
Increased Th1 Cells with Disease Resolution of Active Pulmonary Tuberculosis in Non-Atopic Patients

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Assessment of Atopic Status
2.3. Preparation of Cells in Bronchoalveolar Lavage (BAL) Fluid
2.4. Quantification of Cytokines and Nitrite Produced by Cultured BAL Cells
2.5. Preparation of Peripheral Blood Mononuclear Cells (PBMCs)
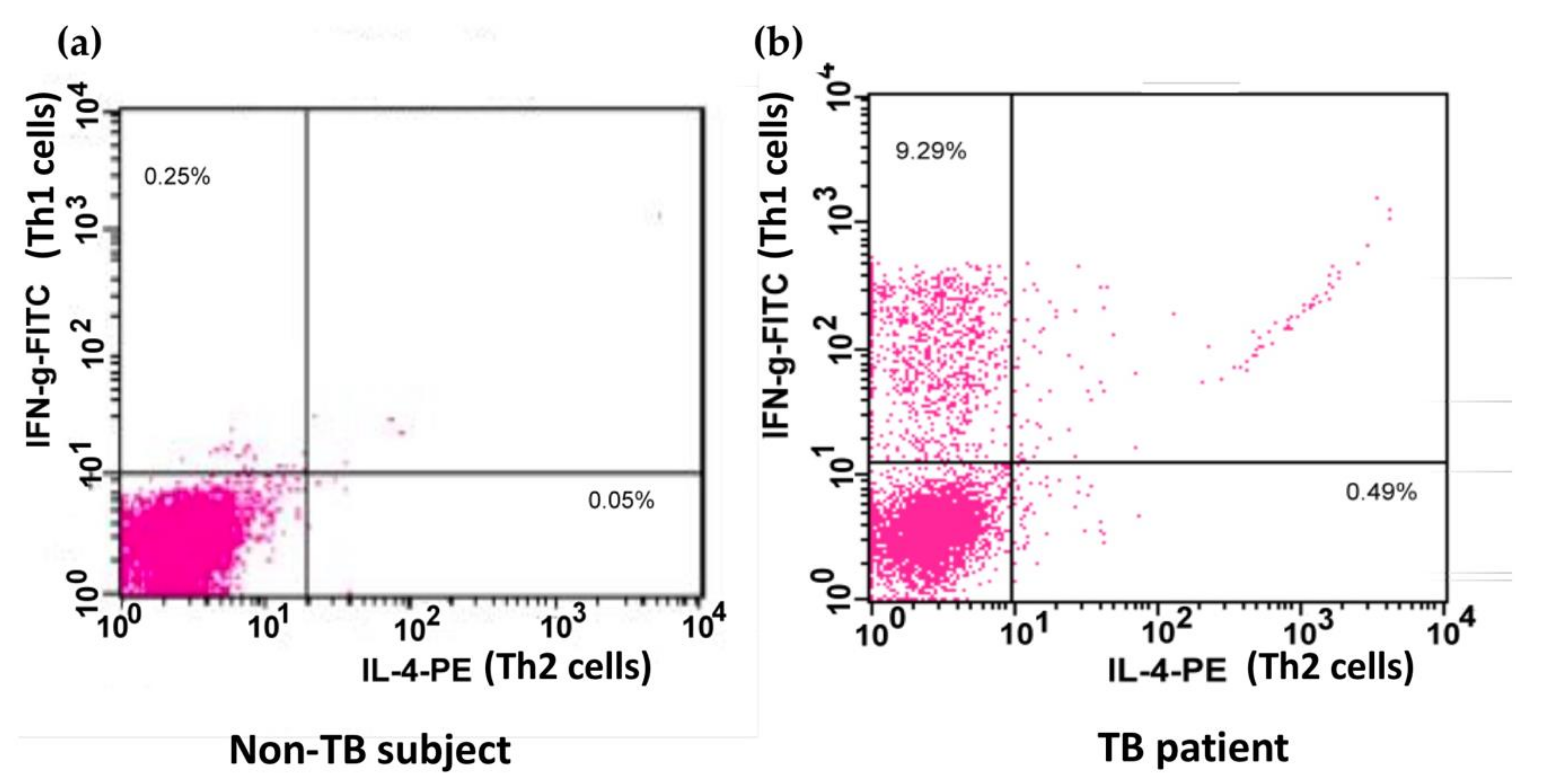
2.6. Identification of Th1 and Th2 Cells in Peripheral Blood and BAL Fluid
2.7. Statistical Analysis
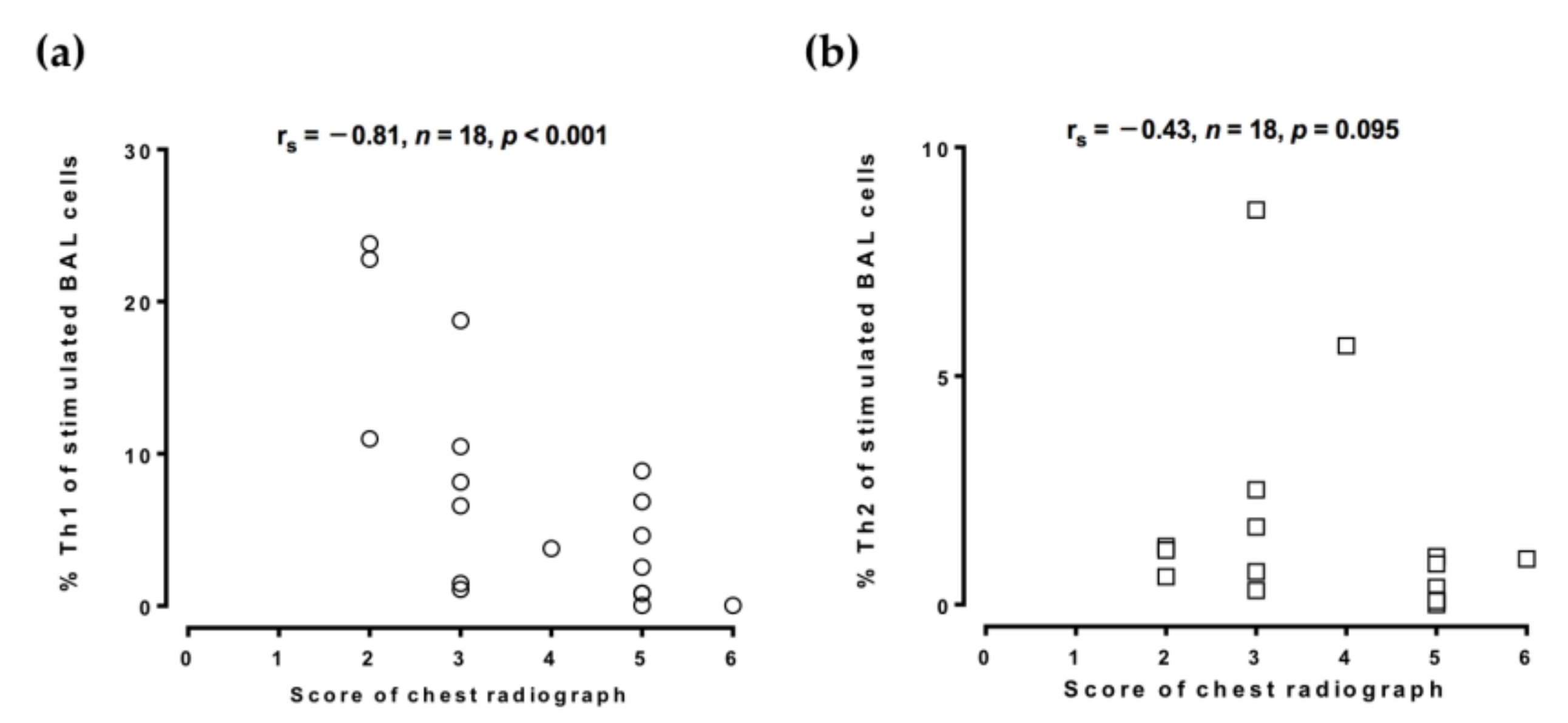
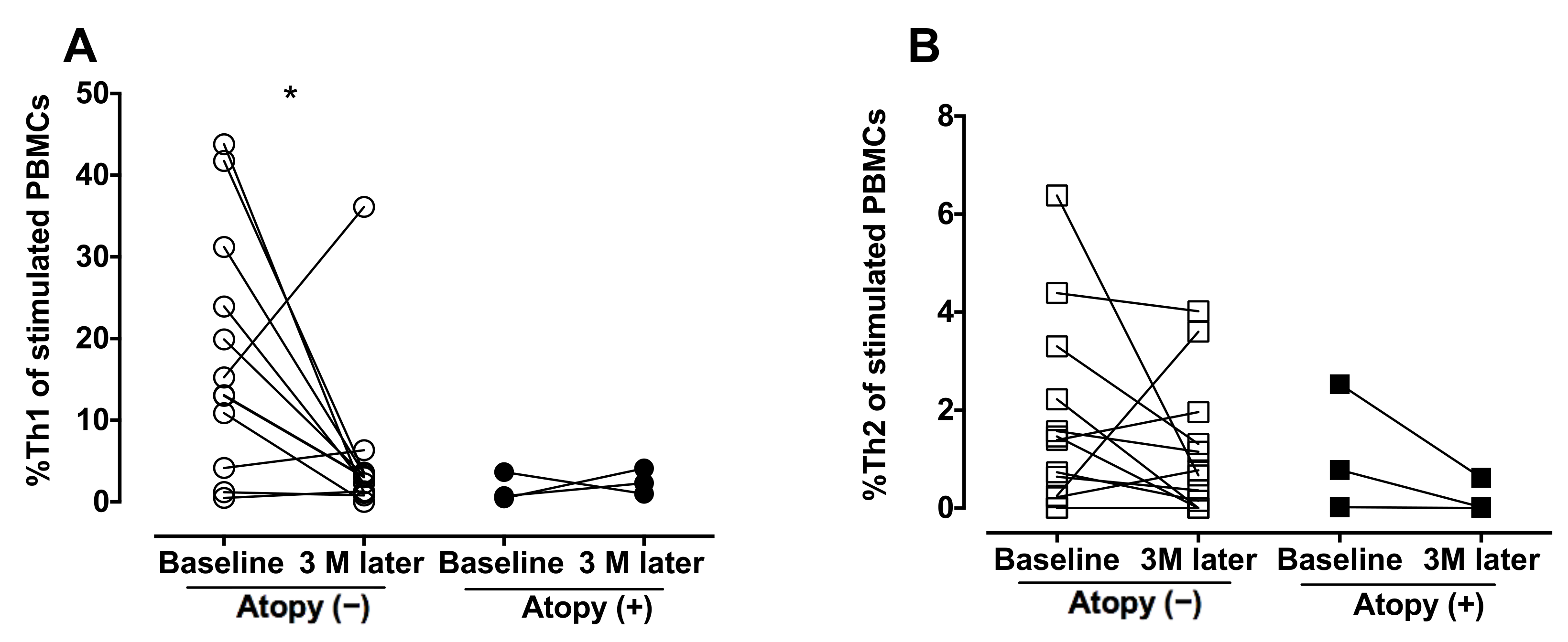
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walsh, K.P.; Mills, K.H. Dendritic cells and other innate determinants of T helper cell polarisation. Trends Immunol. 2013, 34, 521–530. [Google Scholar] [CrossRef]
- Furin, J.; Cox, H.; Pai, M. Tuberculosis. Lancet 2019, 393, 1642–1656. [Google Scholar] [CrossRef]
- Weiss, G.; Schaible, U.E. Macrophage defense mechanisms against intracellular bacteria. Immunol. Rev. 2015, 264, 182–203. [Google Scholar] [CrossRef] [Green Version]
- Flynn, J.L.; Chan, J.; Lin, P.L. Macrophages and control of granulomatous inflammation in tuberculosis. Mucosal Immunol. 2011, 4, 271–278. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.H.; Liu, C.Y.; Lin, H.C.; Yu, C.T.; Chung, K.F.; Kuo, H.P. Increased exhaled nitric oxide in active pulmonary tuberculosis due to inducible NO synthase upregulation in alveolar macrophages. Eur. Respir. J. 1998, 11, 809–815. [Google Scholar] [CrossRef] [Green Version]
- Errante, P.R.; Frazao, J.B.; Condino-Neto, A. The use of interferon-gamma therapy in chronic granulomatous disease. Recent Pat. Antiinfect Drug Discov. 2008, 3, 225–230. [Google Scholar] [CrossRef]
