
Weighted Gene Co-Expression Network Analysis Reveals Key Genes and Potential Drugs in Abdominal Aortic Aneurysm

 ,
, 
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Sources and Preprocessing
2.2. Construction of WGCNA
2.3. Selection of Key Modules Corresponding to Sample Traits
2.4. Functional Enrichment Analysis of the Key Modules
2.5. Identification of Hub Genes in the Key Modules
2.6. Hub Genes Validation and Key Genes Selection
2.7. Predication of Drug–Gene Interaction
2.8. Statistical Analysis
3. Results
3.1. Construction of Weighted Gene Co-Expression Network
3.2. Construction of Module-Trait Relationships and Detection of Key Modules
3.3. Functional Enrichment Analysis of Genes in the Module
3.4. Identification of Hub Genes in the Key Modules
3.5. Hub Genes Validation and Key Genes Selection
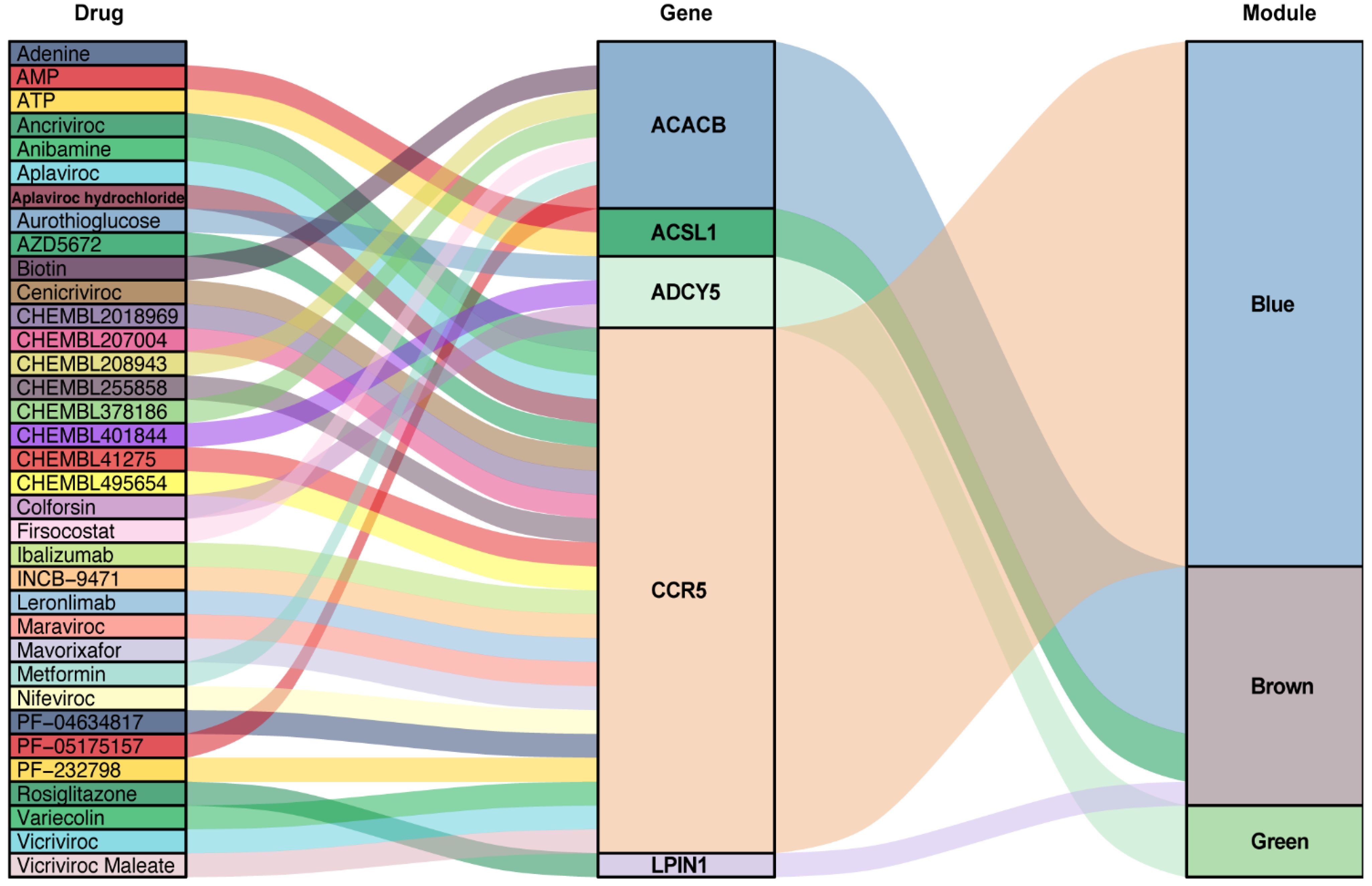
3.6. Predication of Drug-Gene Interaction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Golledge, J.; Muller, J.; Daugherty, A.; Norman, P. Abdominal Aortic Aneurysm: Pathogenesis and Implications for Management. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2605–2613. [Google Scholar] [CrossRef]
- Lucas, R.; Banerjee, A.; Barquera, S.; Blyth, F.M.; Cowie, B.C.; Ding, E.L.; Gunnell, D.; Lan, Q.; J, M.J.; Patton, G.C.; et al. Global, regional, and national age–sex specific all-cause and cause-specific mortality for 240 causes of death, 1990–2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet 2015, 385, 117–171. [Google Scholar] [CrossRef]
- Sampson, U.K.A.; Norman, P.E.; Fowkes, F.G.R.; Aboyans, V.; Song, Y.; Harrell, F.E.; Forouzanfar, M.H.; Naghavi, M.; Denenberg, J.O.; McDermott, M.M.; et al. Global and Regional Burden of Aortic Dissection and Aneurysms: Mortality Trends in 21 World Regions, 1990 to 2010. Glob. Heart 2014, 9, 171–180.e10. [Google Scholar] [CrossRef]
- Toghill, B.J.; Saratzis, A.; Bown, M.J. Abdominal Aortic Aneurysm—an Independent Disease to Atherosclerosis? Cardiovasc. Pathol. 2017, 27, 71–75. [Google Scholar] [CrossRef]
- Chaikof, E.L.; Dalman, R.L.; Eskandari, M.K.; Jackson, B.M.; Lee, W.A.; Mansour, M.A.; Mastracci, T.M.; Mell, M.; Murad, M.H.; Nguyen, L.L.; et al. The Society for Vascular Surgery Practice Guidelines on the Care of Patients with an Abdominal Aortic Aneurysm. J. Vasc. Surg. 2018, 67, 2–77.e2. [Google Scholar] [CrossRef] [PubMed]
