
Effects of Chronic Oral Probiotic Treatment in Paclitaxel-Induced Neuropathic Pain


 , ,
, ,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
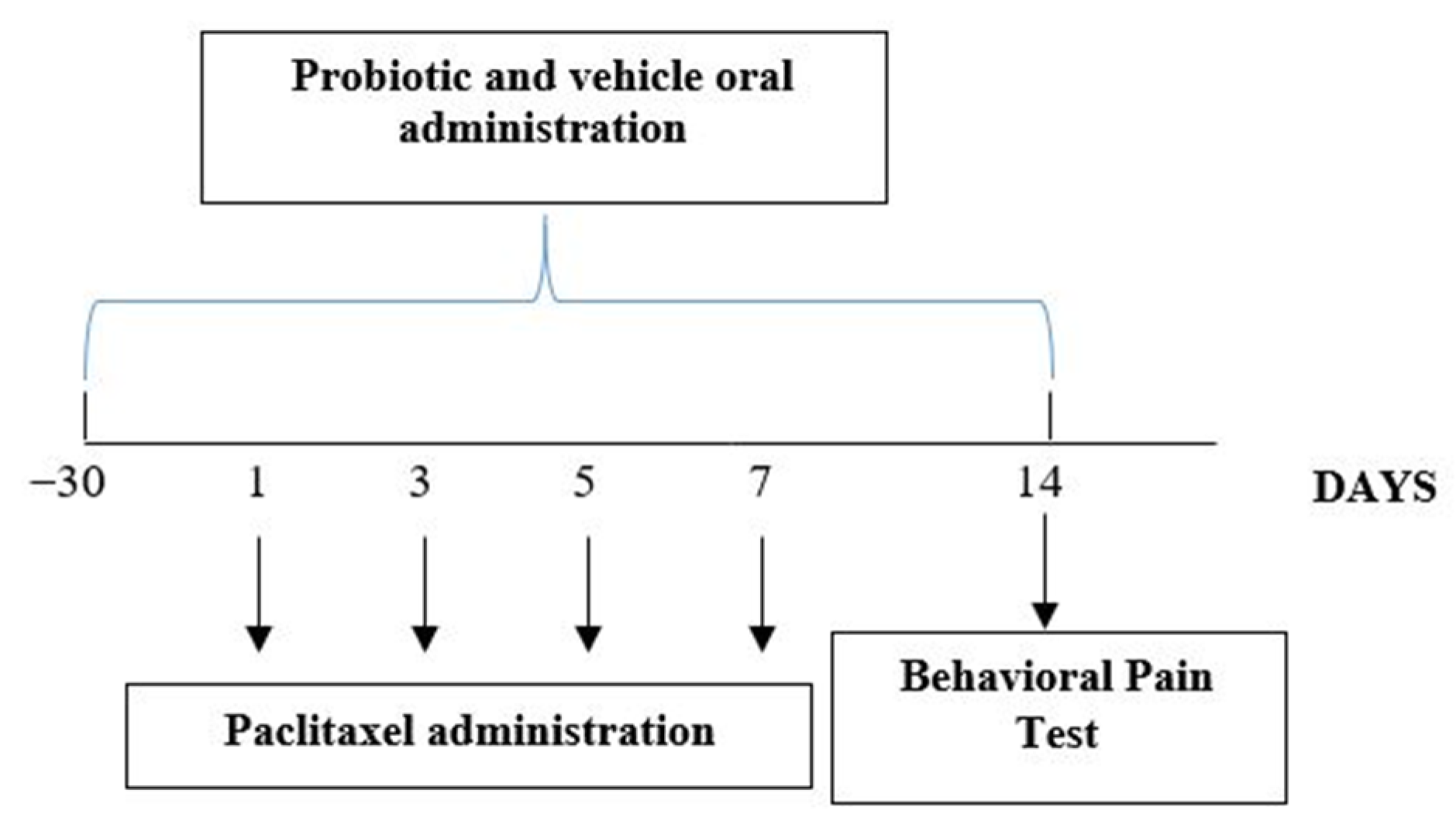
2.2. Experimental Groups and Procedures
- Sham Mice treated with Vehicle (n = 10),
- Sham Mice treated with probiotics (n = 10),
- Mice treated with Paclitaxel + Vehicle (n = 10),
- Mice treated with Paclitaxel + probiotics (n = 10)
2.3. Drug Treatment
2.4. Behavioral Pain Tests
2.4.1. Mechanical Allodynia (Von Frey Test)
2.4.2. Mechanical Hyperalgesia (Randall-Sellitto Test)
2.5. Ex Vivo Experiments
2.5.1. Serum TNF-α, IL-1β, and IL-6 Determination
2.5.2. Western Blot Analysis
2.5.3. Immunohistochemistry Analyses
2.5.4. Statistical Analyses
3. Results
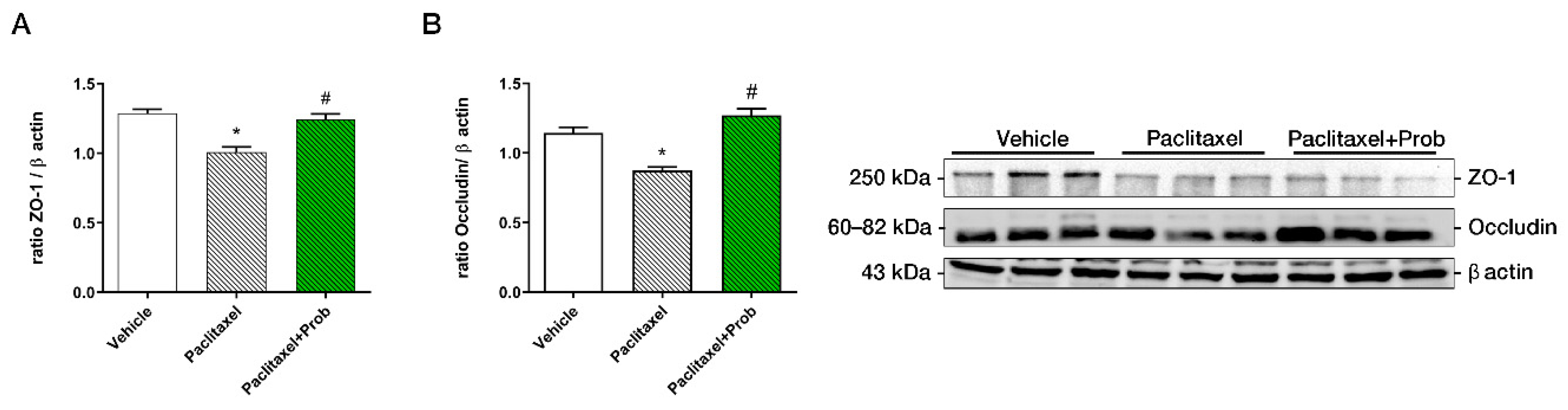
3.1. Probiotic Treatment Reduces Colon Tissue Damage
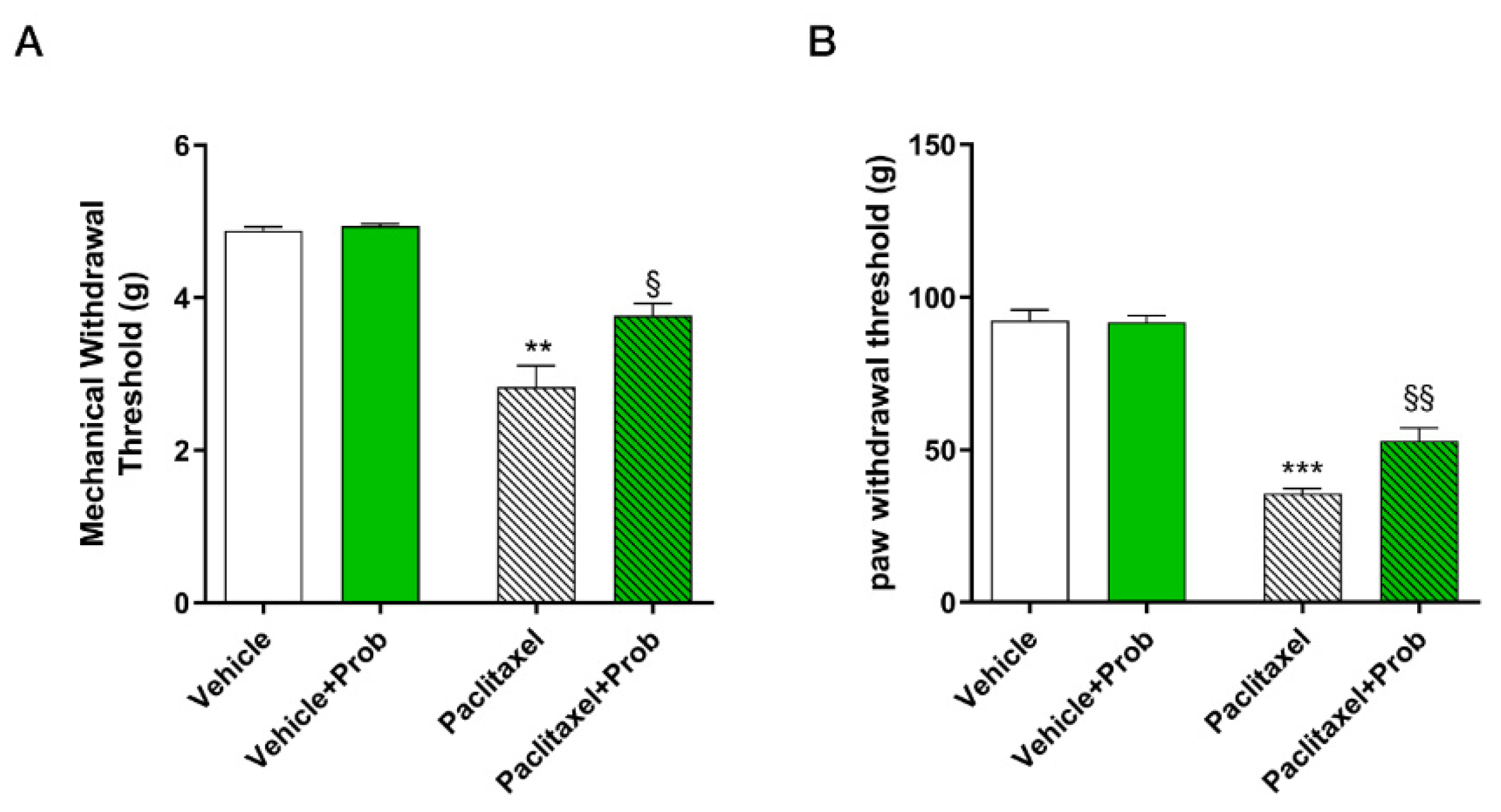
3.2. Effect of Probiotic on Paclitaxel-Induced Mechanical Allodynia and Hyperalgesia
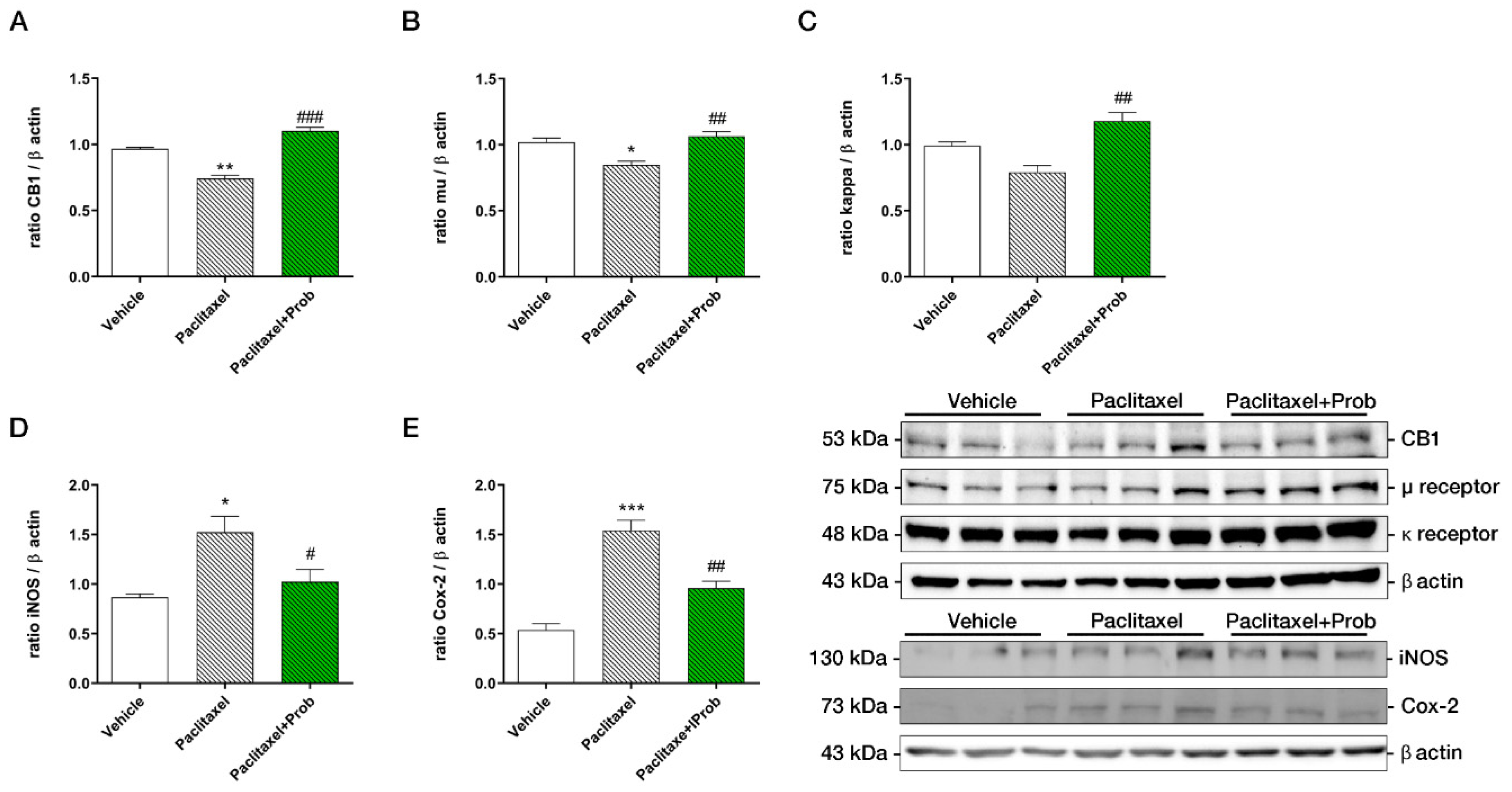
3.3. Probiotic Treatment Modulates CB-1, µ, and ĸ Receptors and Pro-Inflammatory Proteins Expression in the Spinal Cord
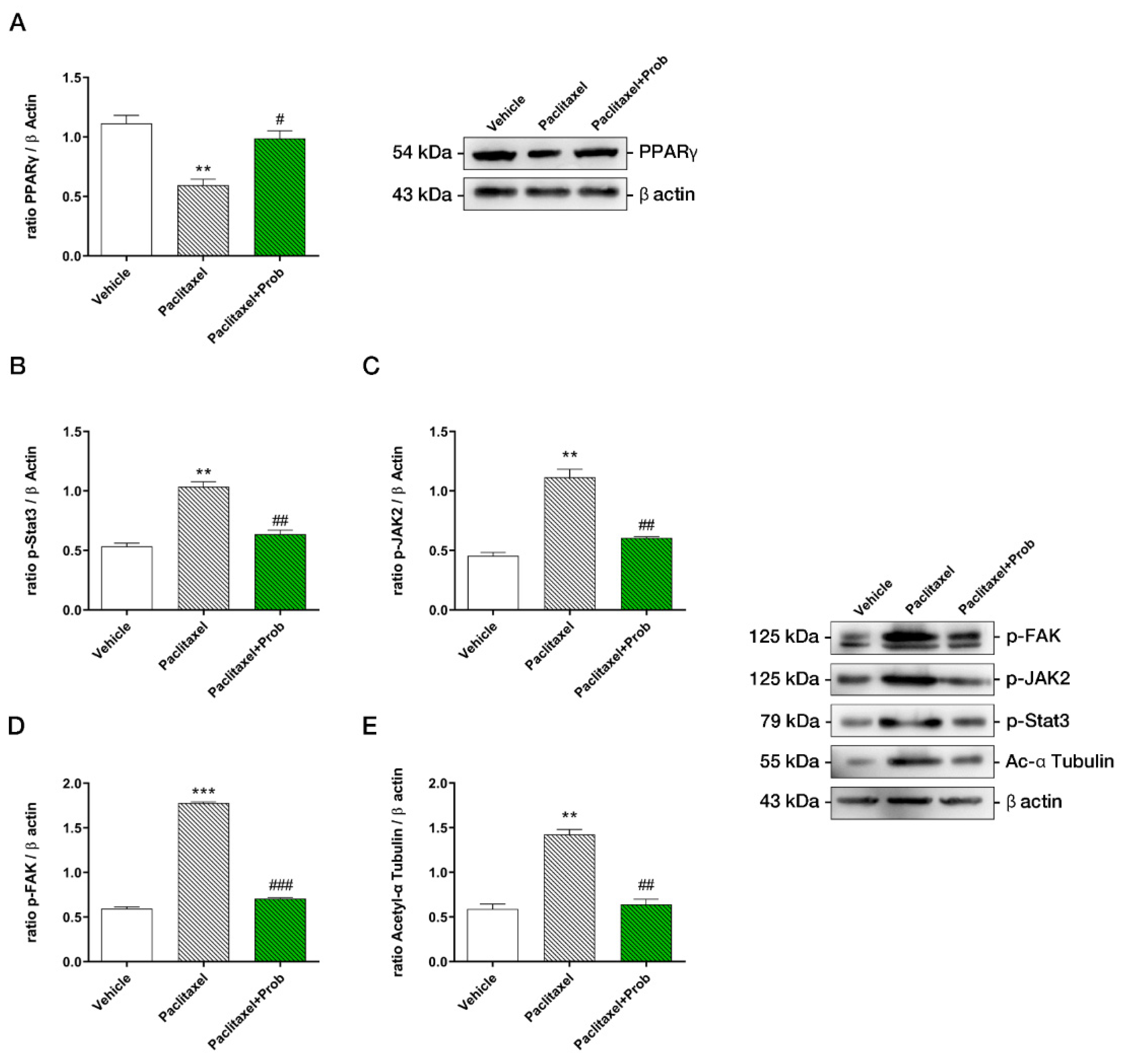
3.4. Probiotic Treatment Counteracts the Increase of Neuropathic Pain Proteins
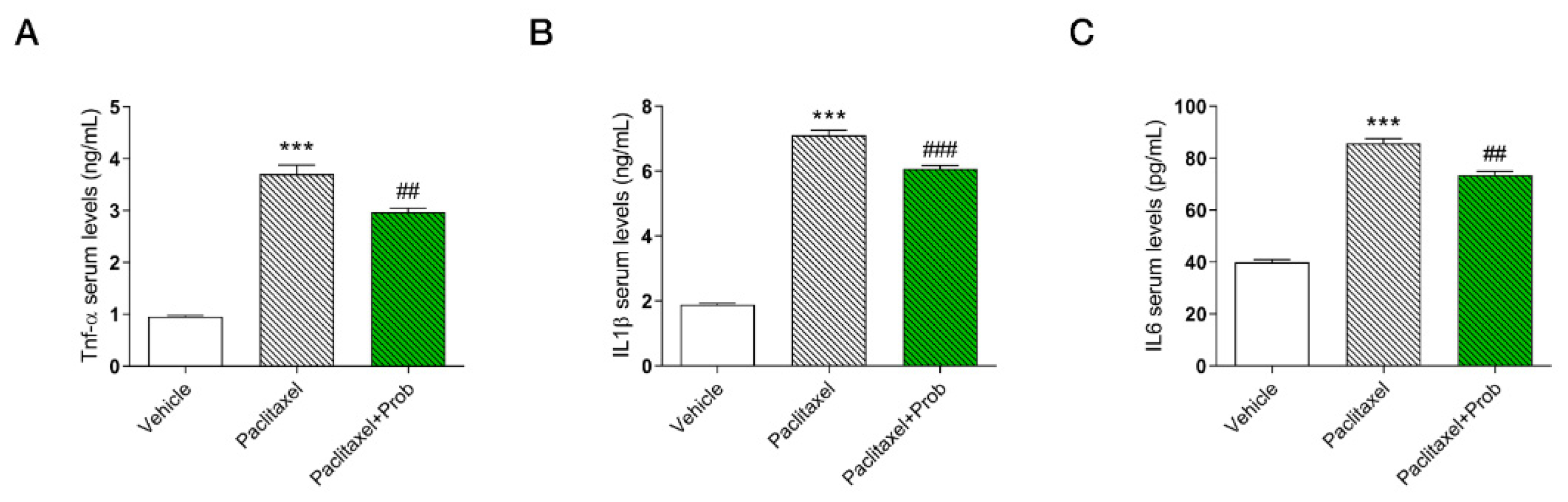
3.5. Effects of the Probiotics on Inflammatory Markers in Serum
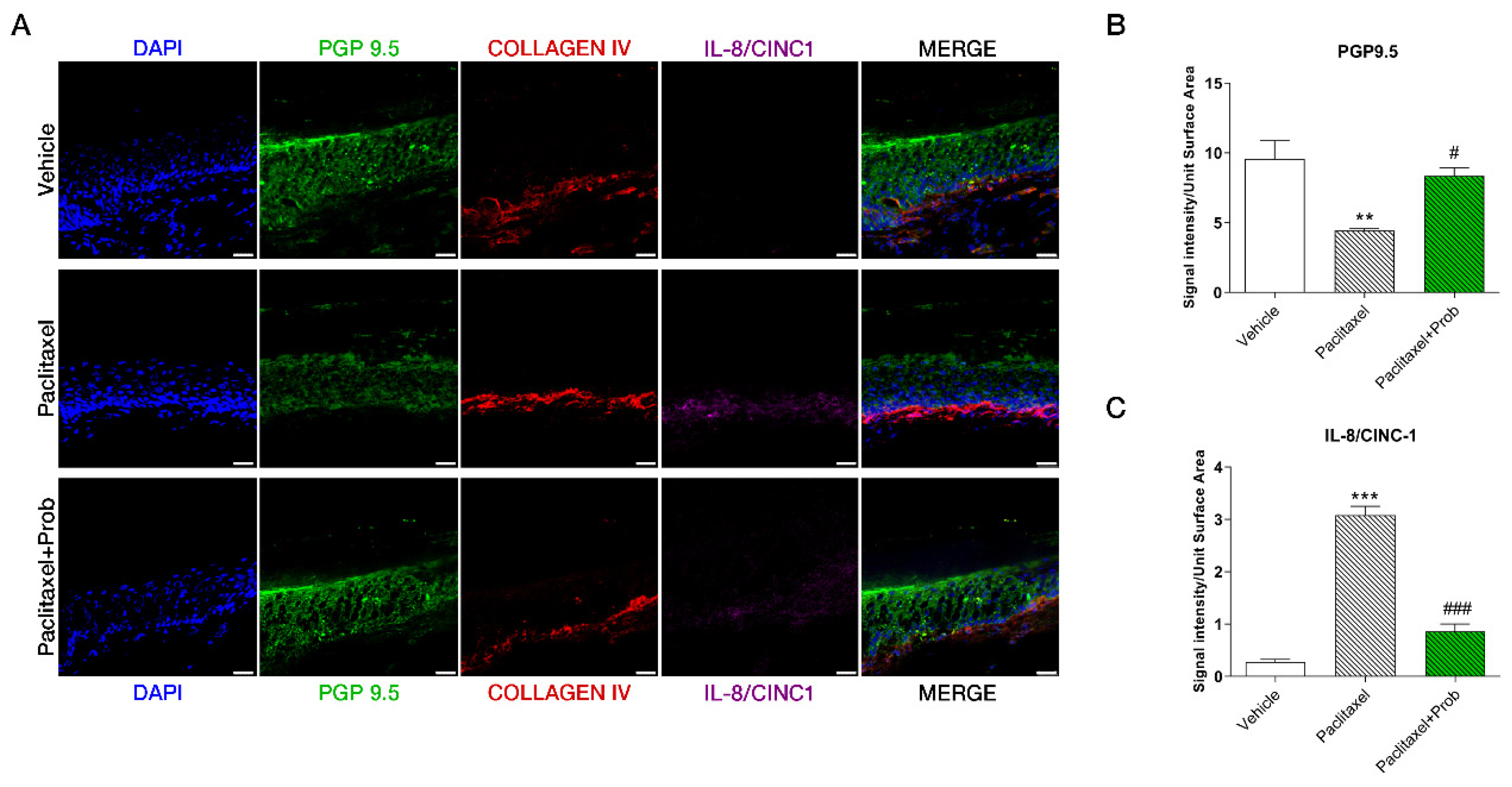
3.6. Probiotic Treatment Prevents the Paclitaxel-Induced Loss of Intra-Epidermal Fiber (IENFs) in the Paw
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miltenburg, N.C.; Boogerd, W. Chemotherapy-Induced Neuropathy: A Comprehensive Survey. Cancer Treat. Rev. 2014, 40, 872–882. [Google Scholar] [CrossRef]
- Starobova, H.; Vetter, I. Pathophysiology of Chemotherapy-Induced Peripheral Neuropathy. Front. Mol. Neurosci. 2017, 10, 174. [Google Scholar] [CrossRef]
- Seretny, M.; Currie, G.L.; Sena, E.S.; Ramnarine, S.; Grant, R.; MacLeod, M.R.; Colvin, L.A.; Fallon, M. Incidence, Prevalence, and Predictors of Chemotherapy-Induced Peripheral Neuropathy: A Systematic Review and Meta-Analysis. Pain 2014, 155, 2461–2470. [Google Scholar] [CrossRef]
- Kerckhove, N.; Collin, A.; Condé, S.; Chaleteix, C.; Pezet, D.; Balayssac, D. Long-Term Effects, Pathophysiological Mechanisms, and Risk Factors of Chemotherapy-Induced Peripheral Neuropathies: A Comprehensive Literature Review. Front. Pharmacol. 2017, 8, 86. [Google Scholar] [CrossRef]
- Scripture, C.D.; Figg, W.D.; Sparreboom, A. Peripheral Neuropathy Induced by Paclitaxel: Recent Insights and Future Perspectives. Curr. Neuropharmacol. 2006, 4, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Abu Samaan, T.M.; Samec, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Paclitaxel’s Mechanistic and Clinical Effects on Breast Cancer. Biomolecules 2019, 9, 789. [Google Scholar] [CrossRef]
- Dichwalkar, T.; Patel, S.; Bapat, S.; Pancholi, P.; Jasani, N.; Desai, B.; Yellepeddi, V.K.; Sehdev, V. Omega-3 Fatty Acid Grafted PAMAM-Paclitaxel Conjugate Exhibits Enhanced Anticancer Activity in Upper Gastrointestinal Cancer Cells. Macromol. Biosci. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-López, J.; Bravo-Caparrós, I.; Cabeza, L.; Nieto, F.R.; Ortiz, R.; Perazzoli, G.; Fernández-Segura, E.; Cañizares, F.J.; Baeyens, J.M.; Melguizo, C.; et al. Paclitaxel Antitumor Effect Improvement in Lung Cancer and Prevention of the Painful Neuropathy Using Large Pegylated Cationic Liposomes. Biomed. Pharmacother. 2021, 133, 111059. [Google Scholar] [CrossRef]
- Hu, L.-Y.; Mi, W.-L.; Wu, G.-C.; Wang, Y.-Q.; Mao-Ying, Q.-L. Prevention and Treatment for Chemotherapy-Induced Peripheral Neuropathy: Therapies Based on CIPN Mechanisms. Curr. Neuropharmacol. 2019, 17, 184–196. [Google Scholar] [CrossRef]
- Gornstein, E.L.; Schwarz, T.L. Neurotoxic Mechanisms of Paclitaxel Are Local to the Distal Axon and Independent of Transport Defects. Exp. Neurol. 2017, 288, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Chen, L.-H.; Xing, C.; Liu, T. Pain Regulation by Gut Microbiota: Molecular Mechanisms and Therapeutic Potential. Br. J. Anaesth. 2019, 123, 637–654. [Google Scholar] [CrossRef]
- Mukaida, N. Intestinal Microbiota: Unexpected Alliance with Tumor Therapy. Immunotherapy 2014, 6, 231–233. [Google Scholar] [CrossRef]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The Intestinal Microbiota Modulates the Anticancer Immune Effects of Cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Montassier, E.; Gastinne, T.; Vangay, P.; Al-Ghalith, G.A.; Bruley des Varannes, S.; Massart, S.; Moreau, P.; Potel, G.; de La Cochetière, M.F.; Batard, E.; et al. Chemotherapy-Driven Dysbiosis in the Intestinal Microbiome. Aliment. Pharmacol. Ther. 2015, 42, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Deleemans, J.M.; Chleilat, F.; Reimer, R.A.; Henning, J.-W.; Baydoun, M.; Piedalue, K.-A.; McLennan, A.; Carlson, L.E. The Chemo-Gut Study: Investigating the Long-Term Effects of Chemotherapy on Gut Microbiota, Metabolic, Immune, Psychological and Cognitive Parameters in Young Adult Cancer Survivors; Study Protocol. BMC Cancer 2019, 19, 1243. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Lim, G.; You, Z.; Ding, W.; Huang, P.; Ran, C.; Doheny, J.; Caravan, P.; Tate, S.; Hu, K.; et al. Gut Microbiota Is Critical for the Induction of Chemotherapy-Induced Pain. Nat. Neurosci. 2017, 20, 1213–1216. [Google Scholar] [CrossRef] [PubMed]
- Castelli, V.; Palumbo, P.; d’Angelo, M.; Moorthy, N.K.; Antonosante, A.; Catanesi, M.; Lombardi, F.; Iannotta, D.; Cinque, B.; Benedetti, E.; et al. Probiotic DSF Counteracts Chemotherapy Induced Neuropathic Pain. Oncotarget 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Rousseaux, C.; Thuru, X.; Gelot, A.; Barnich, N.; Neut, C.; Dubuquoy, L.; Dubuquoy, C.; Merour, E.; Geboes, K.; Chamaillard, M.; et al. Lactobacillus Acidophilus Modulates Intestinal Pain and Induces Opioid and Cannabinoid Receptors. Nat. Med. 2007, 13, 35–37. [Google Scholar] [CrossRef]
- Rhee, S.H.; Pothoulakis, C.; Mayer, E.A. Principles and Clinical Implications of the Brain-Gut-Enteric Microbiota Axis. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 306–314. [Google Scholar] [CrossRef]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of Cannabinoid Receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef]
- Smith, F.L.; Cichewicz, D.; Martin, Z.L.; Welch, S.P. The Enhancement of Morphine Antinociception in Mice by Delta9-Tetrahydrocannabinol. Pharmacol. Biochem. Behav. 1998, 60, 559–566. [Google Scholar] [CrossRef]
- Seely, K.A.; Brents, L.K.; Franks, L.N.; Rajasekaran, M.; Zimmerman, S.M.; Fantegrossi, W.E.; Prather, P.L. AM-251 and Rimonabant Act as Direct Antagonists at Mu-Opioid Receptors: Implications for Opioid/Cannabinoid Interaction Studies. Neuropharmacology 2012, 63, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Cichewicz, D.L. Synergistic Interactions between Cannabinoid and Opioid Analgesics. Life Sci. 2004, 74, 1317–1324. [Google Scholar] [CrossRef]
- Tham, S.M.; Angus, J.A.; Tudor, E.M.; Wright, C.E. Synergistic and Additive Interactions of the Cannabinoid Agonist CP55,940 with Mu Opioid Receptor and Alpha2-Adrenoceptor Agonists in Acute Pain Models in Mice. Br. J. Pharmacol. 2005, 144, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.D.; Gennings, C.; Shih, M. Synergistic Affective Analgesic Interaction between Delta-9-Tetrahydrocannabinol and Morphine. Eur. J. Pharmacol. 2006, 530, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Bushlin, I.; Rozenfeld, R.; Devi, L.A. Cannabinoid-Opioid Interactions during Neuropathic Pain and Analgesia. Curr. Opin. Pharmacol. 2010, 10, 80–86. [Google Scholar] [CrossRef]
- Wilson-Poe, A.R.; Pocius, E.; Herschbach, M.; Morgan, M.M. The Periaqueductal Gray Contributes to Bidirectional Enhancement of Antinociception between Morphine and Cannabinoids. Pharmacol. Biochem. Behav. 2013, 103, 444–449. [Google Scholar] [CrossRef]
- Huang, W.; He, J.; Nisar, M.F.; Li, H.; Wan, C. Phytochemical and Pharmacological Properties of Chaenomeles Speciosa: An Edible Medicinal Chinese Mugua. Evid. Based Complement. Alternat. Med. 2018, 2018, 9591845. [Google Scholar] [CrossRef]
- Catanesi, M.; d’Angelo, M.; Antonosante, A.; Castelli, V.; Alfonsetti, M.; Benedetti, E.; Desideri, G.; Ferri, C.; Cimini, A. Neuroprotective Potential of Choline Alfoscerate against Β-amyloid Injury: Involvement of Neurotrophic Signals. Cell Biol. Int. 2020, 44, 1734–1744. [Google Scholar] [CrossRef] [PubMed]
- Tabrizian, K.; Shahraki, J.; Bazzi, M.; Rezaee, R.; Jahantigh, H.; Hashemzaei, M. Neuro-Protective Effects of Resveratrol on Carbon Monoxide-Induced Toxicity in Male Rats. Phytother. Res. 2017, 31, 1310–1315. [Google Scholar] [CrossRef] [PubMed]
- Catanesi, M.; Caioni, G.; Castelli, V.; Benedetti, E.; d’Angelo, M.; Cimini, A. Benefits under the Sea: The Role of Marine Compounds in Neurodegenerative Disorders. Mar. Drugs 2021, 19, 24. [Google Scholar] [CrossRef]
- Nicola, S.; Amoruso, A.; Deidda, F.; Pane, M.; Allesina, S.; Mogna, L.; Del Piano, M.; Mogna, G. Searching for the Perfect Homeostasis: Five Strains of Bifidobacterium Longum From Centenarians Have a Similar Behavior in the Production of Cytokines. J. Clin. Gastroenterol. 2016, 50, S126–S130. [Google Scholar] [CrossRef]
- Nowak, A.; Paliwoda, A.; Błasiak, J. Anti-Proliferative, pro-Apoptotic and Anti-Oxidative Activity of Lactobacillus and Bifidobacterium Strains: A Review of Mechanisms and Therapeutic Perspectives. Crit. Rev. Food Sci. Nutr. 2019, 59, 3456–3467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-M.; An, J. Cytokines, Inflammation, and Pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, S.K.; El-Bedewy, M.M. Effect of Probiotics on Pro-Inflammatory Cytokines and NF-KappaB Activation in Ulcerative Colitis. World J. Gastroenterol. 2010, 16, 4145–4151. [Google Scholar] [CrossRef]
- Shadnoush, M.; Shaker Hosseini, R.; Mehrabi, Y.; Delpisheh, A.; Alipoor, E.; Faghfoori, Z.; Mohammadpour, N.; Zaringhalam Moghadam, J. Probiotic Yogurt Affects Pro- and Anti-Inflammatory Factors in Patients with Inflammatory Bowel Disease. Iran. J. Pharm. Res. 2013, 12, 929–936. [Google Scholar]
- Plaza-Díaz, J.; Ruiz-Ojeda, F.J.; Vilchez-Padial, L.M.; Gil, A. Evidence of the Anti-Inflammatory Effects of Probiotics and Synbiotics in Intestinal Chronic Diseases. Nutrients 2017, 9, 555. [Google Scholar] [CrossRef]
- Bonfili, L.; Cecarini, V.; Cuccioloni, M.; Angeletti, M.; Berardi, S.; Scarpona, S.; Rossi, G.; Eleuteri, A.M. SLAB51 Probiotic Formulation Activates SIRT1 Pathway Promoting Antioxidant and Neuroprotective Effects in an AD Mouse Model. Mol. Neurobiol. 2018, 55, 7987–8000. [Google Scholar] [CrossRef]
- Castelli, V.; d’Angelo, M.; Lombardi, F.; Alfonsetti, M.; Antonosante, A.; Catanesi, M.; Benedetti, E.; Palumbo, P.; Cifone, M.G.; Giordano, A.; et al. Effects of the Probiotic Formulation SLAB51 in in Vitro and in Vivo Parkinson’s Disease Models. Aging 2020, 12, 4641–4659. [Google Scholar] [CrossRef]
- Toma, W.; Kyte, S.L.; Bagdas, D.; Alkhlaif, Y.; Alsharari, S.D.; Lichtman, A.H.; Chen, Z.-J.; Del Fabbro, E.; Bigbee, J.W.; Gewirtz, D.A.; et al. Effects of Paclitaxel on the Development of Neuropathy and Affective Behaviors in the Mouse. Neuropharmacology 2017, 117, 305–315. [Google Scholar] [CrossRef]
- González-Chávez, S.A.; Pacheco-Tena, C.; Macías-Vázquez, C.E.; Luévano-Flores, E. Assessment of Different Decalcifying Protocols on Osteopontin and Osteocalcin Immunostaining in Whole Bone Specimens of Arthritis Rat Model by Confocal Immunofluorescence. Int. J. Clin. Exp. Pathol. 2013, 6, 1972–1983. [Google Scholar]
- Curtis, M.J.; Alexander, S.; Cirino, G.; Docherty, J.R.; George, C.H.; Giembycz, M.A.; Hoyer, D.; Insel, P.A.; Izzo, A.A.; Ji, Y.; et al. Experimental Design and Analysis and Their Reporting II: Updated and Simplified Guidance for Authors and Peer Reviewers: Editorial. Br. J. Pharmacol. 2018, 175, 987–993. [Google Scholar] [CrossRef]
- Zanardelli, M.; Micheli, L.; Cinci, L.; Failli, P.; Ghelardini, C.; Di Cesare Mannelli, L. Oxaliplatin Neurotoxicity Involves Peroxisome Alterations. PPARγ Agonism as Preventive Pharmacological Approach. PLoS ONE 2014, 9, e102758. [Google Scholar] [CrossRef]
- Brandolini, L.; Benedetti, E.; Ruffini, P.A.; Russo, R.; Cristiano, L.; Antonosante, A.; d’Angelo, M.; Castelli, V.; Giordano, A.; Allegretti, M.; et al. CXCR1/2 Pathways in Paclitaxel-Induced Neuropathic Pain. Oncotarget 2017, 8, 23188–23201. [Google Scholar] [CrossRef] [PubMed]
- Polomano, R.C.; Mannes, A.J.; Clark, U.S.; Bennett, G.J. A Painful Peripheral Neuropathy in the Rat Produced by the Chemotherapeutic Drug, Paclitaxel. Pain 2001, 94, 293–304. [Google Scholar] [CrossRef]
- Topp, K.S.; Tanner, K.D.; Levine, J.D. Damage to the Cytoskeleton of Large Diameter Sensory Neurons and Myelinated Axons in Vincristine-Induced Painful Peripheral Neuropathy in the Rat. J. Comp. Neurol. 2000, 424, 563–576. [Google Scholar] [CrossRef]
- Zajączkowska, R.; Kocot-Kępska, M.; Leppert, W.; Wrzosek, A.; Mika, J.; Wordliczek, J. Mechanisms of Chemotherapy-Induced Peripheral Neuropathy. Int. J. Mol. Sci. 2019, 20, 1451. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Y.; Zhang, H.; Jin, J.; Ma, Y.; Leng, Y. Sinomenine Alleviates Dorsal Root Ganglia Inflammation to Inhibit Neuropathic Pain via the P38 MAPK/CREB Signalling Pathway. Eur. J. Pharmacol. 2021, 897, 173945. [Google Scholar] [CrossRef]
- Fang, J.; Du, J.; Xiang, X.; Shao, X.; He, X.; Jiang, Y.; Liu, B.; Liang, Y.; Fang, J. SNI and CFA Induce Similar Changes in TRPV1 and P2X3 Expressions in the Acute Phase but Not in the Chronic Phase of Pain. Exp. Brain Res. 2021. [Google Scholar] [CrossRef]
- Flatters, S.J.L.; Bennett, G.J. Studies of Peripheral Sensory Nerves in Paclitaxel-Induced Painful Peripheral Neuropathy: Evidence for Mitochondrial Dysfunction. Pain 2006, 122, 245–257. [Google Scholar] [CrossRef]
- LaPointe, N.E.; Morfini, G.; Brady, S.T.; Feinstein, S.C.; Wilson, L.; Jordan, M.A. Effects of Eribulin, Vincristine, Paclitaxel and Ixabepilone on Fast Axonal Transport and Kinesin-1 Driven Microtubule Gliding: Implications for Chemotherapy-Induced Peripheral Neuropathy. Neurotoxicology 2013, 37, 231–239. [Google Scholar] [CrossRef]