- Cooper, A.M.; Dalton, D.K.; Stewart, T.A.; Griffin, J.P.; Russell, D.G.; Orme, I.M. Disseminated tuberculosis in interferon gamma gene-disrupted mice. J. Exp. Med. 1993, 178, 2243–2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, A.M.; Magram, J.; Ferrante, J.; Orme, I.M. Interleukin 12 (IL-12) is crucial to the development of protective immunity in mice intravenously infected with mycobacterium tuberculosis. J. Exp. Med. 1997, 186, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.J.; Ryan, L.; LaCourse, R.; North, R.J. Properties and protective value of the secondary versus primary T helper type 1 response to airborne Mycobacterium tuberculosis infection in mice. J. Exp. Med. 2005, 201, 1915–1924. [Google Scholar] [CrossRef] [Green Version]
- Dlugovitzky, D.; Torres-Morales, A.; Rateni, L.; Farroni, M.A.; Largacha, C.; Molteni, O.; Bottasso, O. Circulating profile of Th1 and Th2 cytokines in tuberculosis patients with different degrees of pulmonary involvement. FEMS Immunol. Med. Microbiol. 1997, 18, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Nachiappan, A.C.; Rahbar, K.; Shi, X.; Guy, E.S.; Mortani Barbosa, E.J., Jr.; Shroff, G.S.; Ocazionez, D.; Schlesinger, A.E.; Katz, S.I.; Hammer, M.M. Pulmonary Tuberculosis: Role of Radiology in Diagnosis and Management. Radiographics 2017, 37, 52–72. [Google Scholar] [CrossRef]
- Sutton, B.J.; Gould, H.J. The human IgE network. Nature 1993, 366, 421–428. [Google Scholar] [CrossRef]
- Justiz Vaillant, A.A.; Jan, A. Atopy. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Thomsen, S.F. Epidemiology and natural history of atopic diseases. Eur. Clin. Respir. J. 2015, 2, 24642. [Google Scholar] [CrossRef] [Green Version]