- Wanhainen, A.; Verzini, F.; Van Herzeele, I.; Allaire, E.; Bown, M.; Cohnert, T.; Dick, F.; van Herwaarden, J.; Karkos, C.; Koelemay, M.; et al. Editor’s Choice - European Society for Vascular Surgery (ESVS) 2019 Clinical Practice Guidelines on the Management of Abdominal Aorto-Iliac Artery Aneurysms. Eur. J. Vasc. Endovasc. Surg. 2019, 57, 8–93. [Google Scholar] [CrossRef]
- Kokje, V.B.C.; Hamming, J.F.; Lindeman, J.H.N. Pharmaceutical Management of Small Abdominal Aortic Aneurysms: A Systematic Review of the Clinical Evidence. J. Vasc. Surg. 2015, 62, 1680. [Google Scholar] [CrossRef]
- Rughani, G.; Robertson, L.; Clarke, M. Medical Treatment for Small Abdominal Aortic Aneurysms. Cochrane Datab. Syst. Rev. 2012, CD009536. [Google Scholar] [CrossRef]
- Daugherty, A.; Cassis, L.A. Mouse Models of Abdominal Aortic Aneurysms. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Golledge, J. Abdominal aortic aneurysm: update on pathogenesis and medical treatments. Nat. Rev. Cardiol. 2019, 16, 225–242. [Google Scholar] [CrossRef]
- Moran, C.S.; Biros, E.; Krishna, S.M.; Wang, Y.; Tikellis, C.; Morton, S.K.; Moxon, J.V.; Cooper, M.E.; Norman, P.E.; Burrell, L.M.; et al. Resveratrol Inhibits Growth of Experimental Abdominal Aortic Aneurysm Associated With Upregulation of Angiotensin-Converting Enzyme 2. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 2195–2203. [Google Scholar] [CrossRef] [PubMed]
- Trachet, B.; Piersigilli, A.; Fraga-Silva, R.A.; Aslanidou, L.; Sordet-Dessimoz, J.; Astolfo, A.; Stampanoni, M.F.M.; Segers, P.; Stergiopulos, N. Ascending Aortic Aneurysm in Angiotensin II-Infused Mice: Formation, Progression, and the Role of Focal Dissections. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 673–681. [Google Scholar] [CrossRef]
- Rateri, D.L.; Howatt, D.A.; Moorleghen, J.J.; Charnigo, R.; Cassis, L.A.; Daugherty, A. Prolonged Infusion of Angiotensin II in apoE−/− Mice Promotes Macrophage Recruitment with Continued Expansion of Abdominal Aortic Aneurysm. Am. J. Pathol. 2011, 179, 1542–1548. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, C.; Zhang, M.; Liang, B.; Zhu, H.; Lee, J.; Viollet, B.; Xia, L.; Zhang, Y.; Zou, M.-H. Activation of AMP-Activated Protein Kinase A2 by Nicotine Instigates Formation of Abdominal Aortic Aneurysms in Mice in Vivo. Nat. Med. 2012, 18, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Daugherty, A.; Manning, M.W.; Cassis, L.A. Angiotensin II promotes atherosclerotic lesions and aneurysms in apolipoprotein E–deficient mice. J. Clin. Investig. 2000, 105, 1605–1612. [Google Scholar] [CrossRef]
- Piacentini, L.; Chiesa, M.; Colombo, G.I. Gene Regulatory Network Analysis of Perivascular Adipose Tissue of Abdominal Aortic Aneurysm Identifies Master Regulators of Key Pathogenetic Pathways. Biomedicines 2020, 8, 288. [Google Scholar] [CrossRef]
- Coscas, R.; Dupont, S.; Mussot, S.; Louedec, L.; Etienne, H.; Morvan, M.; Chiocchia, G.; Massy, Z.; Jacob, M.-P.; Michel, J.-B. Exploring antibody-dependent adaptive immunity against aortic extracellular matrix components in experimental aortic aneurysms. J. Vasc. Surg. 2018, 68, 60S–71S.e3. [Google Scholar] [CrossRef]
- Liljeqvist, M.L.; Hultgren, R.; Bergman, O.; Villard, C.; Kronqvist, M.; Eriksson, P.; Roy, J. Tunica-Specific Transcriptome of Abdominal Aortic Aneurysm and the Effect of Intraluminal Thrombus, Smoking, and Diameter Growth Rate. Arter. Thromb. Vasc. Biol. 2020, 40, 2700–2713. [Google Scholar] [CrossRef]
- Piacentini, L.; Werba, J.P.; Bono, E.; Saccu, C.; Tremoli, E.; Spirito, R.; Colombo, G.I. Genome-Wide Expression Profiling Unveils Autoimmune Response Signatures in the Perivascular Adipose Tissue of Abdominal Aortic Aneurysm. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 237–249. [Google Scholar] [CrossRef]
- Dale, M.A.; Ruhlman, M.K.; Baxter, B.T. Inflammatory Cell Phenotypes in AAAs. Arter. Thromb. Vasc. Biol. 2015, 35, 1746–1755. [Google Scholar] [CrossRef]
- Kuivaniemi, H.; Ryer, E.J.; Elmore, J.R.; Tromp, G. Understanding the pathogenesis of abdominal aortic aneurysms. Expert Rev. Cardiovasc. Ther. 2015, 13, 975–987. [Google Scholar] [CrossRef]
- Raffort, J.; Lareyre, F.; Clément, M.; Hassen-Khodja, F.L.R.; Chinetti, J.R.F.L.R.H.-K.G.; Mallat, J.R.F.L.M.C.Z. Monocytes and macrophages in abdominal aortic aneurysm. Nat. Rev. Cardiol. 2017, 14, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, N.L. Understanding Abdominal Aortic Aneurysm. N. Engl. J. Med. 2009, 361, 1114–1116. [Google Scholar] [CrossRef]
- Pahl, M.C.; Erdman, R.; Kuivaniemi, H.; Lillvis, J.H.; Elmore, J.R.; Tromp, G. Transcriptional (ChIP-Chip) Analysis of ELF1, ETS2, RUNX1 and STAT5 in Human Abdominal Aortic Aneurysm. Int. J. Mol. Sci. 2015, 16, 11229–11258. [Google Scholar] [CrossRef]
- Rush, C.; Nyara, M.; Moxon, J.V.; Trollope, A.; Cullen, B.; Golledge, J. Whole genome expression analysis within the angiotensin II-apolipoprotein E deficient mouse model of abdominal aortic aneurysm. BMC Genom. 2009, 10, 298. [Google Scholar] [CrossRef]
- Spin, J.M.; Hsu, M.; Azuma, J.; Tedesco, M.M.; Deng, A.; Dyer, J.S.; Maegdefessel, L.; Dalman, R.L.; Tsao, P.S. Transcriptional profiling and network analysis of the murine angiotensin II-induced abdominal aortic aneurysm. Physiol. Genom. 2011, 43, 993–1003. [Google Scholar] [CrossRef]
- Zhang, B.; Horvath, S. A General Framework for Weighted Gene Co-Expression Network Analysis. Stat. Appl. Genet. Mol. Biol. 2005, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Zhang, N.; Yan, L.; Lian, Z.; Wang, J.; Lv, F.; Wang, Y.; Cao, X. Weighted gene co-expression network analysis in identification of key genes and networks for ischemic-reperfusion remodeling myocardium. Mol. Med. Rep. 2018, 18, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Nagenborg, J.; Jin, H.; Brennan, E.; Goossens, P.; Donners, M.; Biessen, E. Re-Programming Atherosclerotic Plaque Macrophages Towards An Anti-Atherogenic Phenotype. Atheroscler. 2019, 287, e79. [Google Scholar] [CrossRef]
- Wang, C.-H.; Shi, H.-H.; Chen, L.-H.; Li, X.-L.; Cao, G.-L.; Hu, X.-F. Identification of Key lncRNAs Associated With Atherosclerosis Progression Based on Public Datasets. Front. Genet. 2019, 10, 123. [Google Scholar] [CrossRef]
- Biros, E.; Gäbel, G.; Moran, C.S.; Schreurs, C.; Lindeman, J.H.N.; Walker, P.J.; Nataatmadja, M.; West, M.; Holdt, L.M.; Hinterseher, I.; et al. Differential gene expression in human abdominal aortic aneurysm and aortic occlusive disease. Oncotarget 2015, 6, 12984–12996. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucl. Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Kassambara, A. Ggcorrplot: Visualization of a Correlation Matrix Using “Ggplot2”. 2019. Available online: https://github.com/kassambara/ggcorrplot (accessed on 13 September 2020).
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS: A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: quality-controlled protein–protein association networks, made broadly accessible. Nucl. Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef]
- Cotto, K.C.; Wagner, A.H.; Feng, Y.-Y.; Kiwala, S.; Coffman, A.C.; Spies, G.; Wollam, A.; Spies, N.C.; Griffith, O.L.; Griffith, M. DGIdb 3.0: a redesign and expansion of the drug–gene interaction database. Nucl. Acids Res. 2018, 46, D1068–D1073. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.L.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H.; Dunnington, D. RStudio Ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics. 2020. Available online: https://ggplot2.tidyverse.org/ (accessed on 10 September 2020).
- Brunson, J.C. Ggalluvial: Alluvial Plots in “Ggplot2”. 2019. Available online: https://cran.r-project.org/web/packages/ggalluvial/index.html (accessed on 16 September 2020).
- Butt, H.; Sylvius, N.; Salem, M.; Wild, J.; Dattani, N.; Sayers, R.; Bown, M. Microarray-based Gene Expression Profiling of Abdominal Aortic Aneurysm. Eur. J. Vasc. Endovasc. Surg. 2016, 52, 47–55. [Google Scholar] [CrossRef]
- Cornuz, J.; Pinto, C.S.; Tevaearai, H.; Egger, M. Risk factors for asymptomatic abdominal aortic aneurysm: systematic review and meta-analysis of population-based screening studies. Eur. J. Public Heal. 2004, 14, 343–349. [Google Scholar] [CrossRef]
- Lin, S.; Han, X.; Bi, Y.; Ju, S.; Gu, L. Fluid-Structure Interaction in Abdominal Aortic Aneurysm: Effect of Modeling Techniques. BioMed Res. Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef]
- Sheidaei, A.; Hunley, S.; Zeinali-Davarani, S.; Raguin, L.; Baek, S. Simulation of abdominal aortic aneurysm growth with updating hemodynamic loads using a realistic geometry. Med Eng. Phys. 2011, 33, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.-I.; Fukumoto, Y.; Nochioka, K.; Minami, T.; Kudo, S.; Shiba, N.; Takai, Y.; Williams, C.L.; Liao, J.K.; Shimokawa, H. Statins Exert the Pleiotropic Effects Through Small GTP-Binding Protein Dissociation Stimulator Upregulation With a Resultant Rac1 Degradation. Arter. Thromb. Vasc. Biol. 2013, 33, 1591–1600. [Google Scholar] [CrossRef]
- Yu, D.; Makkar, G.; Strickland, D.K.; Blanpied, T.A.; Stumpo, D.J.; Blackshear, P.J.; Sarkar, R.; Monahan, T.S. Myristoylated Alanine-Rich Protein Kinase Substrate (MARCKS) Regulates Small GTPase Rac1 and Cdc42 Activity and Is a Critical Mediator of Vascular Smooth Muscle Cell Migration in Intimal Hyperplasia Formation. J. Am. Heart Assoc. 2015, 4, e002255. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, M.; Hellström, A.; Henriksson, A.E. Abdominal Aortic Aneurysm and the Association with Serum Levels of Homocysteine, Vitamins B6, B12 and Folate. Am. J. Cardiovasc. Dis. 2012, 2, 318–322. [Google Scholar]
- Wiernicki, I.; Parafiniuk, M.; Kolasa-Wołosiuk, A.; Gutowska, I.; Kazimierczak, A.; Clark, J.; Baranowska-Bosiacka, I.; Szumilowicz, P.; Gutowski, P. Relationship between aortic wall oxidative stress/proteolytic enzyme expression and intraluminal thrombus thickness indicates a novel pathomechanism in the progression of human abdominal aortic aneurysm. FASEB J. 2018, 33, 885–895. [Google Scholar] [CrossRef]
- Berger, E.A.; Murphy, P.M.; Farber, J.M. CHEMOKINE RECEPTORS AS HIV-1 CORECEPTORS: Roles in Viral Entry, Tropism, and Disease. Annu. Rev. Immunol. 1999, 17, 657–700. [Google Scholar] [CrossRef]
- Kohlmeier, J.E.; Reiley, W.W.; Perona-Wright, G.; Freeman, M.L.; Yager, E.J.; Connor, L.M.; Brincks, E.L.; Cookenham, T.; Roberts, A.D.; Burkum, C.E.; et al. Inflammatory chemokine receptors regulate CD8+ T cell contraction and memory generation following infection. J. Exp. Med. 2011, 208, 1621–1634. [Google Scholar] [CrossRef]
- Ghilardi, G.; Biondi, M.L.; Battaglioli, L.; Zambon, A.; Guagnellini, E.; Scorza, R. Genetic risk factor characterizes abdominal aortic aneurysm from arterial occlusive disease in human beings: CCR5 Δ32 deletion. J. Vasc. Surg. 2004, 40, 995–1000. [Google Scholar] [CrossRef]
- Phillips, E.H.; Lorch, A.H.; Durkes, A.C.; Goergen, C.J. Early pathological characterization of murine dissecting abdominal aortic aneurysms. APL Bioeng. 2018, 2, 046106. [Google Scholar] [CrossRef]
- Grarup, N.; Moltke, I.; Andersen, M.K.; Dalby, M.; Vitting-Seerup, K.; Kern, T.; Mahendran, Y.; Jørsboe, E.; Larsen, C.V.L.; Dahl-Petersen, I.K.; et al. Loss-of-function variants in ADCY3 increase risk of obesity and type 2 diabetes. Nat. Genet. 2018, 50, 172–174. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gong, Y.W.; Zhou, X.Q.; Xu, H.X.; Yang, L.; Wu, Y.Y. [Association between single nucleotide polymorphism of adenylyl cyclase 3 and essential hypertension]. Zhonghua Xin Xue Guan Bing Za Zhi 2016, 44, 594–599. [Google Scholar]
- Thompson, A.; Drenos, F.; Hafez, H.M.; E Humphries, S. Candidate Gene Association Studies in Abdominal Aortic Aneurysm Disease: A Review and Meta-Analysis. Eur. J. Vasc. Endovasc. Surg. 2008, 35, 19–30. [Google Scholar] [CrossRef][Green Version]
- Reue, K.; Xu, P.; Wang, X.-P.; Slavin, B.G. Adipose tissue deficiency, glucose intolerance, and increased atherosclerosis result from mutation in the mouse fatty liver dystrophy (fld) gene. J. Lipid Res. 2000, 41, 1067–1076. [Google Scholar] [CrossRef]
- Jama, A.; Huang, D.; A Alshudukhi, A.A.; Chrast, R.; Ren, H. Lipin1 is required for skeletal muscle development by regulating MEF2c and MyoD expression. J. Physiol. 2019, 597, 889–901. [Google Scholar] [CrossRef]
- Koh, J.-H.; Kim, K.-H.; Park, S.-Y.; Kim, Y.-W.; Kim, J.-Y. PPARδ Attenuates Alcohol-Mediated Insulin Resistance by Enhancing Fatty Acid-Induced Mitochondrial Uncoupling and Antioxidant Defense in Skeletal Muscle. Front. Physiol. 2020, 11, 749. [Google Scholar] [CrossRef]
- Silvestri, E.; Senese, R.; De Matteis, R.; Cioffi, F.; Moreno, M.; Lanni, A.; Gentile, A.; Busiello, R.A.; Salzano, A.M.; Scaloni, A.; et al. Absence of uncoupling protein 3 at thermoneutrality influences brown adipose tissue mitochondrial functionality in mice. FASEB J. 2020, 34, 15146–15163. [Google Scholar] [CrossRef]
- Stierwalt, H.D.; Ehrlicher, S.E.; Robinson, M.M.; Newsom, S.A. Skeletal Muscle ACSL Isoforms Relate to Measures of Fat Metabolism in Humans. Med. Sci. Sports Exerc. 2020. [Google Scholar] [CrossRef]
- Alkhouri, N.; Lawitz, E.; Noureddin, M.; DeFronzo, R.; Shulman, G.I. GS-0976 (Firsocostat): an investigational liver-directed acetyl-CoA carboxylase (ACC) inhibitor for the treatment of non-alcoholic steatohepatitis (NASH). Expert Opin. Investig. Drugs 2020, 29, 135–141. [Google Scholar] [CrossRef]
- Huard, K.; Smith, A.C.; Cappon, G.D.; Dow, R.L.; Edmonds, D.J.; El-Kattan, A.; Esler, W.P.; Fernando, D.P.; Griffith, D.A.; Kalgutkar, A.S.; et al. Optimizing the Benefit/Risk of Acetyl-CoA Carboxylase Inhibitors through Liver Targeting. J. Med. Chem. 2020, 63, 10879–10896. [Google Scholar] [CrossRef]
- Chaudhury, A.; Duvoor, C.; Dendi, V.S.R.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T.; Kuriakose, K.; et al. Clinical Review of Antidiabetic Drugs: Implications for Type 2 Diabetes Mellitus Management. Front. Endocrinol. 2017, 8, 6. [Google Scholar] [CrossRef]
- Isoda, K.; Young, J.L.; Zirlik, A.; Macfarlane, L.A.; Tsuboi, N.; Gerdes, N.; Schoönbeck, U.; Libby, P. Metformin Inhibits Proinflammatory Responses and Nuclear Factor-κB in Human Vascular Wall Cells. Arter. Thromb. Vasc. Biol. 2006, 26, 611–617. [Google Scholar] [CrossRef]
- Esfahanian, N.; Shakiba, Y.; Nikbin, B.; Soraya, H.; Maleki-Dizaji, N.; Ghazi-Khansari, M.; Garjani, A. Effect of metformin on the proliferation, migration, and MMP-2 and -9 expression of human umbilical vein endothelial cells. Mol. Med. Rep. 2012, 5, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.A.; Choi, H.C. Metformin inhibits inflammatory response via AMPK–PTEN pathway in vascular smooth muscle cells. Biochem. Biophys. Res. Commun. 2012, 425, 866–872. [Google Scholar] [CrossRef]
- Fujimura, N.; Xiong, J.; Kettler, E.B.; Xuan, H.; Glover, K.J.; Mell, M.W.; Xu, B.; Dalman, R.L. Metformin treatment status and abdominal aortic aneurysm disease progression. J. Vasc. Surg. 2016, 64, 46–54.e8. [Google Scholar] [CrossRef]
- Itoga, N.K.; Rothenberg, K.A.; Suarez, P.; Ho, T.-V.; Mell, M.W.; Xu, B.; Curtin, C.M.; Dalman, R.L. Metformin prescription status and abdominal aortic aneurysm disease progression in the U.S. veteran population. J. Vasc. Surg. 2019, 69, 710–716.e3. [Google Scholar] [CrossRef] [PubMed]
- Afonso, P.; Auclair, M.; Caron-Debarle, M.; Capeau, J. Impact of CCR5, integrase and protease inhibitors on human endothelial cell function, stress, inflammation and senescence. Antivir. Ther. 2017, 22, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Francisci, D.; Pirro, M.; Schiaroli, E.; Mannarino, M.R.; Cipriani, S.; Bianconi, V.; Alunno, A.; Bagaglia, F.; Bistoni, O.; Falcinelli, E.; et al. Maraviroc Intensification Modulates Atherosclerotic Progression in HIV-Suppressed Patients at High Cardiovascular Risk. A Randomized, Crossover Pilot Study. Open Forum Infect. Dis. 2019, 6, ofz112. [Google Scholar] [CrossRef]
- Yue, T.-L.; Chen, J.; Bao, W.; Narayanan, P.K.; Bril, A.; Jiang, W.; Lysko, P.G.; Gu, J.-L.; Boyce, R.; Zimmerman, D.M.; et al. In Vivo Myocardial Protection From Ischemia/Reperfusion Injury by the Peroxisome Proliferator–Activated Receptor-γ Agonist Rosiglitazone. Circulation 2001, 104, 2588–2594. [Google Scholar] [CrossRef]
