
In Vivo and In Vitro Characterization of Primary Human Liver Macrophages and Their Inflammatory State

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Media and Chemicals
2.2. Cell Culture
2.3. Tissue Samples
2.4. Immunohistochemical Staining of Paraffin Embedded Tissue
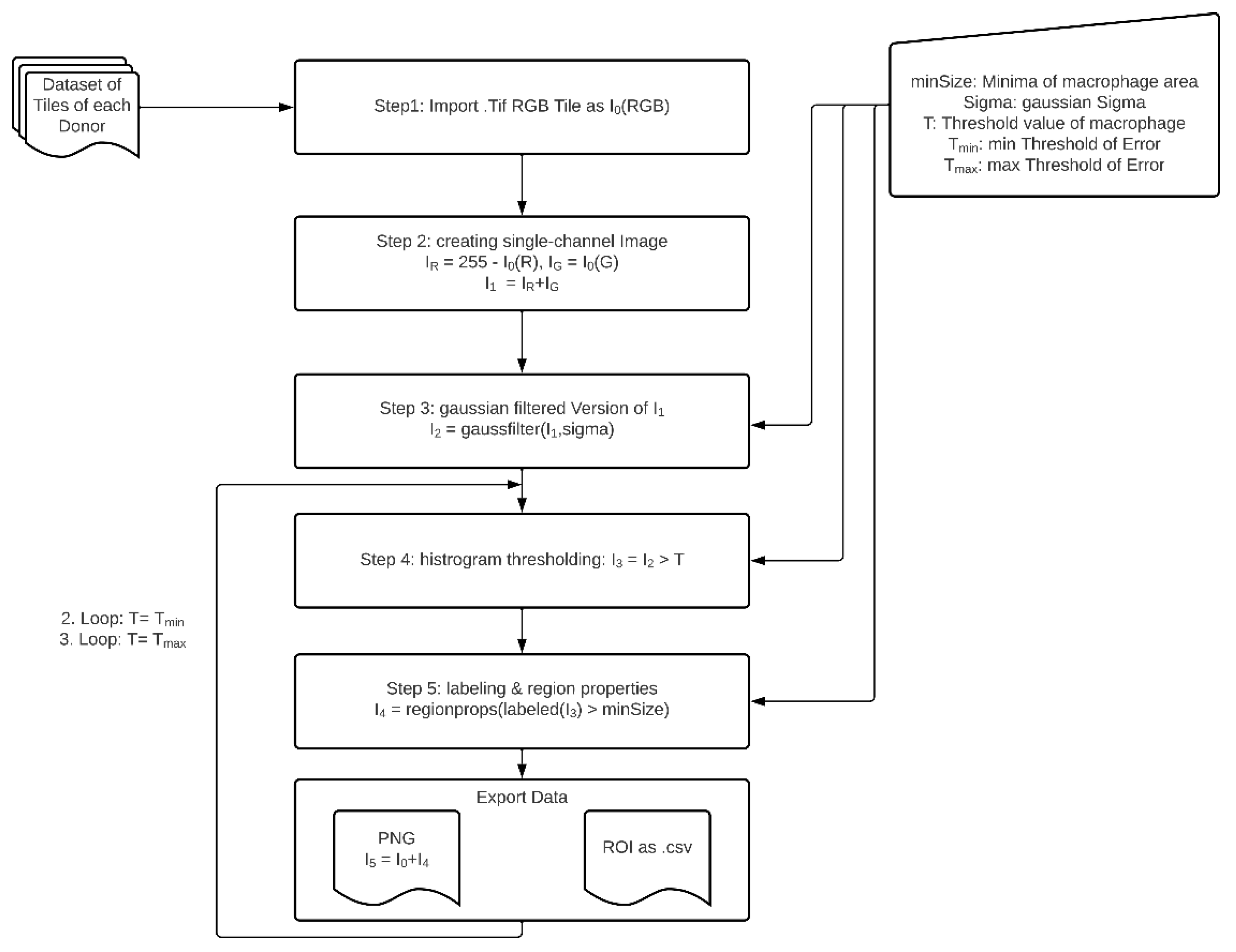
2.5. Image Analysis of Immunohistochemical Stainings
2.6. Isolation of Hepatic Macrophages
2.7. XTT Viability Assay
2.8. DCF Assay for ROI Detection
2.9. Determination of Protein Content
2.10. Immunofluorescence Staining
2.10.1. Cell Staining for Flow Cytometry
2.10.2. Cell Staining of Adherent Cells
2.11. Statistical Analysis
3. Results
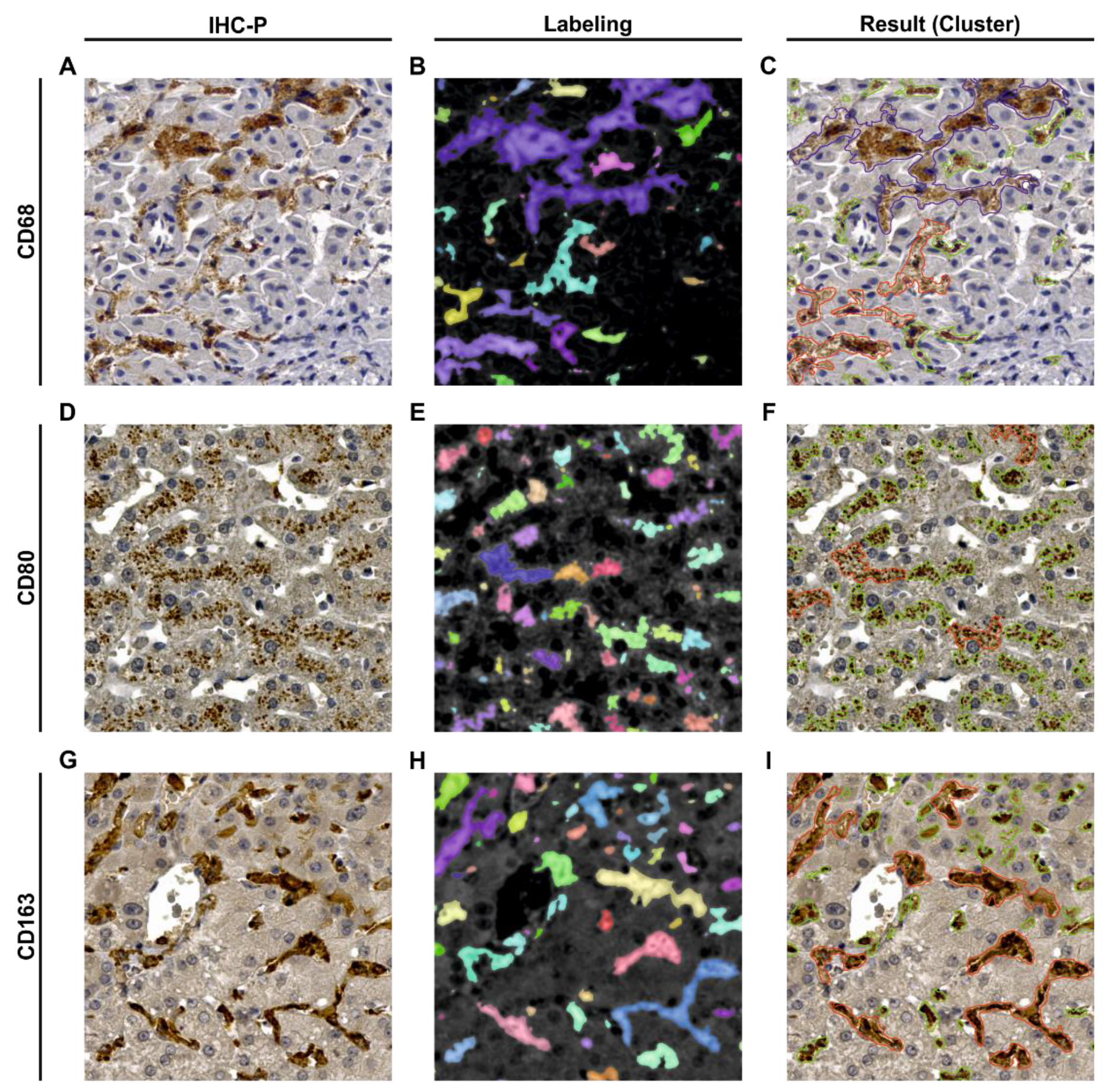
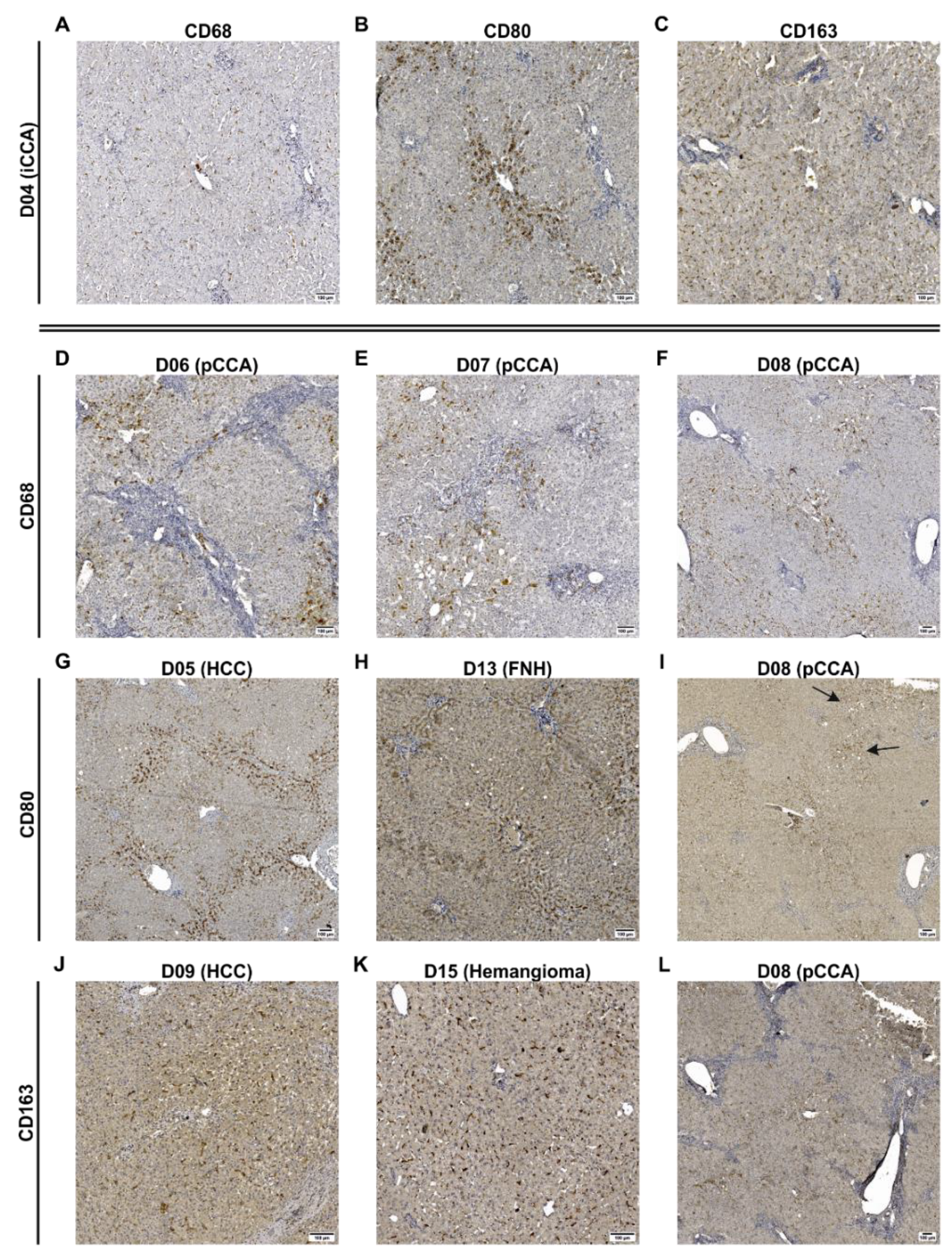
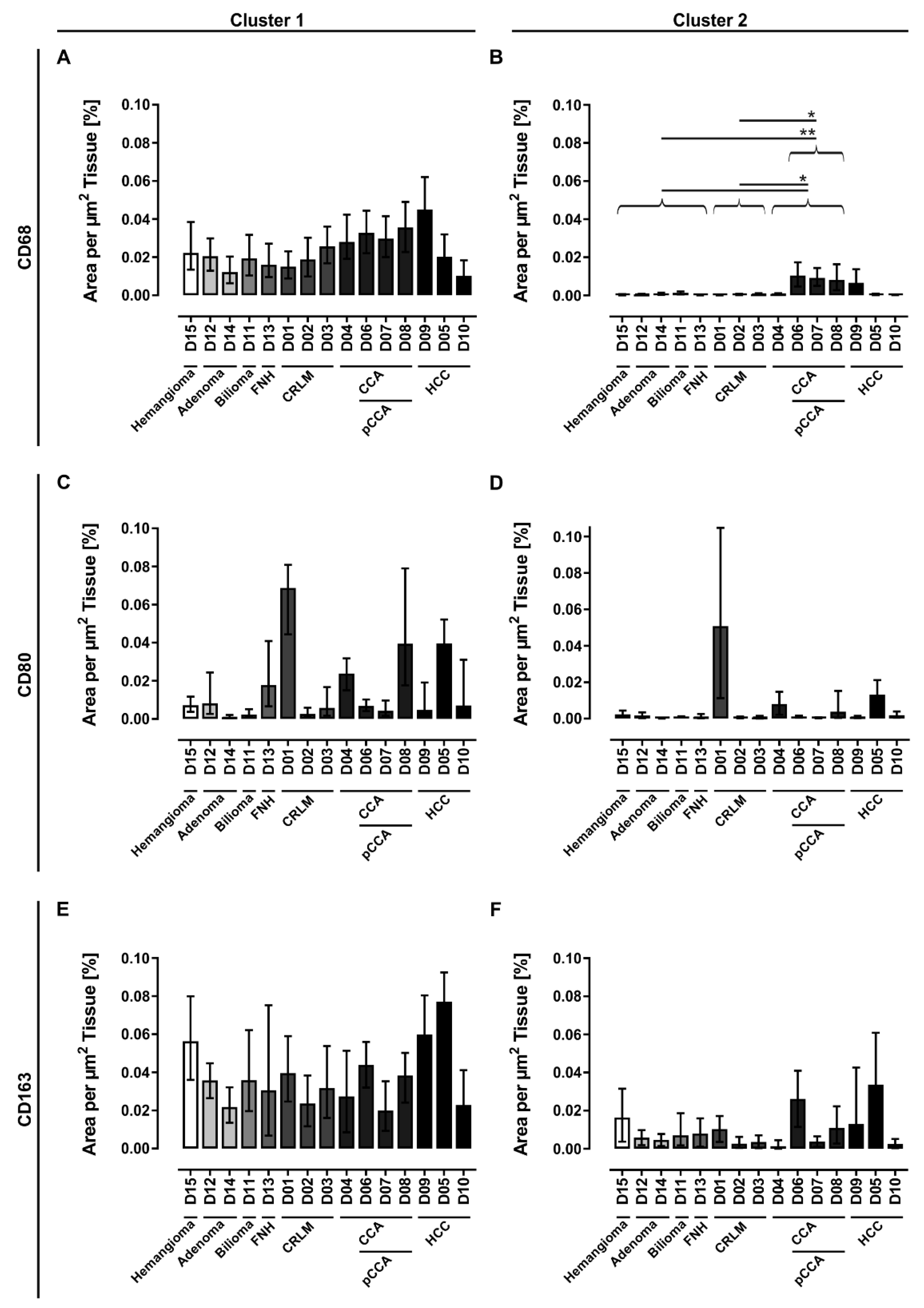
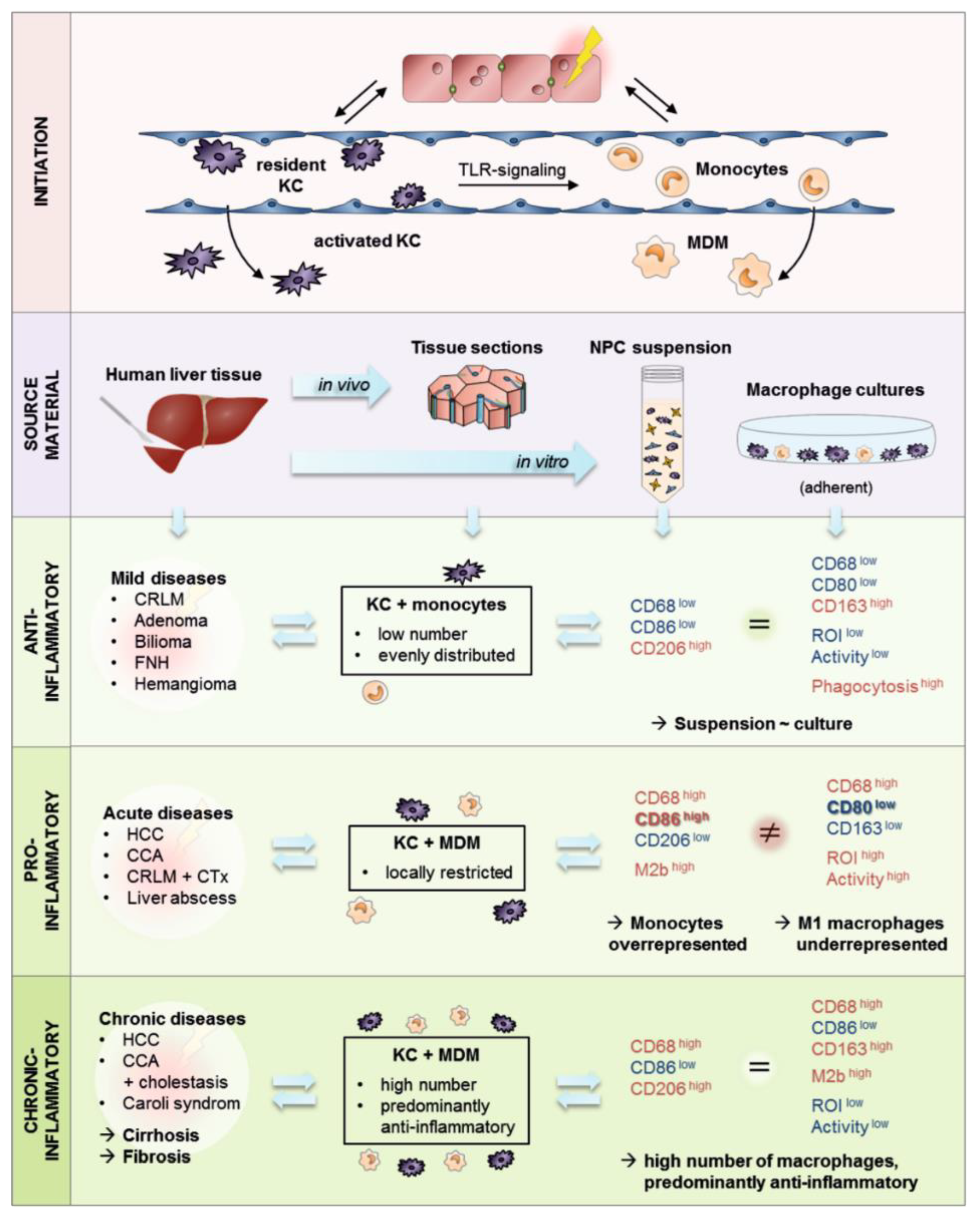
3.1. Human Liver Tissue Sections Showed Disease Specific Patterns of Lobular Macrophage Reactivity
3.2. Bioinformatic Analysis Revealed That Human Liver Macrophages In Vivo Are Characterized by a Predominantly Anti-Inflammatory Activation State
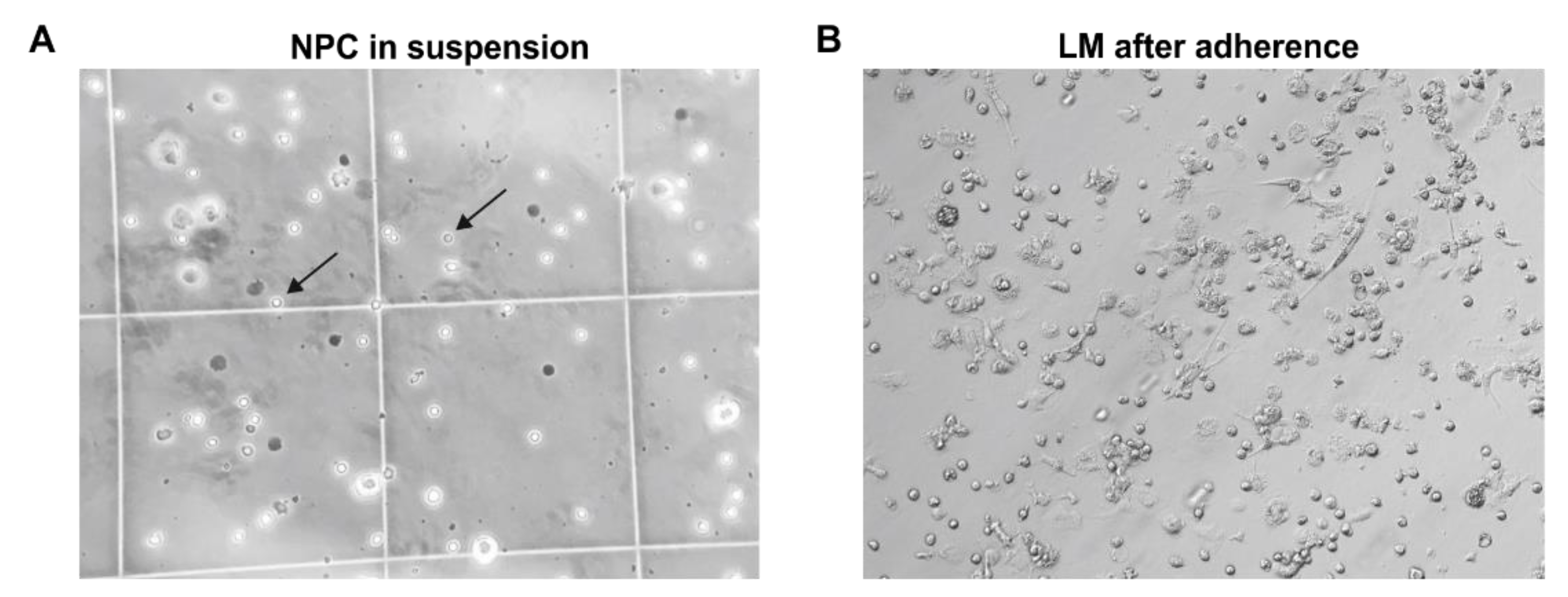
3.3. Isolation of Primary Human Liver Macrophages Results in Cell Suspensions and Cultures of Varying Quality and Quantity
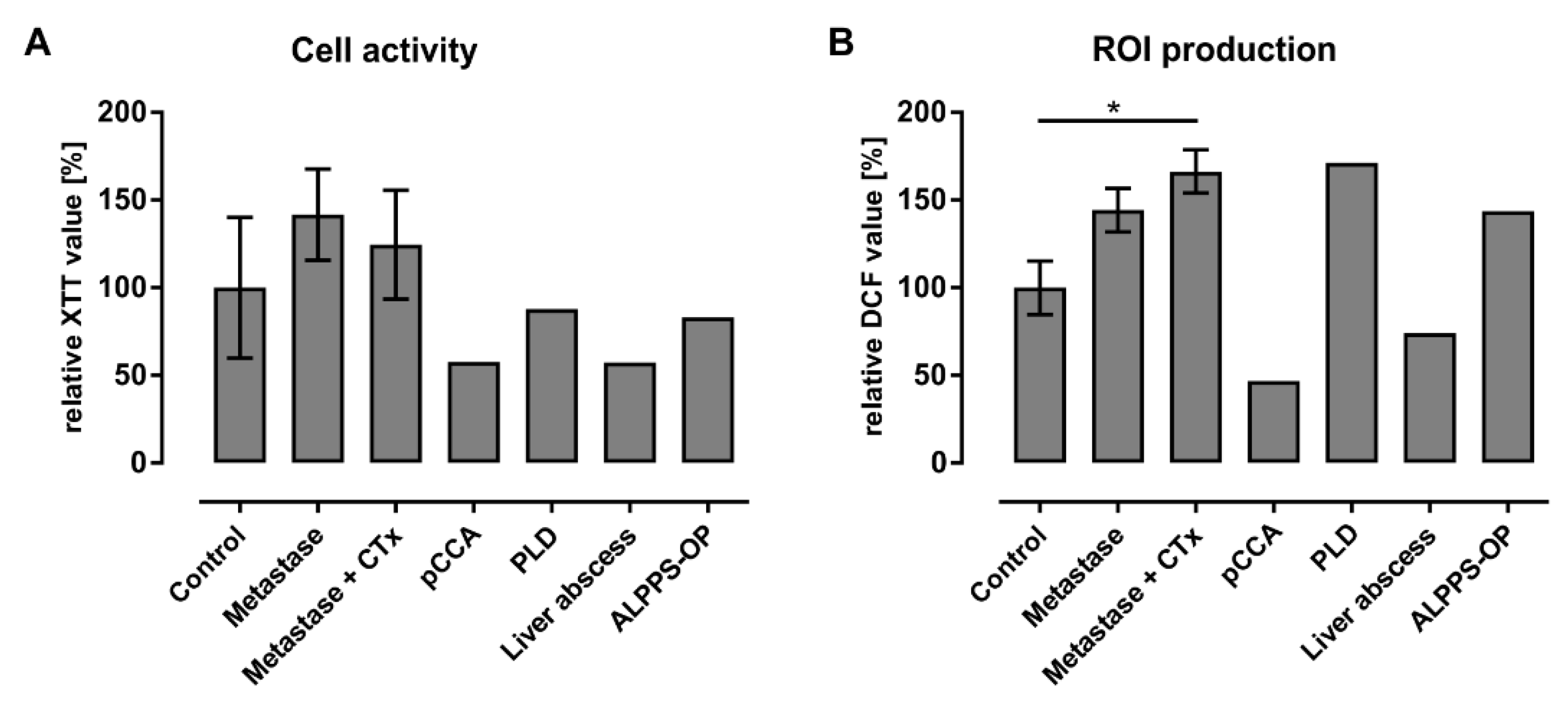
3.4. Initial Characterization of Liver Macrophages Revealed Anamnesis-Dependent Metabolic and Inflammatory Activation
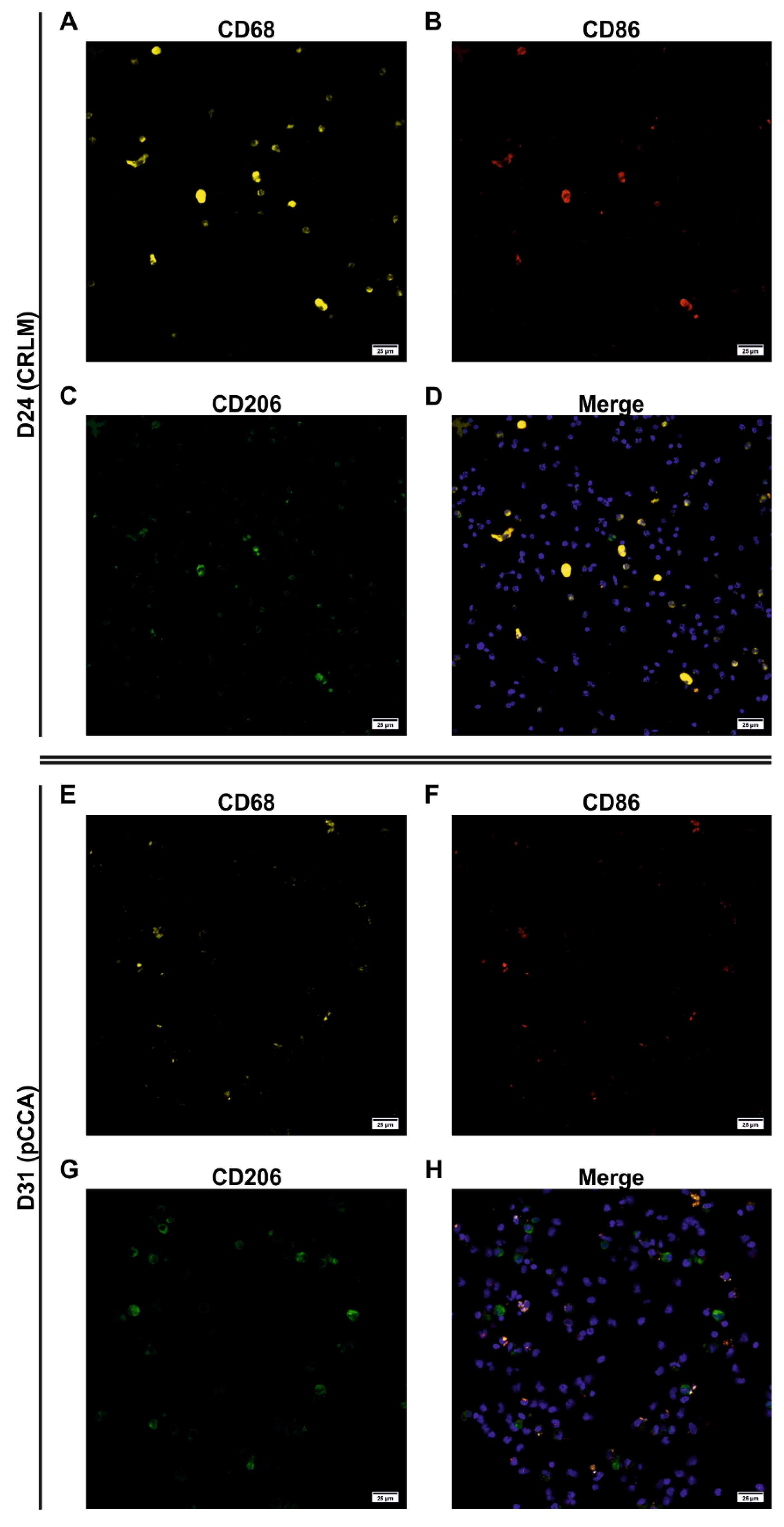
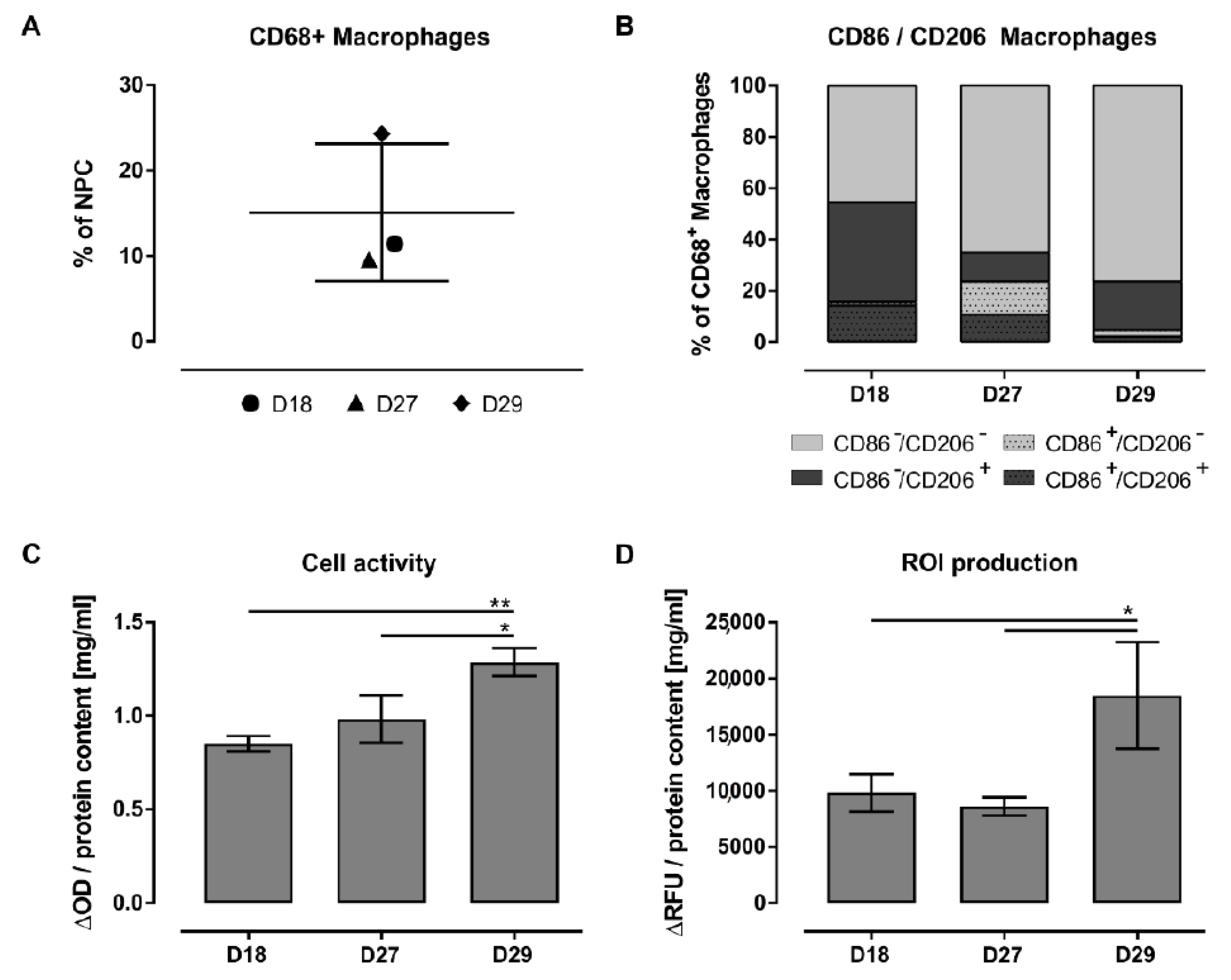
3.5. Characterization of the Adherent NPCs Fraction Showed Various Macrophage Subpopulations
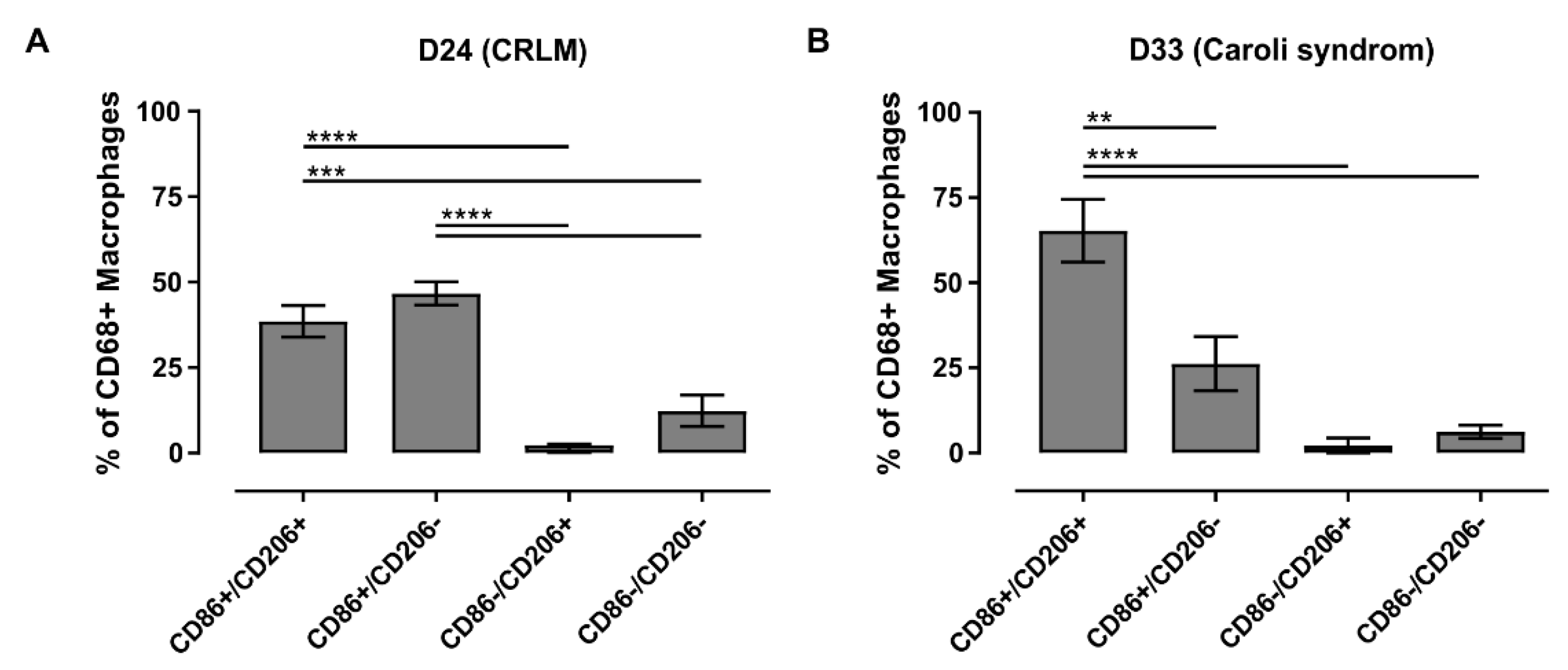
3.6. FACS Analysis Allowed a More Robust Characterization of Macrophage Subpopulations
4. Discussion
4.1. Selectivity of Specific Antigens for KC Characterization
4.2. The majority of Investigated Liver Pathologies Are Characterized by Anti-Inflammatory Macrophages
4.3. Patients Diagnosed with Klatskin Tumors Showed Acute to Chronic Progression of Inflammation
4.4. Isolation of Primary Human Non-Parenchymal Liver Cells Resulted in a Macrophage-Rich Fraction Showing Donor-Related Activation and Reactivity
4.5. Isolated Primary Hepatic Macrophages Are a Heterogeneous Cell Fraction with Predominant Anti-Inflammatory Characteristics
4.6. Limitations of the LM Characterization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbrevations
| ALPPS | “associating liver partition and portal vein ligation for staged hepatectomy” operation |
| APCs | Antigen-presenting cells |
| BCA | Bicinchoninic acid |
| BSA | Bovine serum albumin |
| CC | Cholangiocytes |
| CRLM | Colorectal liver metastasis |
| CRP | C-reactive protein |
| CSV | Comma Separated Values |
| CTx | Chemotherapy |
| CZI | Carl Zeiss Image |
| DAB | 3,3’-Diaminobenzidine |
| DCs | Dendritic cells |
| DCF | 2,7–dichlorofluorescein |
| DCF-DA | 2,7–dichlorofluorescein diacetate |
| DILI | Drug induced liver injuries |
| ECM | Extracellular matrix |
| EDTA | Ethylenediaminetetraacetic acid |
| ER | Endoplasmic reticulum |
| FACS | Fluorescence activated cell sorting |
| FMO | Fluorescence Minus One |
| FNH | Focal nodular hyperplasia |
| HBV | Hepatitis B viruses |
| HCC | Hepatocellular carcinoma |
| HCV | Hepatitis C viruses |
| HRP | Horseradish peroxidase |
| iCCA | Intrahepatic cholangiocarcinoma |
| IHC | Immunohistochemistry |
| KCs | Kupffer cells |
| LMs | Liver macrophages |
| LPS | Lipopolysaccharides |
| LSM | Laser scanning microscope |
| MHC | Major histocompatibility complex |
| MDMs | Monocyte-derived macrophages |
| NAFLD | Non-alcoholic fatty liver disease |
| NKC | Natural killer cells |
| NO | Nitric oxide |
| NPCs | Non-parenchymal cells |
| PBS | Phosphate-buffered saline |
| pCCA | Perihilar cholangiocarcinoma |
| PD-L1 | Programmed cell death ligand-1 |
| PFA | Paraformaldehyde |
| PGE2 | Prostaglandin E2 |
| PHH | Primary human hepatocytes |
| PLD | Polycystic liver disease |
| PMA | Phorbol-12-myristat-13-acetat |
| PNG | Portable Network Graphic |
| PVE | Portal vein embolization |
| ROIs | Reactive oxygen intermediates |
| ROS | Reactive oxygen species |
| RT | Room temperature |
| SD | Standard deviation |
| SDS | Sodium dodecyl sulfate |
| TBS | Tris-buffered saline |
| TIFF | Tagged Image File Format |
| TLR | Toll-like receptor |
| s/p | status post |
| XTT | 2,3-Bis-(2-Methoxy-4-Nitro-5-Sulfophenyl)-2H-Tetrazolium-5-Carboxanilide |
| WSI | Whole Slide Images |
References
- Taub, R. Liver regeneration: From myth to mechanism. Nat. Rev. Mol. Cell Biol. 2004, 5, 836–847. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Plüddemann, A. Tissue macrophages: Heterogeneity and functions. BMC Biol. 2017, 15, 1–18. [Google Scholar] [CrossRef]
- Guillot, A.; Tacke, F. Liver Macrophages: Old Dogmas and New Insights. Hepatol. Commun. 2019, 3, 730–743. [Google Scholar] [CrossRef]
- Blériot, C.; Ginhoux, F. Understanding the Heterogeneity of Resident Liver Macrophages. Front. Immunol. 2019, 10, 2694. [Google Scholar] [CrossRef]
- Dong, P.; Ma, L.; Liu, L.; Zhao, G.; Zhang, S.; Dong, L.; Xue, R.; Chen, S. CD86+/CD206+, Diametrically Polarized Tumor-Associated Macrophages, Predict Hepatocellular Carcinoma Patient Prognosis. Int. J. Mol. Sci. 2016, 17, 320. [Google Scholar] [CrossRef] [PubMed]
- Jenne, C.N.; Kubes, P. Immune surveillance by the liver. Nat. Immunol. 2013, 14, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Ju, C.; Tacke, F. Hepatic macrophages in homeostasis and liver diseases: From pathogenesis to novel therapeutic strategies. Cell. Mol. Immunol. 2016, 13, 316–327. [Google Scholar] [CrossRef]
- Beljaars, L.; Schippers, M.; Reker-Smit, C.; Martinez, F.O.; Helming, L.; Poelstra, K.; Melgert, B.N. Hepatic Localization of Macrophage Phenotypes during Fibrogenesis and Resolution of Fibrosis in Mice and Humans. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Martinez, F.O. Alternative Activation of Macrophages: Mechanism and Functions. Immun. 2010, 32, 593–604. [Google Scholar] [CrossRef]
- Ritz, T.; Krenkel, O.; Tacke, F. Dynamic plasticity of macrophage functions in diseased liver. Cell. Immunol. 2018, 330, 175–182. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Krenkel, O.; Tacke, O.K.F. Liver macrophages in tissue homeostasis and disease. Nat. Rev. Immunol. 2017, 17, 306–321. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage Activation and Polarization: Nomenclature and Experimental Guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Tacke, F. Targeting hepatic macrophages to treat liver diseases. J. Hepatol. 2017, 66, 1300–1312. [Google Scholar] [CrossRef]
- ZEISS Mikroskopsoftware ZEN-Analysieren Sie Bilddaten oder Mikroskopie-Bilder und steuern Sie Mikroskopkomponenten. 2020-12-03T10:11:27.000Z. Available online: https://www.zeiss.de/mikroskopie/produkte/mikroskopsoftware/zen.html (accessed on 3 December 2020).
