
Establishment of a Patient-Derived Xenograft Model of Colorectal Cancer in CIEA NOG Mice and Exploring Smartfish Liquid Diet as a Source of Omega-3 Fatty Acids

 , , ,
, , , 
Abstract
1. Introduction
2. Experimental Section
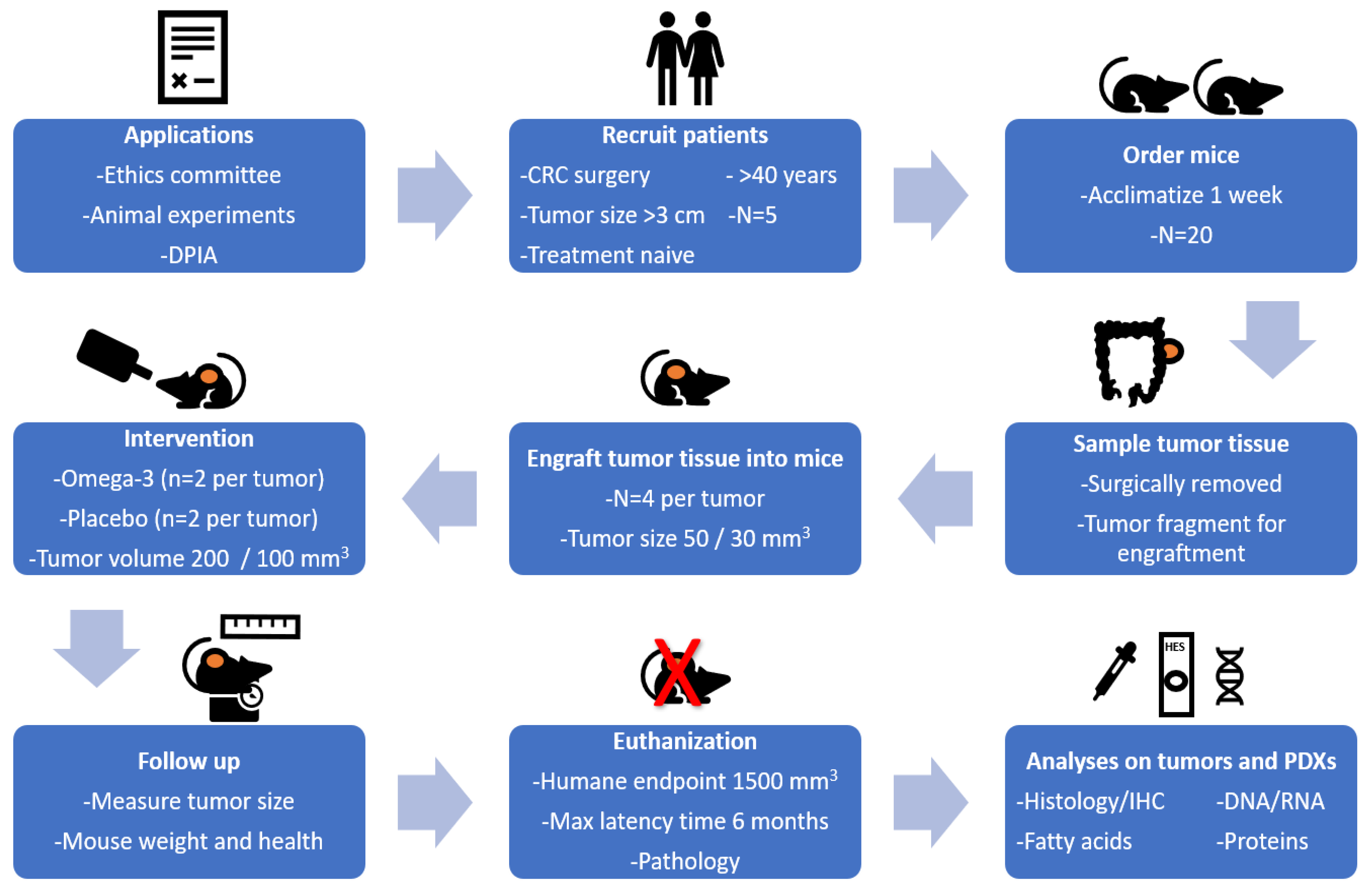
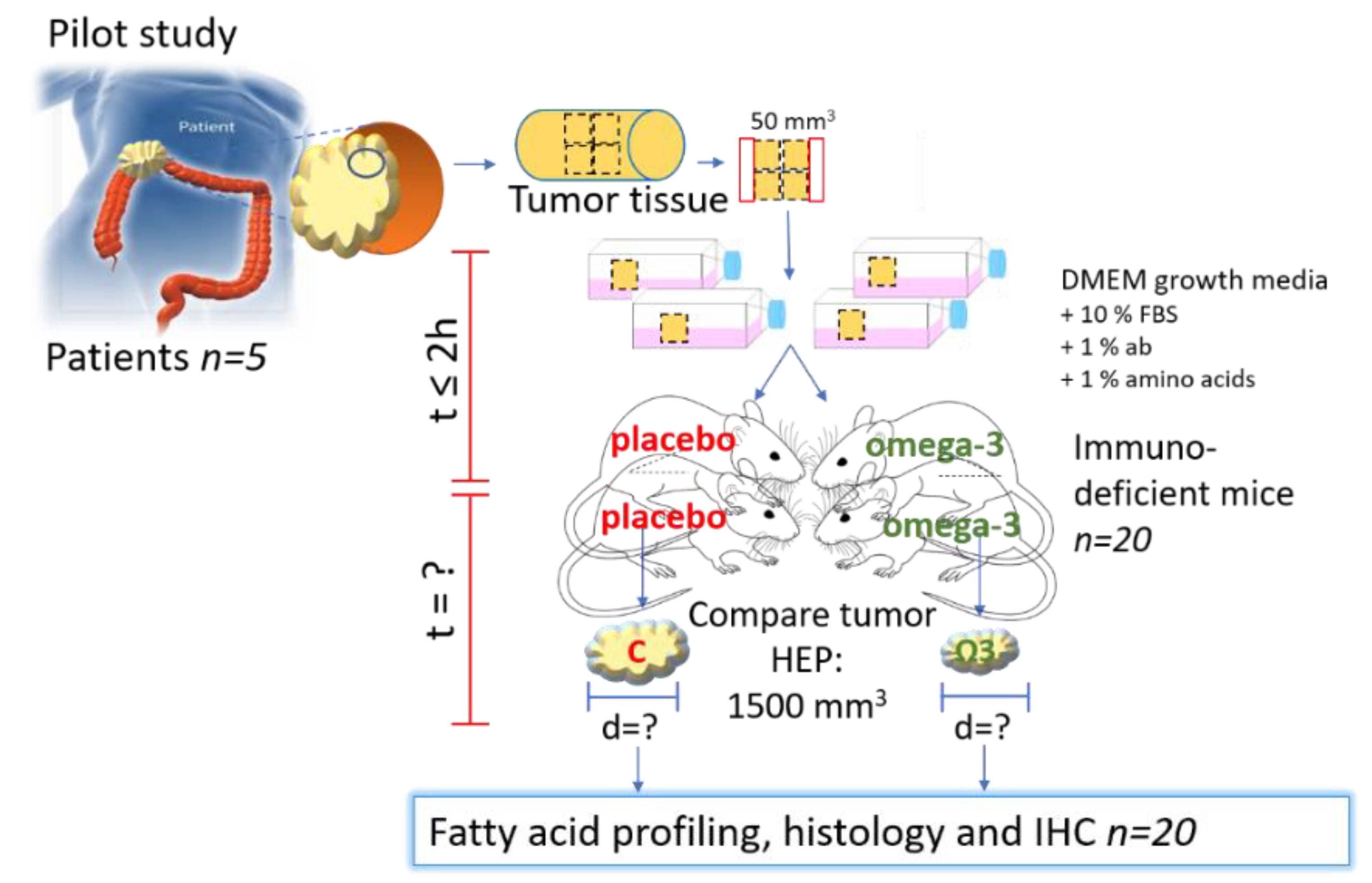
2.1. Study Design
2.2. Patient Characteristics and Inclusion Criteria
2.3. Patient Blood Sampling
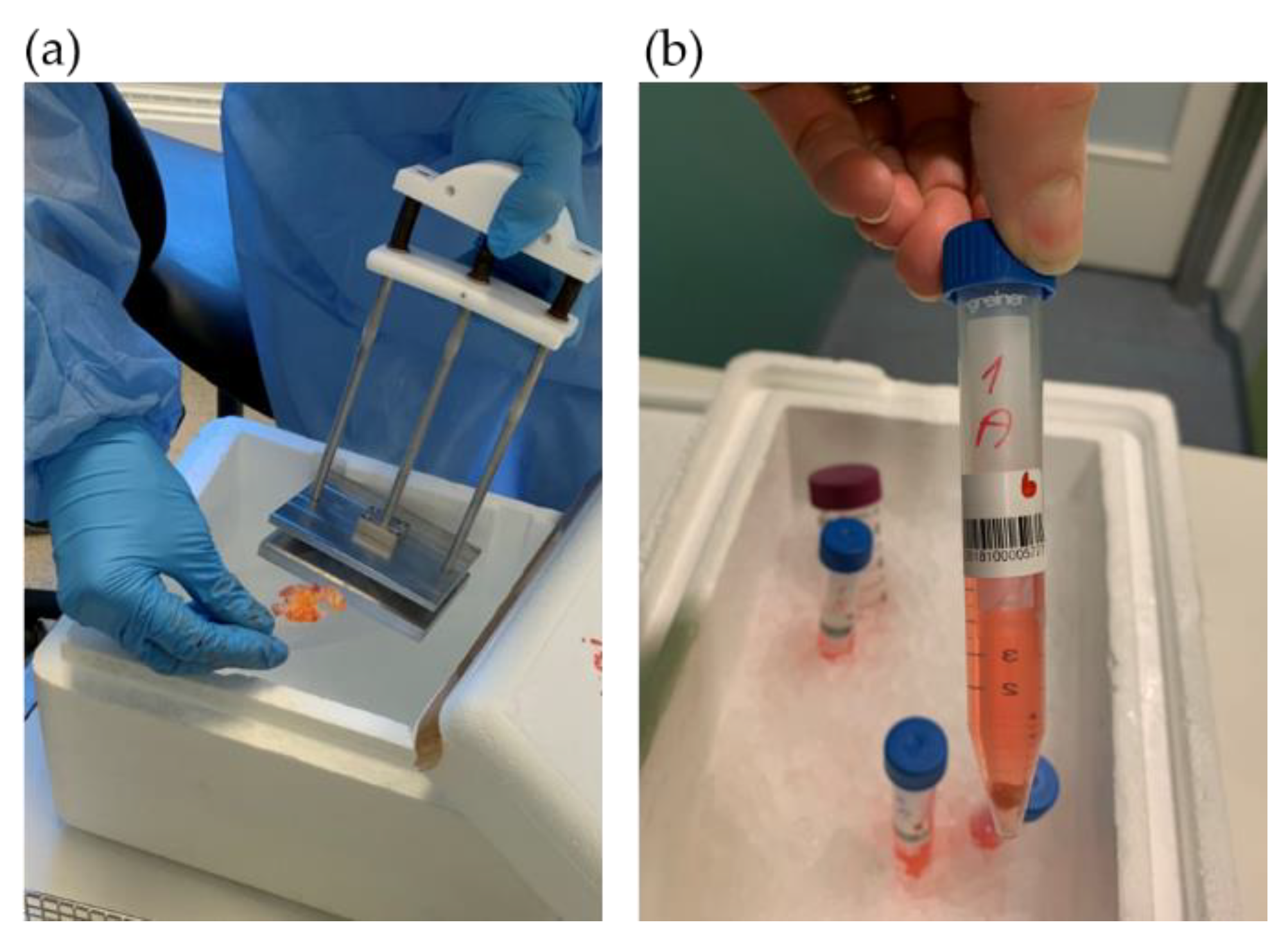
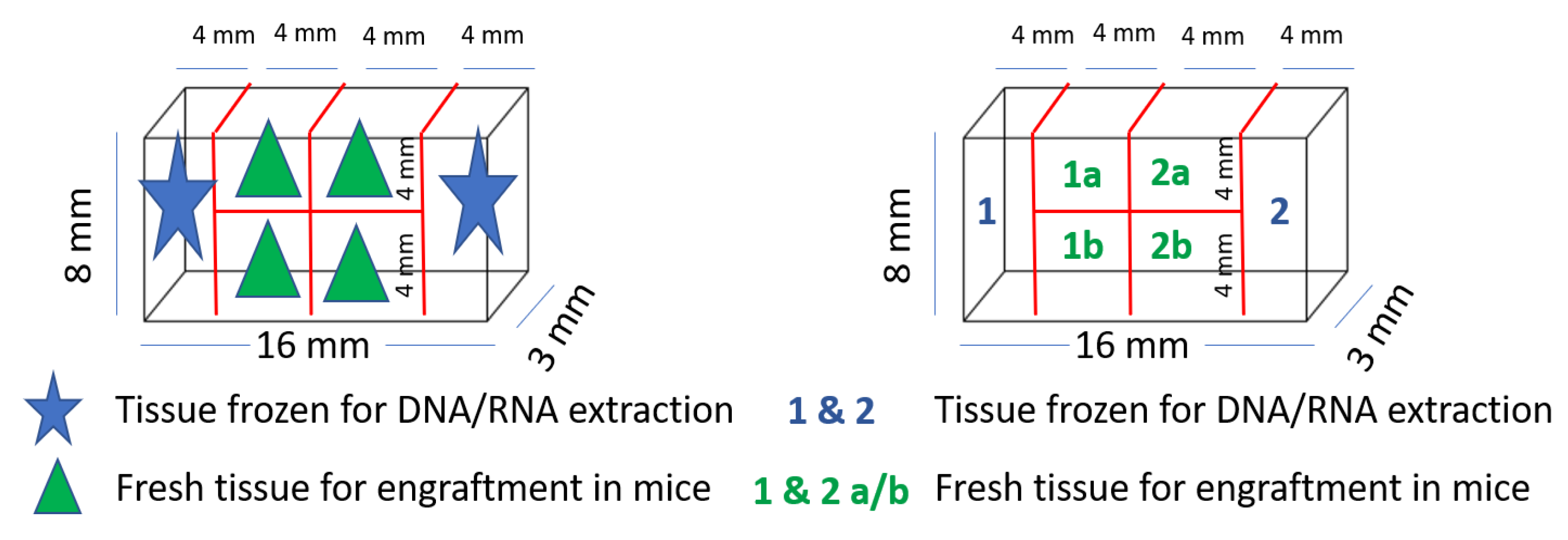
2.4. Collection of Colorectal Tumor and Healthy Tissue
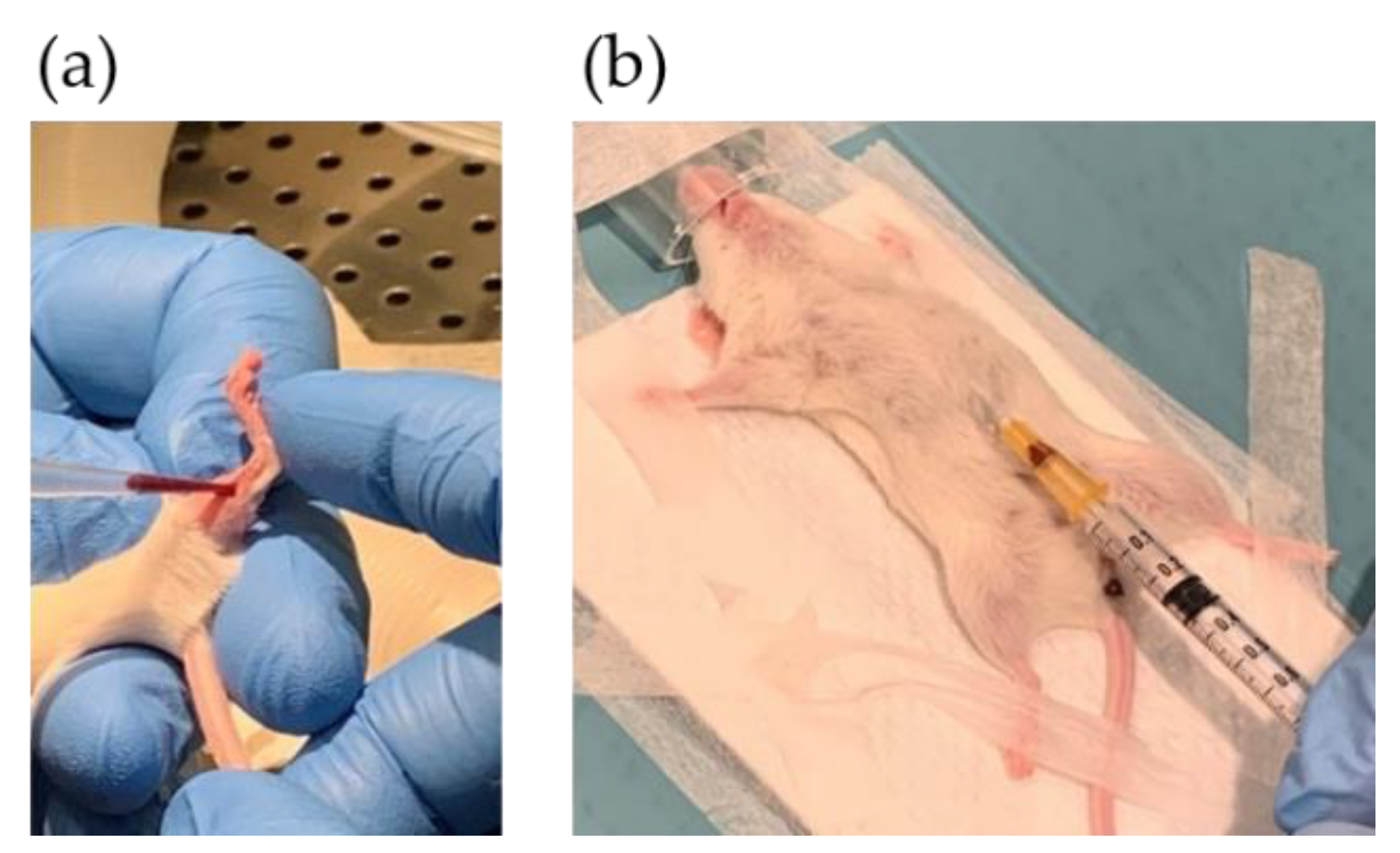
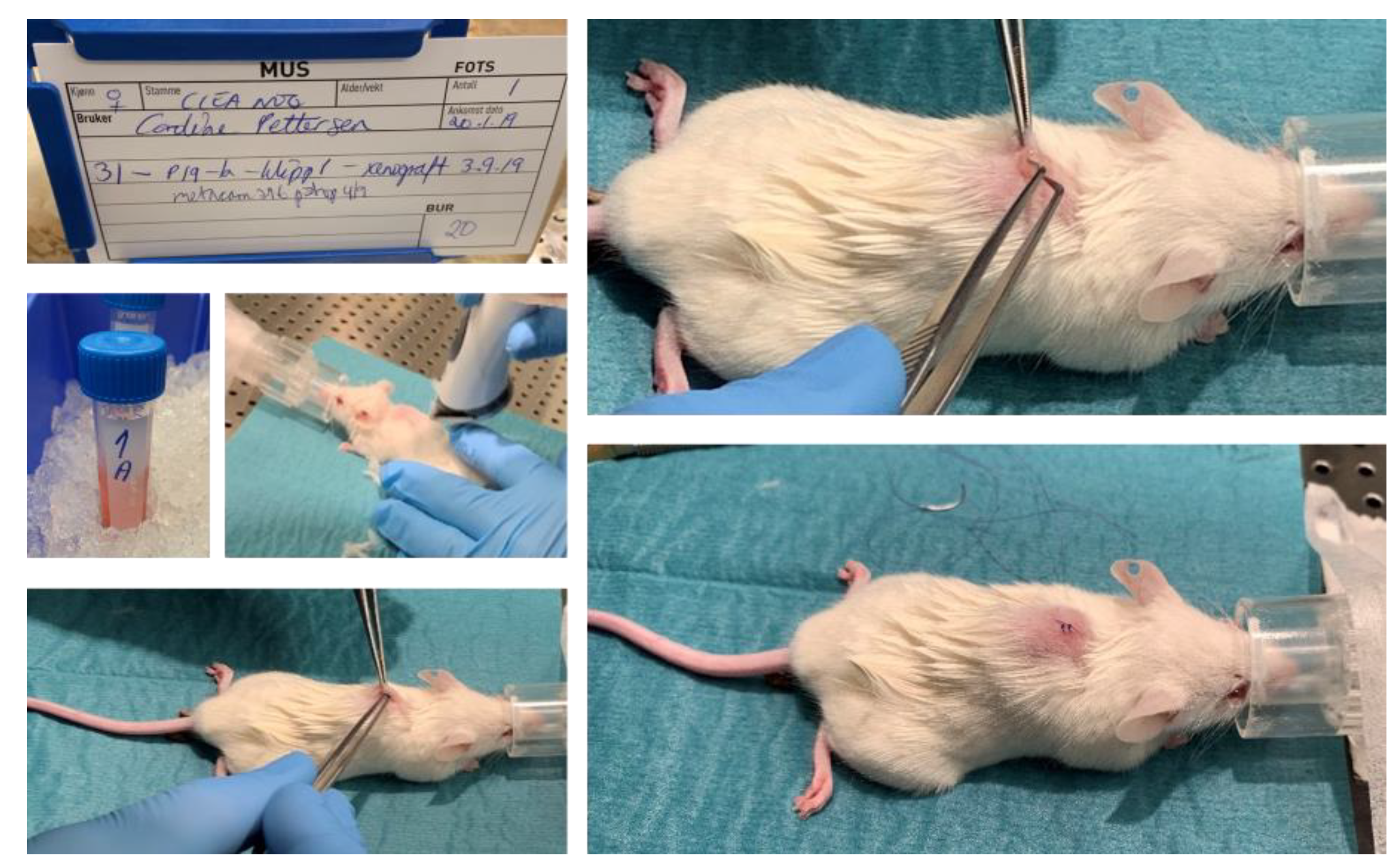
2.5. PDX Procedure in Immunodeficient Mice
2.6. Postoperative Follow-Up
2.7. Mouse Blood Sampling
2.8. Treatment with Liquid Diets
2.9. Euthanasia, Necropsy and Tissue Sampling
2.10. Fatty Acid Profiling and Omega-3 Index in Blood