- Yazdanbakhsh, M.; Kremsner, P.G.; van Ree, R. Allergy, parasites, and the hygiene hypothesis. Science 2002, 296, 490–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Q.; Xie, W.P.; Min, R.; Dai, G.Q.; Xu, C.C.; Pan, H.Q.; Miao, C.D.; Yang, Z.; Xu, W.G.; Wang, H. Characterization of Th1- and Th2-type immune response in human multidrug-resistant tuberculosis. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1233–1242. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.; De Haro, S.A.; Master, S.S.; Keane, J.; Roberts, E.A.; Delgado, M.; Deretic, V. T helper 2 cytokines inhibit autophagic control of intracellular Mycobacterium tuberculosis. Immunity 2007, 27, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Rook, G.A. Th2 cytokines in susceptibility to tuberculosis. Curr. Mol. Med. 2007, 7, 327–337. [Google Scholar] [CrossRef]
- Huang, K.H.; Wang, C.H.; Lee, K.Y.; Lin, S.M.; Lin, C.H.; Kuo, H.P. NF-κB repressing factor inhibits chemokine synthesis by peripheral blood mononuclear cells and alveolar macrophages in active pulmonary tuberculosis. PLoS ONE 2013, 8, e77789. [Google Scholar] [CrossRef]
- Simon, G. Radiology in epidemiological studies and some therapeutic trials. Br. Med. J. 1966, 2, 491–494. [Google Scholar] [CrossRef] [Green Version]
- Krouwels, F.H.; Nocker, R.E.; Snoek, M.; Lutter, R.; van der Zee, J.S.; Weller, F.R.; Jansen, H.M.; Out, T.A. Immunocytochemical and flow cytofluorimetric detection of intracellular IL-4, IL-5 and IFN-gamma: Applications using blood- and airway-derived cells. J. Immunol. Methods 1997, 203, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Barnes, P.F.; Lu, S.; Abrams, J.S.; Wang, E.; Yamamura, M.; Modlin, R.L. Cytokine production at the site of disease in human tuberculosis. Infect. Immun. 1993, 61, 3482–3489. [Google Scholar] [CrossRef] [Green Version]