- Lisse, T.S.; Middleton, L.J.; Pellegrini, A.D.; Martin, P.B.; Spaulding, E.L.; Lopes, O.; Brochu, E.A.; Carter, E.V.; Waldron, A.; Rieger, S. Paclitaxel-Induced Epithelial Damage and Ectopic MMP-13 Expression Promotes Neurotoxicity in Zebrafish. Proc. Natl. Acad. Sci. USA 2016, 113, E2189–E2198. [Google Scholar] [CrossRef]
- Forsythe, P.; Bienenstock, J.; Kunze, W.A. Vagal Pathways for Microbiome-Brain-Gut Axis Communication. Adv. Exp. Med. Biol. 2014, 817, 115–133. [Google Scholar] [CrossRef]
- Foster, J.A.; McVey Neufeld, K.-A. Gut-Brain Axis: How the Microbiome Influences Anxiety and Depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef]
- Bai, J.; Behera, M.; Bruner, D.W. The Gut Microbiome, Symptoms, and Targeted Interventions in Children with Cancer: A Systematic Review. Support. Care Cancer 2018, 26, 427–439. [Google Scholar] [CrossRef]
- Jordan, K.R.; Loman, B.R.; Bailey, M.T.; Pyter, L.M. Gut Microbiota-Immune-Brain Interactions in Chemotherapy-Associated Behavioral Comorbidities. Cancer 2018, 124, 3990–3999. [Google Scholar] [CrossRef]
- Jacola, L.M.; Edelstein, K.; Liu, W.; Pui, C.-H.; Hayashi, R.; Kadan-Lottick, N.S.; Srivastava, D.; Henderson, T.; Leisenring, W.; Robison, L.L.; et al. Cognitive, Behaviour, and Academic Functioning in Adolescent and Young Adult Survivors of Childhood Acute Lymphoblastic Leukaemia: A Report from the Childhood Cancer Survivor Study. Lancet Psychiatry 2016, 3, 965–972. [Google Scholar] [CrossRef]
- Schulte, F.; Brinkman, T.M.; Li, C.; Fay-McClymont, T.; Srivastava, D.K.; Ness, K.K.; Howell, R.M.; Mueller, S.; Wells, E.; Strother, D.; et al. Social Adjustment in Adolescent Survivors of Pediatric Central Nervous System Tumors: A Report from the Childhood Cancer Survivor Study. Cancer 2018, 124, 3596–3608. [Google Scholar] [CrossRef] [PubMed]
- Iacob, S.; Iacob, D.G. Infectious Threats, the Intestinal Barrier, and Its Trojan Horse: Dysbiosis. Front. Microbiol. 2019, 10, 1676. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, M.F.; Lee, C.Y.; Chen, W.N.; Shaikh, F.A. The Gut-Brain-Axis on the Manifestation of Depressive Symptoms in Epilepsy: An Evidence-Driven Hypothesis. Front. Pharmacol. 2020, 11, 465. [Google Scholar] [CrossRef] [PubMed]
- Zmora, N.; Zilberman-Schapira, G.; Suez, J.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Kotler, E.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.-Z.; et al. Personalized Gut Mucosal Colonization Resistance to Empiric Probiotics Is Associated with Unique Host and Microbiome Features. Cell 2018, 174, 1388–1405.e21. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.; Neish, A. Role of Gut Microbiota in Intestinal Wound Healing and Barrier Function. Tissue Barriers 2018, 6, 1539595. [Google Scholar] [CrossRef]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal Barrier and Gut Microbiota: Shaping Our Immune Responses throughout Life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Scharl, M.; Rogler, G. Inflammatory Bowel Disease Pathogenesis: What Is New? Curr. Opin. Gastroenterol. 2012, 28, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Dogra, N.; Mani, R.J.; Katare, D.P. The Gut-Brain Axis: Two Ways Signaling in Parkinson’s Disease. Cell Mol. Neurobiol. 2021. [Google Scholar] [CrossRef]
- Kriss, M.; Hazleton, K.Z.; Nusbacher, N.M.; Martin, C.G.; Lozupone, C.A. Low Diversity Gut Microbiota Dysbiosis: Drivers, Functional Implications and Recovery. Curr. Opin. Microbiol. 2018, 44, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.W.M.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of Intestinal Microbiota from Lean Donors Increases Insulin Sensitivity in Individuals with Metabolic Syndrome. Gastroenterology 2012, 143, 913–916.e7. [Google Scholar] [CrossRef] [PubMed]
- Dabke, K.; Hendrick, G.; Devkota, S. The Gut Microbiome and Metabolic Syndrome. J. Clin. Investig. 2019, 129, 4050–4057. [Google Scholar] [CrossRef]
- Ramakrishna, C.; Corleto, J.; Ruegger, P.M.; Logan, G.D.; Peacock, B.B.; Mendonca, S.; Yamaki, S.; Adamson, T.; Ermel, R.; McKemy, D.; et al. Dominant Role of the Gut Microbiota in Chemotherapy Induced Neuropathic Pain. Sci. Rep. 2019, 9, 20324. [Google Scholar] [CrossRef]
- Yang, C.; Fang, X.; Zhan, G.; Huang, N.; Li, S.; Bi, J.; Jiang, R.; Yang, L.; Miao, L.; Zhu, B.; et al. Key Role of Gut Microbiota in Anhedonia-like Phenotype in Rodents with Neuropathic Pain. Transl. Psychiatry 2019, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Cristiano, C.; Avagliano, C.; De Caro, C.; La Rana, G.; Raso, G.M.; Canani, R.B.; Meli, R.; Calignano, A. Gut-Brain Axis: Role of Lipids in the Regulation of Inflammation, Pain and CNS Diseases. Curr. Med. Chem. 2018, 25, 3930–3952. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, J.M.; Cheer, J.F. Endocannabinoid Regulation of Reward and Reinforcement through Interaction with Dopamine and Endogenous Opioid Signaling. Neuropsychopharmacology 2018, 43, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Kienzl, M.; Storr, M.; Schicho, R. Cannabinoids and Opioids in the Treatment of Inflammatory Bowel Diseases. Clin. Transl. Gastroenterol. 2020, 11, e00120. [Google Scholar] [CrossRef]
- Theodorou, V.; Ait Belgnaoui, A.; Agostini, S.; Eutamene, H. Effect of Commensals and Probiotics on Visceral Sensitivity and Pain in Irritable Bowel Syndrome. Gut Microbes 2014, 5, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Amaral, F.A.; Sachs, D.; Costa, V.V.; Fagundes, C.T.; Cisalpino, D.; Cunha, T.M.; Ferreira, S.H.; Cunha, F.Q.; Silva, T.A.; Nicoli, J.R.; et al. Commensal Microbiota Is Fundamental for the Development of Inflammatory Pain. Proc. Natl. Acad. Sci. USA 2008, 105, 2193–2197. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-M.; Lehky, T.J.; Brell, J.M.; Dorsey, S.G. Discovering Cytokines as Targets for Chemotherapy-Induced Painful Peripheral Neuropathy. Cytokine 2012, 59, 3–9. [Google Scholar] [CrossRef]
- Jaggi, A.S.; Singh, N. Mechanisms in Cancer-Chemotherapeutic Drugs-Induced Peripheral Neuropathy. Toxicology 2012, 291, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ledeboer, A.; Jekich, B.M.; Sloane, E.M.; Mahoney, J.H.; Langer, S.J.; Milligan, E.D.; Martin, D.; Maier, S.F.; Johnson, K.W.; Leinwand, L.A.; et al. Intrathecal Interleukin-10 Gene Therapy Attenuates Paclitaxel-Induced Mechanical Allodynia and Proinflammatory Cytokine Expression in Dorsal Root Ganglia in Rats. Brain Behav. Immun. 2007, 21, 686–698. [Google Scholar] [CrossRef]
- Woolf, C.J.; Allchorne, A.; Safieh-Garabedian, B.; Poole, S. Cytokines, Nerve Growth Factor and Inflammatory Hyperalgesia: The Contribution of Tumour Necrosis Factor Alpha. Br. J. Pharmacol. 1997, 121, 417–424. [Google Scholar] [CrossRef]
- Özaktay, C.; Cavanaugh, J.; Asik, I.; DeLeo, J.; Weinstein, J. Dorsal Root Sensitivity to Interleukin-1 Beta, Interleukin-6 and Tumor Necrosis Factor in Rats. Eur. Spine J. 2002, 11, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Binshtok, A.M.; Wang, H.; Zimmermann, K.; Amaya, F.; Vardeh, D.; Shi, L.; Brenner, G.J.; Ji, R.-R.; Bean, B.P.; Woolf, C.J.; et al. Nociceptors Are Interleukin-1 Sensors. J. Neurosci. 2008, 28, 14062–14073. [Google Scholar] [CrossRef] [PubMed]
- Boyette-Davis, J.; Xin, W.; Zhang, H.; Dougherty, P.M. Intraepidermal Nerve Fiber Loss Corresponds to the Development of Taxol-Induced Hyperalgesia and Can Be Prevented by Treatment with Minocycline. Pain 2011, 152, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Boyette-Davis, J.; Dougherty, P.M. Protection against Oxaliplatin-Induced Mechanical Hyperalgesia and Intraepidermal Nerve Fiber Loss by Minocycline. Exp. Neurol. 2011, 229, 353–357. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuozzo, M.; Castelli, V.; Avagliano, C.; Cimini, A.; d’Angelo, M.; Cristiano, C.; Russo, R. Effects of Chronic Oral Probiotic Treatment in Paclitaxel-Induced Neuropathic Pain. Biomedicines 2021, 9, 346. https://doi.org/10.3390/biomedicines9040346
Cuozzo M, Castelli V, Avagliano C, Cimini A, d’Angelo M, Cristiano C, Russo R. Effects of Chronic Oral Probiotic Treatment in Paclitaxel-Induced Neuropathic Pain. Biomedicines. 2021; 9(4):346. https://doi.org/10.3390/biomedicines9040346
Chicago/Turabian StyleCuozzo, Mariarosaria, Vanessa Castelli, Carmen Avagliano, Annamaria Cimini, Michele d’Angelo, Claudia Cristiano, and Roberto Russo. 2021. "Effects of Chronic Oral Probiotic Treatment in Paclitaxel-Induced Neuropathic Pain" Biomedicines 9, no. 4: 346. https://doi.org/10.3390/biomedicines9040346
APA StyleCuozzo, M., Castelli, V., Avagliano, C., Cimini, A., d’Angelo, M., Cristiano, C., & Russo, R. (2021). Effects of Chronic Oral Probiotic Treatment in Paclitaxel-Induced Neuropathic Pain. Biomedicines, 9(4), 346. https://doi.org/10.3390/biomedicines9040346