2.11. Histology and Immunohistochemistry Analysis
2.12. In Situ Hybridization
2.13. Data Analysis
3. Results
3.1. Patient and Tumor Characteristics
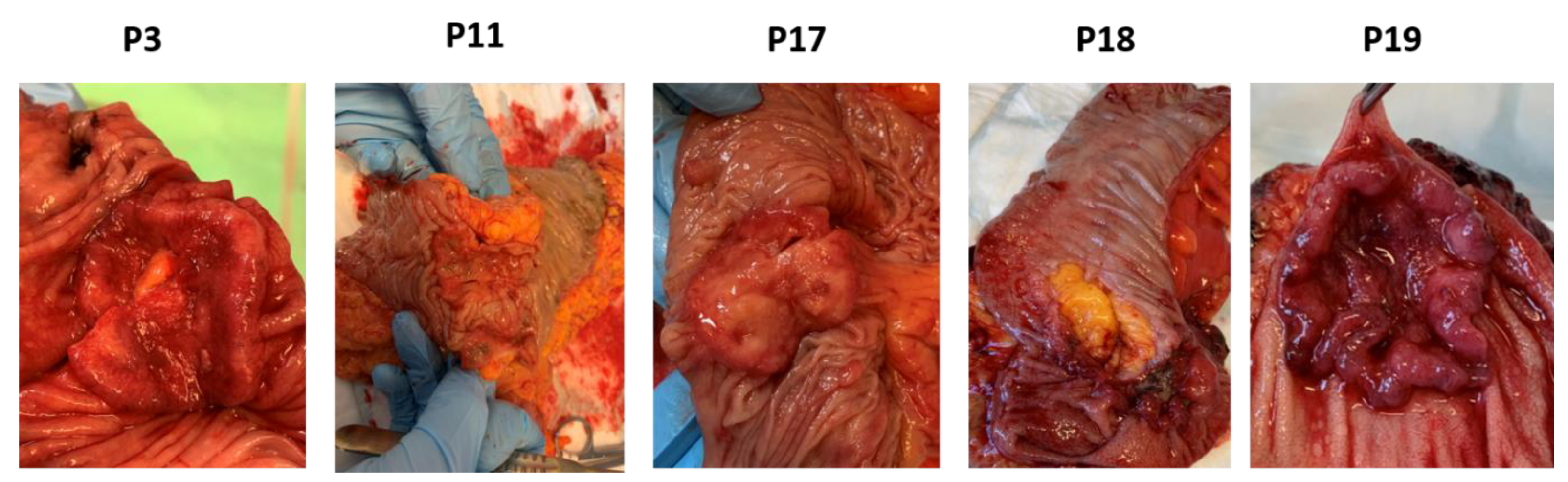
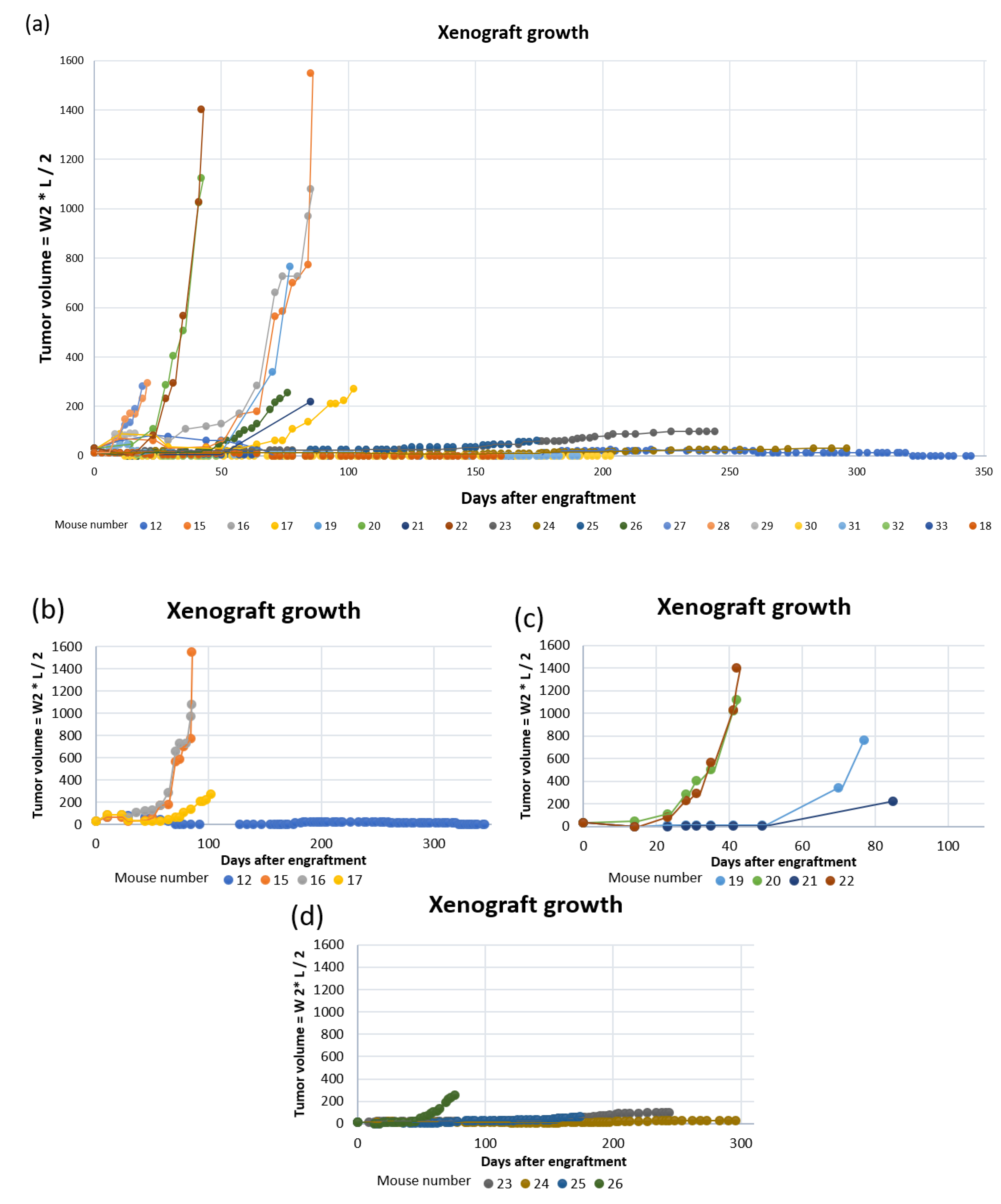
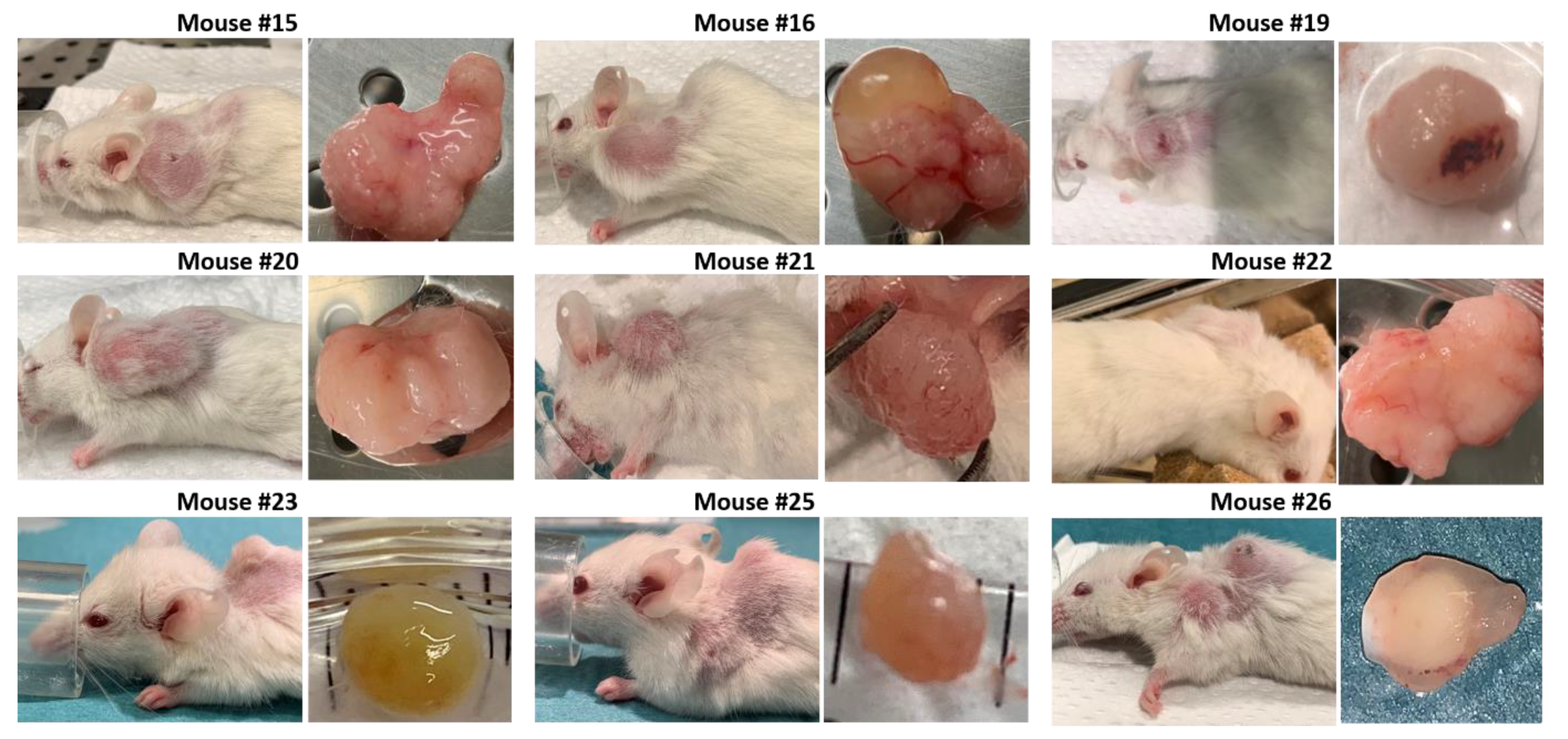
3.2. Establishment of Patient-Derived Xenografts of Colorectal Cancer
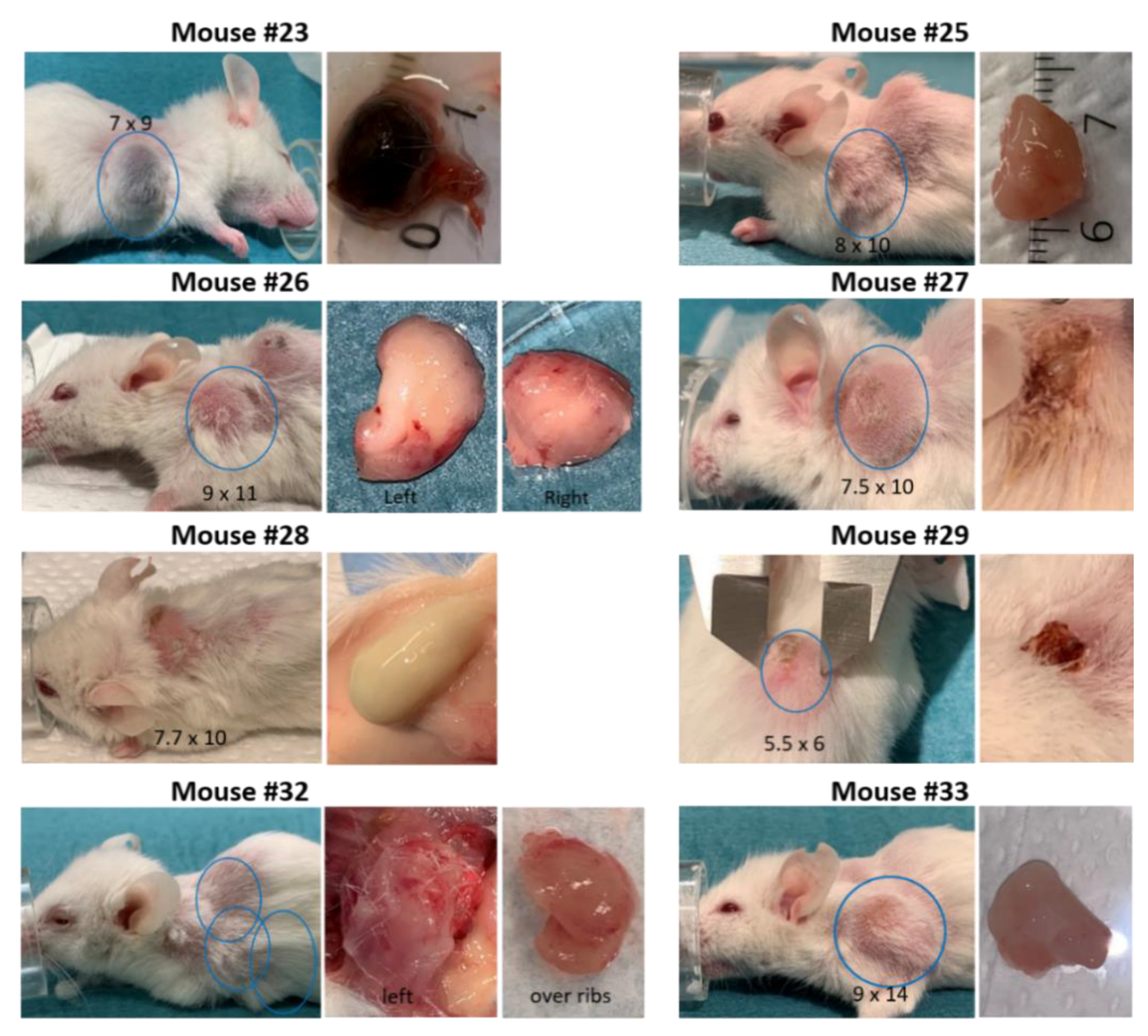
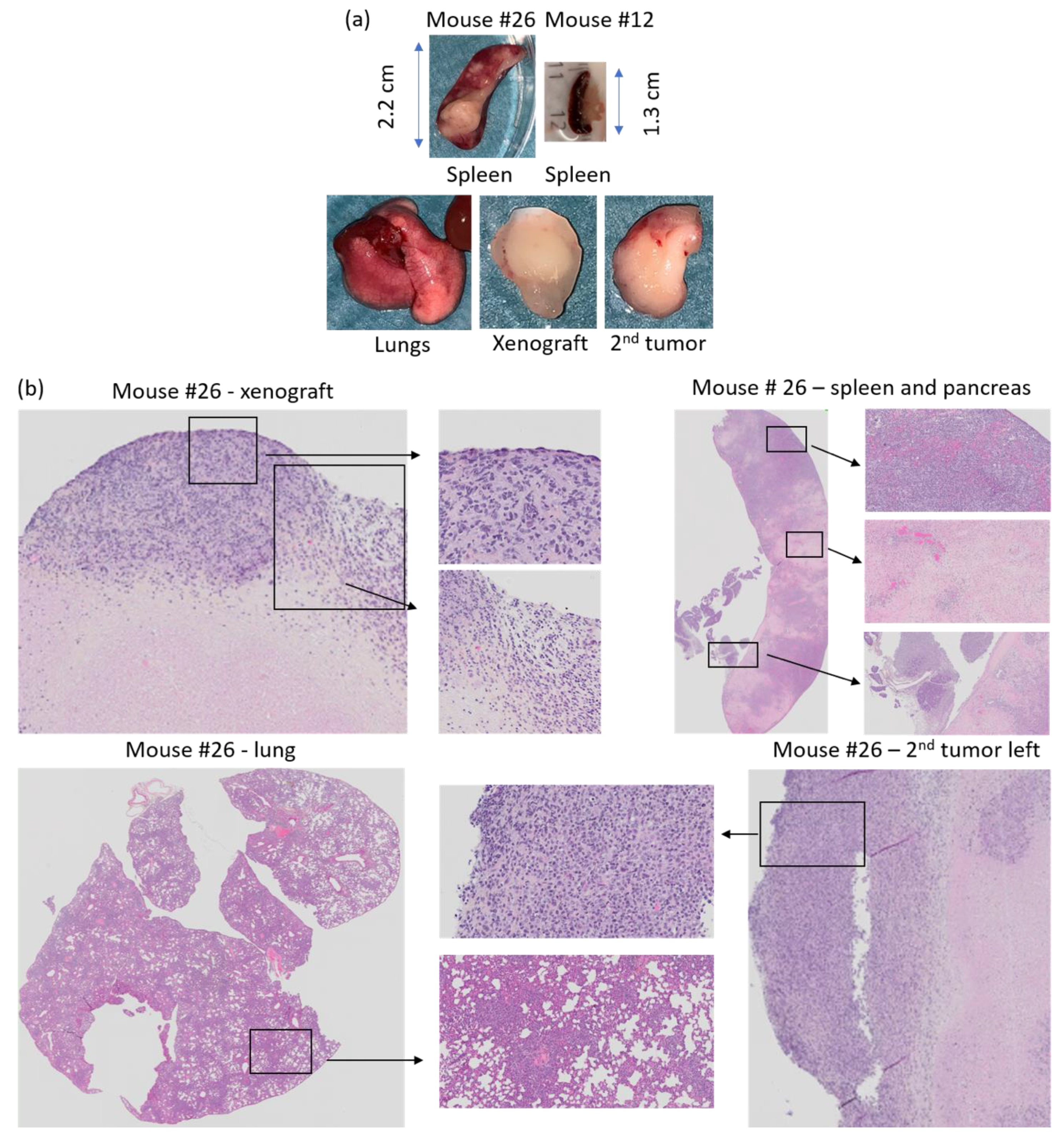
3.3. Development of Secondary Tumors and Abscesses
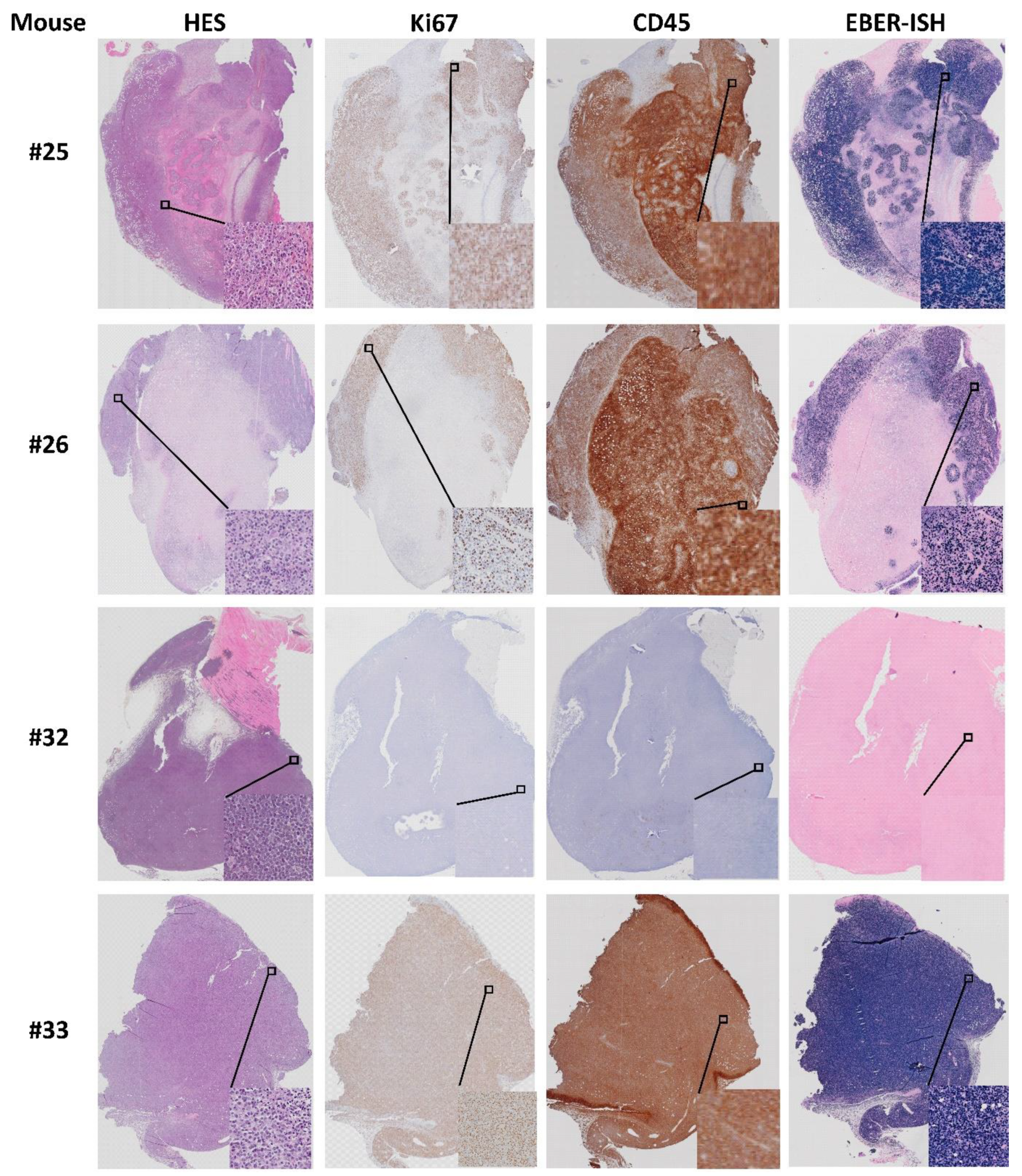
3.4. Histological Similarity between Patient Tumors and Patient-Derived Xenografts
3.5. Histology of Secondary Tumors and Affected Organs
3.6. Origin of Cells Found in Lymphomas and Presence of Epstein–Barr Virus
3.7. Intake of Liquid Diet
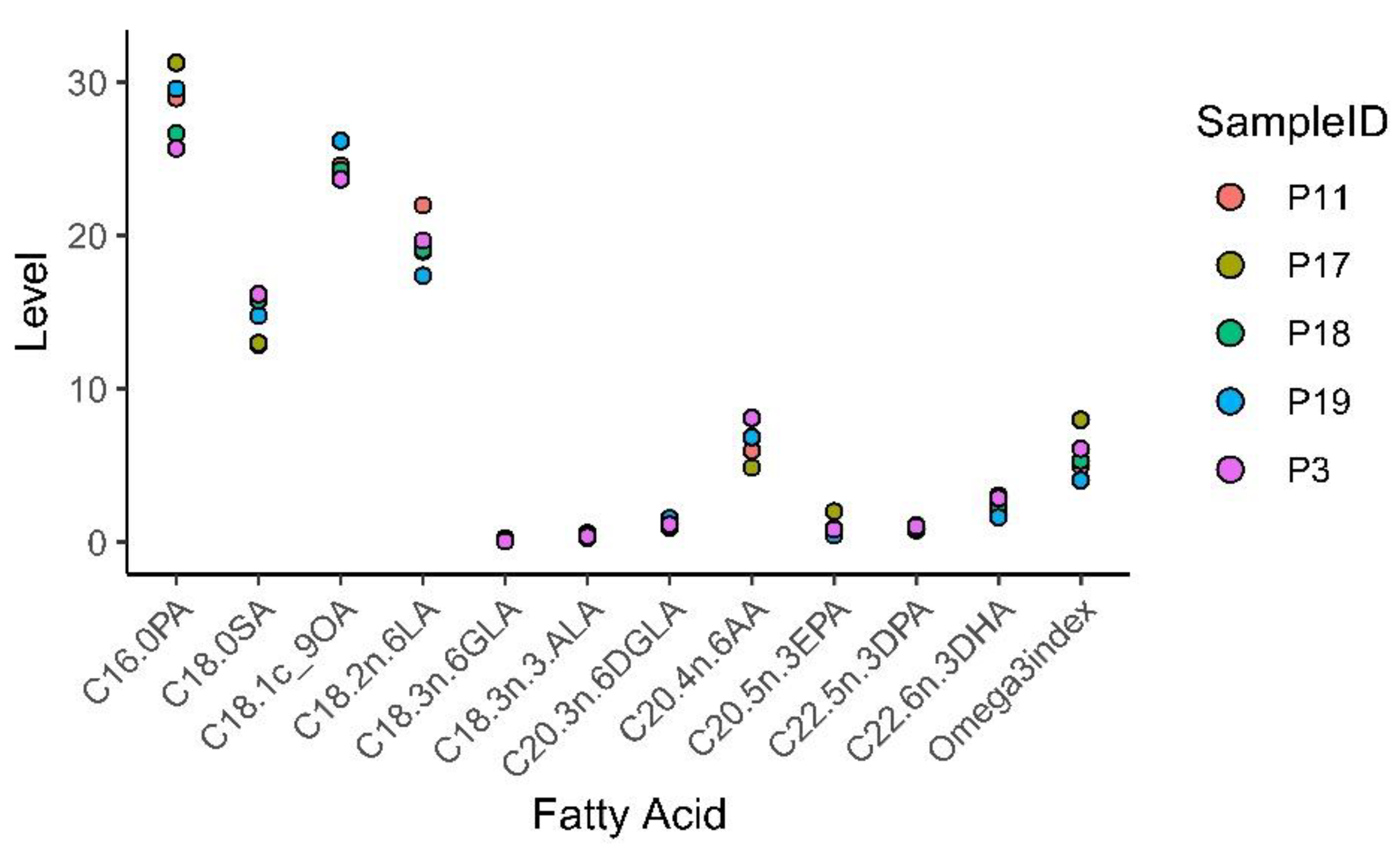
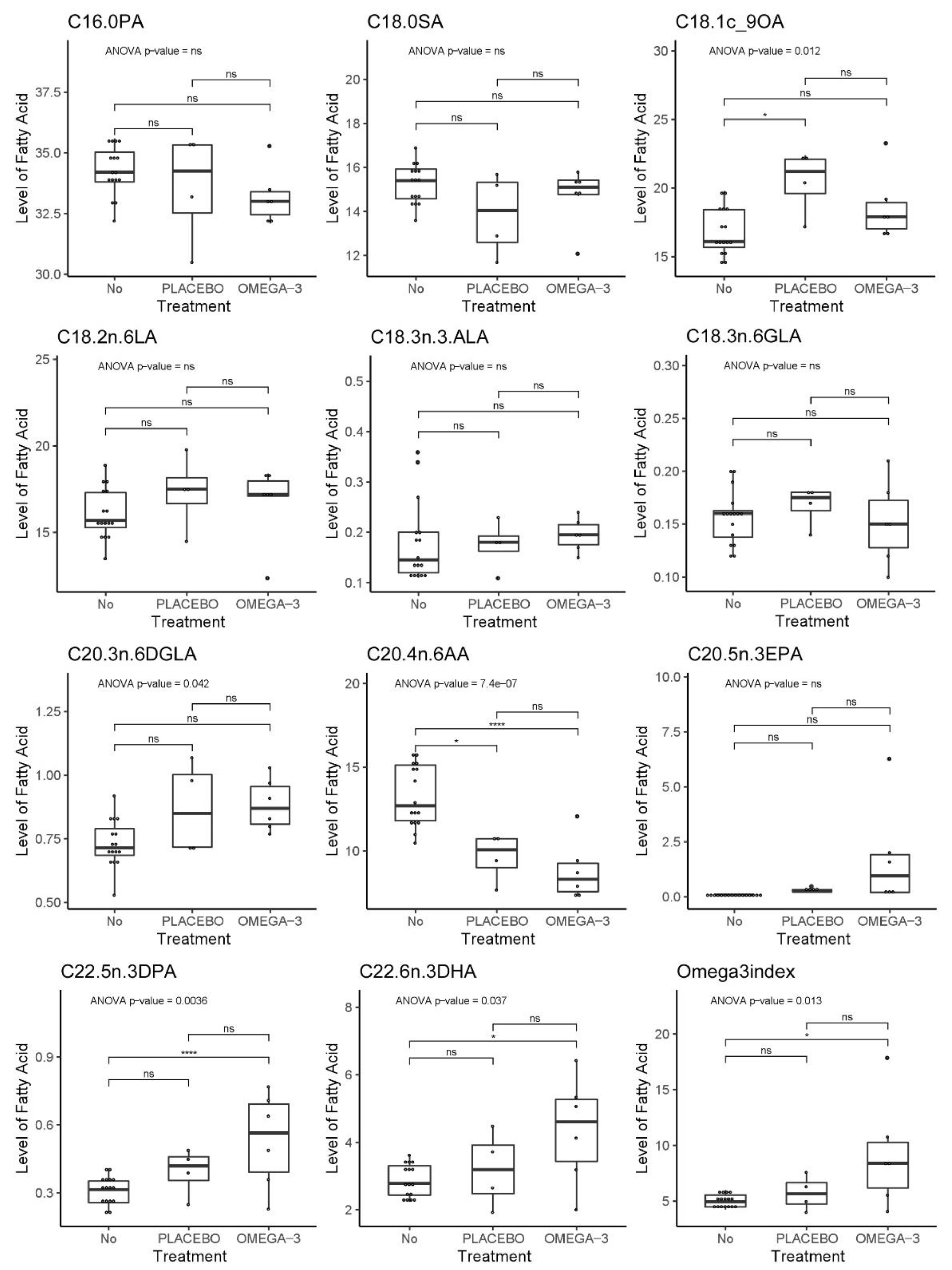
3.8. Fatty acid Profiling of Patient and Mouse Whole Blood
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Inoue, A.; Deem, A.K.; Kopetz, S.; Heffernan, T.P.; Draetta, G.F.; Carugo, A. Current and Future Horizons of Patient-Derived Xenograft Models in Colorectal Cancer Translational Research. Cancers 2019, 11, 1321. [Google Scholar] [CrossRef]
- Yoshida, G.J. Applications of patient-derived tumor xenograft models and tumor organoids. J. Hematol. Oncol. 2020, 13, 4. [Google Scholar] [CrossRef] [PubMed]
- Fujii, E.; Kato, A.; Suzuki, M. Patient-derived xenograft (PDX) models: Characteristics and points to consider for the process of establishment. J. Toxicol. Pathol. 2020, 33, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Cobb, L.M. The behaviour of carcinoma of the large bowel in man following transplantation into immune deprived mice. Br. J. Cancer 1973, 28, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Pickard, R.G.; Cobb, L.M.; Steel, G.G. The growth kinetics of xenografts of human colorectal tumours in immune deprived mice. Br. J. Cancer 1975, 31, 36–45. [Google Scholar] [CrossRef][Green Version]
- Brown, K.M.; Xue, A.; Mittal, A.; Samra, J.S.; Smith, R.; Hugh, T.J. Patient-derived xenograft models of colorectal cancer in pre-clinical research: A systematic review. Oncotarget 2016, 7, 66212–66225. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Hiramatsu, H.; Kobayashi, K.; Suzue, K.; Kawahata, M.; Hioki, K.; Ueyama, Y.; Koyanagi, Y.; Sugamura, K.; Tsuji, K.; et al. NOD/SCID/gamma(c)(null) mouse: An excellent recipient mouse model for engraftment of human cells. Blood 2002, 100, 3175–3182. [Google Scholar] [CrossRef]
