
The Potential Role of Dopamine in Mediating Motor Function and Interpersonal Synchrony


{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Motor Functioning and Planning
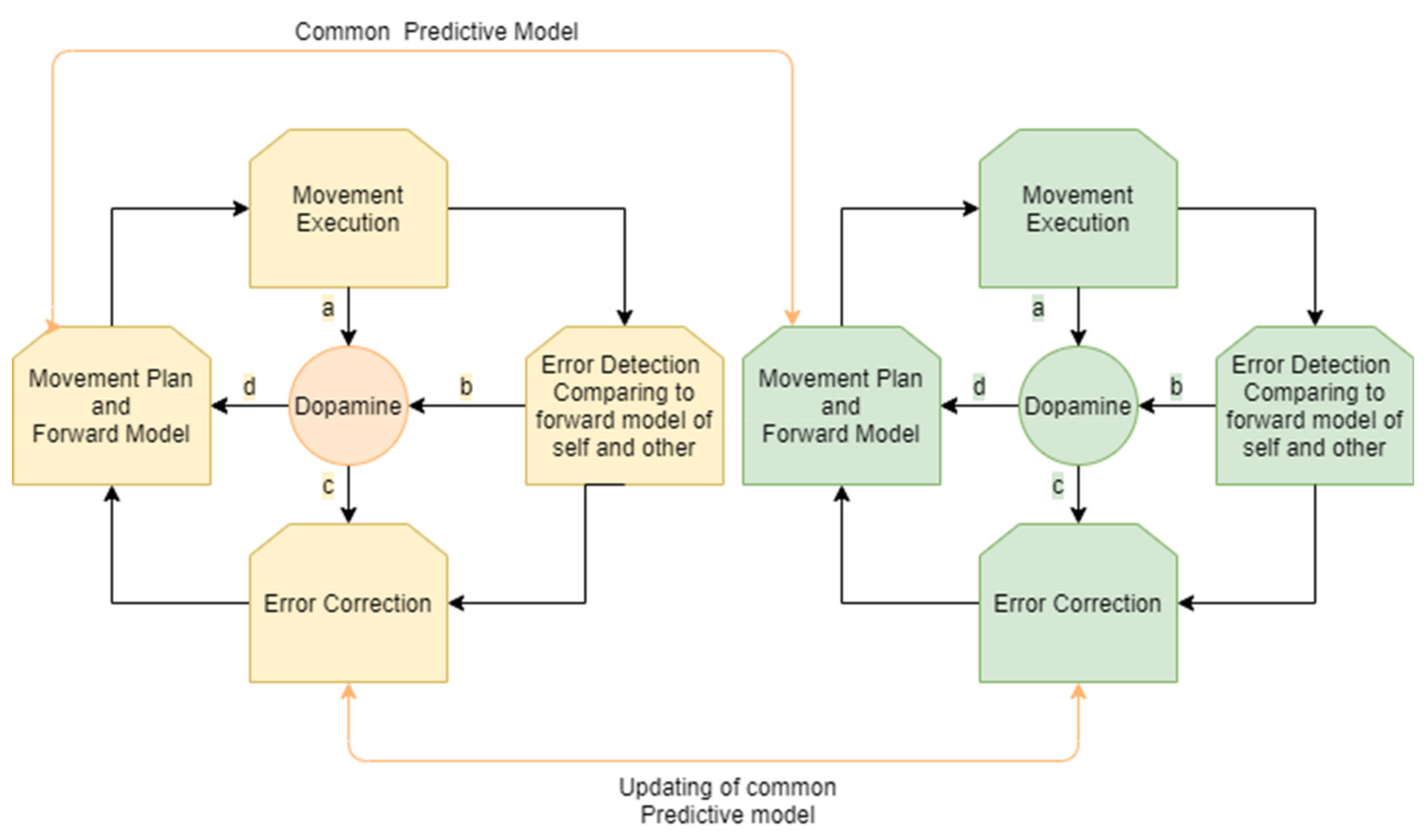
1.2. Interpersonal Synchronization
1.3. Predictive Coding Supports the Link Between Motor and Social Dysfunctions
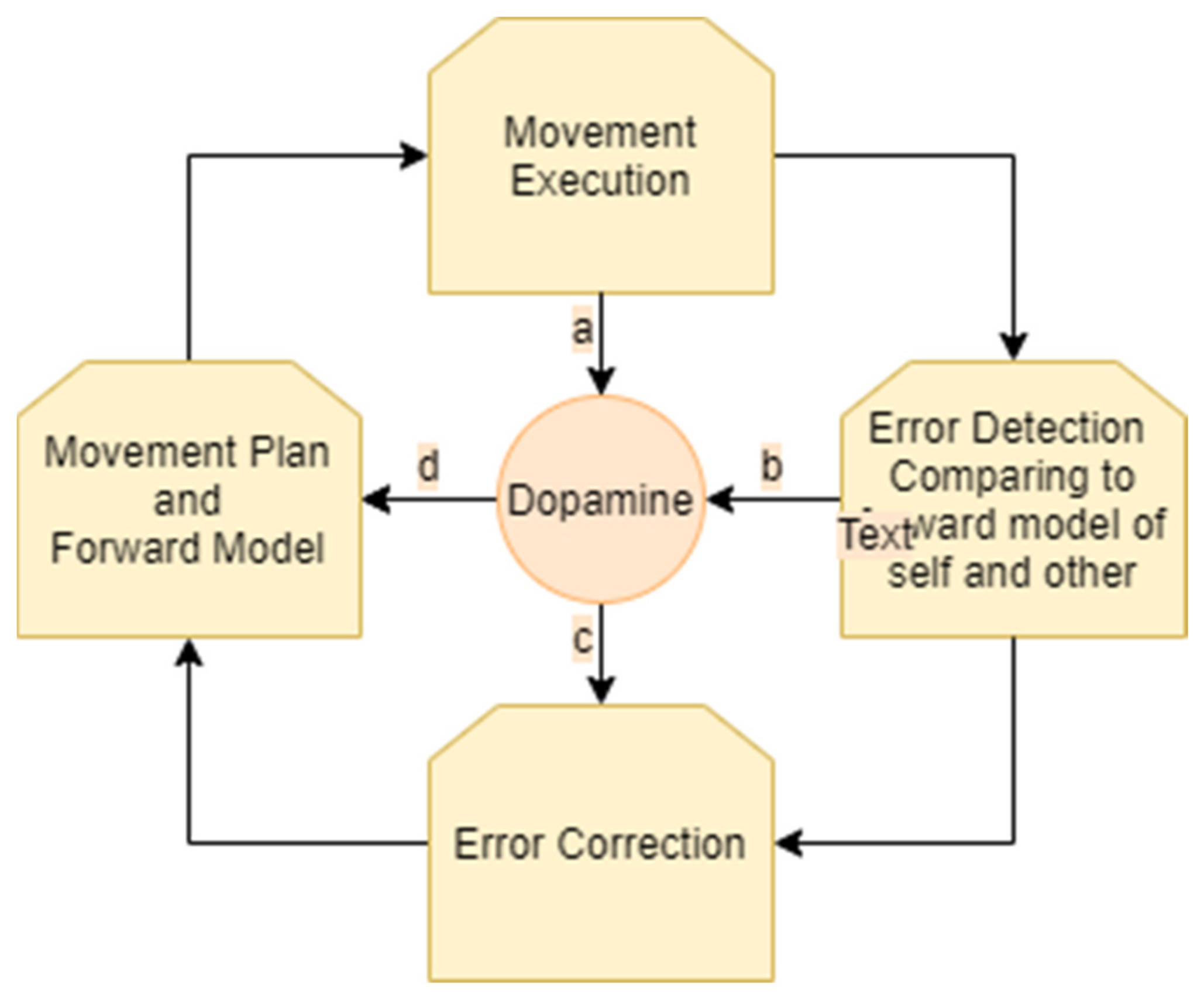
2. Dopamine and the Neural Mechanism That Governs Error Detection
3. Motor and Social Deficiencies in Developmental Disorders That Involve the Dopaminergic System
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Delaherche, E.; Chetouanic, M.; Mahdhaoui, A.; Saint-Georges, C.; Viaux, S.; Cohen, D. Interpersonal Synchrony: A Survey of Evaluation Methods across Disciplines. IEEE Trans. Affect. Comput. 2012, 3, 349–365. Available online: http://ieeexplore.ieee.org/document/6322986/ (accessed on 30 April 2017). [CrossRef]
- Feldman, R. From biological rhythms to social rhythms: Physiological precursors of mother-infant synchrony. Dev. Psychol. 2006, 42, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Leclère, C.; Viaux, S.; Avril, M.; Achard, C.; Chetouani, M.; Missonnier, S.; Cohen, D. Why Synchrony Matters during Mother-Child Interactions: A Systematic Review. PLoS ONE 2014, 9, e113571. [Google Scholar] [CrossRef] [PubMed]
- Sylos-Labini, F.; D’Avella, A.; Lacquaniti, F.; Ivanenko, Y. Human-Human Interaction Forces and Interlimb Coordination During Side-by-Side Walking With Hand Contact. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Varlet, M.; Marin, L.; Lagarde, J.; Bardy, B.G. Social postural coordination. J. Exp. Psychol. Hum. Percept. Perform. 2011, 37, 473–483. [Google Scholar] [CrossRef]
- Mu, Y.; Cerritos, C.; Khan, F. Neural mechanisms underlying interpersonal coordination: A review of hyperscanning research. Soc. Pers. Psychol. Compass 2018, 12, e12421. [Google Scholar] [CrossRef]
- Kaur, M.; Srinivasan, S.M.; Bhat, A.N. Comparing motor performance, praxis, coordination, and interpersonal synchrony betweenchildren with and without Autism Spectrum Disorder (ASD). Res. Dev. Disabil. 2018, 72, 79–95. [Google Scholar] [CrossRef]
- Feldman, R. Sensitive periods in human social development: New insights from research on oxytocin, synchrony, and high-risk parenting. Dev. Psychopathol. 2015, 27, 369–395. [Google Scholar] [CrossRef]
- Feldman, R. Parent? Infant synchrony and the construction of shared timing; physiological precursors, developmental outcomes, and risk conditions. J. Child Psychol. Psychiatry 2007, 48, 329–354. [Google Scholar] [CrossRef]
- Xavier, J.; Tilmont, E.; Bonnot, O. Children’s synchrony and rhythmicity in imitation of peers: Toward a developmental model of empathy. J. Physiol. 2013, 107, 291–297. [Google Scholar] [CrossRef]
- Brezis, R.-S.; Noy, L.; Alony, T.; Gotlieb, R.; Cohen, R.; Golland, Y.; Levit-Binnun, N. Patterns of Joint Improvisation in Adults with Autism Spectrum Disorder. Front. Psychol. 2017, 8. [Google Scholar] [CrossRef]
- Fitzpatrick, P.; Romero, V.; Amaral, J.L.; Duncan, A.; Barnard, H.; Richardson, M.J.; Schmidt, R.C. Evaluating the importance of social motor synchronization and motor skill for understanding autism. Autism Res. 2017, 10, 1687–1699. [Google Scholar] [CrossRef]
- Fulceri, F.; Tonacci, A.; Lucaferro, A.; Apicella, F.; Narzisi, A.; Vincenti, G.; Muratori, F.; Contaldo, A. Interpersonal motor coordination during joint actions in children with and without autism spectrum disorder: The role of motor information. Res. Dev. Disabil. 2018, 80, 13–23. [Google Scholar] [CrossRef]
- Reddan, M.C.; Young, H.; Falkner, J.; López-Solà, M.; Wager, T.D. Touch and social support influence interpersonal synchrony and pain. Soc. Cogn. Affect. Neurosci. 2020, 15, 1064–1075. [Google Scholar] [CrossRef]
- Klein, M.O.; Battagello, D.S.; Cardoso, A.R.; Hauser, D.N.; Bittencourt, J.C.; Correa, R.G. Dopamine: Functions, Signaling, and Association with Neurological Diseases. Cell. Mol. Neurobiol. 2019, 39, 31–59. [Google Scholar] [CrossRef]
- Dahan, A.; Ryder, C.H.; Reiner, M.; Anat, D.; Hanna, R.C.; Miriam, R. Components of Motor Deficiencies in ADHD and Possible Interventions. Neuroscience 2018, 378, 34–53. [Google Scholar] [CrossRef]
- Li, N.; Daie, K.; Svoboda, K.; Druckmann, S. Robust neuronal dynamics in premotor cortex during motorplanning. Nature 2016, 532, 459–464. [Google Scholar] [CrossRef]
- Svoboda, K.; Li, N. Neural mechanisms of movement planning: Motor cortex and beyond. Curr. Opin. Neurobiol. 2018, 49, 33–41. [Google Scholar] [CrossRef]
