
Myeloid HIF1α Is Involved in the Extent of Orthodontically Induced Tooth Movement

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiment
2.2. RNA Analysis
2.3. MicroCT Analysis
2.4. Data Analysis and Statistics
3. Results
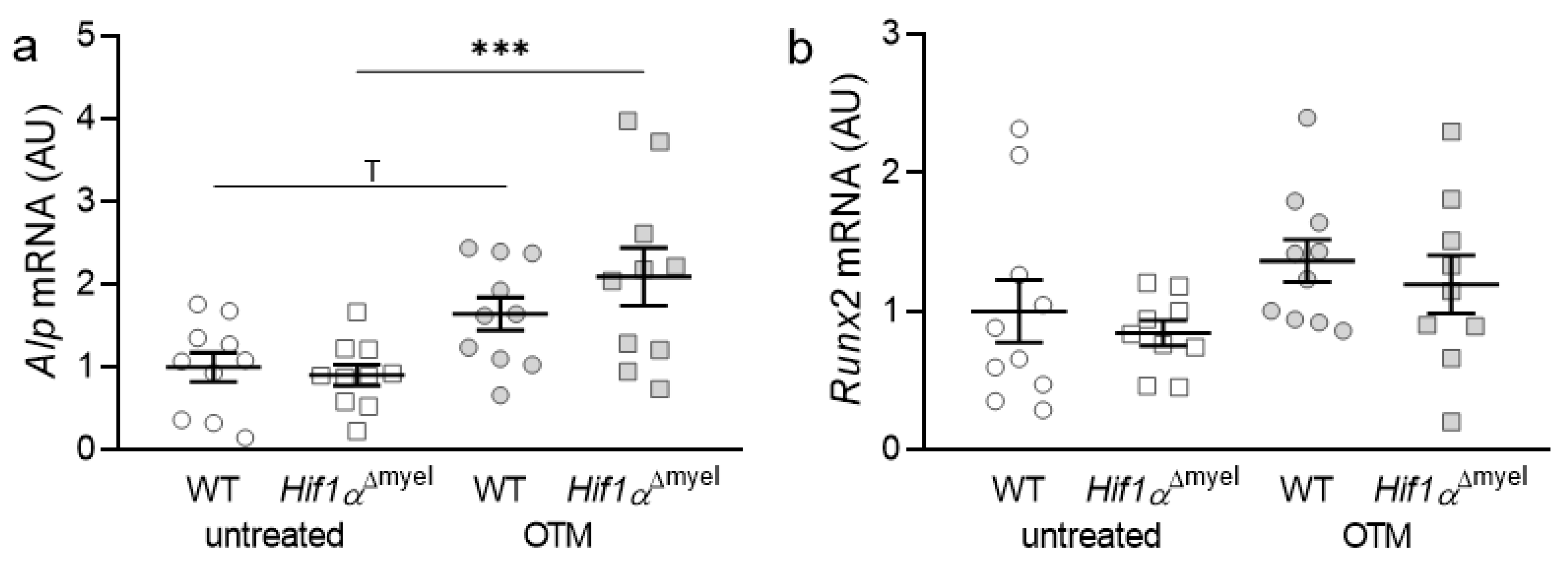
3.1. Effects of Myeloid Hif1α Deletion and Orthodontic Treatment on Genes Involved in Bone Formation
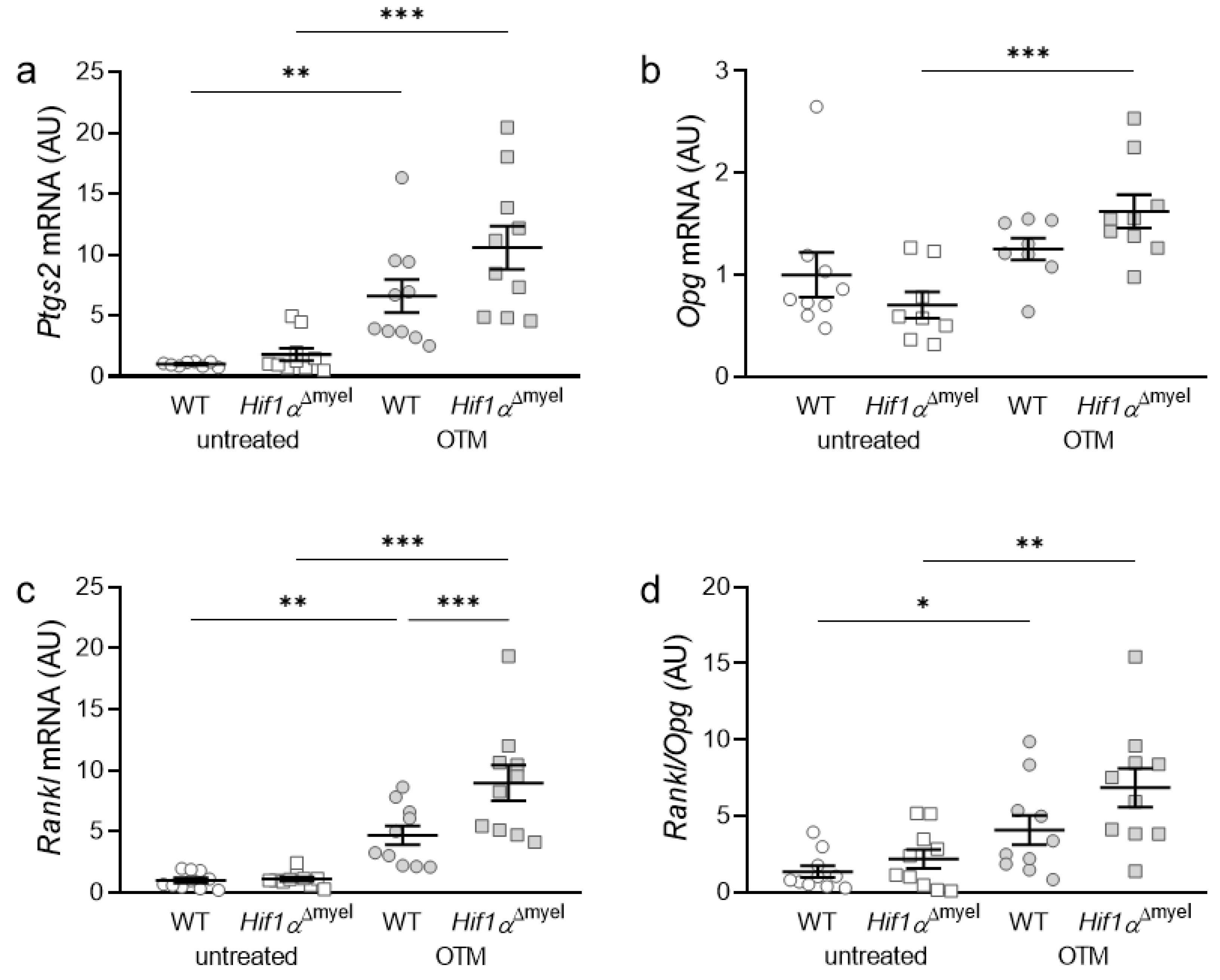
3.2. Effects of Myeloid Hif1α Deletion and Orthodontic Treatment on Genes Involved in Bone Modulation
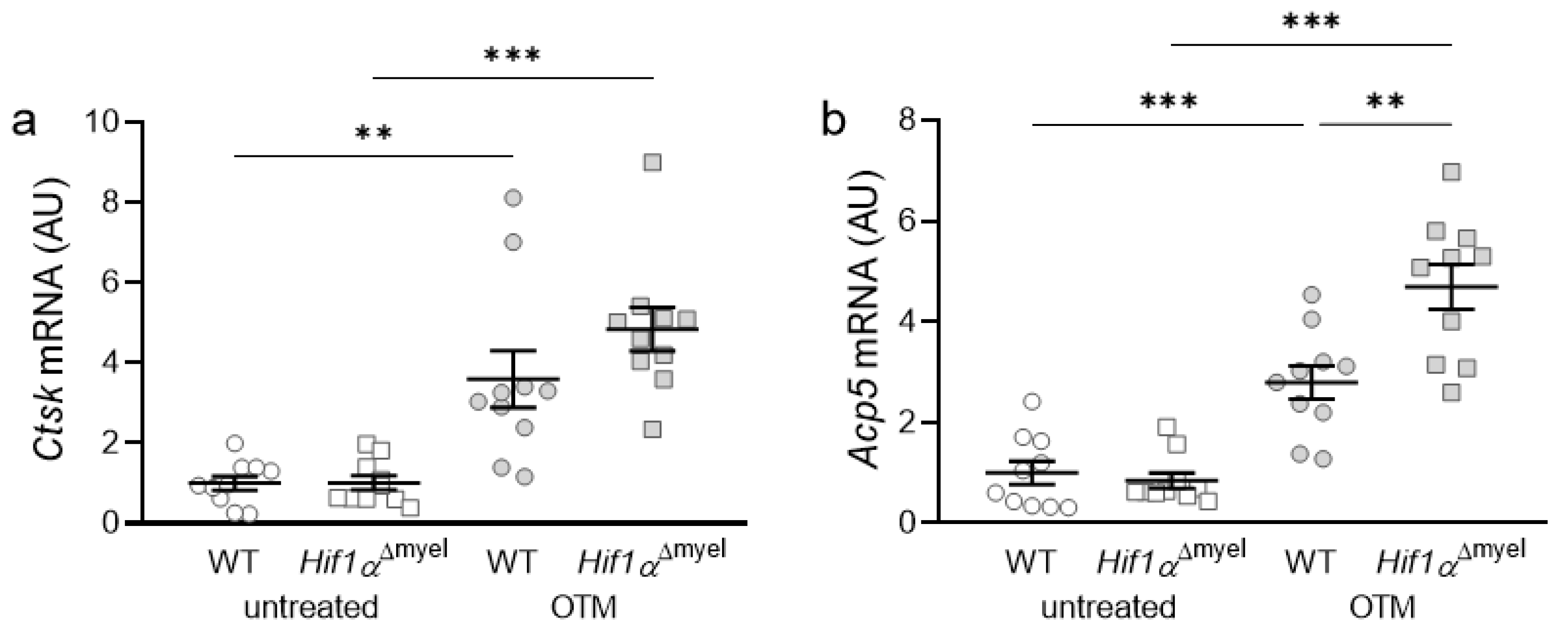
3.3. Effects of Myeloid Hif1α Deletion and Orthodontic Treatment on Genes Involved in Bone Resorption
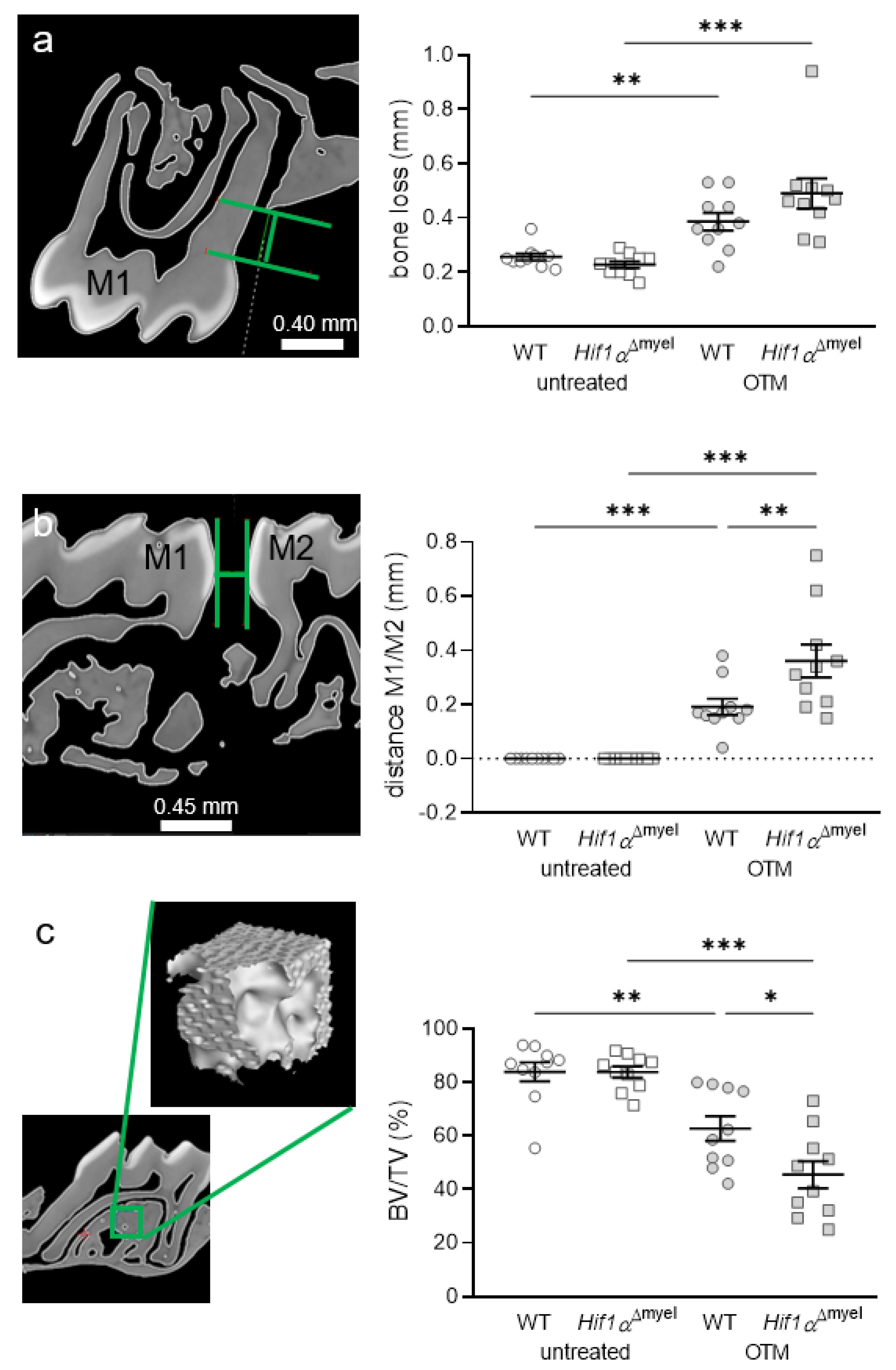
3.4. Effects of Myeloid Hif1α Deletion and Orthodontic Treatment on Periodontal Bone Loss and Orthodontic Tooth Movement
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.; Jacox, L.A.; Little, S.H.; Ko, C.-C. Orthodontic tooth movement: The biology and clinical implications. Kaohsiung J. Med. Sci. 2018, 34, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meikle, M.C. The tissue, cellular, and molecular regulation of orthodontic tooth movement: 100 years after Carl Sandstedt. Eur. J. Orthod. 2006, 28, 221–240. [Google Scholar] [CrossRef] [PubMed]
- Kirschneck, C.; Küchler, E.C.; Wolf, M.; Spanier, G.; Proff, P.; Schröder, A. Effects of the Highly COX-2-Selective Analgesic NSAID Etoricoxib on Human Periodontal Ligament Fibroblasts during Compressive Orthodontic Mechanical Strain. Mediat. Inflamm. 2019, 2019, 2514956. [Google Scholar] [CrossRef] [PubMed]
