
Proteomic Profiling and Artificial Intelligence for Hepatocellular Carcinoma Translational Medicine

 ,
, 
Abstract
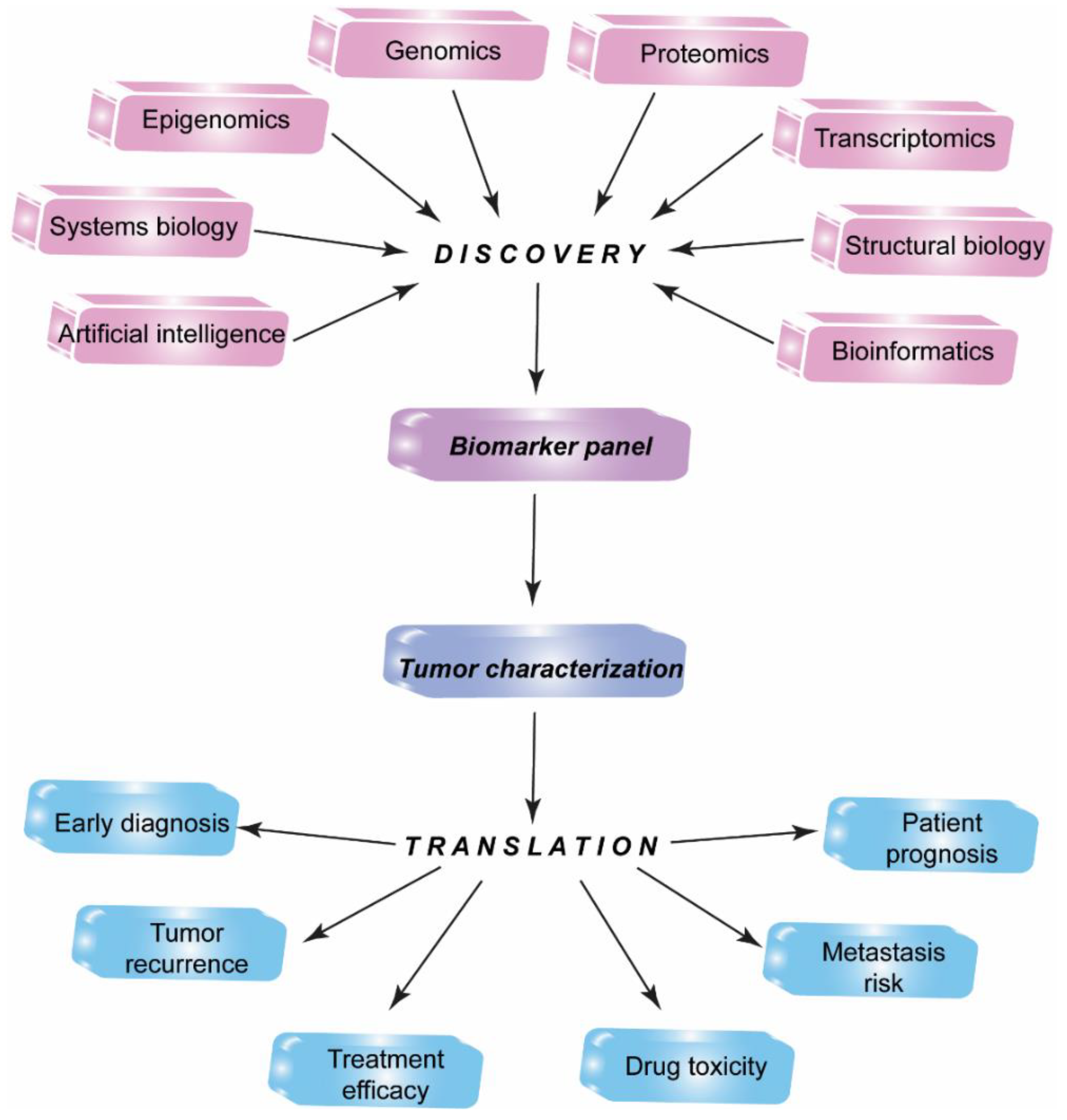
1. Introduction
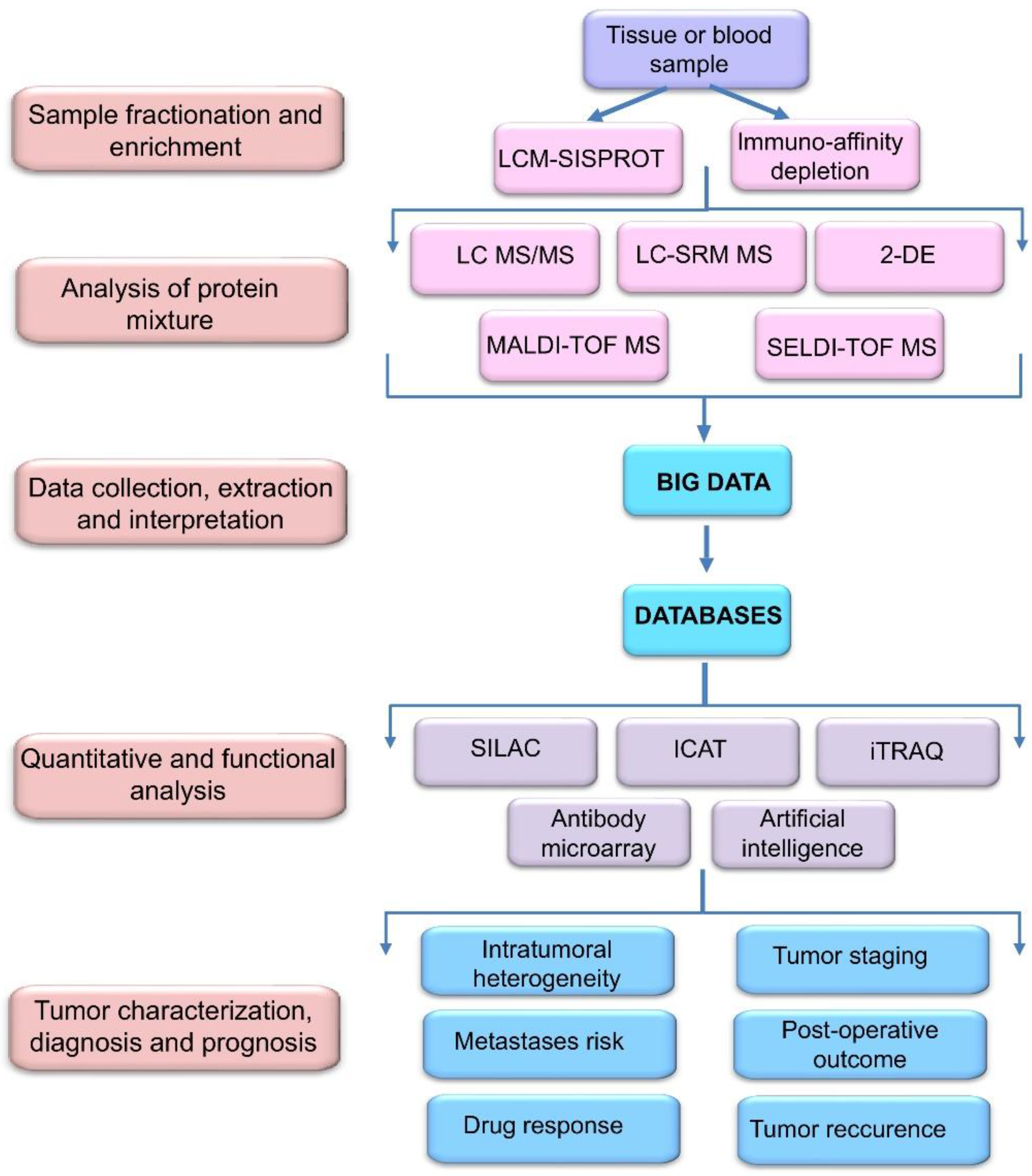
2. Proteomic Profiling Technologies and Big Data
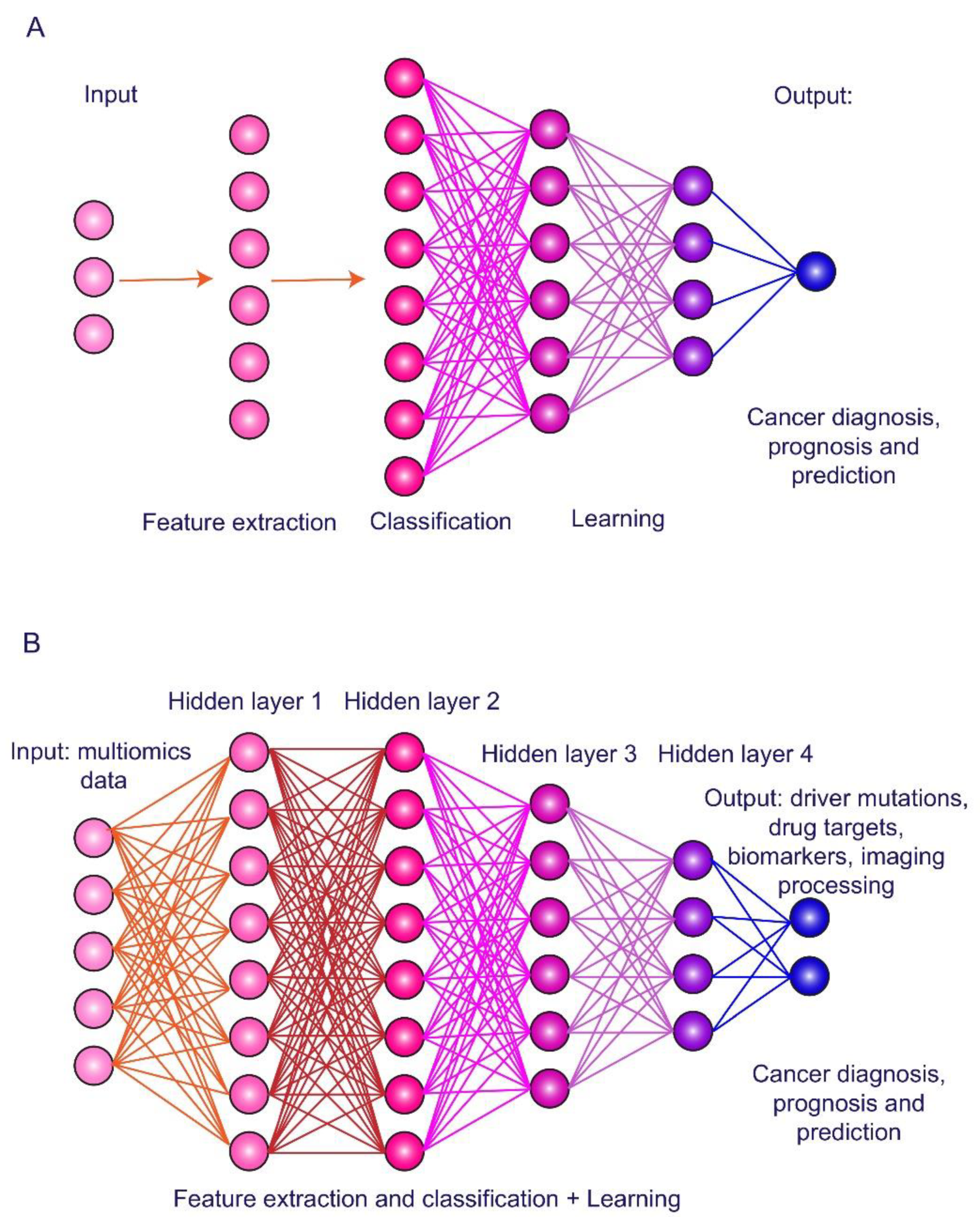
3. Artificial Intelligence in HCC Imaging and Biomarker Exploring
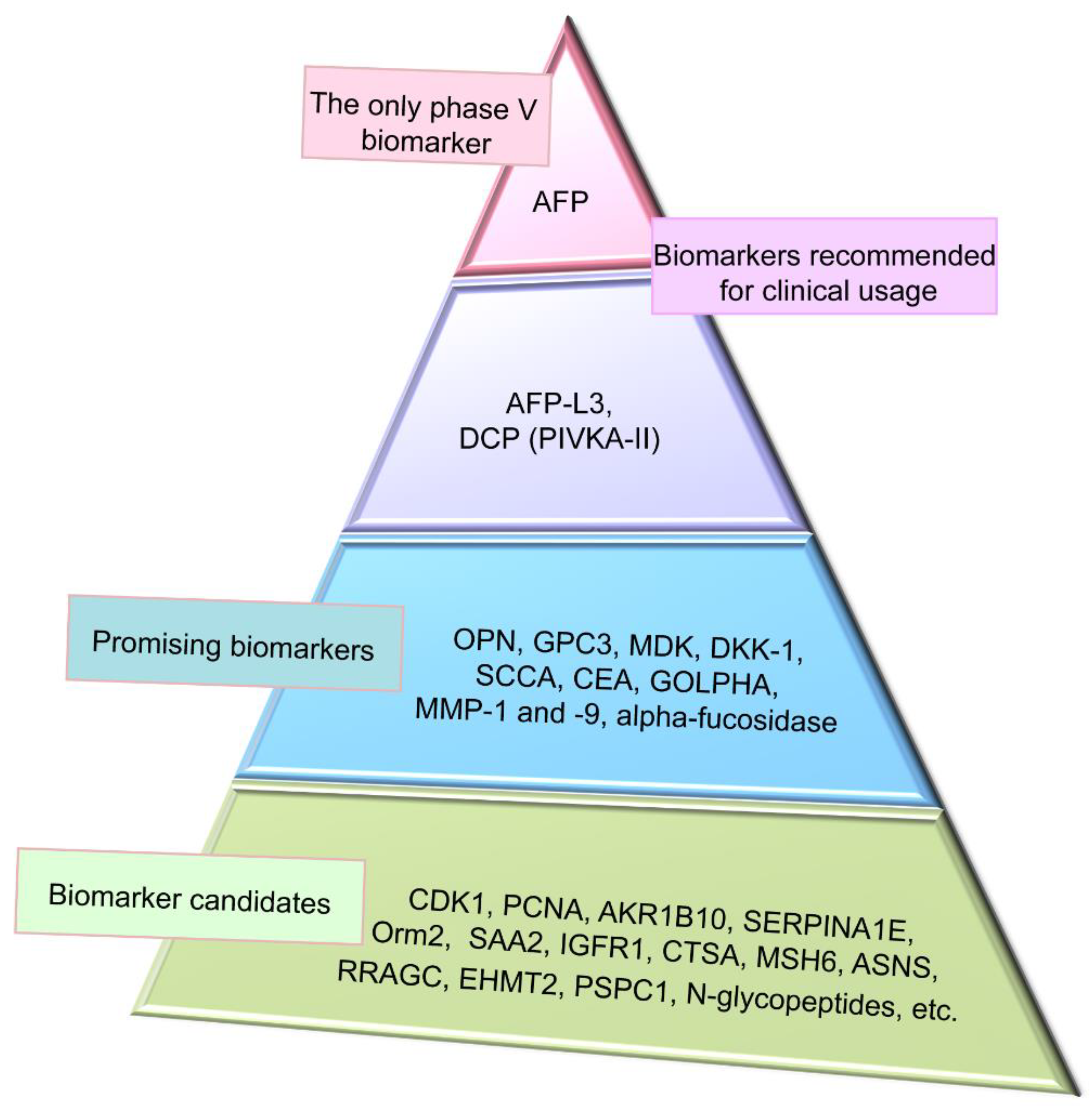
4. Conventional Biomarkers of Hepatocellular Carcinoma
4.1. Alpha-Fetoprotein and Its Glycoform
4.2. Des-Gamma-Carboxyprothrombin
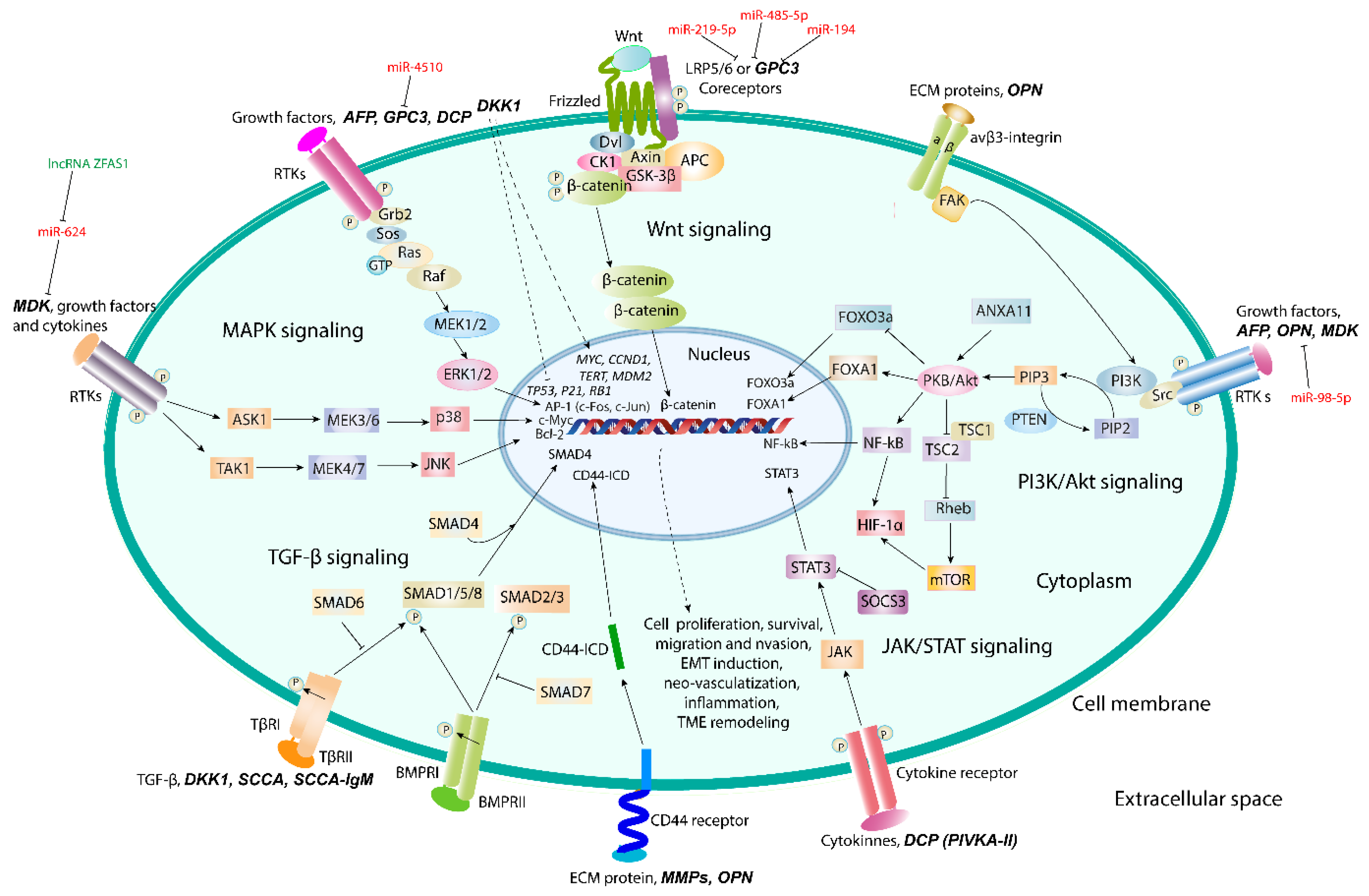
5. Promising Proteomic Biomarkers of HCC
5.1. Osteopontin
5.2. Glypican-3
5.3. Midkine
5.4. Dickkopf-1
5.5. Squamous Cell Carcinoma Antigen
6. Screening for Novel HCC Proteomic Biomarker Candidates
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kim, D.Y.; Han, K.H. Epidemiology and surveillance of hepatocellular carcinoma. Liver Cancer 2012, 1, 2–14. [Google Scholar] [CrossRef]