- Cisek, P.; Kalaska, J.F. Neural Mechanisms for Interacting with a World Full of Action Choices. Annu. Rev. Neurosci. 2010, 33, 269–298. [Google Scholar] [CrossRef]
- Cisek, P.; Kalaska, J.F. Neural Correlates of Reaching Decisions in Dorsal Premotor Cortex: Specification of Multiple Direction Choices and Final Selection of Action. Neuron 2005, 45, 801–814. [Google Scholar] [CrossRef]
- Platt, M.L.; Glimcher, P.W. Responses of Intraparietal Neurons to Saccadic Targets and Visual Distractors. J. Neurophysiol. 1997, 78, 1574–1589. [Google Scholar] [CrossRef]
- Brandi, M.-L.; Wohlschläger, A.; Sorg, C.; Hermsdörfer, J. The Neural Correlates of Planning and Executing Actual Tool Use. J. Neurosci. 2014, 34, 13183–13194. [Google Scholar] [CrossRef]
- Bernier, P.-M.; Cieslak, M.; Grafton, S.T. Effector selection precedes reach planning in the dorsal parietofrontal cortex. J. Neurophysiol. 2012, 108, 57–68. [Google Scholar] [CrossRef]
- Potok, W.; Maskiewicz, A.; Króliczak, G.; Marangon, M. The temporal involvement of the left supramarginal gyrus in planning functional grasps: A neuronavigated TMS study. Cortex 2019, 111, 16–34. [Google Scholar] [CrossRef]
- Buchwald, M.; Przybylski, Ł.; Króliczak, G. Decoding Brain States for Planning Functional Grasps of Tools: A Functional Magnetic Resonance Imaging Multivoxel Pattern Analysis Study. J. Int. Neuropsychol. Soc. 2018, 24, 1013–1025. [Google Scholar] [CrossRef]
- Buccino, G.; Lui, F.; Canessa, N.; Patteri, I.; LaGravinese, G.; Benuzzi, F.; Porro, C.A.; Rizzolatti, G. Neural Circuits Involved in the Recognition of Actions Performed by Nonconspecifics: An fMRI Study. J. Cogn. Neurosci. 2004, 16, 114–126. [Google Scholar] [CrossRef]
- Gehring, W.J.; Goss, B.; Coles, M.G.H.; Meyer, D.E.; Donchin, E. The Error-Related Negativity. Perspect. Psychol. Sci. 2018, 13, 200–204. [Google Scholar] [CrossRef]
- Wolpert, D.M.; Diedrichsen, J.; Flanagan, J.R. Principles of sensorimotor learning. Nat. Rev. Neurosci. 2011, 12, 739–751. [Google Scholar] [CrossRef]
- Wolpert, D.M.; Flanagan, J.R. Motor prediction. Curr. Biol. 2001, 11, R729–R732. [Google Scholar] [CrossRef]
- Adolph, K.E.; Franchak, J.M. The development of motor behavior. Wiley Interdiscip. Rev. Cogn. Sci. 2017, 8, e1430. [Google Scholar] [CrossRef]
- Orban, G.A.; Caruana, F. The neural basis of human tool use. Front. Psychol. 2014, 5, 310. [Google Scholar] [CrossRef] [PubMed]
- Styrkowiec, P.P.; Nowik, A.M.; Króliczak, G. The neural underpinnings of haptically guided functional grasping of tools: An fMRI study. NeuroImage 2019, 194, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Cabaraux, P.; Gandini, J.; Kakei, S.; Manto, M.; Mitoma, H.; Tanaka, H. Dysmetria and Errors in Predictions: The Role of Internal Forward Model. Int. J. Mol. Sci. 2020, 21, 6900. [Google Scholar] [CrossRef] [PubMed]
- Shadmehr, R.; Smith, M.A.; Krakauer, J.W. Error Correction, Sensory Prediction, and Adaptation in Motor Control. Annu. Rev. Neurosci. 2010, 33, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Klichowski, M.; Kroliczak, G. Mental Shopping Calculations: A Transcranial Magnetic Stimulation Study. Front. Psychol. 2020, 11, 1930. [Google Scholar] [CrossRef] [PubMed]
- Yokozuka, T.; Ono, E.; Inoue, Y.; Ogawa, K.-I.; Miyake, Y. The Relationship Between Head Motion Synchronization and Empathy in Unidirectional Face-to-Face Communication. Front. Psychol. 2018, 9. [Google Scholar] [CrossRef]
- Kokal, I.; Engel, A.; Kirschner, S.; Keysers, C. Synchronized Drumming Enhances Activity in the Caudate and Facilitates Prosocial Commitment—If the Rhythm Comes Easily. PLoS ONE 2011, 6, e27272. [Google Scholar] [CrossRef]
- Cirelli, L.K. How interpersonal synchrony facilitates early prosocial behavior. Curr. Opin. Psychol. 2018, 20, 35–39. [Google Scholar] [CrossRef]
- Ramseyer, F.; Tschacher, W. Nonverbal synchrony of head- and body-movement in psychotherapy: Different signals have different associations with outcome. Front. Psychol. 2014, 5, 979. [Google Scholar] [CrossRef]
- Shamay-Tsoory, S.G.; Saporta, N.; Marton-Alper, I.Z.; Gvirts, H.Z. Herding Brains: A Core Neural Mechanism for Social Alignment. Trends Cogn. Sci. 2019, 23, 174–186. [Google Scholar] [CrossRef]
- Hove, M.J.; Risen, J.L. It’s All in the Timing: Interpersonal Synchrony Increases Affiliation. Soc. Cogn. 2009, 27, 949–960. [Google Scholar] [CrossRef]
- Vacharkulksemsuk, T.; Fredrickson, B. Strangers in sync: Achieving embodied rapport through shared movements. J. Exp. Soc. Psychol. 2012, 48, 399–402. [Google Scholar] [CrossRef]
- Launay, J.; Dean, R.T.; Bailes, F. Synchronization Can Influence Trust Following Virtual Interaction. Exp. Psychol. 2013, 60, 53–63. [Google Scholar] [CrossRef]
- Koehne, S.; Hatri, A.; Cacioppo, J.T.; Dziobek, I. Perceived interpersonal synchrony increases empathy: Insights from autism spectrum disorder. Cognition 2016, 146, 8–15. [Google Scholar] [CrossRef]
- Valdesolo, P.; Ouyang, J.; DeSteno, D. The rhythm of joint action: Synchrony promotes cooperative ability. J. Exp. Soc. Psychol. 2010, 46, 693–695. [Google Scholar] [CrossRef]
- MacLean, P.C.; Rynes, K.N.; Aragón, C.; Caprihan, A.; Phillips, J.P.; Lowe, J.R. Mother–infant mutual eye gaze supports emotion regulation in infancy during the Still-Face paradigm. Infant Behav. Dev. 2014, 37, 512–522. [Google Scholar] [CrossRef]
- Bell, H.C. Behavioral Variability in the Service of Constancy. Int. J. Comp. Psychol. 2014, 27, 217–220. [Google Scholar]
- Krabben, K.; Orth, D.; Van Der Kamp, J. Combat as an Interpersonal Synergy: An Ecological Dynamics Approach to Combat Sports. Sports Med. 2019, 49, 1825–1836. [Google Scholar] [CrossRef]
- Blanchard, R.J.; Blanchard, D.; Takahashi, T.; Kelley, M.J. Attack and defensive behaviour in the albino rat. Anim. Behav. 1977, 25, 622–634. [Google Scholar] [CrossRef]
- Baimel, A.; Birch, S.A.; Norenzayan, A. Coordinating bodies and minds: Behavioral synchrony fosters mentalizing. J. Exp. Soc. Psychol. 2018, 74, 281–290. [Google Scholar] [CrossRef]
- Hu, Y.; Hu, Y.; Li, X.; Pan, Y.; Cheng, X. Brain-to-brain synchronization across two persons predicts mutual prosociality. Soc. Cogn. Affect. Neurosci. 2017, 12, 1835–1844. [Google Scholar] [CrossRef] [PubMed]
- Osaka, N.; Minamoto, T.; Yaoi, K.; Azuma, M.; Shimada, Y.M.; Osaka, M. How Two Brains Make One Synchronized Mind in the Inferior Frontal Cortex: fNIRS-Based Hyperscanning During Cooperative Singing. Front. Psychol. 2015, 6, 1–11. [Google Scholar] [CrossRef]
- Nummenmaa, L.; Lahnakoski, J.M.; Glerean, E. Sharing the social world via intersubject neural synchronisation. Curr. Opin. Psychol. 2018, 24, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Chartrand, T.L.; Lakin, J.L. The Antecedents and Consequences of Human Behavioral Mimicry. Annu. Rev. Psychol. 2013, 64, 285–308. [Google Scholar] [CrossRef] [PubMed]
- Gvirts Provolovski, H.Z.; Perlmutter, R. How Can We Prove the Causality of Interbrain Synchronization? Front. Hum. Neurosci. 2021, 15, 651949. [Google Scholar] [CrossRef] [PubMed]
- Asai, T. Know thy agency in predictive coding: Meta-monitoring over forward modeling. Conscious. Cogn. 2017, 51, 82–99. [Google Scholar] [CrossRef]
- Wolpert, D.M.; Miall, R.; Kawato, M. Internal models in the cerebellum. Trends Cogn. Sci. 1998, 2, 338–347. [Google Scholar] [CrossRef]
- Shipp, S.; Adams, R.A.; Friston, K.J. Reflections on agranular architecture: Predictive coding in the motor cortex. Trends Neurosci. 2013, 36, 706–716. [Google Scholar] [CrossRef]
- Monaco, S.; Malfatti, G.; Zendron, A.; Pellencin, E.; Turella, L. Predictive coding of action intentions in dorsal and ventral visual stream is based on visual anticipations, memory-based information and motor preparation. Beiträge und Referate zur Anatomie und Entwickelungsgeschichte 2019, 224, 3291–3308. [Google Scholar] [CrossRef] [PubMed]
- Kilner, J.M.; Friston, K.J.; Frith, C.D. Predictive coding: An account of the mirror neuron system. Cogn. Process. 2007, 8, 159–166. [Google Scholar] [CrossRef]
- Brenner, E.; Smeets, J.B.J. Continuously updating one’s predictions underlies successful interception. J. Neurophysiol. 2018, 120, 3257–3274. [Google Scholar] [CrossRef] [PubMed]
- Gredebäck, G.; Lindskog, M.; Juvrud, J.C.; Green, D.; Marciszko, C. Action Prediction Allows Hypothesis Testing via Internal Forward Models at 6 Months of Age. Front. Psychol. 2018, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Koban, L.; Ramamoorthy, A.; Konvalinka, I. Why do we fall into sync with others? Interpersonal synchronization and the brain’s optimization principle. Soc. Neurosci. 2019, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Heggli, O.A.; Konvalinka, I.; Kringelbach, M.L.; Vuust, P. Musical interaction is influenced by underlying predictive models and musical expertise. Sci. Rep. 2019, 9, 11048. [Google Scholar] [CrossRef]
- Friston, K. Does predictive coding have a future? Nat. Neurosci. 2018, 21, 1019–1021. [Google Scholar] [CrossRef]
- Wolf, J. Predictive coding: Neuroscience and art. In Progress in Brain Research; Elsevier BV: Amsterdam, The Netherlands, 2020; pp. 139–167. [Google Scholar]
- Spratling, M.W. A review of predictive coding algorithms. Brain Cogn. 2017, 112, 92–97. [Google Scholar] [CrossRef]
- Gebauer, L.; Witek, M.A.G.; Hansen, N.C.; Thomas, J.; Konvalinka, I.; Vuust, P. Oxytocin improves synchronisation in leader-follower interaction. Sci. Rep. 2016, 6, 38416. [Google Scholar] [CrossRef]
- Era, V.; Boukarras, S.; Candidi, M. Neural Correlates of Action Monitoring and Mutual Adaptation during Interpersonal Motor Coordination. Phys. Life Rev. 2019, 28, 43–45. [Google Scholar] [CrossRef]
- Schultz, W. Dopamine reward prediction error coding. Dialog Clin. Neurosci. 2016, 18, 23–32. [Google Scholar]
- Bonini, F.; Burle, B.; Liégeois-Chauvel, C.; Régis, J.; Chauvel, P.; Vidal, F. Action Monitoring and Medial Frontal Cortex: Leading Role of Supplementary Motor Area. Science 2014, 343, 888–891. [Google Scholar] [CrossRef]
- Ullsperger, M.; Fischer, A.G.; Nigbur, R.; Endrass, T. Neural mechanisms and temporal dynamics of performance monitoring. Trends Cogn. Sci. 2014, 18, 259–267. [Google Scholar] [CrossRef]
- Fu, Z.; Wu, D.-A.J.; Ross, I.; Chung, J.M.; Mamelak, A.N.; Adolphs, R.; Rutishauser, U. Single-Neuron Correlates of Error Monitoring and Post-Error Adjustments in Human Medial Frontal Cortex. Neuron 2019, 101, 165–177.e5. [Google Scholar] [CrossRef]
- Frank, M.J.; Woroch, B.S.; Curran, T. Error-Related Negativity Predicts Reinforcement Learning and Conflict Biases. Neuron 2005, 47, 495–501. [Google Scholar] [CrossRef]
- Quilodran, R.; Rothé, M.; Procyk, E. Behavioral Shifts and Action Valuation in the Anterior Cingulate Cortex. Neuron 2008, 57, 314–325. [Google Scholar] [CrossRef]
- Bush, G.; Vogt, B.A.; Holmes, J.; Dale, A.M.; Greve, D.; Jenike, M.A.; Rosen, B.R. Dorsal anterior cingulate cortex: A role in reward-based decision making. Proc. Natl. Acad. Sci. USA 2002, 99, 523–528. [Google Scholar] [CrossRef]
- Joch, M.; Hegele, M.; Maurer, H.; Müller, H.; Maurer, L.K. Accuracy of Motor Error Predictions for Different Sensory Signals. Front. Psychol. 2018, 9. [Google Scholar] [CrossRef]
- Burgio-Murphy, A.; Klorman, R.; Shaywitz, S.E.; Fletcher, J.M.; Marchione, K.E.; Holahan, J.; Stuebing, K.K.; Thatcher, J.E.; Shaywitz, B.A. Error-related event-related potentials in children with attention-deficit hyperactivity disorder, oppositional defiant disorder, reading disorder, and math disorder. Biol. Psychol. 2007, 75, 75–86. [Google Scholar] [CrossRef][Green Version]
- Morris, S.E.; Yee, C.M.; Nuechterlein, K.H. Electrophysiological analysis of error monitoring in schizophrenia. J. Abnorm. Psychol. 2006, 115, 239–250. [Google Scholar] [CrossRef]
- Franken, I.H.; Rassin, E.; Muris, P. The assessment of anhedonia in clinical and non-clinical populations: Further validation of the Snaith–Hamilton Pleasure Scale (SHAPS). J. Affect. Disord. 2007, 99, 83–89. [Google Scholar] [CrossRef]
- Vassena, E.; Holroyd, C.B.; Alexander, W.H. Computational Models of Anterior Cingulate Cortex: At the Crossroads between Prediction and Effort. Front. Neurosci. 2017, 11, 316. [Google Scholar] [CrossRef]
- Holroyd, C.B.; Coles, M.G.H. The neural basis of human error processing: Reinforcement learning, dopamine, and the error-related negativity. Psychol. Rev. 2002, 109, 679–709. [Google Scholar] [CrossRef]
- Nachev, P.; Kennard, C.; Husain, M. Functional role of the supplementary and pre-supplementary motor areas. Nat. Rev. Neurosci. 2008, 9, 856–869. [Google Scholar] [CrossRef]
- Jahanshahi, M.; Obeso, I.; Rothwell, J.C.; Obeso, J.A. A fronto–striato–subthalamic–pallidal network for goal-directed and habitual inhibition. Nat. Rev. Neurosci. 2015, 16, 719–732. [Google Scholar] [CrossRef]
- Holroyd, C.B.; Coles, M.G. Dorsal anterior cingulate cortex integrates reinforcement history to guide voluntary behavior. Cortex 2008, 44, 548–559. [Google Scholar] [CrossRef]
- Posner, M.I.; Petersen, S.E.; Fox, P.T.; Raichle, M.E. Localization of cognitive operations in the human brain. Science 1988, 240, 1627–1631. [Google Scholar] [CrossRef]
- Frith, C.D.; Friston, K.; Liddle, P.F.; Frackowiak, R.S.J. Willed action and the prefrontal cortex in man: A study with PET. Proc. Royal. Soc. B: Biol. Sci. 1991, 244, 241–246. [Google Scholar]
- Somerville, L.H.; Heatherton, T.F.; Kelley, W.M. Anterior cingulate cortex responds differentially to expectancy violation and social rejection. Nat. Neurosci. 2006, 9, 1007–1008. [Google Scholar] [CrossRef]
- Chudasama, Y.; Daniels, T.E.; Gorrin, D.P.; Rhodes, S.E.; Rudebeck, P.H.; Murray, E.A. The Role of the Anterior Cingulate Cortex in Choices based on Reward Value and Reward Contingency. Cereb. Cortex 2013, 23, 2884–2898. [Google Scholar] [CrossRef]
- Hadland, K.A.; Rushworth, M.; Gaffan, D.; Passingham, R.E. The Anterior Cingulate and Reward-Guided Selection of Actions. J. Neurophysiol. 2003, 89, 1161–1164. [Google Scholar] [CrossRef]
- Kolling, N.; Behrens, T.; Wittmann, M.; Rushworth, M. Multiple signals in anterior cingulate cortex. Curr. Opin. Neurobiol. 2016, 37, 36–43. [Google Scholar] [CrossRef]
- Mansouri, F.A.; Egner, T.; Buckley, M.J. Monitoring Demands for Executive Control: Shared Functions between Human and Nonhuman Primates. Trends Neurosci. 2017, 40, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Rizzolatti, G.; Craighero, L. The mirror-neuron system. Annu. Rev. Neurosci. 2004, 27, 169–192. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.F.D.C.; Grafton, S.T. Goal representation in human anterior intraparietal sulcus. J Neurosci. 2006, 26, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Frith, C.D.; Frith, U. Interacting minds—A biological basis. Science 1999, 286, 1692–1695. [Google Scholar] [CrossRef]
- Iacoboni, M.; Molnar-Szakacs, I.; Gallese, V.; Buccino, G.; Mazziotta, J.C.; Rizzolatti, G. Grasping the Intentions of Others with One’s Own Mirror Neuron System. PLoS Biol. 2005, 3, e79. [Google Scholar] [CrossRef]
- Kikuchi, T.; Sugiura, M.; Yamamoto, Y.; Sasaki, Y.; Hanawa, S.; Sakuma, A.; Matsumoto, K.; Matsuoka, H.; Kawashima, R. Neural responses to action contingency error in different cortical areas are attributable to forward prediction or sensory processing. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Shergill, S.S.; White, T.; Joyce, D.; Bays, P.M.; Wolpert, D.M.; Frith, C. Modulation of somatosensory processing by action. NeuroImage 2013, 70, 356–362. [Google Scholar] [CrossRef]
- Chao, H.H.; Luo, X.; Chang, J.L.; Li, C.-S.R. Activation of the pre-supplementary motor area but not inferior prefrontal cortex in association with short stop signal reaction time—An intra-subject analysis. BMC Neurosci. 2009, 10, 75. [Google Scholar] [CrossRef]
- Duann, J.-R.; Ide, J.S.; Luo, X.; Li, C.-S.R. Functional connectivity delineates distinct roles of the inferior frontal cortex and presupplementary motor area in stop signal inhibition. J. Neurosci. 2009, 29, 10171–10179. [Google Scholar] [CrossRef]
- Seidler, R.D.; Kwak, Y.; Fling, B.W.; Bernard, J.A. Neurocognitive mechanisms of error-based motor learning. Adv. Exp. Med. Biol. 2013, 782, 39–60. [Google Scholar]
- Stuphorn, V.; Brown, J.W.; Schall, J.D. Role of Supplementary Eye Field in Saccade Initiation: Executive, Not Direct, Control. J. Neurophysiol. 2010, 103, 801–816. [Google Scholar] [CrossRef]
- Chen, X.; Scangos, K.W.; Stuphorn, V. Supplementary Motor Area Exerts Proactive and Reactive Control of Arm Movements. J. Neurosci. 2010, 30, 14657–14675. [Google Scholar] [CrossRef]
- Keller, P.E.; Novembre, G.; Hove, M.J. Rhythm in joint action: Psychological and neurophysiological mechanisms for real-time interpersonal coordination. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130394. [Google Scholar] [CrossRef]
- Vicaria, I.M.; Dickens, L. Meta-Analyses of the Intra- and Interpersonal Outcomes of Interpersonal Coordination. J. Nonverbal Behav. 2016, 40, 335–361. [Google Scholar] [CrossRef]
- Wang, Y.; Ramsey, R.; Hamilton, A.F.D.C. The Control of Mimicry by Eye Contact Is Mediated by Medial Prefrontal Cortex. J. Neurosci. 2011, 31, 12001–12010. [Google Scholar] [CrossRef]
- Goldenberg, G.; Karnath, H.-O. The Neural Basis of Imitation is Body Part Specific. J. Neurosci. 2006, 26, 6282–6287. [Google Scholar] [CrossRef]
- Caspers, S.; Zilles, K.; Laird, A.R.; Eickhoff, S.B. ALE meta-analysis of action observation and imitation in the human brain. NeuroImage 2010, 50, 1148–1167. [Google Scholar] [CrossRef]
- Sasaki, A.T.; Okamoto, Y.; Kochiyama, T.; Kitada, R.; Sadato, N. Distinct sensitivities of the lateral prefrontal cortex and extrastriate body area to contingency between executed and observed actions. Cortex 2018, 108, 234–251. [Google Scholar] [CrossRef]
- Jiang, J.; Dai, B.; Peng, D.; Zhu, C.; Liu, L.; Lu, C. Neural Synchronization during Face-to-Face Communication. J. Neurosci. 2012, 32, 16064–16069. [Google Scholar] [CrossRef]
- Jasmin, K.M.; Mcgettigan, C.; Agnew, Z.K.; LaVan, N.; Josephs, O.; Cummins, F.; Scott, S.K. Cohesion and Joint Speech: Right Hemisphere Contributions to Synchronized Vocal Production. J. Neurosci. 2016, 36, 4669–4680. [Google Scholar] [CrossRef]
- Gvirts, H.Z.; Perlmutter, R. What Guides Us to Neurally and Behaviorally Align With Anyone Specific? A Neurobiological Model Based on fNIRS Hyperscanning Studies. Neuroscience 2020, 26, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Steinman, M.Q.; Duque-Wilckens, N.; Trainor, B.C. Complementary Neural Circuits for Divergent Effects of Oxytocin: Social Approach Versus Social Anxiety. Biol. Psychiatry 2019, 85, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Shamay-Tsoory, S.G.; Abu-Akel, A. The Social Salience Hypothesis of Oxytocin. Biol. Psychiatry 2016, 79, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Wiltermuth, S. Synchrony and destructive obedience. Soc. Influ. 2012, 7, 78–89. [Google Scholar] [CrossRef]
- Roca, M.; Torralva, T.; Gleichgerrcht, E.; Chade, A.; Arévalo, G.G.; Gershanik, O.; Manes, F. Impairments in Social Cognition in Early Medicated and Unmedicated Parkinson Disease. Cogn. Behav. Neurol. 2010, 23, 152–158. [Google Scholar] [CrossRef]
- Lloyd, M.; Macdonald, M.; Lord, C. Motor skills of toddlers with autism spectrum disorders. Autism 2013, 17, 133–146. [Google Scholar] [CrossRef]
- Kenny, L.; Hill, E.; Hamilton, A.F.D.C. The Relationship between Social and Motor Cognition in Primary School Age-Children. Front. Psychol. 2016, 7. [Google Scholar] [CrossRef][Green Version]
- Marsh, K.L.; Isenhower, R.W.; Richardson, M.J.; Helt, M.; Verbalis, A.D.; Schmidt, R.C.; Fein, D. Autism and social disconnection in interpersonal rocking. Front. Integr. Neurosci. 2013, 7, 4. [Google Scholar] [CrossRef]
- Fitzpatrick, P.; Frazier, J.A.; Cochran, D.M.; Mitchell, T.; Coleman, C.; Schmidt, R.C. Impairments of Social Motor Synchrony Evident in Autism Spectrum Disorder. Front. Psychol. 2016, 7, 1323. [Google Scholar] [CrossRef]
- Scharoun, S.; Bryden, P.; Otipkova, Z.; Musalek, M.; Lejcarova, A. Motor skills in Czech children with attention-deficit/hyperactivity disorder and their neurotypical counterparts. Res. Dev. Disabil. 2013, 34, 4142–4153. [Google Scholar] [CrossRef]
- Brossard-Racine, M.; Shevell, M.; Snider, L.; Bélanger, S.A.; Majnemer, A. Motor skills of children newly diagnosed with Attention Deficit Hyperactivity Disorder prior to and following treatment with stimulant medication. Res. Dev. Disabil. 2012, 33, 2080–2087. [Google Scholar] [CrossRef]
- Fenollar-Cortés, J.; Gallego-Martínez, A.; Fuentes, L.J. The role of inattention and hyperactivity/impulsivity in the fine motor coordination in children with ADHD. Res. Dev. Disabil. 2017, 69, 77–84. [Google Scholar] [CrossRef]
- Carte, E.T.; Nigg, J.T.; Hinshaw, S.P. Neuropsychological functioning, motor speed, and language processing in boys with and without ADHD. J. Abnor. Child Psychol. 1996, 24, 481–498. [Google Scholar] [CrossRef]
- Harvey, W.J.; Reid, G.; Bloom, G.A.; Staples, K.; Grizenko, N.; Mbekou, V.; Ter-Stepanian, M.; Joober, R. Physical activity experiences of boys with and without ADHD. Adapt. Phys. Act. Q. 2009, 26, 131–150. [Google Scholar] [CrossRef]
- Sweeney, K.L.; Ryan, M.; Schneider, H.; Ferenc, L.; Denckla, M.B.; Mahone, E.M. Developmental Trajectory of Motor Deficits in Preschool Children with ADHD. Dev. Neuropsychol. 2018, 43, 419–429. [Google Scholar] [CrossRef]
- Papadopoulos, N.; Stavropoulos, V.; McGinley, J.; Bellgrove, M.; Tonge, B.; Murphy, A.; Cornish, K.; Rinehart, N. Moderating Effect of Motor Proficiency on the Relationship Between ADHD Symptoms and Sleep Problems in Children With Attention Deficit Hyperactivity Disorder–Combined Type. Behav. Sleep Med. 2018, 17, 646–656. [Google Scholar] [CrossRef]
- Kaiser, M.-L.; Schoemaker, M.; Albaret, J.-M.; Geuze, R. What is the evidence of impaired motor skills and motor control among children with attention deficit hyperactivity disorder (ADHD)? Systematic review of the literature. Res. Dev. Disabil. 2015, 36, 338–357. [Google Scholar] [CrossRef]
- Nijmeijer, J.S.; Minderaa, R.B.; Buitelaar, J.K.; Mulligan, A.; Hartman, C.A.; Hoekstra, P.J. Attention-deficit/hyperactivity disorder and social dysfunctioning. Clin. Psychol. Rev. 2008, 28, 692–708. [Google Scholar] [CrossRef]
- McQuade, J.D.; Hoza, B. Peer problems in Attention Deficit Hyperactivity Disorder: Current status and future directions. Dev. Disabil. Res. Rev. 2008, 14, 320–324. [Google Scholar] [CrossRef]
- Gardner, D.M.; Gerdes, A.C. A Review of Peer Relationships and Friendships in Youth With ADHD. J. Atten. Disord. 2015, 19, 844–855. [Google Scholar] [CrossRef]
- Rokeach, A.; Wiener, J. Friendship Quality in Adolescents With ADHD. J. Atten. Disord. 2017, 24, 1156–1168. [Google Scholar] [CrossRef]
- Young, S.; Toone, B.; Tyson, C. Comorbidity and psychosocial profile of adults with Attention Deficit Hyperactivity Disorder. Pers. Individ. Differ. 2003, 35, 743–755. [Google Scholar] [CrossRef]