- Sohaskey, C.D.; Wayne, L.G. Role of narK2X and narGHJI in hypoxic upregulation of nitrate reduction by Mycobacterium tuberculosis. J. Bacteriol. 2003, 185, 7247–7256. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.; Xing, Y.; Magliozzo, R.S.; Bloom, B.R. Killing of virulent Mycobacterium tuberculosis by reactive nitrogen intermediates produced by activated murine macrophages. J. Exp. Med. 1992, 175, 1111–1122. [Google Scholar] [CrossRef]
- Ghadimi, D.; de Vrese, M.; Heller, K.J.; Schrezenmeir, J. Lactic acid bacteria enhance autophagic ability of mononuclear phagocytes by increasing Th1 autophagy-promoting cytokine (IFN-gamma) and nitric oxide (NO) levels and reducing Th2 autophagy-restraining cytokines (IL-4 and IL-13) in response to Mycobacterium tuberculosis antigen. Int. Immunopharmacol. 2010, 10, 694–706. [Google Scholar] [PubMed]
- Deepak, P.; Kumar, S.; Acharya, P. Interleukin-13-induced type II polarization of inflammatory macrophages is mediated through suppression of nuclear factor-κB and preservation of IκBα in a T cell lymphoma. Clin. Exp. Immunol. 2007, 149, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Sozzani, P.; Cambon, C.; Vita, N.; Séguélas, M.H.; Caput, D.; Ferrara, P.; Pipy, B. Interleukin-13 inhibits protein kinase C-triggered respiratory burst in human monocytes. Role of calcium and cyclic AMP. J. Biol. Chem. 1995, 270, 5084–5088. [Google Scholar] [CrossRef] [Green Version]
- Newport, M.J.; Huxley, C.M.; Huston, S.; Hawrylowicz, C.M.; Oostra, B.A.; Williamson, R.; Levin, M. A mutation in the interferon-gamma-receptor gene and susceptibility to mycobacterial infection. N. Engl. J. Med. 1996, 335, 1941–1949. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.L.; Chan, J.; Triebold, K.J.; Dalton, D.K.; Stewart, T.A.; Bloom, B.R. An essential role for interferon gamma in resistance to Mycobacterium tuberculosis infection. J. Exp. Med. 1993, 178, 2249–2254. [Google Scholar] [CrossRef] [Green Version]
- Shalaby, M.R.; Waage, A.; Espevik, T. Cytokine regulation of interleukin 6 production by human endothelial cells. Cell Immunol. 1989, 121, 372–382. [Google Scholar] [CrossRef]
- Barry, S.; Breen, R.; Lipman, M.; Johnson, M.; Janossy, G. Impaired antigen-specific CD4(+) T lymphocyte responses in cavitary tuberculosis. Tuberculosis (Edinb) 2009, 89, 48–53. [Google Scholar] [CrossRef]
- Zhu, J.; Yamane, H.; Cote-Sierra, J.; Guo, L.; Paul, W.E. GATA-3 promotes Th2 responses through three different mechanisms: Induction of Th2 cytokine production, selective growth of Th2 cells and inhibition of Th1 cell-specific factors. Cell Res. 2006, 16, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Pooran, A.; Davids, M.; Nel, A.; Shoko, A.; Blackburn, J.; Dheda, K. IL-4 subverts mycobacterial containment in Mycobacterium tuberculosis-infected human macrophages. Eur. Respir. J. 2019, 54, 1802242. [Google Scholar] [CrossRef]
- Kutlu, A.; Bozkurt, B.; Ciftci, F.; Bozkanat, E.; Gorur, R.; Sonmez, G.; Ozturk, S.; Taskapan, O.; Kartaloglu, Z. Does atopy affect the radiological extent of pulmonary tuberculosis? Allergol. Immunopathol. (Madr) 2008, 36, 187–190. [Google Scholar] [CrossRef]
- Kuo, H.C.; Liu, C.A.; Ou, C.Y.; Hsu, T.Y.; Yang, K.D. Correlation between atopy and tuberculin/Candida skin test reactivity in a bacillus Calmette- Gue’rin-vaccinated cohort. Allergy 2010, 65, 1625–1626. [Google Scholar] [CrossRef]
- Shirakawa, T.; Enomoto, T.; Shimazu, S.; Hopkin, J.M. The inverse association between tuberculin responses and atopic disorder. Science 1997, 275, 77–79. [Google Scholar] [CrossRef]
- George, P.J.; Anuradha, R.; Kumar, N.P.; Sridhar, R.; Banurekha, V.V.; Nutman, T.B.; Babu, S. Helminth infections coincident with active pulmonary tuberculosis inhibit mono- and multifunctional CD4+ and CD8+ T cell responses in a process dependent on IL-10. PLoS Pathog. 2014, 10, e1004375. [Google Scholar] [CrossRef] [PubMed]
- Sartono, E.; Kruize, Y.C.; Kurniawan, A.; van der Meide, P.H.; Partono, F.; Maizels, R.M.; Yazdanbakhsh, M. Elevated cellular immune responses and interferon-gamma release after long-term diethylcarbamazine treatment of patients with human lymphatic filariasis. J. Infect. Dis. 1995, 171, 1683–1687. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Lee, S.H.; Yim, J.J. Serial interferon-gamma release assays after chemoprophylaxis in a tuberculosis outbreak cohort. Infection 2012, 40, 431–435. [Google Scholar] [CrossRef]
- Li, G.; Yang, F.; He, X.; Liu, Z.; Pi, J.; Zhu, Y.; Ke, X.; Liu, S.; Ou, M.; Guo, H.; et al. Anti-tuberculosis (TB) chemotherapy dynamically rescues Th1 and CD8+ T effector levels in Han Chinese pulmonary TB patients. Microbes Infect. 2020, 22, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Zaibi, H.; Fekih, L.; Akroutm, I.; Tangour, E.; Hassene, H.; Greb, D.; Fenniche, S.; Abdelghaffar, H.B.; Belahbib, D.; Megdiche, M.L. What about asthma in tuberculosis patients. Eur. Respir. J. 2011, 38, 2610. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KERRYPNX | Control Subjects (n = 7) | TB Patients | ||
|---|---|---|---|---|
| Total (n = 19) | Non-Atopy (n = 12) | Atopy (n = 7) | ||
| Age, years | 45.7 ± 5.4 | 50.5 ± 3.7 | 52.8 ± 5.0 | 46.4 ± 5.6 |
| Male/female | 3/4 | 12/7 | 8/4 | 4/3 |
| IgE, IU/mL | 56.1 ± 15.4 | 78.1 ± 13.5 | 38.1 ± 5.6 ¶ | 146.6 ± 11.7 ** |
| Bacterial load on sputum | N/A | 3.5 ± 1.8 | 4.6 ± 1.1 | 1.7 ± 1.0 |
| Bronchoalveolar lavage | ||||
| Cellularity, 105 cells/mL | 1.6 ± 0.2 | 10.6 ± 3.0 ** | 12.9 ± 4.5 ¶,** | 6.6 ± 2.7 |
| Recovery rate, % | 50.1 ± 7.1 | 44.4 ± 3.1 | 43.2 ± 4.1 | 46.5 ± 5.1 |
| Viability, % | 95.4 ± 1.3 | 91.1 ±1.5 | 92.3 ± 1.8 | 88.9 ± 2.7 |
| AMs, % | 97.1 ± 0.6 | 83.8 ± 2.3 ** | 80.3 ± 4.0 ** | 88.6 ± 2.3 * |
| Lymphocytes, % | 1.7 ± 0.4 | 7.3 ± 1.3 * | 6.8 ± 1.6 * | 8.2 ± 2.3 * |
| Neutrophils, % | 0.8 ± 0.3 | 9.8 ± 2.9 ** | 14.2 ± 4.2 §,** | 2.2 ± 1.0 * |
| Eosinophils, % | 0.3 ± 0.3 | 0.8 ± 0.2 | 0.8 ± 0.3 | 0.9 ± 0.4 |
| Control Subjects (n = 5) | TB Patients (n = 9) | |||
|---|---|---|---|---|
| Unstimulated | Stimulated | Unstimulated | Stimulated | |
| IFN-γ (ng/mL) | 48 ± 32 | 26 ± 13 | 377 ± 116 ¶¶ | 4649 ± 1512 *,¶¶¶ |
| Nitrite (μM) | 2995 ± 918 | 1731 ± 184 | 3036 ± 336 | 4669 ± 610 *,¶¶¶ |
| IL-4 (ng/mL) | Undetectable | Undetectable | Undetectable | Undetectable |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo, C.-Y.; Huang, Y.-C.; Huang, H.-Y.; Chung, F.-T.; Lin, C.-W.; Chung, K.F.; Wang, C.-H. Increased Th1 Cells with Disease Resolution of Active Pulmonary Tuberculosis in Non-Atopic Patients. Biomedicines 2021, 9, 724. https://doi.org/10.3390/biomedicines9070724
Lo C-Y, Huang Y-C, Huang H-Y, Chung F-T, Lin C-W, Chung KF, Wang C-H. Increased Th1 Cells with Disease Resolution of Active Pulmonary Tuberculosis in Non-Atopic Patients. Biomedicines. 2021; 9(7):724. https://doi.org/10.3390/biomedicines9070724
Chicago/Turabian StyleLo, Chun-Yu, Yu-Chen Huang, Hung-Yu Huang, Fu-Tsai Chung, Chang-Wei Lin, Kian Fan Chung, and Chun-Hua Wang. 2021. "Increased Th1 Cells with Disease Resolution of Active Pulmonary Tuberculosis in Non-Atopic Patients" Biomedicines 9, no. 7: 724. https://doi.org/10.3390/biomedicines9070724
APA StyleLo, C.-Y., Huang, Y.-C., Huang, H.-Y., Chung, F.-T., Lin, C.-W., Chung, K. F., & Wang, C.-H. (2021). Increased Th1 Cells with Disease Resolution of Active Pulmonary Tuberculosis in Non-Atopic Patients. Biomedicines, 9(7), 724. https://doi.org/10.3390/biomedicines9070724