- Ferlay, J.; Colomber, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñerous, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed]
- El-Serag, H.B.; Rudolf, K.L. Hepatocellular carcinoma: Epidemiology and molecular carcinogenesis. Gastroenterology 2007, 132, 2557–2576. [Google Scholar] [CrossRef]
- Caldwell, S.; Park, S.H. The epidemiology of hepatocellular cancer: From the perspectives of public health problem to tumor biology. J. Gastroenterol. 2009, 44, 96–101. [Google Scholar] [CrossRef]
- Ayoub, W.S.; Steggerda, J.; Yang, J.D.; Kuo, A.; Sundaram, B.; Lu, S.C. Current status of hepatocellular carcinoma detection: Screening strategies and novel biomarkers. Ther. Adv. Med. Oncol. 2019, 11, 1758835919869120. [Google Scholar] [CrossRef]
- Osho, A.; Rich, N.E.; Singal, A.G. Role of imaging in management of hepatocellular carcinoma: Surveillance, diagnosis, and treatment response. Hepatoma Res. 2020, 6, 55. [Google Scholar] [CrossRef]
- Becker, A.K.; Tso, D.K.; Harris, A.C.; Malfair, D.; Chang, S.D. Extrahepatic metastases of hepatocellular carcinoma: A spectrum of imaging findings. Can. Assoc. Radiol. J. 2014, 65, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, D.R.; Fraum, T.J.; Cannella, R.; Tsai, R.; Naeem, M.; LeBlanc, M.; Salter, A.; Tsung, A.; Fleckenstein, J.; Shetty, A.S.; et al. Expanding the Liver Imaging Reporting and Data System (LI-RADS) v2018 diagnostic population: Performance and reliability of LI-RADS for distinguishing hepatocellular carcinoma (HCC) from non-HCC primary liver carcinoma in patients who do not meet strict LI-RADS high-risk criteria. HPB 2019, 21, 1697–1706. [Google Scholar]
- Nakamura, I.; Hatano, E.; Tada, M.; Kawabata, Y.; Tamagawa, S.; Kurimoto, A.; Iwama, H.; Toriguchi, K.; Sueoka, H.; Iida, K.; et al. Enhanced patterns on intraoperative contrast-enhanced ultrasonography predicts outcomes after curative liver resection in patients with hepatocellular carcinoma. Surg. Today 2020. [Google Scholar] [CrossRef] [PubMed]