- Shultz, L.D.; Goodwin, N.; Ishikawa, F.; Hosur, V.; Lyons, B.L.; Greiner, D.L. Human cancer growth and therapy in immunodeficient mouse models. Cold Spring Harb. Protoc. 2014, 2014, 694–708. [Google Scholar] [CrossRef]
- Kato, C.; Fujii, E.; Chen, Y.J.; Endaya, B.B.; Matsubara, K.; Suzuki, M.; Ohnishi, Y.; Tamaoki, N. Spontaneous thymic lymphomas in the non-obese diabetic/Shi-scid, IL-2R gamma (null) mouse. Lab. Anim. 2009, 43, 402–404. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, M.; Ogura, T.; Goto, T.; Yagoto, M.; Kamai, Y.; Shimomura, C.; Hayashimoto, N.; Kiyokawa, Y.; Shinohara, H.; Takahashi, R.; et al. Incidence of spontaneous lymphomas in non-experimental NOD/Shi-scid, IL-2Rgamma(null) (NOG) mice. Exp. Anim. 2017, 66, 425–435. [Google Scholar] [CrossRef]
- Katsiampoura, A.; Raghav, K.; Jiang, Z.Q.; Menter, D.G.; Varkaris, A.; Morelli, M.P.; Manuel, S.; Wu, J.; Sorokin, A.V.; Rizi, B.S.; et al. Modeling of Patient Derived Xenografts in Colorectal Cancer. Mol. Cancer Ther. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Larsen, I.K. (Ed.) Cancer in Norway 2016: Cancer Incidence, Mortality, Survival and Prevalence in Norway; Cancer Registry of Norway: Oslo, Norway, 2017. [Google Scholar]
- Constant, S.; Huang, S.; Wiszniewski, L.; Mas, C. Colon Cancer: Current Treatments and Preclinical Models for the Discovery and Development of New Therapies. In Drug Discovery; El-Shemy, H.A., Ed.; IntechOpen: Rijeka, Croatia, 2013. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef]
- Covelli, V. Guide to the Necroscopy of the Mouse. Available online: http://eulep.pdn.cam.ac.uk/Necropsy_of_the_Mouse/printable.php (accessed on 2 January 2020).