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Yen, J.-C.; Chang, F.-J.; Chang, S. A new criterion for automatic multilevel thresholding. IEEE Trans. Image Process. 1995, 4, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Kegel, V.; Deharde, D.; Pfeiffer, E.; Zeilinger, K.; Seehofer, D.; Damm, G. Protocol for Isolation of Primary Human Hepatocytes and Corresponding Major Populations of Non-parenchymal Liver Cells. J. Vis. Exp. 2016, e53069. [Google Scholar] [CrossRef]
- Damm, G.; Schicht, G.; Zimmermann, A.; Rennert, C.; Fischer, N.; Kießig, M.; Wagner, T.; Kegel, V.; Seehofer, D. Effect of glucose and insulin supplementation on the isolation of primary human hepatocytes. EXCLI J. 2019, 18, 1071–1091. [Google Scholar]
- Kegel, V.; Pfeiffer, E.; Burkhardt, B.; Liu, J.L.; Zeilinger, K.; Nüssler, A.K.; Seehofer, D.; Damm, G. Subtoxic Concentrations of Hepatotoxic Drugs Lead to Kupffer Cell Activation in a HumanIn VitroLiver Model: An Approach to Study DILI. Mediat. Inflamm. 2015, 2015, 1–14. [Google Scholar] [CrossRef]
- Pfeiffer, E.; Kegel, V.; Zeilinger, K.; Hengstler, J.G.; Nüssler, A.K.; Seehofer, D.; Damm, G. Featured Article: Isolation, characterization, and cultivation of human hepatocytes and non-parenchymal liver cells. Exp. Biol. Med. 2014, 240, 645–656. [Google Scholar] [CrossRef]
- Schulz, D.; Severin, Y.; Zanotelli, V.R.T.; Bodenmiller, B. In-Depth Characterization of Monocyte-Derived Macrophages using a Mass Cytometry-Based Phagocytosis Assay. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- A Chistiakov, D.; Killingsworth, M.C.; A Myasoedova, V.; Orekhov, A.N.; Bobryshev, Y.V. CD68/macrosialin: Not just a histochemical marker. Lab. Investig. 2017, 97, 4–13. [Google Scholar] [CrossRef]
- Elchaninov, A.V.; Fatkhudinov, T.K.; Vishnyakova, P.A.; Lokhonina, A.V.; Sukhikh, G.T. Phenotypical and Functional Polymorphism of Liver Resident Macrophages. Cells 2019, 8, 1032. [Google Scholar] [CrossRef] [PubMed]
- Betjes, M.G.; Haks, M.C.; Tuk, C.W.; Beelen, R.H. Monoclonal Antibody EBM11 (Anti-CD68) Discriminates between Dendritic Cells and Macrophages after Short-Term Culture. Immunobiology 1991, 183, 79–87. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Myasoedova, V.A.; Revin, V.V.; Orekhov, A.N.; Bobryshev, Y.V. The impact of interferon-regulatory factors to macrophage differentiation and polarization into M1 and M2. Immunobiology 2018, 223, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Romo-Tena, J.; Gómez-Martín, D.; Alcocer-Varela, J. CTLA-4 and autoimmunity: New insights into the dual regulator of tolerance. Autoimmun. Rev. 2013, 12, 1171–1176. [Google Scholar] [CrossRef] [PubMed]
- Said, E.A.; Al-Reesi, I.; Al-Riyami, M.; Al-Naamani, K.; Al-Sinawi, S.; Al-Balushi, M.S.; Koh, C.Y.; Al-Busaidi, J.Z.; Idris, M.A.; Al-Jabri, A.A. Increased CD86 but Not CD80 and PD-L1 Expression on Liver CD68+ Cells during Chronic HBV Infection. PLOS ONE 2016, 11, e0158265. [Google Scholar] [CrossRef]
- Bobryshev, Y.V. Dendritic cells and their role in atherogenesis. Lab. Investig. 2010, 90, 970–984. [Google Scholar] [CrossRef]
- Leifeld, L.; Trautwein, C.; Dumoulin, F.L.; Manns, M.P.; Sauerbruch, T.; Spengler, U. Enhanced Expression of CD80 (B7-1), CD86 (B7-2), and CD40 and Their Ligands CD28 and CD154 in Fulminant Hepatic Failure. Am. J. Pathol. 1999, 154, 1711–1720. [Google Scholar] [CrossRef]
- Nielsen, M.C.; Gantzel, R.H.; Clària, J.; Trebicka, J.; Møller, H.J.; Grønbæk, H. Macrophage Activation Markers, CD163 and CD206, in Acute-on-Chronic Liver Failure. Cells 2020, 9, 1175. [Google Scholar] [CrossRef] [PubMed]
- Pulford, K.; Micklem, K.; McCarthy, S.; Cordell, J.; Jones, M.; Mason, D.Y. A monocyte/macrophage antigen recognized by the four antibodies GHI/61, Ber-MAC3, Ki-M8 and SM4. Immunology 1992, 75, 588–595. [Google Scholar] [PubMed]
- A Wynn, T.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef] [PubMed]
- Tacke, F.; Zimmermann, H.W. Macrophage heterogeneity in liver injury and fibrosis. J. Hepatol. 2014, 60, 1090–1096. [Google Scholar] [CrossRef]
- Shan, Z.; Ju, C. Hepatic Macrophages in Liver Injury. Front. Immunol. 2020, 11, 322. [Google Scholar] [CrossRef]
- Rudnick, D.A.; Davidson, N.O. Functional Relationships between Lipid Metabolism and Liver Regeneration. Int. J. Hepatol. 2012, 2012, 1–8. [Google Scholar] [CrossRef]
- Della Fazia, M.A.; Servillo, G. Foie gras and liver regeneration: A fat dilemma. Cell Stress 2018, 2, 162–175. [Google Scholar] [CrossRef]
- Millonig, G.; Reimann, F.M.; Friedrich, S.; Fonouni, H.; Mehrabi, A.; Büchler, M.W.; Seitz, H.K.; Mueller, S. Extrahepatic cholestasis increases liver stiffness (FibroScan) irrespective of fibrosis. Hepatology 2008, 48, 1718–1723. [Google Scholar] [CrossRef]
- Karlmark, K.R.; Weiskirchen, R.; Zimmermann, H.W.; Gassler, N.; Ginhoux, F.; Weber, C.; Merad, M.; Luedde, T.; Trautwein, C.; Tacke, F. Hepatic recruitment of the inflammatory Gr1+monocyte subset upon liver injury promotes hepatic fibrosis. Hepatology 2009, 50, 261–274. [Google Scholar] [CrossRef]
- Gehring, S.; Dickson, E.M.; Martin, M.E.S.; Van Rooijen, N.; Papa, E.F.; Harty, M.W.; Tracy, T.F.; Gregory, S.H. Kupffer Cells Abrogate Cholestatic Liver Injury in Mice. Gastroenterology 2006, 130, 810–822. [Google Scholar] [CrossRef]
- Hosomura, N.; Kono, H.; Tsuchiya, M.; Ishii, K.; Ogiku, M.; Matsuda, M.; Fujii, H. HCV-Related Proteins Activate Kupffer Cells Isolated from Human Liver Tissues. Dig. Dis. Sci. 2010, 56, 1057–1064. [Google Scholar] [CrossRef]
- Uchikura, K.; Wada, T.; Hoshino, S.; Nagakawa, Y.; Aiko, T.; Bulkley, G.B.; Klein, A.S.; Sun, Z. Lipopolysaccharides induced increases in Fas ligand expression by Kupffer cells via mechanisms dependent on reactive oxygen species. Am. J. Physiol. Liver Physiol. 2004, 287, G620–G626. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, H.L.; Prats, M.M.; Sasatomi, E. Chemotherapy-induced Sinusoidal Injury (CSI) score: A novel histologic assessment of chemotherapy-related hepatic sinusoidal injury in patients with colorectal liver metastasis. BMC Cancer 2017, 17, 35. [Google Scholar] [CrossRef] [PubMed]
- Axelrod, H.D.; Pienta, K.J.; Valkenburg, K.C. Optimization of Immunofluorescent Detection of Bone Marrow Disseminated Tumor Cells. Biol. Proced. Online 2018, 20, 13. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M. The many faces of macrophage activation. J. Leukoc. Biol. 2003, 73, 209–212. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, S.; Wu, H.; Rong, X.; Guo, J. M2b macrophage polarization and its roles in diseases. J. Leukoc. Biol. 2019, 106, 345–358. [Google Scholar] [CrossRef]
- Taylor, P.; Martinez-Pomares, L.; Stacey, M.; Lin, H.-H.; Brown, G.; Gordon, S. MACROPHAGE RECEPTORS AND IMMUNE RECOGNITION. Annu. Rev. Immunol. 2005, 23, 901–944. [Google Scholar] [CrossRef]
- Kosters, A.; Karpen, S.J. The Role of Inflammation in Cholestasis: Clinical and Basic Aspects. Semin. Liver Dis. 2010, 30, 186–194. [Google Scholar] [CrossRef]
- Bilzer, M.; Roggel, F.; Gerbes, A.L. Role of Kupffer cells in host defense and liver disease. Liver Int. 2006, 26, 1175–1186. [Google Scholar] [CrossRef]
- Liu, Y.; Ji, X.; Kang, N.; Zhou, J.; Liang, X.; Li, J.; Han, T.; Zhao, C.; Yang, T. Tumor necrosis factor α inhibition overcomes immunosuppressive M2b macrophage-induced bevacizumab resistance in triple-negative breast cancer. Cell Death Dis. 2020, 11, 993. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Summary | Donor ID | Age, Sex | Disease | Steatosis | Portal Fibrosis | Portal Inflammation |
|---|---|---|---|---|---|---|
| Primary liver tumors (N = 7) | D05 | 77, f 1 | HCC 2, low cholestasis | 5% | moderate | mild |
| D09 | 59, m 3 | HCC, cirrhosis | 5% | slight to moderate | mild to moderate | |
| D10 | 79, f | HCC, Child A cirrhosis | 60%, NAFLD 4/AFLD 5 | cirrhosis | moderate | |
| D04 | 46, f | iCCA 6, low cholestasis | 15% | moderate | moderate, mild florid | |
| D06 | 77, f | pCCA 7, moderate cholestasis | 5% | partly distinct, also portoportal | mild, also periportal | |
| D07 | 65, f | pCCA, moderate cholangitis and cholestasis | 5% | slight | mild | |
| D08 | 75, m | pCCA, moderate cholestasis | 5% | moderate | mild to moderate, also periportal | |
| Secondary liver tumors (N = 3) | D02 | 74, f | CRLM 8 + CTx 9 | 5% | - | mild |
| D03 | 50, m | CRLM + CTx | NAFLD/AFLD | slight | mild | |
| D01 (control) | 38, f | CRLM | - | low | mild to moderate | |
| Benign liver tumors (N = 5) | D11 (control) | 69, m | Bilioma (infected) | - | highly chronic, partly granulating | moderate florid |
| D12 (control) | 24, f | Adenoma | - | slight | mild | |
| D13 (control) | 41, m | FNH 10 | 5% | slight | mild | |
| D14 (control) | 26, f | Adenoma | 75% | slight to moderate | mild | |
| D15 (control) | 55, f | Hemangioma | 5% | slight | mild |
| Donor ID | Age, Sex | Disease | Medical History |
|---|---|---|---|
| D16 | 59, f 1 | Angiomyolipoma | - |
| D17 | 28, f | FNH 2 | IgA-nephropathy |
| D18 | 28, m 3 | Echinococcosis | - |
| D19 | 45, m | CRLM 4 | ulcerative colitis, s/p 5 colectomy |
| D20 | 39, m | CRLM | recurrence |
| D21 | 64, m | CRLM | CTx 6 |
| D22 | 61, f | Metastasized cervix carcinoma | CTx, steatohepatitis |
| D23 | 46, f | CRLM | CTx, steatohepatitis |
| D24 | 78, m | CRLM | recurrence, CTx |
| D25 | 72, m | pCCA 7 | s/p prostate cancer |
| D26 | 42, f | Hereditary polycystic liver and kidney disease | immunosuppressive therapy |
| D27 | 47, m | Liver abscess | infectious genesis, s/p adenocarcinoma |
| D28 | 49, m | CRLM | in-situ split liver resection, avital tissue |
| D29 | 66, m | Bile duct carcinoma | cholestasis, s/p PVE |
| D30 | 74, m | pCCA | cholangitis |
| D31 | 69, f | pCCA | s/p PVE 8, cholangitis |
| D32 | 77, m | HCC 9 | urothelial carcinoma, obesity, intravesical CTx |
| D33 | 47, f | Caroli disease | cholangitis, s/p acute hepatitis, s/p intraabdominal abscess, obesity |
| Primary Antibodies 1 | ||||||
| Antigen | Macrophage Type | Clone | Isotype, Host Species | Dilution | Incubation Time | Incubation Temperature |
| CD68 | Miscellaneous | KP1 | IgG1, mouse | 1:150 | overnight | 4 °C |
| CD80 | M1 | EPR1157(2) | IgG, rabbit | 1:1000 | overnight | 4 °C |
| CD163 | M2 | OTI2G12 | IgG1, mouse | 1:150 | overnight | 4 °C |
| Secondary Antibodies 2 | ||||||
| Antigen | Conjugate | Clone | Host Species | Dilution | Incubation Time | Incubation Temperature |
| Mouse-IgG | Peroxidase | polyclonal | rabbit | 1:200 | 2 h | RT 3 |
| Rabbit-IgG | Peroxidase | polyclonal | goat | 1:200 | 2 h | RT |
| Conjugated Antibodies 1 | |||||||
|---|---|---|---|---|---|---|---|
| Antigen | Fluorophore | Macrophage Type, Localization | Clone | Isotype | Dilution | Incubation Time | Incubation Temperature |
| CD68 | PE | Miscellaneous, intracellular | REA886 | IgG1 (human recombinant) | 1:50 | 10 min | RT 2 |
| CD86 | APC-Vio700 | M1, extracellular | REA968 | IgG1 (human recombinant) | 1:50 | 10 min | 4 °C |
| CD86 | PerCP-Vio700 | M1, extracellular | FM95 | IgG1κ (mouse) | 1:11 | 10 min | 4 °C |
| CD206 | FITC | M2, extracellular | DCN228 | IgG1κ (mouse) | 1:11 | 10 min | 4 °C |
| Summary | Donor ID | Age, Sex | Disease | Medical History |
|---|---|---|---|---|
| Control (N = 3) | D16 | 59, f 1 | Angiomyolipoma | - |
| D17 | 28, f | FNH 2 | IgA-nephropathy | |
| D18 | 28, m 3 | Echinococcosis | - | |
| Metastase (N = 2) | D19 | 45, m | CRLM 4 | ulcerative colitis, s/p 5 colectomy |
| D20 | 39, m | CRLM | recurrence | |
| Metastase + CTx6 (N = 4) | D21 | 64, m | CRLM | - |
| D22 | 61, f | Metastasized cervix carcinoma | steatohepatitis | |
| D23 | 46, f | CRLM | steatohepatitis | |
| D24 | 78, m | CRLM | recurrence | |
| pCCA7 (N = 1) | D25 | 72, m | Klatskin tumor | s/p prostate cancer |
| PLD8 (N = 1) | D26 | 42, f | Hereditary polycystic liver and kidney disease | immunosuppressive therapy |
| Liver abscess (N = 1) | D27 | 47, m | Liver abscess | infectious genesis, s/p adenocarcinoma |
| ALPPS-OP9 (N = 1) | D28 | 49, m | CRLM | in-situ split liver resection, avital tissue |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimmermann, A.; Hänsel, R.; Gemünden, K.; Kegel-Hübner, V.; Babel, J.; Bläker, H.; Matz-Soja, M.; Seehofer, D.; Damm, G. In Vivo and In Vitro Characterization of Primary Human Liver Macrophages and Their Inflammatory State. Biomedicines 2021, 9, 406. https://doi.org/10.3390/biomedicines9040406
Zimmermann A, Hänsel R, Gemünden K, Kegel-Hübner V, Babel J, Bläker H, Matz-Soja M, Seehofer D, Damm G. In Vivo and In Vitro Characterization of Primary Human Liver Macrophages and Their Inflammatory State. Biomedicines. 2021; 9(4):406. https://doi.org/10.3390/biomedicines9040406
Chicago/Turabian StyleZimmermann, Andrea, René Hänsel, Kilian Gemünden, Victoria Kegel-Hübner, Jonas Babel, Hendrik Bläker, Madlen Matz-Soja, Daniel Seehofer, and Georg Damm. 2021. "In Vivo and In Vitro Characterization of Primary Human Liver Macrophages and Their Inflammatory State" Biomedicines 9, no. 4: 406. https://doi.org/10.3390/biomedicines9040406
APA StyleZimmermann, A., Hänsel, R., Gemünden, K., Kegel-Hübner, V., Babel, J., Bläker, H., Matz-Soja, M., Seehofer, D., & Damm, G. (2021). In Vivo and In Vitro Characterization of Primary Human Liver Macrophages and Their Inflammatory State. Biomedicines, 9(4), 406. https://doi.org/10.3390/biomedicines9040406