
Dendrobine Inhibits γ-Irradiation-Induced Cancer Cell Migration, Invasion and Metastasis in Non-Small Cell Lung Cancer Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Chemicals
2.2. Cell Culture and Treatment
2.3. Cell Viability Assay
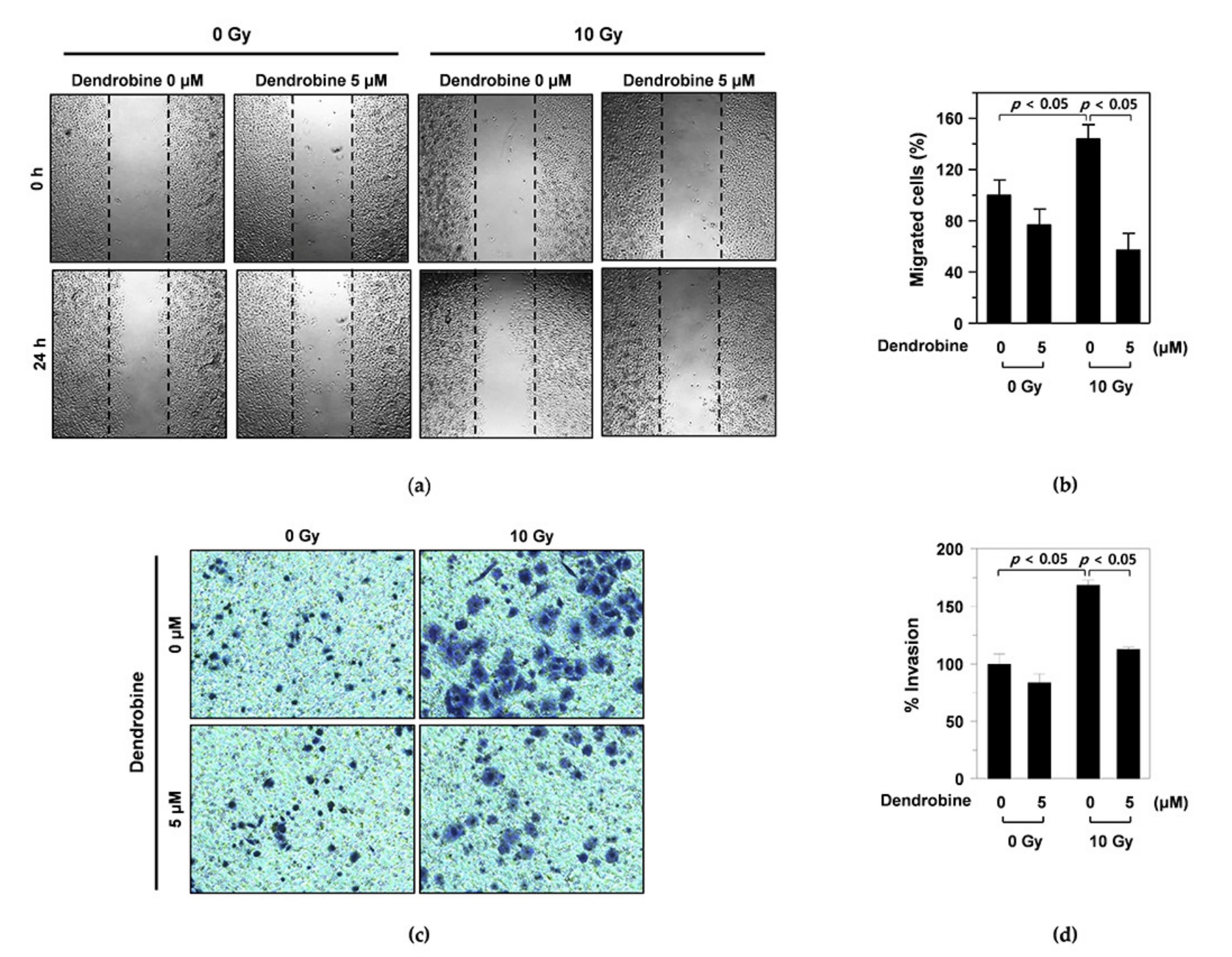
2.4. Wound-Healing Assay
2.5. Invasion Assay
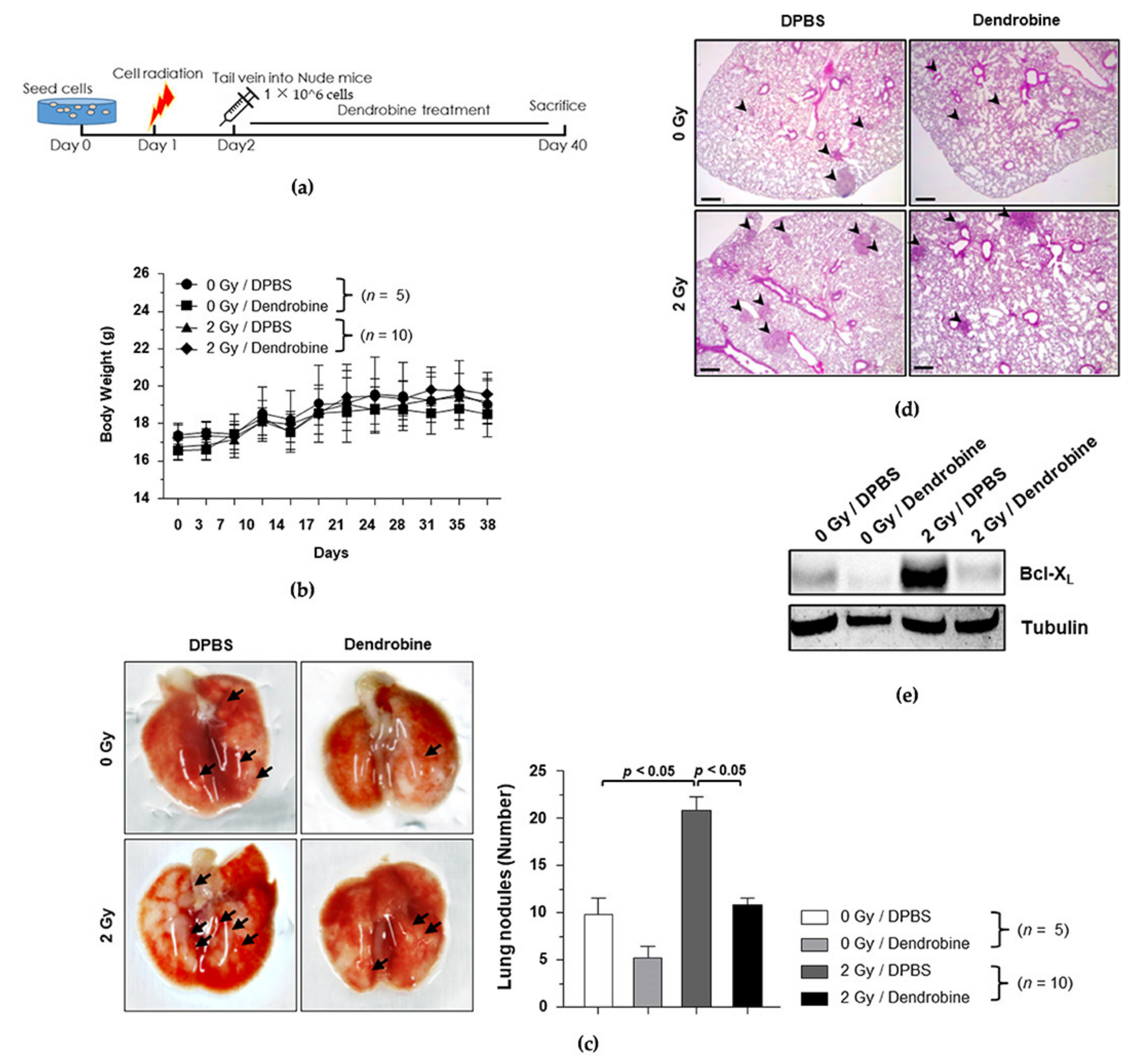
2.6. Animal Study
2.7. IR-Induced Metastasis Mouse Model
2.8. A549 Xenograft Model
2.9. Metastasis Assay in the IR-Induced Metastasis Mouse Model
2.10. Western Blotting
2.11. Enzyme-Linked Immunosorbent Assay (ELISA)
2.12. qRT-PCR
2.13. Statistical Analysis
3. Results
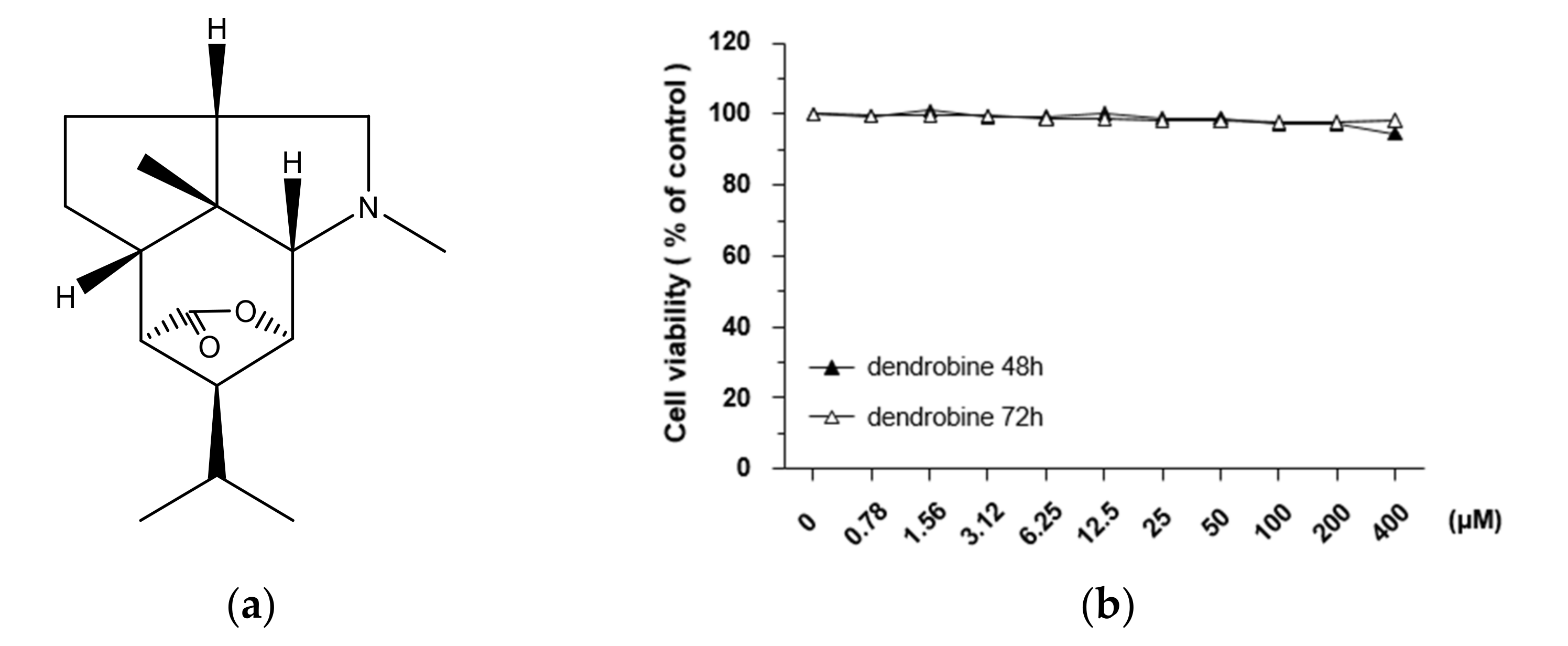
3.1. Effects of Dendrobine on A549 Cell Viability
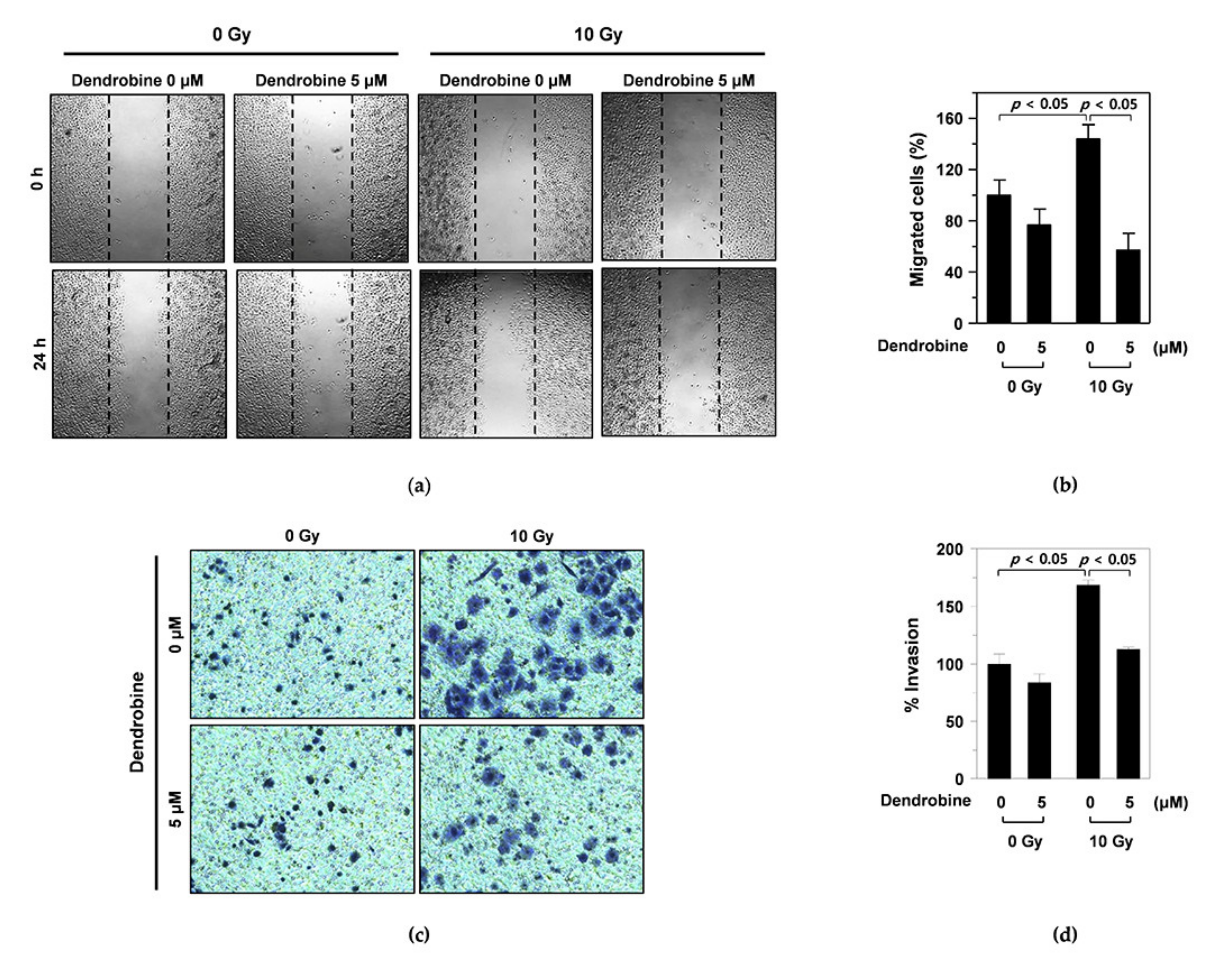
3.2. Inhibitory Effects of Dendrobine on IR-Induced Migration and Invasion in A549 Cells
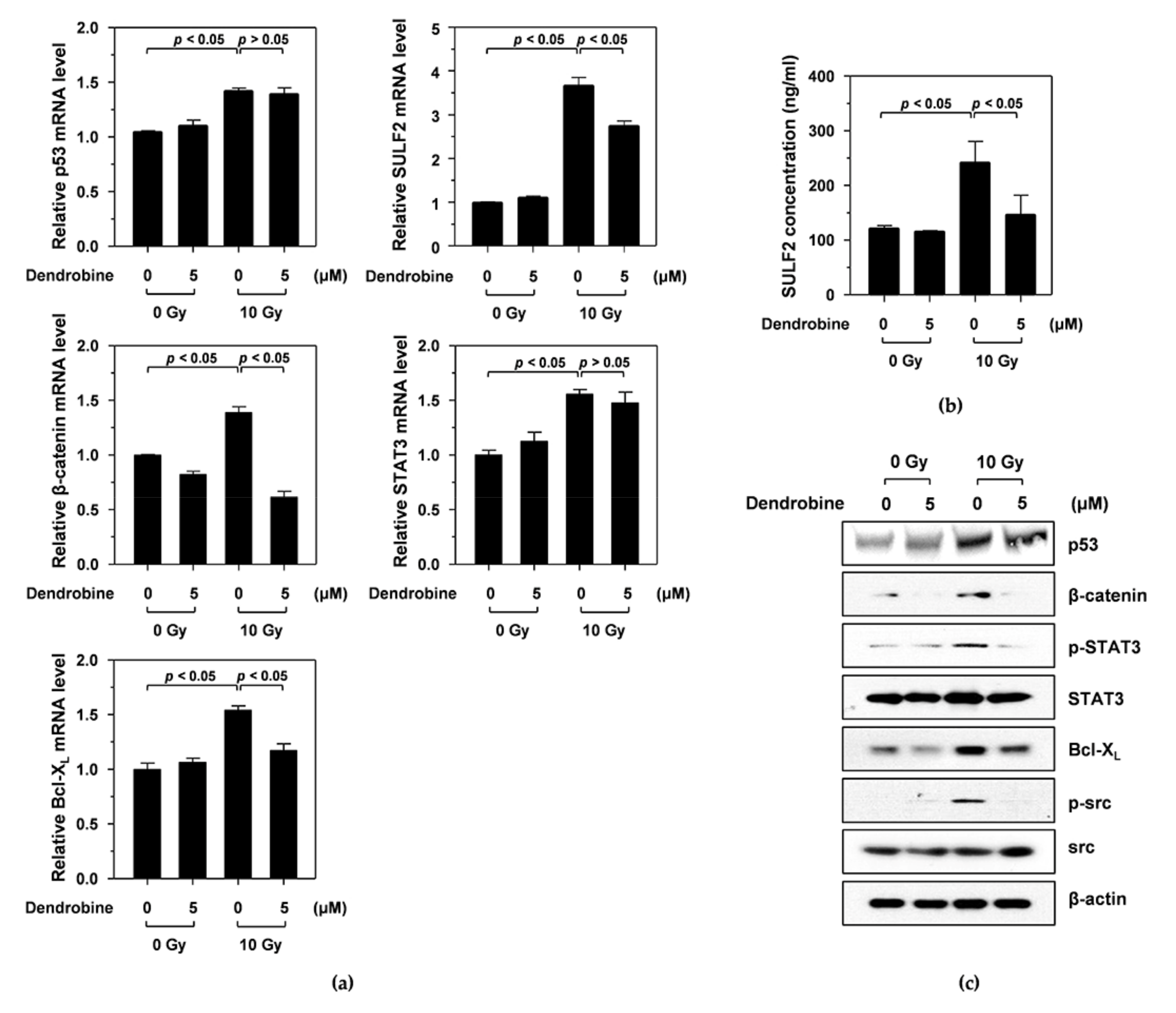
3.3. Dendrobine Inhibits Ir-Induced Cancer Cell Migration and Invasion by Suppressing SULF2 mRNA Levels
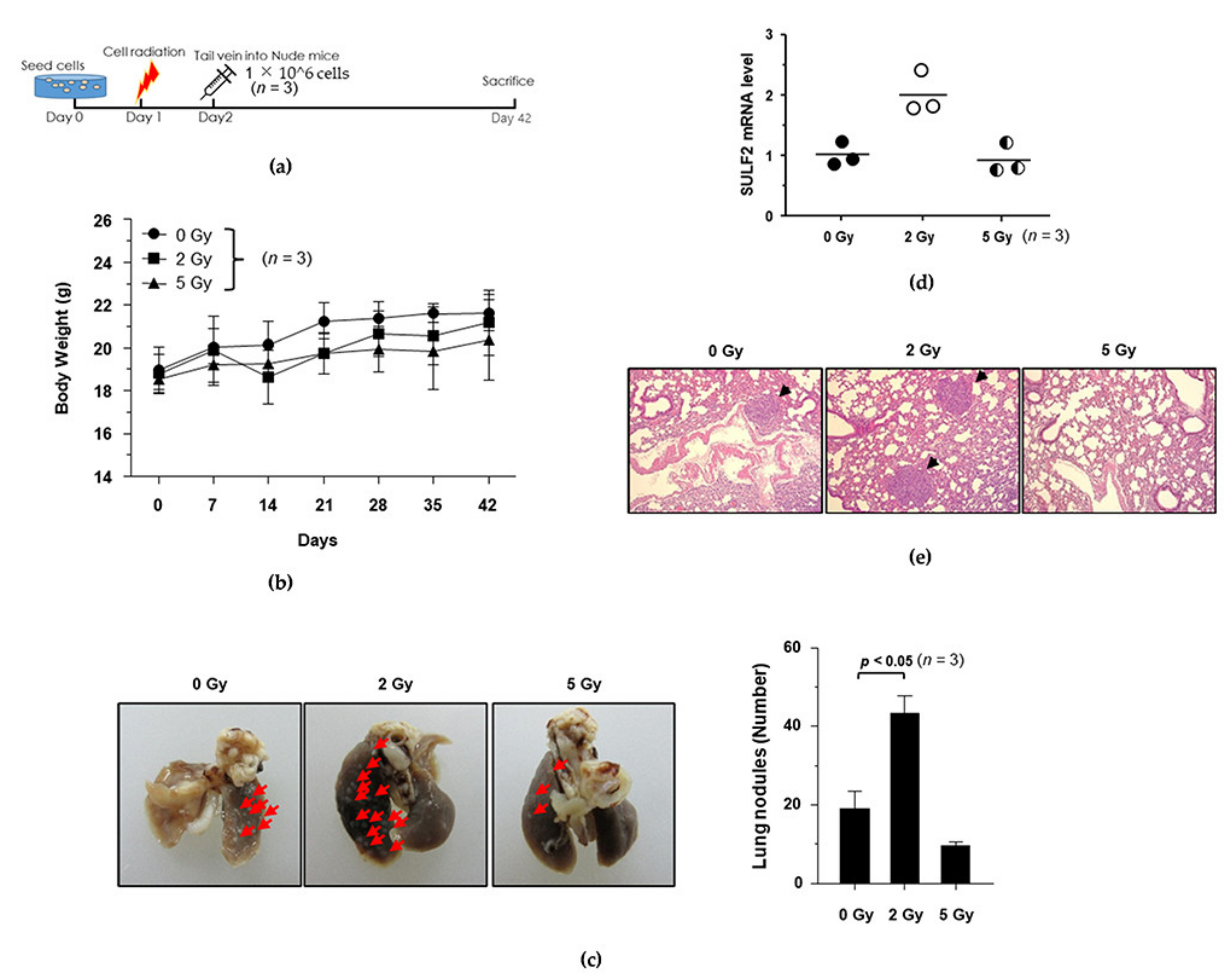
3.4. Establishment of the IR-Induced Metastasis Mouse Model
3.5. Effects of Dendrobine on Tumor Growth in the Xenograft Model