- Wu, H.; Yang, Y.; Zheng, B.; Chen, K. Decreased PPAR-γ expression after internal carotid artery stenting is associated with vascular lesions induced by smooth muscle cell proliferation and systemic inflammation in a minipig model. Int. J. Clin. Exp. Pathol. 2017, 10, 7375–7383. [Google Scholar]
- Jones, A.; Deb, R.; Torsney, E.; Howe, F.; Dunkley, M.; Gnaneswaran, Y.; Gaze, D.; Nasr, H.; Loftus, I.M.; Thompson, M.M.; et al. Rosiglitazone Reduces the Development and Rupture of Experimental Aortic Aneurysms. Circulation 2009, 119, 3125–3132. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Catalog. | GSE Dataset | Organism | Sample Number * | PMID |
|---|---|---|---|---|
| Explore dataset | GSE17901 | Mouse | AAA day7: 7, AAA day14: 5, AAA day28: 6 | 21712436 |
| Validate dataset | GSE12591 | Mouse | Control: 6, AAA: 5 | 19580648 |
| GSE7084 | Human | Donor: 10, AAA: 9 | 17634102 | |
| GSE47472 | Human | Donor: 8, AAA: 14 | NA | |
| GSE57691 | Human | Donor: 10, AAA: 49 | NA |
| Catalog | Key Modules | ||
|---|---|---|---|
| Blue | Green | Brown | |
| Top 10 Gene | Ccr5 | Gnai1 | Lpl |
| Fpr2 | Adcy5 | Dgat2 | |
| Ccr2 | Adcy3 | Fasn | |
| Fpr1 | Rnase2a | Acacb | |
| P2ry12 | Cxcl13 | Lpin1 | |
| Hcar1 | Clca1 | Acsl1 | |
| Ppbp | Ear10 | Mogat1 | |
| Aif1 | Ear1 | Lep | |
| Sirpb1b | Npr1 | Ucp3 | |
| Clec4n | Ccl11 | Pdk4 | |
| Datasets | Key Modules | ||
|---|---|---|---|
| Blue | Green | Brown | |
| GSE7084 | CCR5, CCR2, FPR2, FPR1, AIF1 | GNAI1, RNASE2, NPR1 | NA |
| GSE47472 | CCR2, FPR2, PPBP | GNAI1, RNASE2, CLCA1, LYVE1 | LPIN1, UCP3 |
| GSE57691 | CCR2, FPR2, PPBP, CLEC6A, SIRPB1 | ADCY5, ADCY3, CXCL13, CLCA1, CCL11 | ACACB, LPIN1, ACSL1, LEP |
| Human | CCR5, CCR2, FPR2, PPBP, AIF1, CLEC6A, SIRPB1, FPR1 | GNAI1, RNASE2, NPR1, CLCA1, LYVE1, ADCY5, ADCY3, CXCL13, CCL11 | ACACB, LPIN1, ACSL1, LEP, UCP3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kan, K.-J.; Guo, F.; Zhu, L.; Pallavi, P.; Sigl, M.; Keese, M. Weighted Gene Co-Expression Network Analysis Reveals Key Genes and Potential Drugs in Abdominal Aortic Aneurysm. Biomedicines 2021, 9, 546. https://doi.org/10.3390/biomedicines9050546
Kan K-J, Guo F, Zhu L, Pallavi P, Sigl M, Keese M. Weighted Gene Co-Expression Network Analysis Reveals Key Genes and Potential Drugs in Abdominal Aortic Aneurysm. Biomedicines. 2021; 9(5):546. https://doi.org/10.3390/biomedicines9050546
Chicago/Turabian StyleKan, Ke-Jia, Feng Guo, Lei Zhu, Prama Pallavi, Martin Sigl, and Michael Keese. 2021. "Weighted Gene Co-Expression Network Analysis Reveals Key Genes and Potential Drugs in Abdominal Aortic Aneurysm" Biomedicines 9, no. 5: 546. https://doi.org/10.3390/biomedicines9050546
APA StyleKan, K.-J., Guo, F., Zhu, L., Pallavi, P., Sigl, M., & Keese, M. (2021). Weighted Gene Co-Expression Network Analysis Reveals Key Genes and Potential Drugs in Abdominal Aortic Aneurysm. Biomedicines, 9(5), 546. https://doi.org/10.3390/biomedicines9050546