- Fujii, E.; Kato, A.; Chen, Y.J.; Matsubara, K.; Ohnishi, Y.; Suzuki, M. Characterization of EBV-related lymphoproliferative lesions arising in donor lymphocytes of transplanted human tumor tissues in the NOG mouse. Exp. Anim. 2014, 63, 289–296. [Google Scholar] [CrossRef]
- Dierge, E.; Larondelle, Y.; Feron, O. Cancer diets for cancer patients: Lessons from mouse studies and new insights from the study of fatty acid metabolism in tumors. Biochimie 2020. [Google Scholar] [CrossRef] [PubMed]
- Abdirahman, S.M.; Christie, M.; Preaudet, A.; Burstroem, M.C.U.; Mouradov, D.; Lee, B.; Sieber, O.M.; Putoczki, T.L. A Biobank of Colorectal Cancer Patient-Derived Xenografts. Cancers 2020, 12, 2340. [Google Scholar] [CrossRef]
- Cho, Y.B.; Hong, H.K.; Choi, Y.L.; Oh, E.; Joo, K.M.; Jin, J.; Nam, D.H.; Ko, Y.H.; Lee, W.Y. Colorectal cancer patient-derived xenografted tumors maintain characteristic features of the original tumors. J. Surg. Res. 2014, 187, 502–509. [Google Scholar] [CrossRef]
- Chijiwa, T.; Kawai, K.; Noguchi, A.; Sato, H.; Hayashi, A.; Cho, H.; Shiozawa, M.; Kishida, T.; Morinaga, S.; Yokose, T.; et al. Establishment of patient-derived cancer xenografts in immunodeficient NOG mice. Int. J. Oncol. 2015, 47, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Wimsatt, J.H.; Montgomery, C.; Thomas, L.S.; Savard, C.; Tallman, R.; Innes, K.; Jrebi, N. Assessment of a mouse xenograft model of primary colorectal cancer with special reference to perfluorooctane sulfonate. PeerJ 2018, 6, e5602. [Google Scholar] [CrossRef]
- Fujii, E.; Suzuki, M.; Matsubara, K.; Watanabe, M.; Chen, Y.J.; Adachi, K.; Ohnishi, Y.; Tanigawa, M.; Tsuchiya, M.; Tamaoki, N. Establishment and characterization of in vivo human tumor models in the NOD/SCID/gamma(c)(null) mouse. Pathol. Int. 2008, 58, 559–567. [Google Scholar] [CrossRef]
- Collins, A.T.; Lang, S.H. A systematic review of the validity of patient derived xenograft (PDX) models: The implications for translational research and personalised medicine. PeerJ 2018, 6, e5981. [Google Scholar] [CrossRef]
- Fleming, M.; Ravula, S.; Tatishchev, S.F.; Wang, H.L. Colorectal carcinoma: Pathologic aspects. J. Gastrointest. Oncol. 2012, 3, 153–173. [Google Scholar] [CrossRef]
- Itoh, T.; Shiota, M.; Takanashi, M.; Hojo, I.; Satoh, H.; Matsuzawa, A.; Moriyama, T.; Watanabe, T.; Hirai, K.; Mori, S. Engraftment of human non-Hodgkin lymphomas in mice with severe combined immunodeficiency. Cancer 1993, 72, 2686–2694. [Google Scholar] [CrossRef]
- Bondarenko, G.; Ugolkov, A.; Rohan, S.; Kulesza, P.; Dubrovskyi, O.; Gursel, D.; Mathews, J.; O’Halloran, T.V.; Wei, J.J.; Mazar, A.P. Patient-Derived Tumor Xenografts Are Susceptible to Formation of Human Lymphocytic Tumors. Neoplasia 2015, 17, 735–741. [Google Scholar] [CrossRef]
- Butler, K.A.; Hou, X.; Becker, M.A.; Zanfagnin, V.; Enderica-Gonzalez, S.; Visscher, D.; Kalli, K.R.; Tienchaianada, P.; Haluska, P.; Weroha, S.J. Prevention of Human Lymphoproliferative Tumor Formation in Ovarian Cancer Patient-Derived Xenografts. Neoplasia 2017, 19, 628–636. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Lee, J.E.; Kim, H.; Sim, M.H.; Kim, K.K.; Lee, G.; Kim, H.I.; An, J.Y.; Hyung, W.J.; Kim, C.B.; et al. Establishment and characterisation of patient-derived xenografts as paraclinical models for gastric cancer. Sci. Rep. 2016, 6, 22172. [Google Scholar] [CrossRef]
- Mukohyama, J.; Iwakiri, D.; Zen, Y.; Mukohara, T.; Minami, H.; Kakeji, Y.; Shimono, Y. Evaluation of the risk of lymphomagenesis in xenografts by the PCR-based detection of EBV BamHI W region in patient cancer specimens. Oncotarget 2016, 7, 50150–50160. [Google Scholar] [CrossRef]
- Radaelli, E.; Hermans, E.; Omodho, L.; Francis, A.; Vander Borght, S.; Marine, J.C.; van den Oord, J.; Amant, F. Spontaneous Post-Transplant Disorders in NOD.Cg- Prkdcscid Il2rgtm1Sug/JicTac (NOG) Mice Engrafted with Patient-Derived Metastatic Melanomas. PLoS ONE 2015, 10, e0124974. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, Q.; Gupta, I.; Vranic, S.; Al Moustafa, A.E. Human Papillomaviruses and Epstein-Barr Virus Interactions in Colorectal Cancer: A Brief Review. Pathogens 2020, 9, 300. [Google Scholar] [CrossRef] [PubMed]
- Moyer, A.M.; Yu, J.; Sinnwell, J.P.; Dockter, T.J.; Suman, V.J.; Weinshilboum, R.M.; Boughey, J.C.; Goetz, M.P.; Visscher, D.W.; Wang, L. Spontaneous murine tumors in the development of patient-derived xenografts: A potential pitfall. Oncotarget 2019, 10, 3924–3930. [Google Scholar] [CrossRef] [PubMed]
- Samdal, H.; Sandmoe, M.A.; Olsen, L.C.; Jarallah, E.A.H.; Hoiem, T.S.; Schonberg, S.A.; Pettersen, C.H.H. Basal level of autophagy and MAP1LC3B-II as potential biomarkers for DHA-induced cytotoxicity in colorectal cancer cells. FEBS J. 2018. [Google Scholar] [CrossRef]
- Pettersen, K.; Monsen, V.T.; Hakvag Pettersen, C.H.; Overland, H.B.; Pettersen, G.; Samdal, H.; Tesfahun, A.N.; Lundemo, A.G.; Bjorkoy, G.; Schonberg, S.A. DHA-induced stress response in human colon cancer cells—Focus on oxidative stress and autophagy. Free. Radic. Biol. Med. 2016, 90, 158–172. [Google Scholar] [CrossRef]
- Eltweri, A.M.; Thomas, A.L.; Metcalfe, M.; Calder, P.C.; Dennison, A.R.; Bowrey, D.J. Potential applications of fish oils rich in omega-3 polyunsaturated fatty acids in the management of gastrointestinal cancer. Clin. Nutr. 2017, 36, 65–78. [Google Scholar] [CrossRef]
- Bathen, T.F.; Holmgren, K.; Lundemo, A.G.; Hjelstuen, M.H.; Krokan, H.E.; Gribbestad, I.S.; Schonberg, S.A. Omega-3 fatty acids suppress growth of SW620 human colon cancer xenografts in nude mice. Anticancer Res. 2008, 28, 3717–3723. [Google Scholar] [PubMed]
- Zou, S.; Meng, X.; Meng, Y.; Liu, J.; Liu, B.; Zhang, S.; Ding, W.; Wu, J.; Zhou, J. Microarray analysis of anti-cancer effects of docosahexaenoic acid on human colon cancer model in nude mice. Int. J. Clin. Exp. Med. 2015, 8, 5075–5084. [Google Scholar] [PubMed]
- Busquets, S.; Marmonti, E.; Oliva, F.; Simoes, E.; Luna, D.; Mathisen, J.S.; López-Soriano, F.J.; Öhlander, M.; Argilés, J.M. Omega-3 and omega-3/curcumin-enriched fruit juices decrease tumour growth and reduce muscle wasting in tumour-bearing mice. JCSM Rapid Commun. 2018, 1, 1–10. [Google Scholar] [CrossRef]
- Larsson, S.C.; Kumlin, M.; Ingelman-Sundberg, M.; Wolk, A. Dietary long-chain n-3 fatty acids for the prevention of cancer: A review of potential mechanisms. Am. J. Clin. Nutr. 2004, 79, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; Spencer, J.A.; Quirke, P.; Toogood, G.J.; Lawton, C.L.; et al. A randomised trial of the effect of omega-3 polyunsaturated fatty acid supplements on the human intestinal microbiota. Gut 2018, 67, 1974–1983. [Google Scholar] [CrossRef]
- Norwegian Food Safety Authority. Rapeseed Oil, The Norwegian Food Composition Table. Available online: www.matvaretabellen.no (accessed on 5 January 2020).
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health implications of high dietary omega-6 polyunsaturated Fatty acids. J. Nutr. Metab. 2012, 2012, 539426. [Google Scholar] [CrossRef]
- Rise, P.; Eligini, S.; Ghezzi, S.; Colli, S.; Galli, C. Fatty acid composition of plasma, blood cells and whole blood: Relevance for the assessment of the fatty acid status in humans. Prostaglandins Leukot. Essent. Fat. Acids 2007, 76, 363–369. [Google Scholar] [CrossRef]
- Liu, G.; Muhlhausler, B.S.; Gibson, R.A. A method for long term stabilisation of long chain polyunsaturated fatty acids in dried blood spots and its clinical application. Prostaglandins Leukot. Essent. Fat. Acids 2014, 91, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.L.; Polreis, J. Measurement of the Omega-3 Index in Dried Blood Spots. Ann. Clin. Lab. Res. 2016, 4, 1–7. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient 3 | Patient 11 | Patient 17 | Patient 18 | Patient 19 | |
|---|---|---|---|---|---|
| Gender (Male/Female) | M | F | M | M | M |
| Age | 69 | 65 | 77 | 62 | 65 |
| Tumor site | Colon | Colon | Colon | Rectum | Rectum |
| Tumor anatomy | Exophytic | Stenosing, ulcerating | Stricturating, ulcerating | Ulcerating | Exophytic, ulcerating |
| Tumor type | Adenocarcinoma | Signet cell carcinoma | Adenocarcinoma with mucus | Adenocarcinoma | Adenocarcinoma |
| Tumor stage (TMN) | T3N0M0 | T3N2M0 | T4N0M0 | T4aN1M0 | T3N0M0 |
| Tumor differentiation grade | Medium | Medium | Medium | Medium | |
| Tumor size (cm) | 4 | 5 | 4 | 4 | 3.7 |
| Tumor MSI status | MSI-high | MSS | MSS | MSS | |
| Mutations | BRAF | BRAF | KRAS | ||
| Pre-operative cancer treatment | No | No | No | No | No |
| Previous cancer diagnoses | Yes | No | |||
| Intake of omega-3 supplements | 0–3/month | 0–3/month | 0–3/month | 0–3/month | 0–3/month |
| Intake of cod liver oil | 0–3/month | 0–3/month | Daily | 0–3/month | Daily 1 |
| Intake of fish | 1–3/week | 1–3/week | 1–3/week | 1–3/week | 1–3/week |
| Use of lipid reducing medication | No | No | No | Yes | No |
| Patient ID | Patient 3 | Patient 11 | Patient 17 | Patient 18 | Patient 19 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mouse ID | 12 | 15 | 16 | 17 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 18 |
| Tumor fragment ID | 1a | 1b | 2a | 2b | 1a | 1b | 2a | 2b | 1a | 1b | 2a | 2b | 1a | 1b | 2a | 2b | 1a | 1b | 2a | 2b |
| Ear clip | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 |
| Age at engraftment (week) | 11 | 11 | 11 | 11 | 17 | 17 | 17 | 17 | 31 | 31 | 31 | 31 | 31 | 31 | 31 | 31 | 33 | 33 | 33 | 33 |
| Tumor fragment size (mm3) | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 50 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 |
| Days to established growth | - | 64 | 64 | 99 | 70 | 34 | 91 | 34 | 185 | - | 174 | 52 | - | - | - | - | - | - | - | - |
| Mouse # | Clinical Symptoms & Comments | Necropsy Findings | Body Weight | PDX Size (mm) | Second Tumor Size (mm) |
|---|---|---|---|---|---|
| 12 | PDX not growing after 6 months. | Only a spot at engraftment side. Dark cystic structure close to pancreas. | Stable | - | - |
| 15 | Placebo treatment until PDX max size. | Large solid tumor. | Stable | 10 × 15.5 | - |
| 16 | Smartfish Remune treatment until PDX max size. | Large tumor, partly with liquid. Visible blood veins to tumor. | Stable | 11.5 × 13 | - |
| 17 | Smartfish Remune treatment until it died during anesthesia. | No samples taken. | Stable | 7.5 × 7.5 | - |
| 19 | Ulcerating xenograft. Smartfish Remune treatment. | Large solid tumor with blood traces. | Slightly reduced | 11.3 × 12 | - |
| 20 | Placebo treatment until PDX reached max size. | Large solid tumor. | −10–20% | 11.5 × 17 | - |
| 21 | Reduced general health and reduced weight. Large PDX. | Solid tumor. Low blood volume. No samples taken. | −10–15% | 7 × 9 | - |
| 22 | Smartfish Remune treatment until PDX max size. | Large solid tumor w/visible blood veins. | Stable | 11 × 17 | - |
| 23 | Reduced weight. Placebo treatment (8 weeks). Possibly rectal prolapse. | Small spleen. Second tumor with dark liquid inside. Clog of fur and food in stomach. | −10–20% | 5.8 × 5.9 | 7 × 9 |
| 24 | Did not reach “established growth” 3 months after animals without growing xenografts were euthanized. | Small slowly growing PDX. Whitish lungs. Normal organs. | Stable | 3.9 × 4 | - |
| 25 | Reduced general health. Large second tumor | Established growth of PDX. Whitish lungs. Large second tumor left shoulder. | Stable | 5 × 5 | 8 × 10 |
| 26 | Ulcerating xenograft. Smartfish Remune treatment. Large second tumor. | Enlarged spleen w/white fields. Whitish lungs. Two second tumors; left and right shoulder. | −10% | 8 × 8 | 9 × 11 (left) |
| 27 | Abscess mistaken for PDX until it burst. Smartfish Remune treatment. | Wound at the abscess site. Small xenograft under the skin. | Stable | - | - |
| 28 | Abscess mistaken as PDX in the beginning. Placebo treatment | Intact abscess 9.5 × 10 mm containing green liquid. Only a spot at engraftment site | Stable | - | - |
| 29 | Abscess mistaken as PDX until it burst. Smartfish Remune treatment | Wound where abscess has burst. Small xenograft under the skin | Stable | - | - |
| 30 | Did not reach “established growth” after 6 months. | Normal organs. A small bump in the liver. Trace of PDX under the skin | Stable | - | - |
| 31 | Did not reach “established growth” after 6 months. | Small xenograft under the skin. Normal organs. | Stable | - | - |
| 32 | Reduced general health. Second tumor. Did not reach “established growth”. Liquid in the eye. | Thick wounded skin at the neck. Whitish lungs. Enlarged spleen >2 cm. Red/swollen legs, shoulders and spine second tumor over the ribs. | Stable | - | - |
| 33 | Did not reach “established growth”. Large second tumor. | Large second tumor. No visible xenograft. Enlarged spleen. | Stable | - | 9 × 14 |
| 18 | Did not reach “established growth”. Rectal prolapse. | Swollen, bloody anal opening. Whitish part of one lung. Traces of xenograft under skin. Enlarged spleen ca 2.5 cm | −10% | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samdal, H.; Olsen, L.C.; Grøn, K.S.; Røyset, E.S.; Høiem, T.S.; Nervik, I.; Sætrom, P.; Wibe, A.; Schønberg, S.A.; Pettersen, C.H.H. Establishment of a Patient-Derived Xenograft Model of Colorectal Cancer in CIEA NOG Mice and Exploring Smartfish Liquid Diet as a Source of Omega-3 Fatty Acids. Biomedicines 2021, 9, 282. https://doi.org/10.3390/biomedicines9030282
Samdal H, Olsen LC, Grøn KS, Røyset ES, Høiem TS, Nervik I, Sætrom P, Wibe A, Schønberg SA, Pettersen CHH. Establishment of a Patient-Derived Xenograft Model of Colorectal Cancer in CIEA NOG Mice and Exploring Smartfish Liquid Diet as a Source of Omega-3 Fatty Acids. Biomedicines. 2021; 9(3):282. https://doi.org/10.3390/biomedicines9030282
Chicago/Turabian StyleSamdal, Helle, Lene C Olsen, Knut S Grøn, Elin S Røyset, Therese S Høiem, Ingunn Nervik, Pål Sætrom, Arne Wibe, Svanhild A Schønberg, and Caroline H H Pettersen. 2021. "Establishment of a Patient-Derived Xenograft Model of Colorectal Cancer in CIEA NOG Mice and Exploring Smartfish Liquid Diet as a Source of Omega-3 Fatty Acids" Biomedicines 9, no. 3: 282. https://doi.org/10.3390/biomedicines9030282
APA StyleSamdal, H., Olsen, L. C., Grøn, K. S., Røyset, E. S., Høiem, T. S., Nervik, I., Sætrom, P., Wibe, A., Schønberg, S. A., & Pettersen, C. H. H. (2021). Establishment of a Patient-Derived Xenograft Model of Colorectal Cancer in CIEA NOG Mice and Exploring Smartfish Liquid Diet as a Source of Omega-3 Fatty Acids. Biomedicines, 9(3), 282. https://doi.org/10.3390/biomedicines9030282