3.6. Anti-Metastasis Efficacy of Dendrobine in the Mouse Model of IR-Induced Metastasis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Riihimäki, M.; Hemminki, A.; Fallah, M.; Thomsen, H.; Sundquist, K.; Sundquist, J.; Hemminki, K. Metastatic sites and survival in lung cancer. Lung Cancer 2014, 86, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Lemjabbar-Alaoui, H.; Hassan, O.U.; Yang, Y.W.; Buchanan, P. Lung cancer: Biology and treatment options. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2015, 1856, 189–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azzam, E.I.; Jay-Gerin, J.P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.N.; Kang, G.Y.; Lee, S.S.; Kim, J.; Bae, I.H.; Hwang, S.-G.; Um, H.-D. Bcl-XL and STAT3 mediate malignant actions of gamma-irradiation in lung cancer cells. Cancer Sci. 2010, 101, 1417–1423. [Google Scholar] [CrossRef]
- Moncharmont, C.; Levy, A.; Guy, J.B.; Falk, A.T.; Guilbert, M.; Trone, J.-C.; Alphonse, G.; Gilormini, M.; Ardail, D.; Toillon, R.A.; et al. Radiation-enhanced cell migration/invasion process: A review. Crit. Rev. Oncol./Hematol. 2014, 92, 133–142. [Google Scholar] [CrossRef]
- Camphausen, K.; Moses, M.A.; Beecken, W.D.; Khan, M.K.; Folkman, J.; O’Reilly, M.S. Radiation therapy to a primary tumor accelerates metastatic growth in mice. Cancer Res. 2001, 61, 2207–2211. [Google Scholar]