- Poon, T.C.; Yip, T.T.; Chan, A.T.; Yap, C.; Yip, V.; Mok, T.S.; Lee, C.C.; Leung, T.W.; Ho, S.K.; Johnson, P.J. Comprehensive proteomic profiling identifies serum proteomic signatures for detection of hepatocellular carcinoma and its subtypes. Clin. Chem. 2003, 49, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yang, Z.; Li, G.; Li, C.; Luo, Y.; Gong, Q.; Wu, X.; Li, T.; Zhang, Z.; Xing, B.; et al. Multi-omics analysis of primary cell culture models reveals genetic and epigenetic basis of intratumoral phenotypic diversity. Genom. Proteom. Bioinform. 2019, 17, 576–589. [Google Scholar] [CrossRef]
- Woo, H.G.; Kim, Y.J. Multiplatform genomic roadmap of hepatocellular carcinoma: A matter of molecular heterogeneity. Hepatology 2018, 68, 2029–2032. [Google Scholar] [CrossRef]
- Ristevski, B.; Chen, M. Big data analytics in medicine and healthcare. J. Integr. Bioinform. 2018, 15, 20170030. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Grabowski, M.M.; Habboub, G.; Mroz, T.E. The impact of artificial intelligence on quality and safety. Glob. Spine J. 2020, 10, 99S–103S. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.K.; George, B.; Rai, V. Artificial intelligence to decode cancer mechanism: Beyond patient stratification for precision oncology. Front. Pharmacol. 2020, 11, 1177. [Google Scholar] [CrossRef] [PubMed]
- Tyers, M.; Mann, M. From genomics to proteomics. Nature 2003, 422, 193–197. [Google Scholar] [CrossRef]
- Duong, V.A.; Park, J.M.; Lee, H. Review of three-dimensional liquid chromatography platforms for bottom-up proteomics. Int. J. Mol. Sci. 2020, 21, 1524. [Google Scholar] [CrossRef] [PubMed]
- Vizcaíno, J.; Deutsch, E.; Wang, R.; Csordas, A.; Reisinger, F.; Rios, D.; Dianes, J.A.; Sun, Z.; Farrah, T.; Bandeira, N.; et al. ProteomeXchange provides globally coordinated proteomics data submission and dissemination. Nat. Biotechnol. 2014, 32, 223–226. [Google Scholar] [CrossRef]
- Martens, L.; Hermjakob, H.; Jones, P.; Adamski, M.; Taylor, C.; States, D.; Gevaert, K.; Vandekerckhove, J.; Apweiler, R. PRIDE: The proteomics identifications database. Proteomics 2005, 5, 3537–3545. [Google Scholar] [CrossRef]
- Deutsch, E.; Eng, J.; Zhang, H.; King, N.; Nesvizhskii, A.I.; Lin, B.; Lee, H.; Yi, E.; Ossola, R.; Aebersold, R. Human plasma peptide atlas. Proteomics 2005, 5, 3497–3500. [Google Scholar] [CrossRef]
- Cohrs, R.J.; Martin, T.; Ghahramani, P.; Bidaut, L.; Higgens, P.J.; Shahzad, A. Translational medicine definition from the European Society for Translational Medicine. New Horiz. Transl. Med. 2015, 2, 86–88. [Google Scholar]
- Meerzaman, D.; Dunn, B.K.; Lee, M.; Chen, Q.; Yan, C.; Ross, S. The promise of omics-based approaches to cancer prevention. Semin. Oncol. 2016, 43, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Poon, T.C.; Chan, A.T.; Zee, B.; Ho, S.K.; Mok, T.S.; Leung, T.W.; Johnson, P.J. Application of classification tree and neural network algorithms to the identification of serological liver marker profiles for the diagnosis of hepatocellular carcinoma. Oncology 2001, 61, 275–283. [Google Scholar] [CrossRef]
- Dayon, L.; Núñez, G.A.; