinata King to changes in geomagnetic field. Electromagn. Biol. Med. 2013, 32, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Wan, G.-J.; Yuan, R.; Wang, W.-J.; Fu, K.-Y.; Zhao, J.-Y.; Jiang, S.-L.; Pan, W.-D.; Sword, G.A.; Chen, F.-J. Reduced geomagnetic field may affect positive phototaxis and flight capacity of a migratory rice planthopper. Anim. Behav. 2016, 121, 107–116. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, W. Removal or component reversal of local geomagnetic field affects foraging orientation preference in migratory insect brown planthopper Nilaparvata lugens. Peerj 2021, 9, e12351. [Google Scholar] [CrossRef] [PubMed]
- Wan, G.-J.; Jiang, S.-L.; Zhao, Z.-C.; Xu, J.-J.; Tao, X.-R.; Sword, G.A.; Gao, Y.-B.; Pan, W.-D.; Chen, F.-J. Bio-effects of near-zero magnetic fields on the growth, development and reproduction of small brown planthopper, Laodelphax striatellus and brown planthopper, Nilaparvata lugens. J. Insect Physiol. 2014, 68, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Mo, W.; Liu, Y.; Cooper, H.M.; He, R.-Q. Altered development of Xenopus embryos in a hypogeomagnetic field. Bioelectromagnetics 2011, 33, 238–246. [Google Scholar] [CrossRef]
- Fesenko, E.E.; Mezhevikina, L.M.; Osipenko, M.A.; Gordon, R.Y.; Khutzian, S.S. Effect of the “zero” magnetic field on early embryogenesis in mice. Electromagn. Biol. Med. 2010, 29, 1–8. [Google Scholar] [CrossRef]
- Xue, X.; Ali, Y.F.; Liu, C.; Hong, Z.; Luo, W.; Nie, J.; Li, B.; Jiao, Y.; Liu, N.-A. Geomagnetic Shielding Enhances Radiation Resistance by Promoting DNA Repair Process in Human Bronchial Epithelial Cells. Int. J. Mol. Sci. 2020, 21, 9304. [Google Scholar] [CrossRef]
- Belyaev, I.Y.; Alipov, Y.D.; Harms-Ringdahl, M. Effects of zero magnetic field on the conformation of chromatin in human cells. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1997, 1336, 465–473. [Google Scholar] [CrossRef]
- Baek, S.; Choi, H.; Park, H.; Cho, B.; Kim, S.; Kim, J. Effects of a hypomagnetic field on DNA methylation during the differentiation of embryonic stem cells. Sci. Rep. 2019, 9, 1333. [Google Scholar] [CrossRef]
- Agliassa, C.; Narayana, R.; Christie, J.M.; Maffei, M.E. Geomagnetic field impacts on cryptochrome and phytochrome signaling. J. Photochem. Photobiol. B Biol. 2018, 185, 32–40. [Google Scholar] [CrossRef]
- Maffei, M.E. Magnetic field effects on plant growth, development, and evolution. Front. Plant Sci. 2014, 5, 445. [Google Scholar] [CrossRef]
- Agliassa, C.; Mannino, G.; Molino, D.; Cavalletto, S.; Contartese, V.; Bertea, C.M.; Secchi, F. A new protein hydrolysate-based biostimulant applied by fertigation promotes relief from drought stress in Capsicum annuum L. Plant Physiol. Biochem. 2021, 166, 1076–1086. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhang, Y.; Yu, Y.; Li, Y.; Wei, S. Suppression of Arabidopsis flowering by near-null magnetic field is mediated by auxin. Bioelectromagnetics 2017, 39, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Yu, Y.; Zhang, Y.; Li, Y.; Wei, S. Gibberellins are involved in effect of near-null magnetic field on Arabidopsis flowering. Bioelectromagnetics 2016, 38, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mo, W.-C.; Zhang, Z.-J.; Liu, Y.; Zhai, G.-J.; Jiang, Y.-D.; He, R.-Q. Effects of a hypogeomagnetic field on gravitropism and germination in soybean. Adv. Space Res. 2011, 47, 1616–1621. [Google Scholar] [CrossRef]
- Xu, C.; Wei, S.; Lu, Y.; Zhang, Y.; Chen, C.; Song, T. Removal of the local geomagnetic field affects reproductive growth inArabidopsis. Bioelectromagnetics 2013, 34, 437–442. [Google Scholar] [CrossRef]
- Islam, M.; Maffei, M.; Vigani, G. The Geomagnetic Field Is a Contributing Factor for an Efficient Iron Uptake in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 325. [Google Scholar] [CrossRef]
- Negishi, Y.; Hashimoto, A.; Tsushima, M.; Dobrota, C.; Yamashita, M.; Nakamura, T. Growth of pea epicotyl in low magnetic field implication for space research. Adv. Space Res. 1999, 23, 2029–2032. [Google Scholar] [CrossRef]
- Xu, C.; Yin, X.; Lv, Y.; Wu, C.; Zhang, Y.; Song, T. A near-null magnetic field affects cryptochrome-related hypocotyl growth and flowering in Arabidopsis. Adv. Space Res. 2012, 49, 834–840. [Google Scholar] [CrossRef]
- Xu, C.; Li, Y.; Yu, Y.; Zhang, Y.; Wei, S. Suppression of Arabidopsis flowering by near-null magnetic field is affected by light. Bioelectromagnetics 2015, 36, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Narayana, R.; Fliegmann, J.; Paponov, I.; Maffei, M.E. Reduction of geomagnetic field (GMF) to near null magnetic field (NNMF) affects Arabidopsis thaliana root mineral nutrition. Life Sci. Space Res. 2018, 19, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Vanderstraeten, J.; Gailly, P.; Malkemper, E.P. Low-light dependence of the magnetic field effect on cryptochromes: Possible relevance to plant ecology. Front. Plant Sci. 2018, 9, 121. [Google Scholar] [CrossRef]
- Soltani, F.; Kashi, A.; Arghavani, M. Effect of magnetic field on Asparagus officinalis L. seed germination and seedling growth. Seed Sci. Technol. 2006, 34, 349–353. [Google Scholar] [CrossRef]
- Tsetlin, V.; Moisa, S.; Levinskikh, M.; Nefedova, E. EFFECT OF VERY SMALL DOSES OF IONIZING RADIATION AND HYPOMAGNETIC FIELD CHANGE PHYSIOLOGICAL CHARACTERISTICS OF HIGHER PLANT SEEDS. Aviakosmicheskaia I Ekol. Meditsina= Aerosp. Environ. Med. 2016, 50, 51–58. [Google Scholar] [CrossRef]
- Zhang, S.-D.; Petersen, N.; Zhang, W.-J.; Cargou, S.; Ruan, J.; Murat, D.; Santini, C.-L.; Song, T.; Kato, T.; Notareschi, P.; et al. Swimming behaviour and magnetotaxis function of the marine bacterium strain MO-1. Environ. Microbiol. Rep. 2013, 6, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Poiata, A.; Creanga, D.E.; Morariu, V.V. Life in zero magnetic field. VE coli resistance to antibiotics. Electromagn. Biol. Med. 2003, 22, 171–182. [Google Scholar] [CrossRef]
- Creanga, D.; Poiata, A.; Morariu, V.; Tupu, P. Zero-magnetic field effect in pathogen bacteria. J. Magn. Magn. Mater. 2004, 272–276, 2442–2444. [Google Scholar] [CrossRef]
- Wang, X.K.; Ma, Q.F.; Jiang, W.; Lv, J.; Pan, W.D.; Song, T.; Wu, L.-F. Effects of Hypomagnetic Field on Magnetosome Formation of Magnetospirillum Magneticum AMB-1. Geomicrobiol. J. 2008, 25, 296–303. [Google Scholar] [CrossRef]
- Ilyin, V.K.; Orlov, O.I.; Morozova, Y.A.; Skedina, M.A.; Vladimirov, S.K.; Plotnikov, E.V.; Artamonov, A.A. Prognostic model for bacterial drug resistance genes horizontal spread in space-crews. Acta Astronaut. 2022, 190, 388–394. [Google Scholar] [CrossRef]


{kind=link}
| Impact on | Effect | Hypomagnetic Field Properties | ||||
|---|---|---|---|---|---|---|
| Organism | Mechanism | B (nT) | Duration | References | ||
| Mineral density of bones | Reduction | Sprague-Dawley rats | Shielding | <300 | 3 days | [30] |
| The concentration of Fe, Mn, Cu, Cr | Reduction | Fur of laboratory rats Wistar | Shielding | <20 | 7 months | [31] |
| Ca2+ dependent proteases | Inactivation | Enzymes from fish and invertebrates | Compensation | 1 h | [32] | |
| The concentration of Co, Ni | No effect | Fur of laboratory rats Wistar | Shielding | <20 | 8 months | [31] |
| Mitochondrial activity | Reduction | Skeletal muscle cells | Compensation | <200 | 7 days | [33] |
| Mitochondrial activity | Reduction | Mouse (C57BL/6) | Compensation | 0–500 | 30 days | [34] |
| ATP levels | Reduction | Skeletal muscle cells | Compensation | <30,000 | 3 days | [35] |
| Cell respiration | Reduction | Drosophila melanogaster | Compensation | 1 | 6 h | [27] |
| Cell respiration | Promotion | Drosophila melanogaster | Compensation | 1 | 1 h | [27] |
| Impact on | Effect | Organism | Hypomagnetic Field Properties | ||||
|---|---|---|---|---|---|---|---|
| Mechanism of Generation | B (nT) | Duration | References | ||||
| Neural system | ROS levels | Reduction | Mouse (C57BL/6 J), males | Shielding | 170 | Every 3 days/150 days | [23] |
| ROS levels | Reduction | Peritoneal mice neutrophils | Shielding | 20 | 1.5 h | [21] | |
| Growth | Promotion | Primary neural progenitor/mouse stem cells | Shielding | 0–200 | 7 days | [42] | |
| ROS levels | Reduction | Human cells of neuroblast | Shielding | 0–200 | 16 h | [44] | |
| ROS genes expression | Reduction | Mouse (C57BL/6 J), males | Shielding | 170 | 3 day/150 days | [23] | |
| Gene expression | Reduction (down-regulation) | Human neuroblast cells | Compensation | <200 | 2 days | [45] | |
| Migratory properties | Reduction | Human cells of neuroblast | Shielding | 0–200 | 48 h | [46] | |
| Proliferation | Promotion | Human cells neuroblast (SH-SY5Y) | Shielding | 0–200 | 3 days | [46] | |
| Memory | Reduction | Drosophila melanogaster | Compensation | 100–680 | 10–19 generations | [47] | |
| Proliferation | Promotion | Human neuroblastoma cells | Shielding | - | - | [48] | |
| Cognitive abilities | Reduction | Human (volunteers) | Compensation | 400 | 45 min | [49] | |
| Proliferation | Promotion | Human neuroblastoma (SHSY5Y) cells | Shielding | <200 | 3 days | [50] | |
| Hippocampal neurogenesis | Inhibition | Mouse (C57BL/6 J), males | Shielding | 170 | every 3 day/150 days | [23] | |
| Cardiovascular system | Blood pressure | Promotion | Human (volunteers) | Compensation | ±10 | 60 min | [51] |
| Blood circulation | Promotion | Human (volunteers) | Compensation | ±10 | 60 min | [52] | |
| Haemolysis | Promotion | Human blood | Compensation | 100 | 72 h | [53] | |
| Haemolysis | Promotion | Blood of rats | Compensation | 192 | 6 h to 4 weeks | [54] | |
| Life cycle and survival | Survival | Reduction | Milnesium inceptum | Shielding | - | 21 days | [29] |
| Survival | Reduction | Tardigada (Echiniscus testudo and Milnesium inceptum) | Shielding | - | 21 days | [29] | |
| Life expectancy | Reduction | Daphninia magne | Compensation | 15 | Generational period | [55] | |
| Larval development | Inhibition | Mythimna separata | Compensation | <500 | 12 h | [56] | |
| Development of eggs and nymphs | Delayed | Nilaparvata lugens | Compensation | 0–1060 | Generational period | [57] | |
| Fertility | Reduction | Nilaparvata lugens | Compensation | 0–1060 | Generational period | [58] | |
| Production of abnormal embryoys | Promotion | Xenopus larvae | Shielding | 104 ± 12.6 | 4 days | [59] | |
| Fertility | Reduction (sterility) | NMRI mouse zygotes | Shielding | 200 | 12 days | [60] | |
| Abortion | Promotion | Pregnant NMRI mice | Shielding | 200 | 3–12 days | [60] | |
| Survival of cells exposed to X-rays | Promotion | Immortalised human bronchial epithelial cells | Shielding | <50 | 24 h | [61] | |
| Chromatic condensation | Changes | Human fibroblasts and lymphocytes | Compensation | 1800 | 20–70 min | [62] | |
| Impact on | Effect | Organism | Hypomagnetic Field Properties | |||
|---|---|---|---|---|---|---|
| Mechanism of Generation | B (nT) | Duration | References | |||
| Growth | Reduction | Glycine max | Shielding | 111 ± 15 | 24 h | [69] |
| Growth | Reduction | Arabidopsis thaliana | Compensation | 0–1330 | 35 days | [70] |
| Growth | Reduction | Arabidopsis thaliana | Compensation | 40–44 | 96 h | [71] |
| Epicotyl elongation | Promotion | Pisum sativum | Shielding | - | 24 h | [72] |
| Gene expansion | Reduction | Arabidopsis thaliana | Compensation | 0–1330 | 33 days | [68] |
| Activity of photoreceptors phyA | Reduction | Arabidopsis thaliana | Compensation | 40 | 3 h | [64] |
| Activity of phyB photoreceptors | Promotion | Arabidopsis thaliana | Compensation | 40 | 3 h | [64] |
| The content of auxin in flower | Reduction | Arabidopsis thaliana | Compensation | 0–1330 | 33 days | [67] |
| Gene expression (associated with flowering) | Promotion | Arabidopsis thaliana | Compensation | 50 | 33 days | [73] |
| Auxin content in roots | Promotion | Arabidopsis thaliana | Compensation | 1–1330 | 33 days | [67] |
| Iron intake by roots | Reduction | Arabidopsis thaliana | Compensation | 40–44 | 96 h | [71] |
| Concentration of Ca2+ ions | Promotion | Pisum sativum (root system) | Shielding | 0.5–2 | 3 days | [41] |
| Impact on | Effect | Organism | Field Properties | |||
|---|---|---|---|---|---|---|
| Mechanism | B (nT) | Duration | References | |||
| Growth and number of cells | Reduction | Magnetotactic bacteria (MO-1) | Shielding | 2 | 2 days | [79] |
| Tolerance to antibiotics | Both reduction and promotion | Escherichia coli | Compensation | - | 6 days | [80] |
| Tolerance to antibiotics | Both reduction and promotion | Pseudomonas and Enterobacter strains | Field compensation | - | 6 days | [81] |
| Magnetosome size | Promotion | Magnetospirillum magneticum AMB-1 | Compensation | 500 | 16 h | [82] |
| Gene expression | Modification | Magnetospirillum magneticum AMB-1 | Compensation | 500 | 16 h | [82] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinčák, M.; Sedlakova-Kadukova, J. Hypomagnetic Fields and Their Multilevel Effects on Living Organisms. Processes 2023, 11, 282. https://doi.org/10.3390/pr11010282
Sinčák M, Sedlakova-Kadukova J. Hypomagnetic Fields and Their Multilevel Effects on Living Organisms. Processes. 2023; 11(1):282. https://doi.org/10.3390/pr11010282
Chicago/Turabian StyleSinčák, Miroslava, and Jana Sedlakova-Kadukova. 2023. "Hypomagnetic Fields and Their Multilevel Effects on Living Organisms" Processes 11, no. 1: 282. https://doi.org/10.3390/pr11010282
APA StyleSinčák, M., & Sedlakova-Kadukova, J. (2023). Hypomagnetic Fields and Their Multilevel Effects on Living Organisms. Processes, 11(1), 282. https://doi.org/10.3390/pr11010282