- Vilalta, M.; Rafat, M.; Graves, E.E. Effects of radiation on metastasis and tumor cell migration. Cell. Mol. Life Sci. 2016, 73, 2999–3007. [Google Scholar] [CrossRef] [Green Version]
- Rosen, S.D.; Lemjabbar-Alaoui, H. Sulf-2: An extracellular modulator of cell signaling and a cancer target candidate. Expert Opin. Ther. Targets 2010, 14, 935–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elgundi, Z.; Papanicolaou, M.; Major, G.; Cox, T.R.; Melrose, J.; Whitelock, J.M.; Farrugia, B.L. Cancer metastasis: The role of the extracellular matrix and the heparan sulfate proteoglycan perlecan. Front. Oncol. 2019, 9, 1482. [Google Scholar] [CrossRef] [Green Version]
- Morimoto-Tomita, M.; Uchimura, K.; Bistrup, A.; Lum, D.H.; Egeblad, M.; Boudreau, N.; Werb, Z.; Rosen, S.D. Sulf-2, a proangiogenic heparan sulfate endosulfatase, is upregulated in breast cancer. Neoplasia 2005, 7, 1001–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.D.; Sun, Z.; Hu, C.; Lai, J.; Dove, R.; Nakamura, I.; Lee, J.-S.; Thorgeirsson, S.S.; Kang, K.J.; Chu, I.