- Schröder, A.; Käppler, P.; Nazet, U.; Jantsch, J.; Proff, P.; Cieplik, F.; Deschner, J.; Kirschneck, C. Effects of Compressive and Tensile Strain on Macrophages during Simulated Orthodontic Tooth Movement. Mediat. Inflamm. 2020, 2020, 2814015. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.; Lossdörfer, S.; Marciniak, J.; Römer, P.; Kirschneck, C.; Craveiro, R.; Deschner, J.; Jäger, A. CD8+ T cells mediate the regenerative PTH effect in hPDL cells via Wnt10b signaling. Innate Immun 2016, 22, 674–681. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M. RANK/RANKL/OPG during orthodontic tooth movement. Orthod. Craniofac. Res. 2009, 12, 113–119. [Google Scholar] [CrossRef]
- Kanzaki, H.; Chiba, M.; Shimizu, Y.; Mitani, H. Periodontal ligament cells under mechanical stress induce osteoclastogenesis by receptor activator of nuclear factor kappaB ligand up-regulation via prostaglandin E2 synthesis. J. Bone Miner. Res. 2002, 17, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Schröder, A.; Küchler, E.C.; Omori, M.; Spanier, G.; Proff, P.; Kirschneck, C. Effects of ethanol on human periodontal ligament fibroblasts subjected to static compressive force. Alcohol 2019, 77, 59–70. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Caldeira, F.I.D.; Hidalgo, M.A.R.; De Carli-Dias, M.L.; Scarel-Caminaga, R.M.; Pigossi, S.C. Systematic review of ratios between disease /health periodontitis modulators and meta-analysis of their levels in gingival tissue and biological fluids. Arch. Oral Biol. 2021, 127, 105147. [Google Scholar] [CrossRef]
- Cavalla, F.; Letra, A.; Silva, R.M.; Garlet, G.P. Determinants of Periodontal/Periapical Lesion Stability and Progression. J. Dent. Res. 2021, 100, 29–36. [Google Scholar] [CrossRef]
- Proff, P.; Römer, P. The molecular mechanism behind bone remodelling: A review. Clin. Oral Investig. 2009, 13, 355–362. [Google Scholar] [CrossRef]
- Behm, C.; Nemec, M.; Weissinger, F.; Rausch, M.A.; Andrukhov, O.; Jonke, E. MMPs and TIMPs Expression Levels in the Periodontal Ligament during Orthodontic Tooth Movement: A Systematic Review of In Vitro and In Vivo Studies. Int. J. Mol. Sci. 2021, 22, 6967. [Google Scholar] [CrossRef]
- Allen, R.K.; Edelmann, A.R.; Abdulmajeed, A.; Bencharit, S. Salivary protein biomarkers associated with orthodontic tooth movement: A systematic review. Orthod. Craniofac. Res. 2019, 22, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Alhadlaq, A.M. Biomarkers of Orthodontic Tooth Movement in Gingival Crevicular Fluid: A Systematic Review. J. Contemp. Dent. Pract. 2015, 16, 578–587. [Google Scholar] [CrossRef]
- He, D.; Kou, X.; Yang, R.; Liu, D.; Wang, X.; Luo, Q.; Song, Y.; Liu, F.; Yan, Y.; Gan, Y.; et al. M1-like Macrophage Polarization Promotes Orthodontic Tooth Movement. J. Dent. Res. 2015, 94, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Schröder, A.; Barschkies, L.; Jantsch, J.; Proff, P.; Gölz, L.; Deschner, J.; Kirschneck, C. Role of Oxygen Supply in Macrophages in a Model of Simulated Orthodontic Tooth Movement. Mediat. Inflamm. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Masoud, G.N.; Li, W. HIF-1α pathway: Role, regulation and intervention for cancer therapy. Acta. Pharm. Sinica. B 2015, 5, 378–389. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Hypoxia-inducible factor 1 (HIF-1) pathway. Sci. STKE 2007, 2007, cm8. [Google Scholar] [CrossRef]
- Wan, C.; Gilbert, S.R.; Wang, Y.; Cao, X.; Shen, X.; Ramaswamy, G.; Jacobsen, K.A.; Alaql, Z.S.; Eberhardt, A.W.; Gerstenfeld, L.C.; et al. Activation of the hypoxia-inducible factor-1alpha pathway accelerates bone regeneration. Proc. Natl. Acad. Sci. USA 2008, 105, 686–691. [Google Scholar] [CrossRef] [Green Version]
- Ullrich, N.; Schröder, A.; Jantsch, J.; Spanier, G.; Proff, P.; Kirschneck, C. The role of mechanotransduction versus hypoxia during simulated orthodontic compressive strain-an in vitro study of human periodontal ligament fibroblasts. Int. J. Oral Sci. 2019, 11, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschneck, C.; Bauer, M.; Gubernator, J.; Proff, P.; Schröder, A. Comparative assessment of mouse models for experimental orthodontic tooth movement. Sci. Rep. 2020, 10, 12154. [Google Scholar] [CrossRef]
- Waldo, C.M.; Rothblatt, J.M. Histologic response to tooth movement in the laboratory rat; procedure and preliminary observations. J. Dent. Res. 1954, 33, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Kirschneck, C.; Proff, P.; Fanghänel, J.; Wolf, M.; Roldán, J.C.; Römer, P. Reference genes for valid gene expression studies on rat dental, periodontal and alveolar bone tissue by means of RT-qPCR with a focus on orthodontic tooth movement and periodontitis. Ann. Anat. 2016, 204, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Koretsi, V.; Kirschneck, C.; Proff, P.; Römer, P. Expression of glutathione peroxidase 1 in the spheno-occipital synchondrosis and its role in ROS-induced apoptosis. Eur. J. Orthod. 2015, 37, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Kirschneck, C.; Wolf, F.; Cieplik, F.; Blanck-Lubarsch, M.; Proff, P.; Schröder, A. Impact of NSAID etoricoxib on side effects of orthodontic tooth movement. Ann. Anat. 2020, 232, 151585. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, R.C.; Costa, A.D.L.L.; Freitas, R.D.A.; Bezerra, B.A.D.A.; Santos, B.R.M.D.; Pinto, L.P.; Gurgel, B.C.D.V. Immunoexpression of HIF-1α and VEGF in Periodontal Disease and Healthy Gingival Tissues. Braz. Dent. J. 2016, 27, 117–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gölz, L.; Memmert, S.; Rath-Deschner, B.; Jäger, A.; Appel, T.; Baumgarten, G.; Götz, W.; Frede, S. Hypoxia and P. gingivalis synergistically induce HIF-1 and NF-κB activation in PDL cells and periodontal diseases. Mediat. Inflamm. 2015, 2015, 438085. [Google Scholar] [CrossRef] [Green Version]
- Cramer, T.; Yamanishi, Y.; Clausen, B.E.; Förster, I.; Pawlinski, R.; Mackman, N.; Haase, V.H.; Jaenisch, R.; Corr, M.; Nizet, V.; et al. HIF-1alpha is essential for myeloid cell-mediated inflammation. Cell 2003, 112, 645–657. [Google Scholar] [CrossRef] [Green Version]
- Jantsch, J.; Chakravortty, D.; Turza, N.; Prechtel, A.T.; Buchholz, B.; Gerlach, R.G.; Volke, M.; Gläsner, J.; Warnecke, C.; Wiesener, M.S.; et al. Hypoxia and hypoxia-inducible factor-1 alpha modulate lipopolysaccharide-induced dendritic cell activation and function. J. Immunol. 2008, 180, 4697–4705. [Google Scholar] [CrossRef]
- Palazon, A.; Goldrath, A.W.; Nizet, V.; Johnson, R.S. HIF transcription factors, inflammation, and immunity. Immunity 2014, 41, 518–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschneck, C.; Thuy, M.; Leikam, A.; Memmert, S.; Deschner, J.; Damanaki, A.; Spanier, G.; Proff, P.; Jantsch, J.; Schröder, A. Role and Regulation of Mechanotransductive HIF-1α Stabilisation in Periodontal Ligament Fibroblasts. Int. J. Mol. Sci. 2020, 21, 9530. [Google Scholar] [CrossRef] [PubMed]
- Corrado, C.; Fontana, S. Hypoxia and HIF Signaling: One Axis with Divergent Effects. Int. J. Mol. Sci. 2020, 21, 5611. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Shao, J.; Gilbert, S.R.; Riddle, R.C.; Long, F.; Johnson, R.S.; Schipani, E.; Clemens, T.L. Role of HIF-1alpha in skeletal development. Ann. N. Y. Acad. Sci. 2010, 1192, 322–326. [Google Scholar] [CrossRef] [Green Version]
- Hulley, P.A.; Bishop, T.; Vernet, A.; Schneider, J.E.; Edwards, J.R.; Athanasou, N.A.; Knowles, H.J. Hypoxia-inducible factor 1-alpha does not regulate osteoclastogenesis but enhances bone resorption activity via prolyl-4-hydroxylase 2. J. Pathol. 2017, 242, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Ballanti, P.; Minisola, S.; Pacitti, M.T.; Scarnecchia, L.; Rosso, R.; Mazzuoli, G.F.; Bonucci, E. Tartrate-resistant acid phosphate activity as osteoclastic marker: Sensitivity of cytochemical assessment and serum assay in comparison with standardized osteoclast histomorphometry. Osteoporos. Int. 1997, 7, 39–43. [Google Scholar] [CrossRef]
- Dai, R.; Wu, Z.; Chu, H.Y.; Lu, J.; Lyu, A.; Liu, J.; Zhang, G. Cathepsin K: The Action in and Beyond Bone. Front. Cell Dev. Biol. 2020, 8, 433. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, Y.; Sato, Y.; Kobayashi, T.; Yoshida, S.; Mori, T.; Kanagawa, H.; Katsuyama, E.; Fujie, A.; Hao, W.; Miyamoto, K.; et al. HIF1α is required for osteoclast activation by estrogen deficiency in postmenopausal osteoporosis. Proc. Natl. Acad. Sci. USA 2013, 110, 16568–16573. [Google Scholar] [CrossRef] [Green Version]
- Tando, T.; Sato, Y.; Miyamoto, K.; Morita, M.; Kobayashi, T.; Funayama, A.; Kanaji, A.; Hao, W.; Watanabe, R.; Oike, T.; et al. Hif1α is required for osteoclast activation and bone loss in male osteoporosis. Biochem. Biophys. Res. Commun. 2016, 470, 391–396. [Google Scholar] [CrossRef]
- Hiraga, T.; Kizaka-Kondoh, S.; Hirota, K.; Hiraoka, M.; Yoneda, T. Hypoxia and hypoxia-inducible factor-1 expression enhance osteolytic bone metastases of breast cancer. Cancer Res. 2007, 67, 4157–4163. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Regulation of osteoblast differentiation by Runx2. Adv. Exp. Med. Biol. 2010, 658, 43–49. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Name | 5′-Forward Primer-3′ | 5′-Reverse Primer-3′ |
|---|---|---|---|
| Acp5 | Acid Phosphatase 5, Tartrate Resistant | ATACGGGGTCACTGCCTACC | TCGTTGATGTCGCACAGAGG |
| Alp | Alkaline Phosphatase | GGGTACAAGGCTAGATGGC | AGTTCAGTGCGGTTCCAGAC |
| Ctsk | Cathepsin K | GACCCATCTCTGTGTCCATCG | CCATAGCCCACCACCAACAC |
| Eef1a1 | Eukaryotic Translation Elongation Factor 1 Alpha 1 | AAAACATGATTACAGGCACATCCC | GCCCGTTCTTGGAGATACCAG |
| Opg | Osteoprotegerin | CCTTGCCCTGACCACTCTTAT | CACACACTCGGTTGTGGGT |
| Ptgs2 | Prostaglandin-Endoperoxide Synthase 2 | TCCCTGAAGCCGTACACATC | TCCCCAAAGATAGCATCTGGAC |
| Rankl | Receptor Activator of NF-κB Ligand | AAACGCAGATTTGCAGGACTC | CCCCACAATGTGTTGCAGTTC |
| Runx2 | Runt-related Transcription Factor 2 | CTCCCTGAACTCTGCACCAAG | GAGTGGATGGATGGGGATGTC |
| Ywhaz | Tryptophan 5-Monooxygenase Activation Protein Zeta | AATGCTTCGCAACCAGAAAGC | TGGTATGCTTGCTGTGACTGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirschneck, C.; Straßmair, N.; Cieplik, F.; Paddenberg, E.; Jantsch, J.; Proff, P.; Schröder, A. Myeloid HIF1α Is Involved in the Extent of Orthodontically Induced Tooth Movement. Biomedicines 2021, 9, 796. https://doi.org/10.3390/biomedicines9070796
Kirschneck C, Straßmair N, Cieplik F, Paddenberg E, Jantsch J, Proff P, Schröder A. Myeloid HIF1α Is Involved in the Extent of Orthodontically Induced Tooth Movement. Biomedicines. 2021; 9(7):796. https://doi.org/10.3390/biomedicines9070796
Chicago/Turabian StyleKirschneck, Christian, Nadine Straßmair, Fabian Cieplik, Eva Paddenberg, Jonathan Jantsch, Peter Proff, and Agnes Schröder. 2021. "Myeloid HIF1α Is Involved in the Extent of Orthodontically Induced Tooth Movement" Biomedicines 9, no. 7: 796. https://doi.org/10.3390/biomedicines9070796
APA StyleKirschneck, C., Straßmair, N., Cieplik, F., Paddenberg, E., Jantsch, J., Proff, P., & Schröder, A. (2021). Myeloid HIF1α Is Involved in the Extent of Orthodontically Induced Tooth Movement. Biomedicines, 9(7), 796. https://doi.org/10.3390/biomedicines9070796