
Ferulic Acid Induces Keratin 6α via Inhibition of Nuclear β-Catenin Accumulation and Activation of Nrf2 in Wound-Induced Inflammation

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Shilajit Preparation and Ferulic Acid
2.2. Cell Cultures
2.3. Real-Time Cell Analysis (RTCA) Measurement
2.4. Real-Time Quantitative PCR
2.5. Western Blotting
2.6. Animals
2.7. Wound Closure Test
2.8. Skin Explant Culture
2.9. Immunofluorescence
2.10. Microarray
2.11. LC-ESI-MS
2.12. Ethics Statement
2.13. Statistical Analysis
3. Results
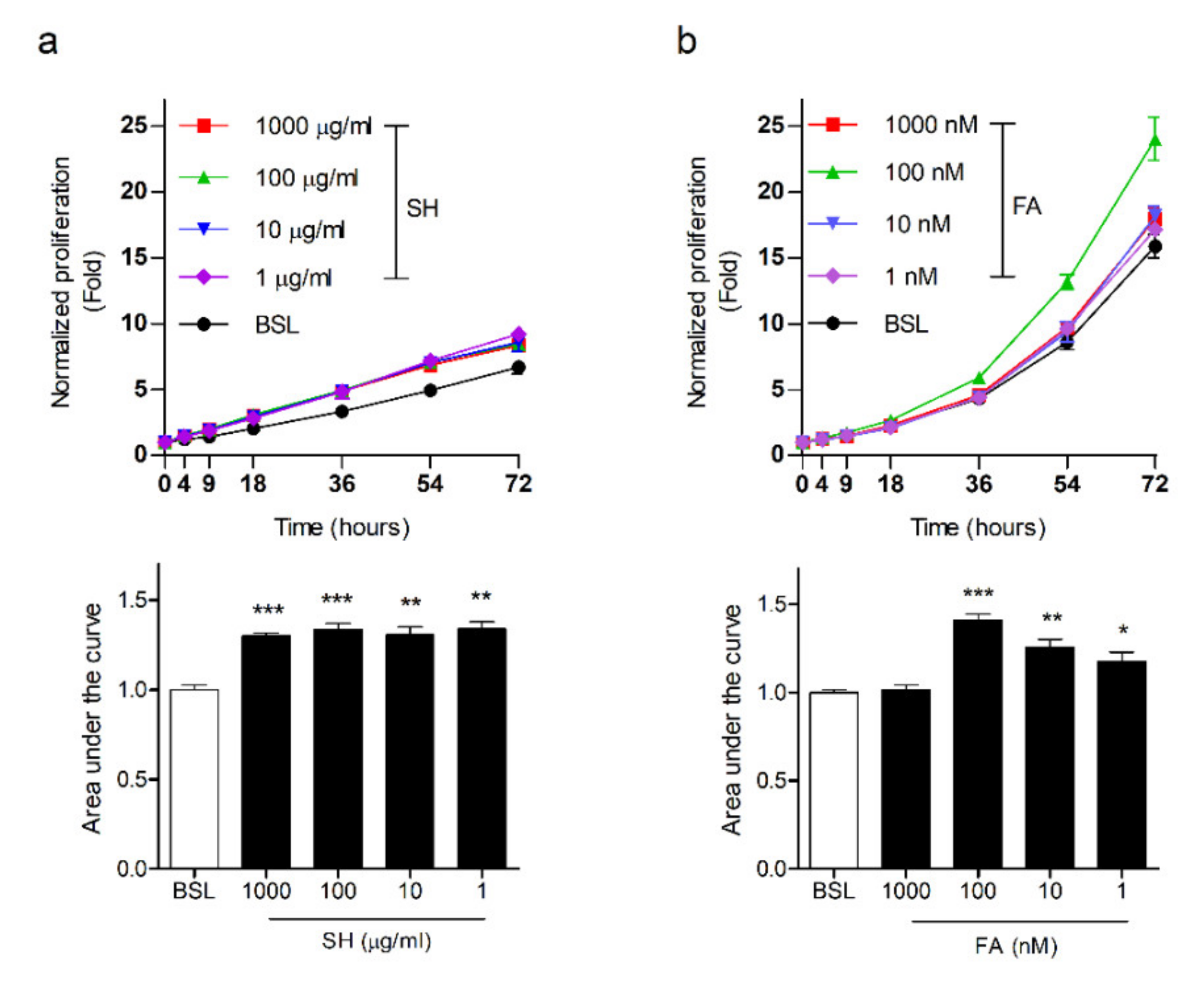
3.1. SH Induces Keratinocyte Migration and Its Active Component Is FA
3.2. SH and FA Induce Rapid Homeostatic Response of β-Catenin in Dermal Fibroblasts but Not in Epidermal Keratinocytes
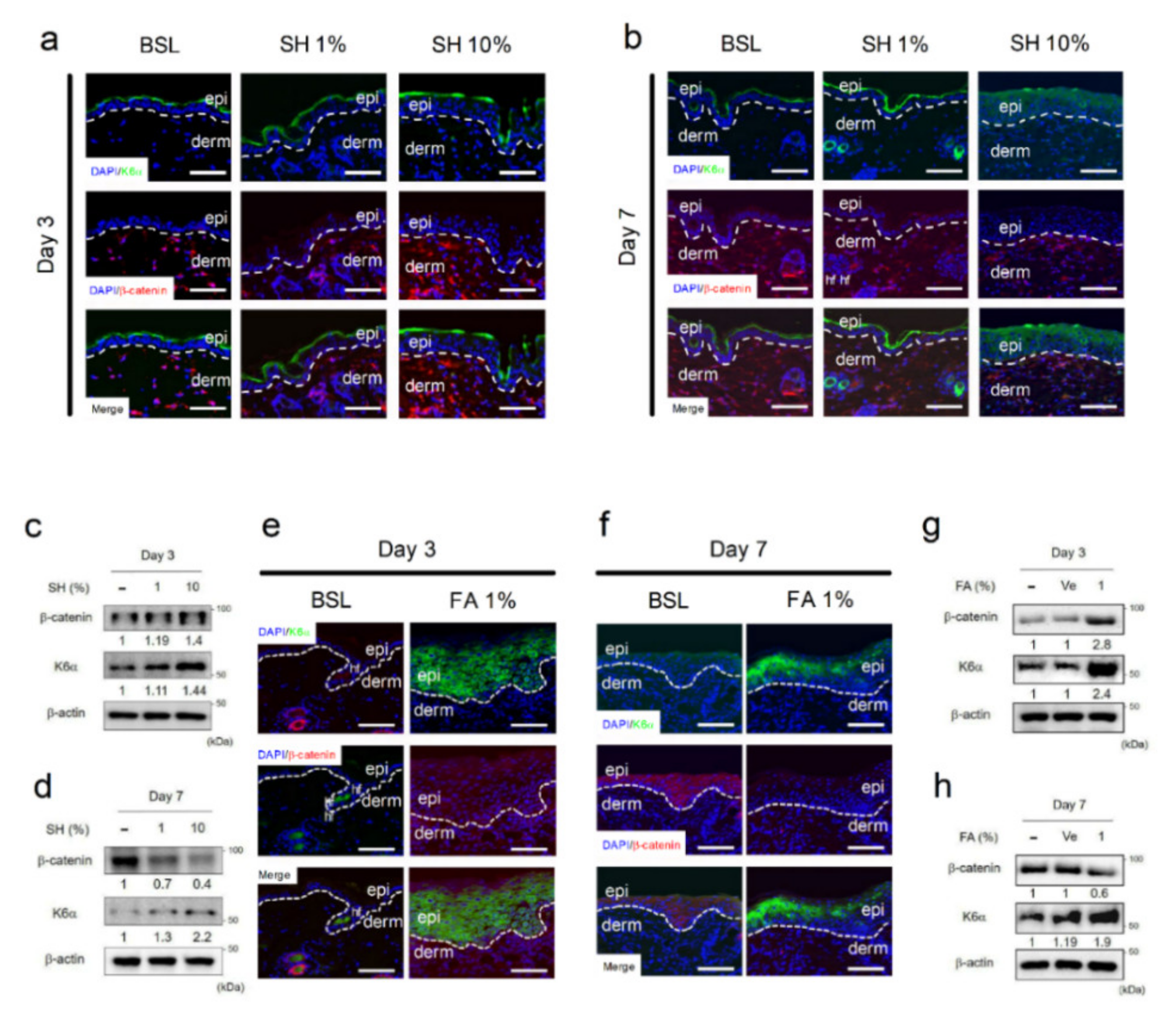
3.3. FA Suppresses Nuclear Accumulation of β-Catenin in Epidermal Keratinocytes
3.4. FA Ameliorated Wound-Induced Inflammation by Activation of NRF2
3.5. FA Induces Proliferation in Keratinocytes
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Demidova-Rice, T.N.; Hamblin, M.R.; Herman, I.M. Acute and Impaired Wound Healing: Pathophysiology and current methods for drug delivery, part 1: Normal and chronic wounds: Biology, causes, and approaches to care. Adv. Ski. Wound Care 2012, 25, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Elliot, S.; Wikramanayake, T.C.; Jozic, I.; Tomic-Canic, M. A Modeling Conundrum: Murine Models for Cutaneous Wound Healing. J. Investig. Dermatol. 2018, 138, 736–740. [Google Scholar] [CrossRef]
- Paladini, R.D.; Takahashi, K.; Bravo, N.S.; Coulombe, P. Onset of re-epithelialization after skin injury correlates with a reorganization of keratin filaments in wound edge keratinocytes: Defining a potential role for keratin 16. J. Cell Biol. 1996, 132, 381–397. [Google Scholar] [CrossRef]
- Takahashi, K.; Yan, B.; Yamanishi, K.; Imamura, S.; Coulombe, P.A. The Two Functional Keratin 6 Genes of Mouse Are Differentially Regulated and Evolved Independently from Their Human Orthologs. Genomics 1998, 53, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.; Coulombe, P.A. Loss of keratin 6 (K6) proteins reveals a function for intermediate filaments during wound repair. J. Cell Biol. 2003, 163, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.S.; Wei, Q.; Gurung, A.; Youn, A.; Bright, T.; Poon, R.; Whetstone, H.; Guha, A.; Alman, B.A. Beta-catenin regulates wound size and mediates the effect of TGF-beta in cutaneous healing. FASEB J. 2006, 20, 692–701. [Google Scholar] [CrossRef]
- Stojadinovic, O.; Brem, H.; Vouthounis, C.; Lee, B.; Fallon, J.; Stallcup, M.; Merchant, A.; Galiano, R.D.; Tomic-Canic, M. Molecular pathogenesis of chronic wounds: The role of beta-catenin and c-myc in the inhibition of epithelialization and wound healing. Am. J. Pathol. 2005, 167, 59–69. [Google Scholar] [CrossRef]
- Barker, N. The Canonical Wnt/β-Catenin Signalling Pathway. Methods Mol. Biol. 2008, 468, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.; Poon, R.; Yu, C.; Khoury, M.; Shenker, R.; Fish, J.; Alman, B. Prolonged β-catenin stabilization and tcf-dependent transcriptional activation in hyperplastic cutaneous wounds. Lab. Investig. 2005, 85, 416–425. [Google Scholar] [CrossRef]
- Veltri, A.; Lang, C.; Lien, W.-H. Concise Review: Wnt Signaling Pathways in Skin Development and Epidermal Stem Cells. Stem Cells 2018, 36, 22–35. [Google Scholar] [CrossRef]
- Cheon, S.S.; Cheah, A.Y.L.; Turley, S.; Nadesan, P.; Poon, R.; Clevers, H.; Alman, B.A. β-Catenin stabilization dysregulates mesenchymal cell proliferation, motility, and invasiveness and causes aggressive fibromatosis and hyperplastic cutaneous wounds. Proc. Natl. Acad. Sci. USA 2002, 99, 6973–6978. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Xie, R.; Shu, B.; Landay, A.L.; Wei, C.; Reiser, J.; Spagnoli, A.; Torquati, A.; Forsyth, C.B.; Keshavarzian, A.; et al. Wnt signaling in bone, kidney, intestine, and adipose tissue and interorgan interaction in aging. Ann. N. Y. Acad. Sci. 2018, 1442, 48–60. [Google Scholar] [CrossRef]
- Takeichi, M. Historical review of the discovery of cadherin, in memory of Tokindo Okada. Dev. Growth Differ. 2017, 60, 3–13. [Google Scholar] [CrossRef]
- Lee, S.-H.; Zahoor, M.; Hwang, J.-K.; Min, S.; Choi, K.-Y. Valproic Acid Induces Cutaneous Wound Healing In Vivo and Enhances Keratinocyte Motility. PLoS ONE 2012, 7, e48791. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.J.; Choi, S.-J.; Yoon, S.-Y.; Lee, J.Y.; Park, W.-S.; Park, P.-J.; Kim, K.H.; Eun, H.C.; Kwon, O. Valproic acid promotes human hair growth in in vitro culture model. J. Dermatol. Sci. 2013, 72, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Yoon, J.; Shin, S.H.; Zahoor, M.; Kim, H.J.; Park, P.J.; Park, W.-S.; Min, D.S.; Kim, H.-Y.; Choi, K.-Y. Valproic Acid Induces Hair Regeneration in Murine Model and Activates Alkaline Phosphatase Activity in Human Dermal Papilla Cells. PLoS ONE 2012, 7, e34152. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.J.; Shin, H.; Park, Y.W.; Paik, S.H.; Park, W.S.; Jeong, Y.S.; Shin, H.J.; Kwon, O. Topical valproic acid increases the hair count in male patients with androgenetic alopecia: A randomized, comparative, clinical feasibility study using phototrichogram analysis. J. Dermatol. 2014, 41, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Gerstner, T.; Bell, N.; König, S. Oral valproic acid for epilepsy—long-term experience in therapy and side effects. Expert Opin. Pharmacother. 2008, 9, 285–292. [Google Scholar] [CrossRef]
- Sztajnkrycer, M.D. Valproic Acid Toxicity: Overview and Management. J. Toxicol. Clin. Toxicol. 2002, 40, 789–801. [Google Scholar] [CrossRef]
- Kim, K.-H.; Chung, W.-S.; Kim, Y.; Kim, K.-S.; Lee, I.-S.; Park, J.Y.; Jeong, H.-S.; Na, Y.-C.; Lee, C.-H.; Jang, H.-J. Transcriptomic Analysis Reveals Wound Healing ofMorus albaRoot Extract by Up-Regulating Keratin Filament and CXCL12/CXCR4 Signaling. Phytother. Res. 2015, 29, 1251–1258. [Google Scholar] [CrossRef]
- Cornejo, A.; Jiménez, J.M.; Caballero, L.; Melo, F.; Maccioni, R.B. Fulvic Acid Inhibits Aggregation and Promotes Disassembly of Tau Fibrils Associated with Alzheimer’s Disease. J. Alzheimer’s Dis. 2011, 27, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Acharya, S.B.; Frotan, M.H.; Goel, R.K.; Tripathi, S.K.; Das, P.K. Pharmacological actions of Shilajit. Indian J. Exp. Boil. 1988, 26, 775–777. [Google Scholar]
- Ozturk, E.; Ocak, N.; Coskun, I.; Turhan, S.; Erener, G. Effects of humic substances supplementation provided through drinking water on performance, carcass traits and meat quality of broilers. J. Anim. Physiol. Anim. Nutr. 2010, 94, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Vucskits, A.V.; Hullár, I.; Bersényi, A.; Andrásofszky, E.; Kulcsár, M.; Szabó, J. Effect of fulvic and humic acids on performance, immune response and thyroid function in rats. J. Anim. Physiol. Anim. Nutr. 2010, 94, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Shalini, S.R.; Srivastav, R. Antifungal Activity Screening and HPLC Analysis of Crude Extract from Tectona grandis, Shilajit, Valeriana wallachi. Internet J. Altern. Med. 2007, 5, 1540–2584. [Google Scholar]
- Bouzaiene, N.N.; Jaziri, S.K.; Kovacic, H.; Chekir-Ghedira, L.; Ghedira, K.; Luis, J. The effects of caffeic, coumaric and ferulic acids on proliferation, superoxide production, adhesion and migration of human tumor cells in vitro. Eur. J. Pharmacol. 2015, 766, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Ghaisas, M.M.; Kshirsagar, S.B.; Sahane, R.S. Evaluation of wound healing activity of ferulic acid in diabetic rats. Int. Wound J. 2012, 11, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Seluanov, A.; Vaidya, A.; Gorbunova, V. Establishing Primary Adult Fibroblast Cultures from Rodents. J. Vis. Exp. 2010, e2033. [Google Scholar] [CrossRef] [PubMed]
- Strickland, J.; Greenhalgh, D.; Koceva-Chyla, A.; Hennings, H.; Restrepo, C.; Balaschak, M.; Yuspa, S.H. Development of murine epidermal cell lines which contain an activated rasHa oncogene and form papillomas in skin grafts on athymic nude mouse hosts. Cancer Res. 1988, 48, 165–169. [Google Scholar] [PubMed]
- Oh, S.W.; Park, S.-H.; Lee, H.S.; Kang, M.; Lee, S.E.; Yoo, J.A.; Cho, J.Y.; Lee, J. Melanogenic mechanism of ethanolic extract of Dalbergia odorifera. Mol. Cell. Toxicol. 2017, 13, 453–459. [Google Scholar] [CrossRef]
- Joo, S.-Y.; Ha, Y.; Seo, D.Y.; Seo, G.Y.; Kim, Y.-J.; Choi, H.-Y. Effect of Prunus yedoensis Matsumura extract on the gene expression in HaCaT cells. Mol. Cell. Toxicol. 2018, 14, 337–345. [Google Scholar] [CrossRef]
- Kim, K.-H.; Park, J.Y.; Lee, I.-S.; Kim, Y.; Jang, H.-J. Proteins derived from Prunus armeniaca kernel are possible to cause Immunoglobulin E reactivity in human sera. Mol. Cell. Toxicol. 2017, 13, 213–220. [Google Scholar] [CrossRef]
- An, E.-J.; Kim, K.-H.; Lee, I.-S.; Park, J.Y.; Kim, Y.; Jung, W.S.; Kwon, D.; Jang, H.-J. Identification of Possibility of Glycyrrhiza uralensis as an Allergen by Protein Analysis. BioChip J. 2018, 12, 75–82. [Google Scholar] [CrossRef]
- Mazzalupo, S.; Wawersik, M.J.; Coulombe, P.A. An Ex Vivo Assay to Assess the Potential of Skin Keratinocytes for Wound Epithelialization. J. Investig. Dermatol. 2002, 118, 866–870. [Google Scholar] [CrossRef]
- Lee, S.E.; Park, H.R.; Yun, H.D.; Kim, H.; Jin, Y.-H.; Ahn, H.-J.; Cho, J.-J. Effect of crotonaldehyde on the induction of HO-1 expression in A549 cells. Mol. Cell. Toxicol. 2017, 13, 221–227. [Google Scholar] [CrossRef]
- Lampiasi, N.; Montana, G. The molecular events behind ferulic acid mediated modulation of IL-6 expression in LPS-activated Raw 264.7 cells. Immunobiology 2016, 221, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Martin, P. Wound Healing—Aiming for Perfect Skin Regeneration. Science 1997, 276, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Steinert, P.M.; Roop, D.R. Molecular and Cellular Biology of Intermediate Filaments. Annu. Rev. Biochem. 1988, 57, 593–625. [Google Scholar] [CrossRef]
- Heins, S.; Aebi, U. Making heads and tails of intermediate filament assembly, dynamics and networks. Curr. Opin. Cell Biol. 1994, 6, 25–33. [Google Scholar] [CrossRef]
- Jacob, J.T.; Coulombe, P.A.; Kwan, R.; Omary, M.B. Types I and II Keratin Intermediate Filaments. Cold Spring Harb. Perspect. Biol. 2018, 10, a018275. [Google Scholar] [CrossRef]
- Ito, M.; Liu, Y.; Yang, Z.; Nguyen, J.; Liang, F.; Morris, R.J.; Cotsarelis, G. Stem cells in the hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis. Nat. Med. 2005, 11, 1351–1354. [Google Scholar] [CrossRef] [PubMed]
- Paladini, R.D.; Coulombe, P.A. Directed Expression of Keratin 16 to the Progenitor Basal Cells of Transgenic Mouse Skin Delays Skin Maturation. J. Cell Biol. 1998, 142, 1035–1051. [Google Scholar] [CrossRef]
- Rotty, J.D.; Coulombe, P.A. A wound-induced keratin inhibits Src activity during keratinocyte migration and tissue repair. J. Cell Biol. 2012, 197, 381–389. [Google Scholar] [CrossRef]
- Wang, F.; Chen, S.; Liu, H.B.; Parent, C.A.; Coulombe, P.A. Keratin 6 regulates collective keratinocyte migration by altering cell–cell and cell–matrix adhesion. J. Cell Biol. 2018, 217, 4314–4330. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Manzanares, M.; Choi, C.K.; Horwitz, A.R. Integrins in cell migration—The actin connection. J. Cell Sci. 2008, 122, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Simpson, C.L.; Patel, D.M.; Green, K.J. Deconstructing the skin: Cytoarchitectural determinants of epidermal morphogenesis. Nat. Rev. Mol. Cell Biol. 2011, 12, 565–580. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Foote, H.P.; Lechler, T. β-Catenin protects the epidermis from mechanical stresses. J. Cell Biol. 2013, 202, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Voronkov, A.; Krauss, S. Wnt/beta-catenin signaling and small molecule inhibitors. Curr. Pharm. Des. 2013, 19, 634–664. [Google Scholar] [CrossRef] [PubMed]
- Geiger, B.; Ayalon, O. Cadherins. Annu. Rev. Cell Biol. 1992, 8, 307–332. [Google Scholar] [CrossRef] [PubMed]
- Hecht, A.; Kemler, R. Curbing the nuclear activities of β-catenin. EMBO Rep. 2000, 1, 24–28. [Google Scholar] [CrossRef]
- Van Mater, D.; Kolligs, F.T.; Dlugosz, A.A.; Fearon, E.R. Transient activation of beta -catenin signaling in cutaneous keratinocytes is sufficient to trigger the active growth phase of the hair cycle in mice. Genes Dev. 2003, 17, 1219–1224. [Google Scholar] [CrossRef]
- Choi, Y.S.; Zhang, Y.; Xu, M.; Yang, Y.; Ito, M.; Peng, T.; Cui, Z.; Nagy, A.; Hadjantonakis, A.-K.; Lang, R.A.; et al. Distinct Functions for Wnt/β-Catenin in Hair Follicle Stem Cell Proliferation and Survival and Interfollicular Epidermal Homeostasis. Cell Stem Cell 2013, 13, 720–733. [Google Scholar] [CrossRef] [PubMed]
- Waikel, R.L.; Kawachi, Y.; Waikel, P.A.; Wang, X.-J.; Roop, D.R. Deregulated expression of c-Myc depletes epidermal stem cells. Nat. Genet. 2001, 28, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Huelsken, J.; Vogel, R.; Erdmann, B.; Cotsarelis, G.; Birchmeier, W. Beta-Catenin controls hair follicle morphogenesis and stem cell differentiation in the skin. Cell 2001, 105, 533–545. [Google Scholar] [CrossRef]
- Arwert, E.N.; Hoste, E.; Watt, F.M. Epithelial stem cells, wound healing and cancer. Nat. Rev. Cancer 2012, 12, 170–180. [Google Scholar] [CrossRef]
- Bergmann, C.; Akhmetshina, A.; Dees, C.; Palumbo, K.; Zerr, P.; Beyer, C.; Zwerina, J.; Distler, O.; Schett, G.; Distler, J.H.W. Inhibition of glycogen synthase kinase 3 induces dermal fibrosis by activation of the canonical Wnt pathway. Ann. Rheum. Dis. 2011, 70, 2191–2198. [Google Scholar] [CrossRef]
- Lee, S.-H.; Kim, M.-Y.; Kim, H.-Y.; Lee, Y.M.; Kim, H.; Nam, K.A.; Roh, M.R.; Min, D.S.; Chung, K.Y.; Choi, K.Y. The Dishevelled-binding protein CXXC5 negatively regulates cutaneous wound healing. J. Exp. Med. 2015, 212, 1061–1080. [Google Scholar] [CrossRef] [PubMed]
- Bos, J.D.; Meinardi, M.M.H.M. The 500 Dalton rule for the skin penetration of chemical compounds and drugs. Exp. Dermatol. 2000, 9, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.; Peters, D.H.; McTavish, D. Valproic acid. A reappraisal of its pharmacological properties and clinical efficacy in epilepsy. Drugs 1994, 47, 332–372. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.; Raviv, S.; Adir, O.; Padmanabhan, K.; Soffer, A.; Luxenburg, C. The Wave complex controls epidermal morphogenesis and proliferation by suppressing Wnt–Sox9 signaling. J. Cell Biol. 2019, 218, 1390–1406. [Google Scholar] [CrossRef] [PubMed]
- Leachman, S.A.; Kaspar, R.L.; Fleckman, P.; Florell, S.R.; Smith, F.J.; McLean, W.I.; Lunny, D.P.; Milstone, L.M.; Van Steensel, M.A.; Munro, C.S.; et al. Clinical and Pathological Features of Pachyonychia Congenita. J. Investig. Dermatol. Symp. Proc. 2005, 10, 3–17. [Google Scholar] [CrossRef] [PubMed]
- De The, H.; Vivanco-Ruiz, M.D.M.; Tiollais, P.; Stunnenberg, H.; Dejean, A. Identification of a retinoic acid responsive element in the retinoic acid receptor & beta;gene. Nat. Cell Biol. 1990, 343, 177–180. [Google Scholar] [CrossRef]
- Zhao, Y.; Gartner, U.; Smith, F.J.; McLean, W.I. Statins Downregulate K6a Promoter Activity: A Possible Therapeutic Avenue for Pachyonychia Congenita. J. Investig. Dermatol. 2011, 131, 1045–1052. [Google Scholar] [CrossRef]
- DiGiovanna, J.J. Fracturing Support for the Role of Systemic Retinoid Therapy as a Cause of Bone Demineralization. Arch. Dermatol. 2010, 146, 551–553. [Google Scholar] [CrossRef] [PubMed]
- Ormerod, A.; Campalani, E.; Goodfield, M. British Association of Dermatologists guidelines on the efficacy and use of acitretin in dermatology. Br. J. Dermatol. 2010, 162, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Moctezuma-Velázquez, C.; Abraldes, J.G.; Montano-Loza, A.J. The Use of Statins in Patients With Chronic Liver Disease and Cirrhosis. Curr. Treat. Options Gastroenterol. 2018, 16, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Robin, N.C.; Agoston, Z.; Biechele, T.L.; James, R.G.; Berndt, J.D.; Moon, R.T. Simvastatin promotes adult hippocampal neurogenesis by enhancing Wnt/β-catenin signaling. Stem Cell Rep. 2014, 2, 9–17. [Google Scholar] [CrossRef]
- Strand, N.S.; Hoi, K.K.; Phan, T.M.; Ray, C.A.; Berndt, J.D.; Moon, R.T. Wnt/β-catenin signaling promotes regeneration after adult zebrafish spinal cord injury. Biochem. Biophys. Res. Commun. 2016, 477, 952–956. [Google Scholar] [CrossRef]
- Zhang, Y.; Talalay, P.; Cho, C.G.; Posner, G.H. A major inducer of anticarcinogenic protective enzymes from broccoli: Isolation and elucidation of structure. Proc. Natl. Acad. Sci. USA 1992, 89, 2399–2403. [Google Scholar] [CrossRef] [PubMed]
- Kerns, M.; DePianto, D.; Yamamoto, M.; Coulombe, P.A. Differential Modulation of Keratin Expression by Sulforaphane Occurs via Nrf2-dependent and -independent Pathways in Skin Epithelia. Mol. Biol. Cell 2010, 21, 4068–4075. [Google Scholar] [CrossRef]
- Kerns, M.L.; DePianto, D.; Dinkova-Kostova, A.T.; Talalay, P.; Coulombe, P.A. Reprogramming of keratin biosynthesis by sulforaphane restores skin integrity in epidermolysis bullosa simplex. Proc. Natl. Acad. Sci. USA 2007, 104, 14460–14465. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe Name | Description | Gene Symbol | Fold Change # |
|---|---|---|---|
| Keratins | |||
| 1422784_at | keratin 6A | Krt6a | 14.6901 |
| 1423227_at | keratin 17 | Krt17 | 7.9747 |
| 1424096_at | keratin 5 | Krt5 | 7.3153 |
| 1423935_x_at | keratin 14 | Krt14 | 8.1810 |
| Probe Name | Description | Gene Symbol | Fold Change # |
|---|---|---|---|
| Collagen | |||
| 1455494_at | collagen, type I, alpha 1 | Col1a1 | 2.5030 |
| 1423110_at | collagen, type I, alpha 2 | Col1a2 | 2.6747 |
| 1427884_at | collagen, type III, alpha 1 | Col3a1 | 3.4892 |
| 1418799_a_at | collagen, type XVII, alpha 1 | Col17a1 | 2.4751 |
| Tight junction | |||
| 1437932_a_at | claudin 1 | Cldn1 | 2.1909 |
| 1434651_a_at | claudin 3 | Cldn3 | 2.0510 |
| Adherens junction | |||
| 1448261_at | cadherin 1 | Cdh1 | 2.2079 |
| Desmosome | |||
| 1435494_s_at | desmoplakin | Dsp | 4.6570 |
| 1434534_at | desmocollin 3 | Dsc3 | 2.1909 |
| Gap junction | |||
| 1415801_at | gap junction protein, alpha 1 | Gja1 | −0.39884 |
| Probe Name | Description | Gene Symbol | Fold Change # |
|---|---|---|---|
| Oncogene | |||
| 1423240_at | Rous sarcoma oncogene | Src | 2.1337 |
| Integrin linked kinases | |||
| 1449942_a_at | Integrin-linked kinase | Ilk | 2.0405 |
| Chemokine ligand | |||
| 1417574_at | chemokine (C-X-C motif) ligand 12 | Cxcl12 | 3.078 |
| Fibroblast growth factor | |||
| 1422916_at | fibroblast growth factor 21 | Fgf21 | 2.0611 |
| A disintegrin and metallopeptidase | |||
| 1450105_at | a disintegrin and metallopeptidase domain 10 | Adam10 | −0.2529 |
| 1421857_at | a disintegrin and metallopeptidase domain 17 | Adam17 | −0.1847 |
| Wnt signaling | |||
| 1448818_at | wingless-related MMTV integration site 5A | Wnt5a | −0.4180 |
| 1430533_a_at | catenin (cadherin associated protein), beta 1 | Ctnnb1 | −0.0786 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.-H.; Jung, J.H.; Chung, W.-S.; Lee, C.-H.; Jang, H.-J. Ferulic Acid Induces Keratin 6α via Inhibition of Nuclear β-Catenin Accumulation and Activation of Nrf2 in Wound-Induced Inflammation. Biomedicines 2021, 9, 459. https://doi.org/10.3390/biomedicines9050459
Kim K-H, Jung JH, Chung W-S, Lee C-H, Jang H-J. Ferulic Acid Induces Keratin 6α via Inhibition of Nuclear β-Catenin Accumulation and Activation of Nrf2 in Wound-Induced Inflammation. Biomedicines. 2021; 9(5):459. https://doi.org/10.3390/biomedicines9050459
Chicago/Turabian StyleKim, Kang-Hoon, Ji Hoon Jung, Won-Seok Chung, Chang-Hun Lee, and Hyeung-Jin Jang. 2021. "Ferulic Acid Induces Keratin 6α via Inhibition of Nuclear β-Catenin Accumulation and Activation of Nrf2 in Wound-Induced Inflammation" Biomedicines 9, no. 5: 459. https://doi.org/10.3390/biomedicines9050459
APA StyleKim, K.-H., Jung, J. H., Chung, W.-S., Lee, C.-H., & Jang, H.-J. (2021). Ferulic Acid Induces Keratin 6α via Inhibition of Nuclear β-Catenin Accumulation and Activation of Nrf2 in Wound-Induced Inflammation. Biomedicines, 9(5), 459. https://doi.org/10.3390/biomedicines9050459