-S.; et al. Sulfatase 1 and sulfatase 2 in hepatocellular carcinoma: Associated signaling pathways, tumor phenotypes, and survival. Genes Chromosomes Cancer 2011, 50, 122–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lui, N.S.; Yang, Y.W.; van Zante, A.; Buchanan, P.; Jablons, D.M.; Lemjabbar-Alaoui, H. SULF2 expression is a potential diagnostic and prognostic marker in lung cancer. PLOS ONE 2016, 11, e0148911. [Google Scholar] [CrossRef]
- Jung, C.-H.; Ho, J.N.; Park, J.K.; Kim, E.M.; Hwang, S.G.; Um, H.D. Involvement of SULF2 in y-irradiation-induced invasion and resistance of cancer cells by inducing IL-6 expression. Oncotarget 2016, 7, 16090–16103. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Avula, B.; Abe, N.; Wei, F.; Wang, M.; Ma, S.-C.; Ali, Z.; Elsohly, M.A.; Khan, I.A. Tandem mass spectrometry for structural identification of sesquiterpene alkaloids from the stems of Dendrobium nobile using LC-QToF. Planta Med. 2016, 82, 662–670. [Google Scholar] [CrossRef] [Green Version]
- Bulpitt, C.J.; Li, Y.; Bulpitt, P.F.; Wang, J. The use of orchids in Chinese medicine. J. R. Soc. Med. 2007, 100, 558–563. [Google Scholar] [CrossRef]
- Kudo, Y.; Tanaka, A.; Yamada, K. Dendrobine, an antagonist of beta-alanine, taurine and of presynaptic inhibition in the frog spinal cord. Br. J. Pharmacol. 1983, 78, 709–715. [Google Scholar] [CrossRef]
- Nie, J.; Tian, Y.; Zhang, Y.; Lu, Y.L.; Li, L.S.; Shi, J.S. Dendrobium alkaloids prevent Aβ25-35-induced neuronal and synaptic loss via promoting neurotrophic factors expression in mice. PeerJ 2016, 4, e2739. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhou, J.; Xu, S.; Li, J.; Liu, J.; Lu, Y.; Shi, J.; Zhou, S.; Wu, Q. Induction of Nrf2 pathway by Dendrobium nobile Lindl. alkaloids protect against carbon tetrachloride induced acute liver injury. Biomed. Pharmacother. 2019, 117, 109073. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Liu, T.; Liu, M.; Chen, F.; Liu, S.; Yang, J. Anti-influenza A virus activity of dendrobine and its mechanism of action. J. Agric. Food Chem. 2017, 65, 3665–3674. [Google Scholar] [CrossRef]
- Feng, Y.; Jia, B.; Feng, Q.; Zhang, Y.; Chen, Y.; Meng, J. Dendrobine attenuates gestational diabetes mellitus in mice by inhibiting Th17 cells. Basic Clin. Pharmacol. Toxicol. 2021, 128, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Song, T.H.; Chen, X.X.; Lee, C.K.; Sze, S.C.; Feng, Y.-B.; Yang, Z.-J.; Chen, H.-Y.; Li, S.-T.; Zhang, L.-Y.; Wei, G.; et al. Dendrobine targeting JNK stress signaling to sensitize chemotoxicity of cisplatin against non-small cell lung cancer cells in vitro and In Vivo. Phytomedicine 2019, 53, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.-H.; Han, A.-R.; Chung, H.-J.; Ha, I.-H.; Um, H.-D. Linarin inhibits radiation-induced cancer invasion by downregulating MMP-9 expression via the suppression of NF-κB activation in human non-small-cell lung cancer A549. Nat. Prod. Res. 2019, 33, 3582–3586. [Google Scholar] [CrossRef]
- Jung, C.-H.; Kim, E.M.; Song, J.Y.; Park, J.K.; Um, H.-D. Mitochondrial superoxide dismutase 2 mediates γ-irradiation-induced cancer cell invasion. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Han, A.-R.; Kang, U.; Kim, J.-B.; Seo, E.K.; Jung, C.-H. Inhibitory effects of furanocoumarins from the roots of Angelica dahurica on ionizing radiation-induced migration of A549 human non-small cell lung cancer cells. Nat. Prod. Commun. 2020, 15, 1–6. [Google Scholar]
- Han, A.-R.; Lee, S.; Han, S.; Lee, Y.J.; Kim, J.-B.; Seo, E.K.; Jung, C.-H. Triterpenoids from the leaves of Centella asiatica inhibit ionizing radiation-induced migration and invasion of human lung cancer cells. Evid.-Based Complementary Altern. Med. 2020, 2020, 3683460. [Google Scholar] [CrossRef]
- Lee, Y.H.; Park, J.D.; Baek, N.I.; Kim, S.I.; Ahn, B.Z. In vitro and in vivo antitumoral phenanthrenes from the aerial parts of Dendrobium nobile. Planta Med. 1995, 61, 178–180. [Google Scholar] [CrossRef]
- Wang, J.H.; Luo, J.P.; Zha, X.Q.; Feng, B.J. Comparison of antitumor activities of different polysaccharide fractions from the stems of Dendrobium nobile Lindl. Carbohydr. Polym. 2010, 79, 114–118. [Google Scholar] [CrossRef]
- Li, Q.; Ding, G.; Li, B.; Guo, S.X. Transcriptome analysis of genes involved in dendrobine biosynthesis in Dendrobium nobile Lindl. infected with mycorrhizal fungus MF23 (Mycena sp.). Sci. Rep. 2017, 7, 316. [Google Scholar] [CrossRef] [Green Version]
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment. Int. J. Oncol. 2019, 54, 407–419. [Google Scholar] [PubMed]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.W. Cancer and radiation therapy: Current advances and future directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | |
|---|---|---|
| p53 | Forward | CAT GAG CGC TGC TCA GAT AG |
| Reverse | TGG TAC AGT CAG AGC CAA CCT | |
| SULF2 | Forward | TGT CAT TGT CTC TCT TGT GTA GC |
| Reverse | AAT CCA TCC TCA AGC TGC TG | |
| β-catenin | Forward | GTC CTC TGT GAA CTT GCT CAG |
| Reverse | CCT CAG ACA TTC GGA ACA AAA | |