- Kooij, S.J.; Bejerot, S.; Blackwell, A.; Caci, H.; Casas-Brugué, M.; Carpentier, P.J.; Edvinsson, D.; Fayyad, J.; Foeken, K.; Fitzgerald, M.; et al. European consensus statement on diagnosis and treatment of adult ADHD: The European Network Adult ADHD. BMC Psychiatry 2010, 10, 67. [Google Scholar] [CrossRef]
- Philipsen, A.; Feige, B.; Hesslinger, B.; Scheel, C.; Ebert, D.; Matthies, S.; Limberger, M.F.; Kleindienst, N.; Bohus, M.; Lieb, K. Borderline typical symptoms in adult patients with attention deficit/hyperactivity disorder. ADHD Atten. Deficit Hyperact. Disord. 2009, 1, 11–18. [Google Scholar] [CrossRef]
- Eakin, L.; Minde, K.; Hechtman, L.; Ochs, E.; Krane, E.; Bouffard, R.; Greenfield, B.; Looper, K. The marital and family functioning of adults with ADHD and their spouses. J. Atten. Disord. 2004, 8, 1–10. [Google Scholar] [CrossRef]
- Uekermann, J.; Kraemer, M.; Abdel-Hamid, M.; Schimmelmann, B.G.; Hebebrand, J.; Daum, I.; Wiltfang, J.; Kis, B. Social cognition in attention-deficit hyperactivity disorder (ADHD). Neurosci. Biobehav. Rev. 2010, 34, 734–743. [Google Scholar] [CrossRef]
- Leong, V.; Schilbach, L. The promise of two-person neuroscience for developmental psychiatry: Using interaction-based sociometrics to identify disorders of social interaction. Br. J. Psychiatry 2019, 215, 636–638. [Google Scholar] [CrossRef]
- Maoz, H.; Gvirts, H.Z.; Sheffer, M.; Bloch, Y. Theory of Mind and Empathy in Children With ADHD. J. Atten. Disord. 2017, 23, 1331–1338. [Google Scholar] [CrossRef]
- Shachar, O.L.; Gvirts, H.Z.; Goldwin, Y.; Bloch, Y.; Shamay-Tsoory, S.; Zagoory-Sharon, O.; Feldman, R.; Maoz, H. The effect of methylphenidate on social cognition and oxytocin in children with attention deficit hyperactivity disorder. Neuropsychopharmacology 2020, 45, 367–373. [Google Scholar] [CrossRef]
- Pineda-Alhucema, W.; Aristizabal, E.; Escudero-Cabarcas, J.; Acosta-López, J.E.; Vélez, J.I. Executive Function and Theory of Mind in Children with ADHD: A Systematic Review. Neuropsychol. Rev. 2018, 28, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Bora, E.; Pantelis, C. Meta-analysis of social cognition in attention-deficit/hyperactivity disorder (ADHD): Comparison with healthy controls and autistic spectrum disorder. Psychol. Med. 2015, 46, 699–716. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hamid, M.; Niklewski, F.; Heßmann, P.; Guberina, N.; Kownatka, M.; Kraemer, M.; Scherbaum, N.; Dziobek, I.; Bartels, C.; Wiltfang, J.; et al. Impaired empathy but no theory of mind deficits in adult attention deficit hyperactivity disorder. Brain Behav. 2019, 9, e01401. [Google Scholar] [CrossRef] [PubMed]
- Varlet, M.; Marin, L.; Raffard, S.; Schmidt, R.C.; Capdevielle, D.; Boulenger, J.-P.; Del-Monte, J.; Bardy, B.G. Impairments of Social Motor Coordination in Schizophrenia. PLoS ONE 2012, 7, e29772. [Google Scholar] [CrossRef]
- Lavelle, M.; Healey, P.G.; McCabe, R. Nonverbal Behavior During Face-to-face Social Interaction in Schizophrenia. J. Nerv. Ment. Dis. 2014, 202, 47–54. [Google Scholar] [CrossRef]
- Marsh, K.L.; Richardson, M.J.; Schmidt, R.C. Social Connection Through Joint Action and Interpersonal Coordination. Top. Cogn. Sci. 2009, 1, 320–339. [Google Scholar] [CrossRef]
- Healey, D.M.; Gopin, C.B.; Grossman, B.R.; Campbell, S.B.; Halperin, J.M. Mother-child dyadic synchrony is associated with better functioning in hyperactive/inattentive preschool children. J. Child Psychol. Psychiatry 2010, 51, 1058–1066. [Google Scholar] [CrossRef]
- Gvirts Problovski, H.Z.; Lavi, D.; Yozevitch, R.; Sherman, M.; Hagay, Y.; Dahan, A. Impairments of interpersonal synchrony evident in attention deficit hyperactivity disorder (ADHD). Acta Psychol. 2021, 212, 103210. [Google Scholar] [CrossRef]
- Molina, B.S.; Hinshaw, S.P.; Swanson, J.M.; Arnold, L.E.; Vitiello, B.; Jensen, P.S.; Epstein, J.N.; Hoza, B.; Hechtman, L.; Abikoff, H.B.; et al. The MTA at 8 Years: Prospective Follow-up of Children Treated for Combined-Type ADHD in a Multisite Study. J. Am. Acad. Child Adolesc. Psychiatry 2009, 48, 484–500. [Google Scholar] [CrossRef]
- Shaw, M.; Hodgkins, P.; Caci, H.; Young, S.; Kahle, J.; Woods, A.G.; Arnold, L.E. A systematic review and analysis of long-term outcomes in attention deficit hyperactivity disorder: Effects of treatment and non-treatment. BMC Med. 2012, 10, 99. [Google Scholar] [CrossRef]
- Maoz, H.; Tsviban, L.; Gvirts, H.Z.; Shamay-Tsoory, S.G.; Levkovitz, Y.; Watemberg, N.; Bloch, Y. Stimulants improve theory of mind in children with attention deficit/hyperactivity disorder. J. Psychopharmacol. 2014, 28, 212–219. [Google Scholar] [CrossRef]
- Golubchik, P.; Weizman, A. The Possible Effect of Methylphenidate Treatment on Empathy in Children Diagnosed with Attention-Deficit/Hyperactivity Disorder, Both With and Without Comorbid Oppositional Defiant Disorder. J. Child Adolesc. Psychopharmacol. 2017, 27, 429–432. [Google Scholar] [CrossRef]
- Jacobi-Polishook, T.; Shorer, Z.; Melzer, I. The effect of methylphenidate on postural stability under single and dual task conditions in children with attention deficit hyperactivity disorder—A double blind randomized control trial. J. Neurol. Sci. 2009, 280, 15–21. [Google Scholar] [CrossRef]
- Leitner, Y.; Barak, R.; Giladi, N.; Peretz, C.; Eshel, R.; Gruendlinger, L.; Hausdorff, J.M. Gait in attention deficit hyperactivity disorder. J. Neurol. 2007, 254, 1330–1338. [Google Scholar] [CrossRef]
- Kim, H.-W.; Joung, Y.-S.; Park, K.J. 1.47 Effect of Pharmacological Treatment for Attention-Deficit/Hyperactivity Disorder (ADHD) on Motor Coordination. J. Am. Acad. Child Adolesc. Psychiatry 2017, 56, S168. [Google Scholar] [CrossRef]
- Flapper, B.C.; Houwen, S.; Schoemaker, M.M. Fine motor skills and effects of methylphenidate in children with attention-deficit-hyperactivity disorder and developmental coordination disorder. Dev. Med. Child Neurol. 2006, 48, 165–169. [Google Scholar] [CrossRef]
- Bart, O.; Podoly, T.; Bar-Haim, Y. A preliminary study on the effect of methylphenidate on motor performance in children with comorbid DCD and ADHD. Res. Dev. Disabil. 2010, 31, 1443–1447. [Google Scholar] [CrossRef]
- Smith, R.; Badcock, P.; Friston, K.J. Recent advances in the application of predictive coding and active inference models within clinical neuroscience. Psychiatry Clin. Neurosci. 2021, 75, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Parr, T.; Friston, K.J. The Anatomy of Inference: Generative Models and Brain Structure. Front. Comput. Neurosci. 2018, 12, 90. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Miao, S.; Han, J.; Zeng, K.; Ouyang, G.; Yang, J.; Li, X. Complexity analysis of fNIRS signals in ADHD children during working memory task. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Diederen, K.M.J.; Fletcher, P.C. Dopamine, Prediction Error and Beyond. Neuroscientist 2021, 27, 30–46. [Google Scholar] [CrossRef] [PubMed]
- Scopa, C.; Palagi, E. Mimic me while playing! Social tolerance and rapid facial mimicry in macaques (Macaca tonkeana and Macaca fuscata). J. Comp. Psychol. 2016, 130, 153–161. [Google Scholar] [CrossRef]
- Palagi, E.; Bertini, M.; Annicchiarico, G.; Cordoni, G. Mirror replication of sexual facial expressions increases the success of sexual contacts in bonobos. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Feniger-Schaal, R.; Noy, L.; Hart, Y.; Koren-Karie, N.; Mayo, A.E.; Alon, U. Would you like to play together? Adults’ attachment and the mirror game. Attach. Hum. Dev. 2015, 18, 33–45. [Google Scholar] [CrossRef]
- Behrends, A.; Müller, S.; Dziobek, I. Moving in and out of synchrony. A concept for a new intervention fostering empathy through interactional movement and dance. Arts Psychother. 2012, 2, 107–116. [Google Scholar] [CrossRef]
- Steinbeis, N. The role of self-other distinction in understanding others’ mental and emotional states: Neurocognitive mechanisms in children and adults. Philos. Trans. Royal Soc. B Biol. Sci. 2016, 1686, 20150074. [Google Scholar] [CrossRef]
- Powers, W.T. Behavior: The Control of Perception; Aldine: Chicago, IL, USA, 1973. [Google Scholar]
- Dereli, E.; Karakuş, Ö. An Examination of Attachment Styles and Social Skills of University Students. Electron. J. Res. Educ. Psychol. 2017, 9. [Google Scholar] [CrossRef][Green Version]
- Noy, L.; Dekel, E.; Alon, U. The mirror game as a paradigm for studying the dynamics of two people improvising motion together. Proc. Nat. Acad. Sci. USA 2011, 108, 20947–20952. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gvirts Probolovski, H.Z.; Dahan, A. The Potential Role of Dopamine in Mediating Motor Function and Interpersonal Synchrony. Biomedicines 2021, 9, 382. https://doi.org/10.3390/biomedicines9040382
Gvirts Probolovski HZ, Dahan A. The Potential Role of Dopamine in Mediating Motor Function and Interpersonal Synchrony. Biomedicines. 2021; 9(4):382. https://doi.org/10.3390/biomedicines9040382
Chicago/Turabian StyleGvirts Probolovski, Hila Z., and Anat Dahan. 2021. "The Potential Role of Dopamine in Mediating Motor Function and Interpersonal Synchrony" Biomedicines 9, no. 4: 382. https://doi.org/10.3390/biomedicines9040382
APA StyleGvirts Probolovski, H. Z., & Dahan, A. (2021). The Potential Role of Dopamine in Mediating Motor Function and Interpersonal Synchrony. Biomedicines, 9(4), 382. https://doi.org/10.3390/biomedicines9040382