
Dietary Iron Intake in Excess of Requirements Impairs Intestinal Copper Absorption in Sprague Dawley Rat Dams, Causing Copper Deficiency in Suckling Pups




Abstract
1. Introduction
2. Materials and Methods
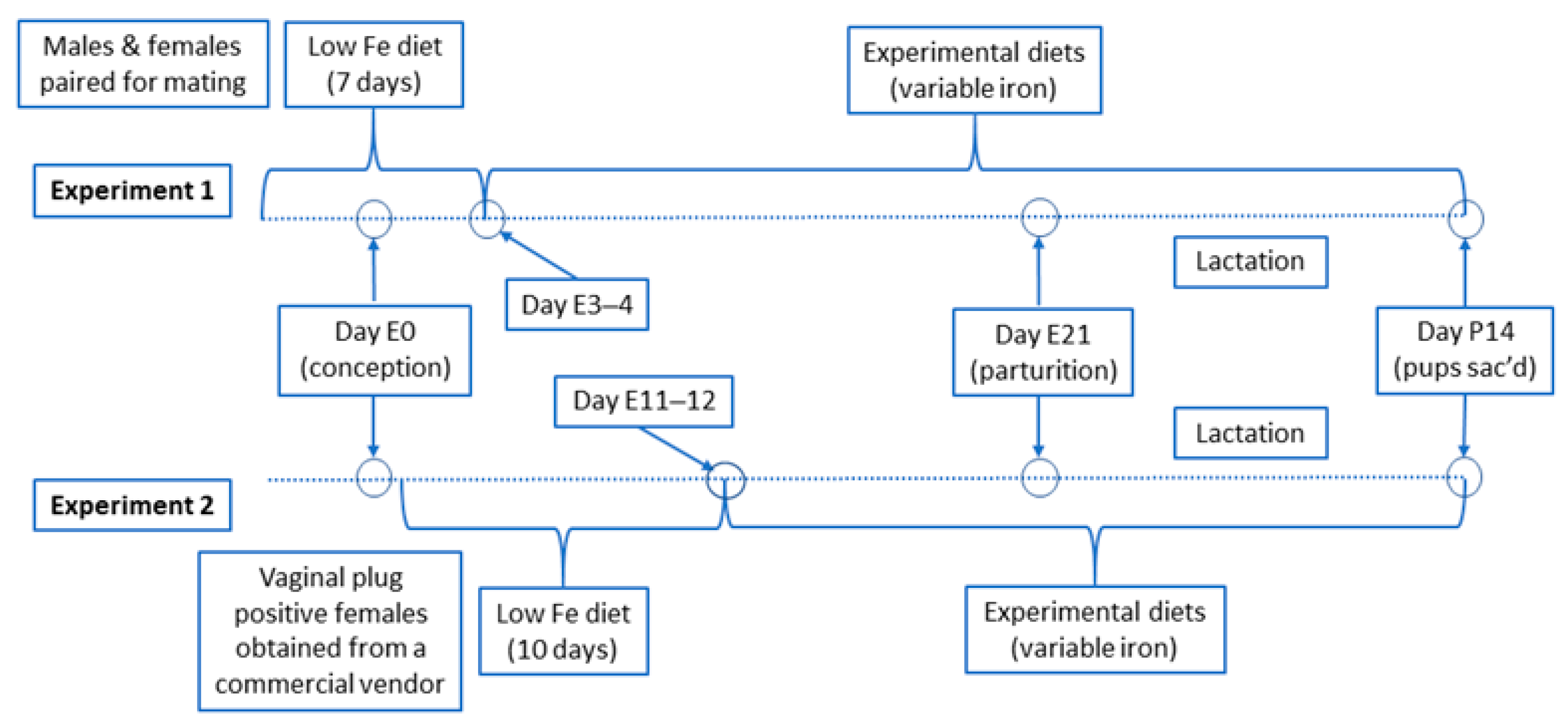
2.1. Experimental Design and Dietary Manipulations
2.2. Quantification of Hematological Parameters, Serum and Tissue Nonheme Fe Levels, and Copper Biomarker Assays
2.3. Quantification of Iron and Copper Concentrations in Experimental Diets and Copper in Liver
2.4. Intestinal Copper Absorption Study
2.5. Statistical Analysis
3. Results
3.1. Iron and Copper Content of Experimental Diets
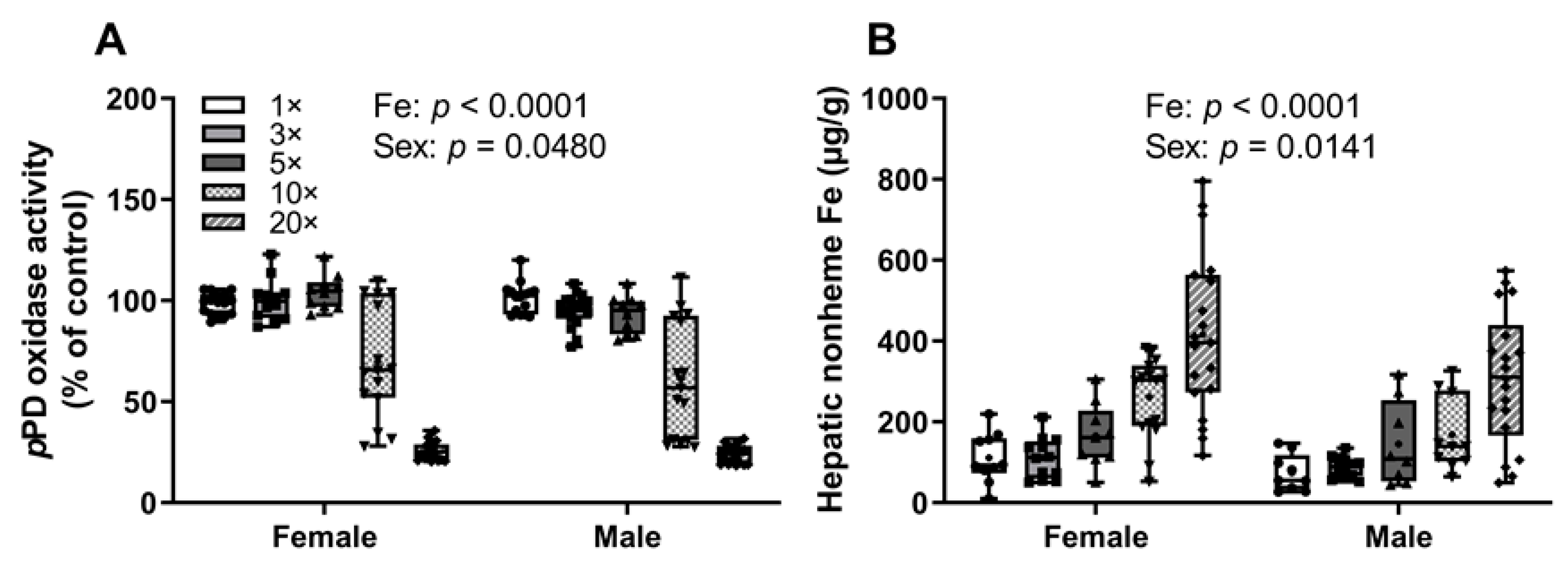
3.1.1. Experiment 1: High-Iron Intake by Rat Dams Suppresses Serum pPD Oxidase Activity and Increases Hepatic Nonheme Iron in Suckling Pups
3.1.2. Experiment 2: Optimized Experimental Design
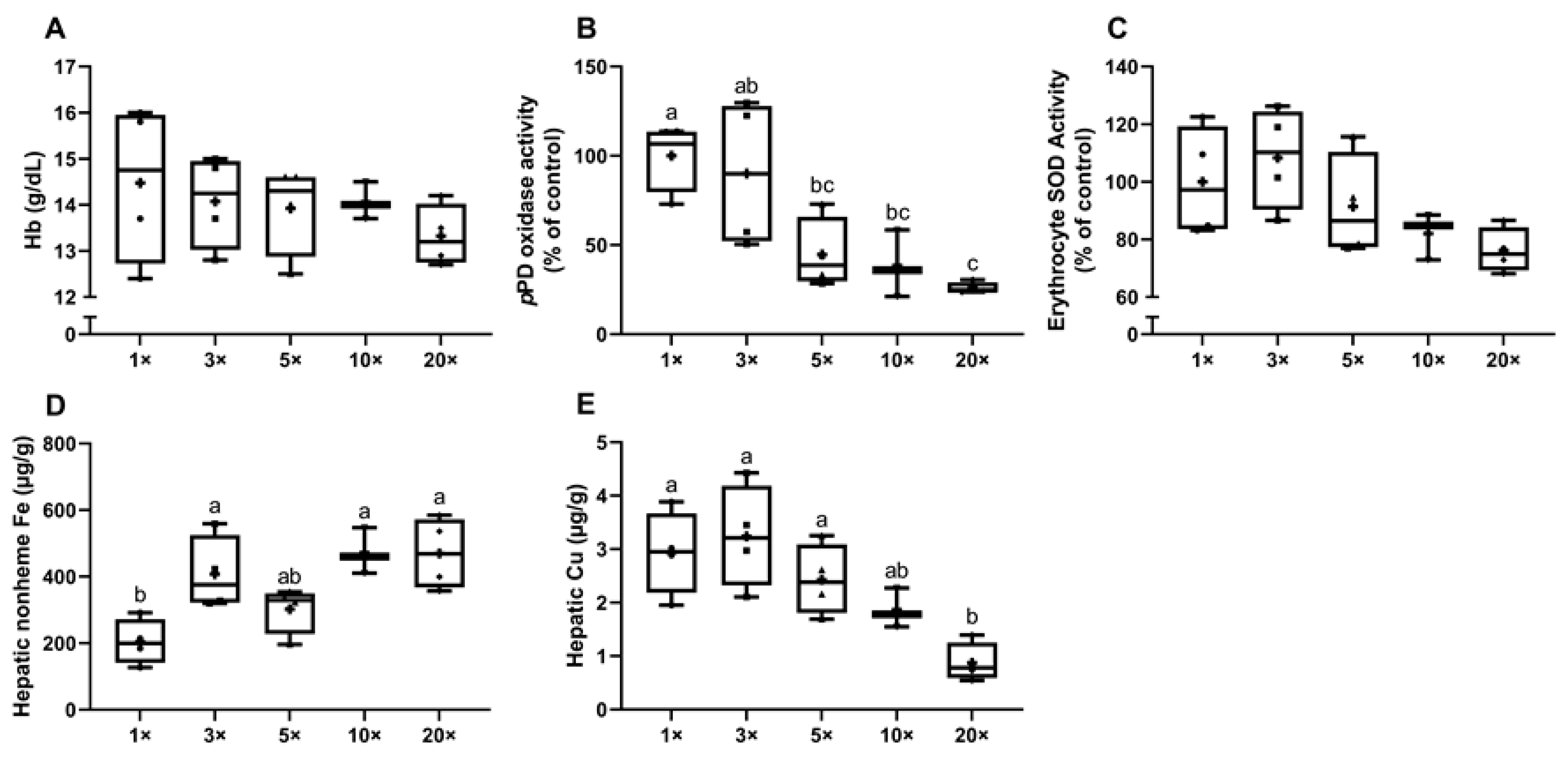
3.1.3. Experiment 2: High-Iron Consumption Perturbs Iron and Copper Homeostasis in Dams
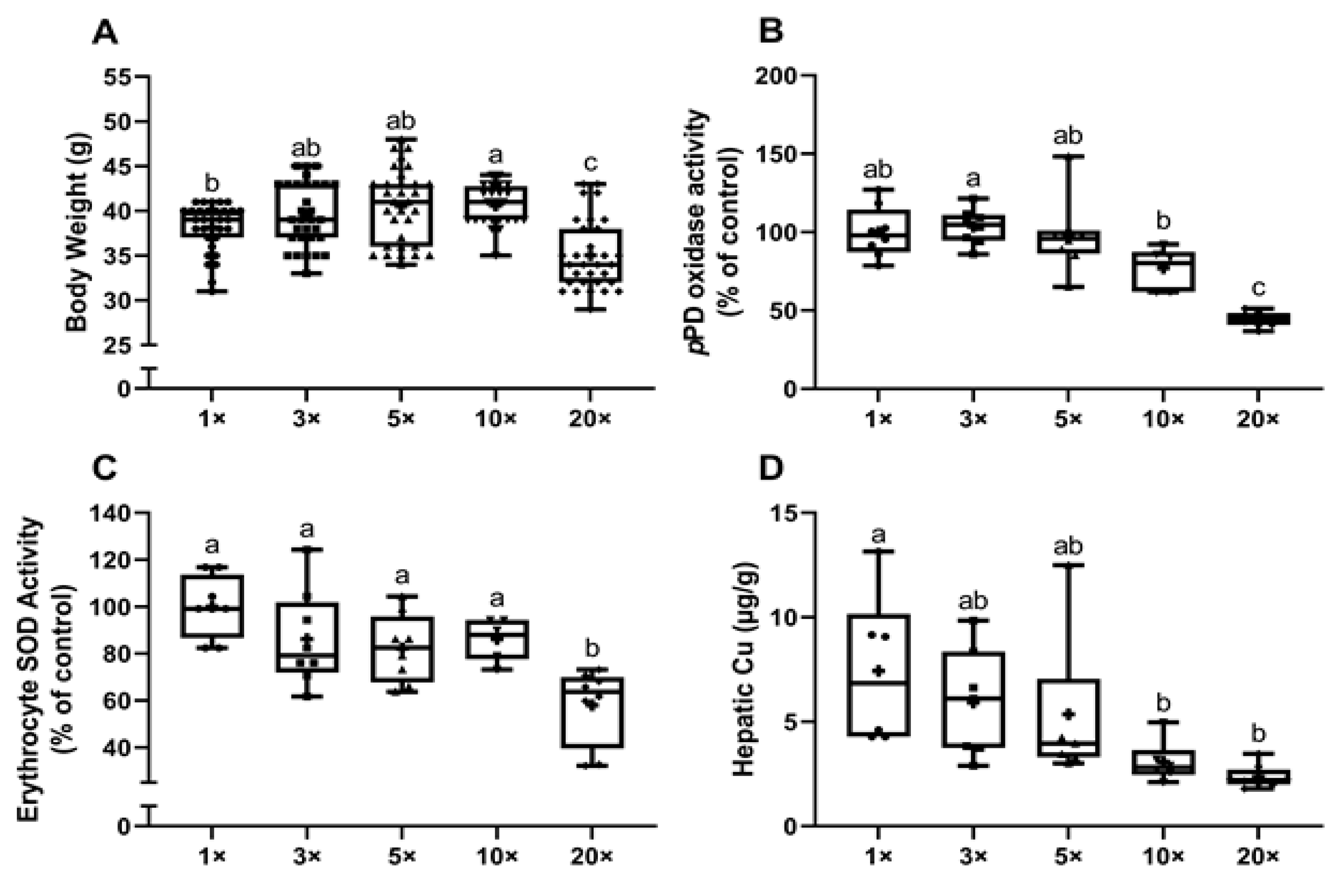
3.1.4. Experiment 2: High-Iron Consumption by Rat Dams Alters Final Body Weights, Serum pPD Oxidase and Erythrocyte SOD1 Activities and Hepatic Copper Content in Suckling Pups
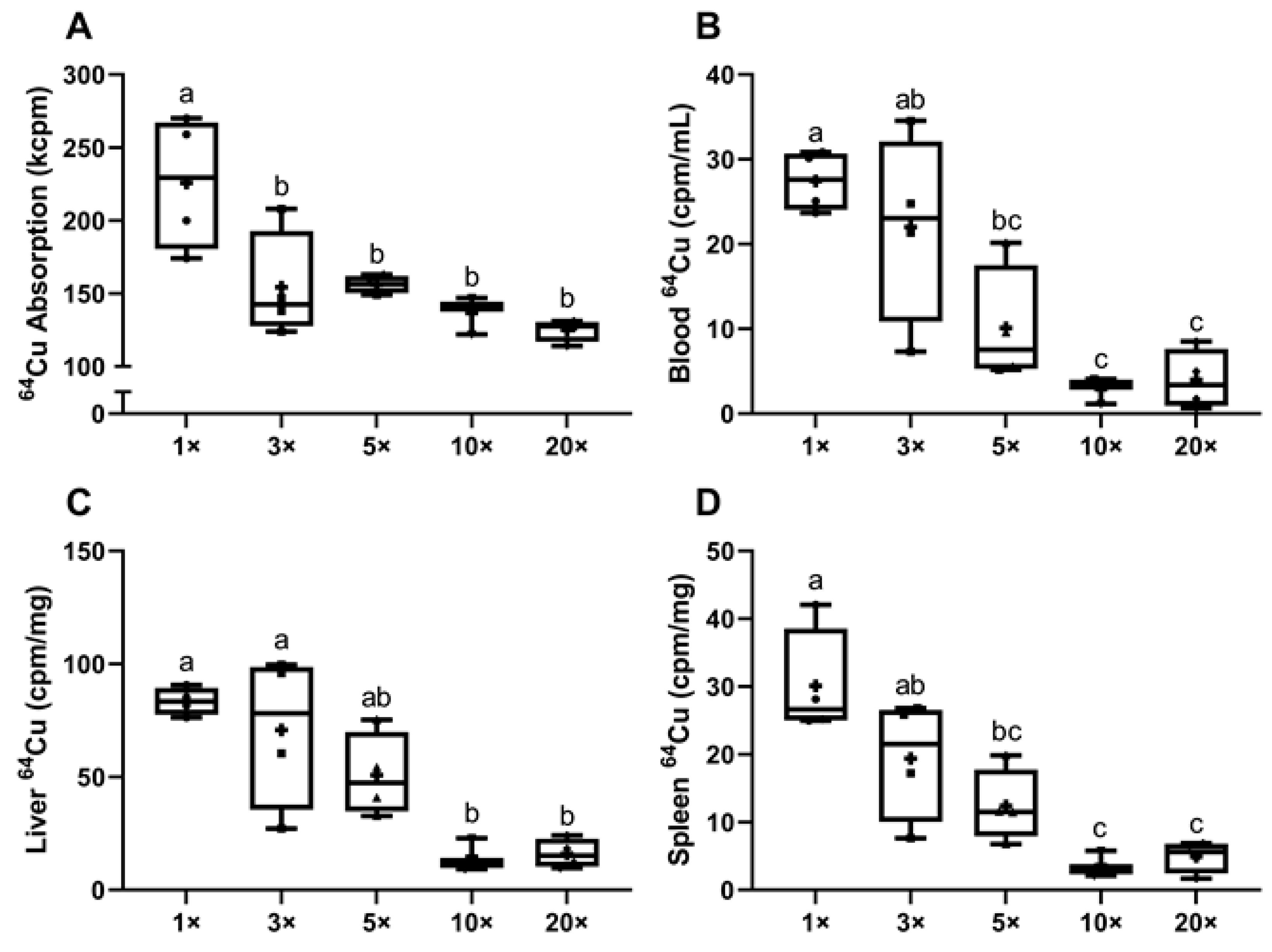
3.1.5. Experiment 2: Increasing Dietary Iron to ~3-Fold above Requirements Decreases Intestinal Copper (64Cu) Absorption in Lactating Rat Dams
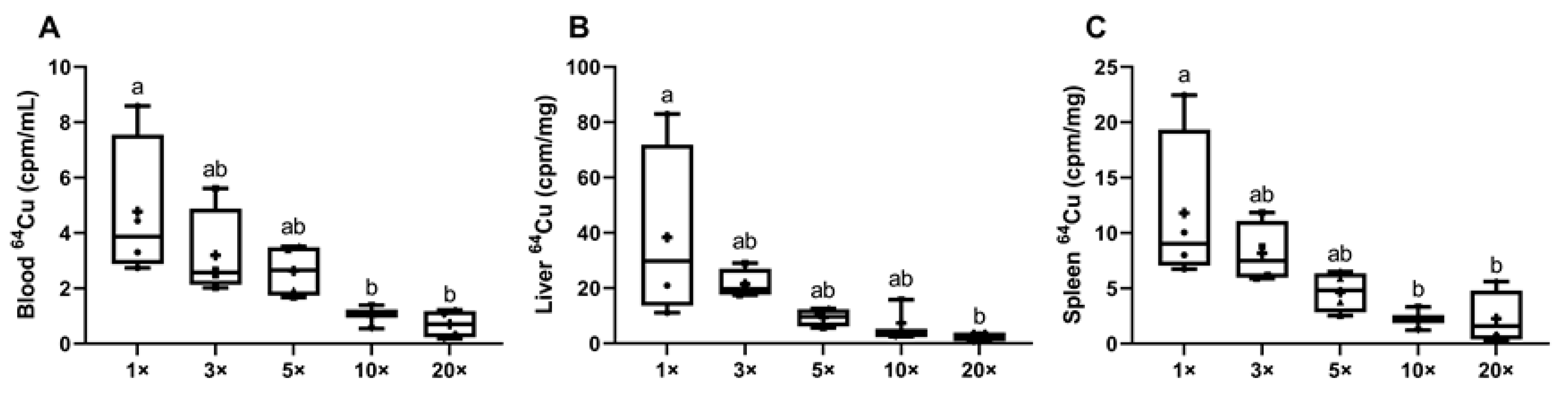
3.1.6. Experiment 2: Consumption of High-Iron Diets by Lactating Dams Decreases Copper Delivery to Suckling Pups
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Camaschella, C. Iron deficiency. Blood 2019, 133, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Pasricha, S.-R.; Tye-Din, J.; Muckenthaler, M.U.; Swinkels, D.W. Iron deficiency. Lancet 2021, 397, 233–248. [Google Scholar] [CrossRef]
- Cappellini, M.D.; Musallam, K.M.; Taher, A.T. Iron deficiency anaemia revisited. J. Intern. Med. 2019, 287, 153–170. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Vichinsky, E. Iron Deficiency: Implications Before Anemia. Pediatr. Rev. 2021, 42, 11–20. [Google Scholar] [CrossRef]
- Cao, C.; O’Brien, K.O. Pregnancy and iron homeostasis: An update. Nutr. Rev. 2013, 71, 35–51. [Google Scholar] [CrossRef]
- Gautam, C.S.; Saha, L.; Sekhri, K.; Saha, P.K. Iron Deficiency in Pregnancy and the Rationality of Iron Supplements Prescribed During Pregnancy. Medscape, J. Med. 2008, 10, 283. [Google Scholar]
- Rioux, F.M.; Leblanc, C.P. Iron supplementation during pregnancy: What are the risks and benefits of current practices? Appl. Physiol. Nutr. Metab. 2007, 32, 282–288. [Google Scholar] [CrossRef]
- Fan, T.; Blitz, J. Screening for Iron Deficiency Anemia and Iron Supplementation in Pregnant Women to Improve Maternal Health and Birth Outcomes: Recommendation Statement. Am. Fam. Physician 2016, 93, 133–136. [Google Scholar]
- Brannon, P.M.; Taylor, C.L. Iron Supplementation during Pregnancy and Infancy: Uncertainties and Implications for Research and Policy. Nutrients 2017, 9, 1327. [Google Scholar] [CrossRef] [PubMed]
- Peña-Rosas, J.P.; De-Regil, L.M.; Garcia-Casal, M.N.; Dowswell, T. Daily oral iron supplementation during pregnancy. Cochrane Database Syst. Rev. 2015, CD004736. [Google Scholar] [CrossRef]
- Klevay, L.M. IHD from copper deficiency: A unified theory. Nutr. Res. Rev. 2016, 29, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Klevay, L. Iron overload can induce mild copper deficiency. J. Trace Elements Med. Biol. 2001, 14, 237–240. [Google Scholar] [CrossRef]
- Ha, J.-H.; Doguer, C.; Flores, S.R.L.; Wang, T.; Collins, J.F. Progressive Increases in Dietary Iron Are Associated with the Emergence of Pathologic Disturbances of Copper Homeostasis in Growing Rats. J. Nutr. 2018, 148, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.-H.; Doguer, C.; Wang, X.; Flores, S.R.; Collins, J.F. High-Iron Consumption Impairs Growth and Causes Copper-Deficiency Anemia in Weanling Sprague-Dawley Rats. PLoS ONE 2016, 11, e0161033. [Google Scholar] [CrossRef]
- Ha, J.-H.; Doguer, C.; Collins, J.F. Consumption of a High-Iron Diet Disrupts Homeostatic Regulation of Intestinal Copper Absorption in Adolescent Mice. Am. J. Physiol. Liver Physiol. 2017, 313, G353–G360. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xiang, P.; Ha, J.-H.; Wang, X.; Doguer, C.; Flores, S.R.; Kang, Y.J.; Collins, J.F. Copper supplementation reverses dietary iron overload-induced pathologies in mice. J. Nutr. Biochem. 2018, 59, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Klevay, L.M. Is the Western diet adequate in copper? J. Trace Elements Med. Biol. 2011, 25, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Gambling, L.; Kennedy, C.; McArdle, H.J. Iron and copper in fetal development. Semin. Cell Dev. Biol. 2011, 22, 637–644. [Google Scholar] [CrossRef]
- Uriu-Adams, J.Y.; Scherr, R.E.; LaNoue, L.; Keen, C.L. Influence of copper on early development: Prenatal and postnatal considerations. BioFactors 2010, 36, 136–152. [Google Scholar] [CrossRef] [PubMed]
- Gulec, S.; Collins, J.F. Investigation of Iron Metabolism in Mice Expressing a Mutant Menke’s Copper Transporting ATPase (Atp7a) Protein with Diminished Activity (Brindled; MoBr/y). PLoS ONE 2013, 8, e66010. [Google Scholar] [CrossRef]
- Harvey, L.J.; McArdle, H.J. Biomarkers of copper status: A brief update. Br. J. Nutr. 2008, 99 (Suppl. 3), S10–S13. [Google Scholar] [CrossRef]
- Olivares, M.; Méndez, M.A.; Astudillo, P.A.; Pizarro, F. Present situation of biomarkers for copper status. Am. J. Clin. Nutr. 2008, 88, 859S–862S. [Google Scholar] [CrossRef] [PubMed]
- Markowitz, H.; Gubler, C.J.; Mahoney, J.P.; Cartwright, G.E.; Wintrobe, M.M. Studies on copper metabolism. xiv. copper, cerulo-plasmin and oxidase activity in sera of normal human subjects, pregnant women, and patients with infection, hepatolenticular degeneration and the nephrotic syndrome 1. J. Clin. Investig. 1955, 34, 1498–1508. [Google Scholar] [CrossRef] [PubMed]
- Broderius, M.A.; Prohaska, J.R. Differential impact of copper deficiency in rats on blood cuproproteins. Nutr. Res. 2009, 29, 494–502. [Google Scholar] [CrossRef]
- Prohaska, J.R.; Broderius, M. Plasma peptidylglycine alpha-amidating monooxygenase (PAM) and ceruloplasmin are affected by age and copper status in rats and mice. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2006, 143, 360–366. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sunderman, F.W.; Nomoto, S. Measurement of Human Serum Ceruloplasmin by Its p-Phenylenediamine Oxidase Activity. Clin. Chem. 1970, 16, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.F.; Flores, R.L.S.; Wang, X.; Anderson, G.J. Mechanisms and Regulation of Intestinal Iron Transport. In Physiology of the Gastrointestinal Tract, 6th ed.; Said, H., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 1451–1483. [Google Scholar]
- Woodman, A.G.; Care, A.S.; Mansour, Y.; Cherak, S.J.; Panahi, S.; Gragasin, F.S.; Bourque, S.L. Modest and Severe Maternal Iron Deficiency in Pregnancy are Associated with Fetal Anaemia and Organ-Specific Hypoxia in Rats. Sci. Rep. 2017, 7, 46573. [Google Scholar] [CrossRef] [PubMed]
- Broderius, M.; Mostad, E.; Wendroth, K.; Prohaska, J.R. Levels of plasma ceruloplasmin protein are markedly lower following dietary copper deficiency in rodents. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 151, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.C.; Houle, P.A.; Isaacs, E.; Moor, J.R.; Scott, L.E. Copper Regulation of Ceruloplasmin in Copper-Deficient Rats. Enzyme 1979, 24, 23–35. [Google Scholar] [CrossRef]
- Kono, S.; Suzuki, H.; Takahashi, K.; Takahashi, Y.; Shirakawa, K.; Murakawa, Y.; Yamaguchi, S.; Miyajima, H. Hepatic Iron Overload Associated with a Decreased Serum Ceruloplasmin Level in a Novel Clinical Type of Aceruloplasminemia. Gastroenterology 2006, 131, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Hellman, N.E.; Gitlin, J.D. CERULOPLASMINMETABOLISM ANDFUNCTION. Annu. Rev. Nutr. 2002, 22, 439–458. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.C. Ceruloplasmin and other copper binding components of blood plasma and their functions: An update. Metallomics 2016, 8, 887–905. [Google Scholar] [CrossRef]
- Prohaska, J.R. Impact of copper deficiency in humans. Ann. N. Y. Acad. Sci. 2014, 1314, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Holtzman, N.A.; Gaumnitz, B.M. Studies on the Rate of Release and Turnover of Ceruloplasmin and Apoceruloplasmin in Rat Plasma. J. Biol. Chem. 1970, 245, 2354–2358. [Google Scholar] [CrossRef]
- Gulec, S.; Collins, J.F. Molecular Mediators Governing Iron-Copper Interactions. Annu. Rev. Nutr. 2014, 34, 95–116. [Google Scholar] [CrossRef]
- Jaeggi, T.; Kortman, G.A.M.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in Kenyan infants. Gut 2015, 64, 731–742. [Google Scholar] [CrossRef]
- Paganini, D.; Uyoga, M.A.; Kortman, G.A.M.; Cercamondi, C.I.; Moretti, D.; Barth-Jaeggi, T.; Schwab, C.; Boekhorst, J.; Timmerman, H.M.; Lacroix, C.; et al. Prebiotic galacto-oligosaccharides mitigate the adverse effects of iron fortification on the gut microbiome: A randomised controlled study in Kenyan infants. Gut 2017, 66, 1956–1967. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.F.; Kordas, K.; Stoltzfus, R.J.; Black, R.E. Interactive effects of iron and zinc on biochemical and functional outcomes in supplementation trials. Am. J. Clin. Nutr. 2005, 82, 5–12. [Google Scholar] [CrossRef]
- Harvey, L.J.; Dainty, J.R.; Hollands, W.J.; Bull, V.J.; Hoogewerff, J.A.; Foxall, R.J.; McAnena, L.; Strain, J.J.; Fairweather-Tait, S.J. Effect of high-dose iron supplements on fractional zinc absorption and status in pregnant women. Am. J. Clin. Nutr. 2007, 85, 131–136. [Google Scholar] [CrossRef]
- Whittaker, P. Iron and zinc interactions in humans. Am. J. Clin. Nutr. 1998, 68, 442S–446S. [Google Scholar] [CrossRef]
- Yip, R.; Reeves, J.D.; Lönnerdal, B.; Keen, C.L.; Dallman, P.R. Does iron supplementation compromise zinc nutrition in healthy infants? Am. J. Clin. Nutr. 1985, 42, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.F. Copper. In Present Knowledge in Nutrition, 11th ed.; Marriott, B., Birt, D.F., Stalling, V., Yates, A., Eds.; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Nose, Y.; Kim, B.-E.; Thiele, D.J. Ctr1 drives intestinal copper absorption and is essential for growth, iron metabolism, and neonatal cardiac function. Cell Metab. 2006, 4, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Shawki, A.; Anthony, S.R.; Nose, Y.; Engevik, M.A.; Niespodzany, E.J.; Barrientos, T.; Öhrvik, H.; Worrell, R.T.; Thiele, D.J.; MacKenzie, B. Intestinal DMT1 is critical for iron absorption in the mouse but is not required for the absorption of copper or manganese. Am. J. Physiol. Liver Physiol. 2015, 309, G635–G647. [Google Scholar] [CrossRef] [PubMed]
- Arredondo, M.; Muñoz, P.; Mura, C.V.; Núñez, M.T. DMT1, a physiologically relevant apical Cu1+transporter of intestinal cells. Am. J. Physiol. Physiol. 2003, 284, C1525–C1530. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Zhang, Z.; Wang, T.; Chen, C.; Kang, Y.J. Copper uptake by DMT1: A compensatory mechanism for CTR1 deficiency in human umbilical vein endothelial cells. Metallomics 2015, 7, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Flores, S.R.; Ha, J.-H.; Doguer, C.; Woloshun, R.R.; Xiang, P.; Grosche, A.; Vidyasagar, S.; Collins, J.F. Intestinal DMT1 Is Essential for Optimal Assimilation of Dietary Copper in Male and Female Mice with Iron-Deficiency Anemia. J. Nutr. 2018, 148, 1244–1252. [Google Scholar] [CrossRef]
- Wyman, S.; Simpson, R.J.; McKie, A.T.; Sharp, P.A. Dcytb (Cybrd1) functions as both a ferric and a cupric reductase in vitro. FEBS Lett. 2008, 582, 1901–1906. [Google Scholar] [CrossRef]
- Sangkhae, V.; Fisher, A.L.; Wong, S.; Koenig, M.D.; Tussing-Humphreys, L.; Chu, A.; Lelić, M.; Ganz, T.; Nemeth, E. Effects of maternal iron status on placental and fetal iron homeostasis. J. Clin. Investig. 2019, 130, 625–640. [Google Scholar] [CrossRef] [PubMed]
- Sangkhae, V.; Nemeth, E. Placental iron transport: The mechanism and regulatory circuits. Free. Radic. Biol. Med. 2019, 133, 254–261. [Google Scholar] [CrossRef]
- McArdle, H.J.; Andersen, H.S.; Jones, H.; Gambling, L. Copper and Iron Transport Across the Placenta: Regulation and Interactions. J. Neuroendocr. 2008, 20, 427–431. [Google Scholar] [CrossRef]
- Gadde, U.; Oh, S.T.; Lee, Y.S.; Davis, E.; Zimmerman, N.; Rehberger, T.; Lillehoj, H.S. The Effects of Direct-fed Microbial Supplementation, as an Alternative to Antibiotics, on Growth Performance, Intestinal Immune Status, and Epithelial Barrier Gene Expression in Broiler Chickens. Probiotics Antimicrob. Proteins 2017, 9, 397–405. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Amount (g/kg) |
|---|---|
| Cornstarch Casein Dyetrose | 397 200 132 |
| Sucrose 1 | 100 |
| Soybean oil | 70 |
| Cellulose (micro) | 50 |
| Mineral Mix 2 | 35 |
| Vitamin Mix 3 | 10 |
| L-Cystine Choline Bitartrate t-Butylhydroquinone | 3 2.5 0.014 |
| Ingredient | 1× Diet | 3× Diet | 5× Diet | 10× Diet | 20× Diet |
|---|---|---|---|---|---|
| Ferric citrate 1 | 80 | 80 | 80 | 80 | 80 |
| Carbonyl Fe 1 | 0 | 160 | 320 | 720 | 1520 |
| Cupric carbonate 1 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 |
| Mineral & Energy Content | |||||
| Iron 1,2 | 88 | 235 | 422 | 886 | 1598 |
| Copper 1,2 | 2.79 | 2.75 | 2.44 | 3.01 | 2.18 |
| Energy (kcal/kg) | 3760 | 3760 | 3759 | 3757 | 3754 |
| Experiment 1: Dam 1 | ||||||
|---|---|---|---|---|---|---|
| Parameter | p2 | 1× Fe | 3× Fe | 5× Fe | 10× Fe | 20× Fe |
| Hb (g/dL) | ns | 12.8 ± 0.9 | 12.6 ± 1.0 | 13.9 ± 0.8 | 11.8 ± 1.1 | 10.3 ± 1.8 |
| Serum NH Fe (µg/dL) | ns | 429.7 ± 41.3 | 331.2 ± 67.9 | 414.2 ± 64.2 | 263.6 ± 103.8 | 358.3 ± 52.2 |
| TSAT (%) | ns | 65.3 ± 8.8 | 72.0 ± 18.5 | 92.7 ± 14.1 | 59.6 ± 25.0 | 77.4 ± 8.3 |
| pPD oxidase activity | ns | 133.3 ± 89.2 | 67.2 ± 1.4 | 48.6 ± 22.4 | 137.5 ± 94.8 | 69.3 ± 2.2 |
| TIBC (µg/dL) | < 0.0001 | 660.6 ± 28.8 a | 466.1 ± 43.1 b | 446.8 ± 1.3 b | 448.9 ± 31.3 b | 461.9 ± 25.9 b |
| Dams 1 | Suckling Pups 2 | ||||
|---|---|---|---|---|---|
| Organ | % of BW 3 | Organ | % of BW 3 | Fe (p) 4 | Sex (p) 4 |
| Liver | 5.46 ± 0.47 | Liver | 3.24 ± 0.31 | p < 0.0001 | p = 0.0237 |
| Kidney | 0.69 ± 0.04 | Kidney | 1.00 ± 0.09 | p = 0.0201 | p = 0.0225 |
| Spleen | 0.26 ± 0.03 | Spleen | 0.46 ± 0.08 | p = 0.0005 | ns |
| Heart | 0.42 ± 0.02 | Heart | 0.57 ± 0.07 | p < 0.0001 | ns |
| Organ 1 | % of BW 2 | Liver % of BW 1–3 | Diet |
|---|---|---|---|
| Kidney | 0.73 ± 0.06 | 4.77 ± 0.41 b | 1× |
| Spleen | 0.23 ± 0.03 | 4.63 ± 0.41 b | 3× |
| Heart | 0.37 ± 0.03 | 5.64 ± 0.57 ab | 5× |
| 5.77 ± 0.18 ab | 10× | ||
| 5.96 ± 0.70 a | 20× |
| Organ 1 | % of BW 2 | Heart % of BW 1−3 | Diet |
|---|---|---|---|
| Liver | 3.28 ± 0.29 | 0.62 ± 0.04 ab | 1× |
| Kidney | 1.28 ± 0.11 | 0.56 ± 0.07 b | 3× |
| Spleen | 0.48 ± 0.08 | 0.56 ± 0.08 b | 5× |
| 0.61± 0.07 ab | 10× | ||
| 0.72 ± 0.09 a | 20× |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.K.; Ha, J.-H.; Collins, J.F. Dietary Iron Intake in Excess of Requirements Impairs Intestinal Copper Absorption in Sprague Dawley Rat Dams, Causing Copper Deficiency in Suckling Pups. Biomedicines 2021, 9, 338. https://doi.org/10.3390/biomedicines9040338
Lee JK, Ha J-H, Collins JF. Dietary Iron Intake in Excess of Requirements Impairs Intestinal Copper Absorption in Sprague Dawley Rat Dams, Causing Copper Deficiency in Suckling Pups. Biomedicines. 2021; 9(4):338. https://doi.org/10.3390/biomedicines9040338
Chicago/Turabian StyleLee, Jennifer K., Jung-Heun Ha, and James F. Collins. 2021. "Dietary Iron Intake in Excess of Requirements Impairs Intestinal Copper Absorption in Sprague Dawley Rat Dams, Causing Copper Deficiency in Suckling Pups" Biomedicines 9, no. 4: 338. https://doi.org/10.3390/biomedicines9040338
APA StyleLee, J. K., Ha, J.-H., & Collins, J. F. (2021). Dietary Iron Intake in Excess of Requirements Impairs Intestinal Copper Absorption in Sprague Dawley Rat Dams, Causing Copper Deficiency in Suckling Pups. Biomedicines, 9(4), 338. https://doi.org/10.3390/biomedicines9040338