| STAT3 | Forward | CCC CGC ACT TTA GAT TCA TTG |
| Reverse | AGG TCA ACT CCA TGT CAA AGG | |
| GAPDH | Forward | ACT CCA CTC ACG GCA AAT TC |
| Reverse | TCT CCA TGG TGG TGA AGA CA | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.-R.; Han, A.-R.; Kim, J.-B.; Jung, C.-H. Dendrobine Inhibits γ-Irradiation-Induced Cancer Cell Migration, Invasion and Metastasis in Non-Small Cell Lung Cancer Cells. Biomedicines 2021, 9, 954. https://doi.org/10.3390/biomedicines9080954
Kim Y-R, Han A-R, Kim J-B, Jung C-H. Dendrobine Inhibits γ-Irradiation-Induced Cancer Cell Migration, Invasion and Metastasis in Non-Small Cell Lung Cancer Cells. Biomedicines. 2021; 9(8):954. https://doi.org/10.3390/biomedicines9080954
Chicago/Turabian StyleKim, Ye-Ram, Ah-Reum Han, Jin-Baek Kim, and Chan-Hun Jung. 2021. "Dendrobine Inhibits γ-Irradiation-Induced Cancer Cell Migration, Invasion and Metastasis in Non-Small Cell Lung Cancer Cells" Biomedicines 9, no. 8: 954. https://doi.org/10.3390/biomedicines9080954
APA StyleKim, Y.-R., Han, A.-R., Kim, J.-B., & Jung, C.-H. (2021). Dendrobine Inhibits γ-Irradiation-Induced Cancer Cell Migration, Invasion and Metastasis in Non-Small Cell Lung Cancer Cells. Biomedicines, 9(8), 954. https://doi.org/10.3390/biomedicines9080954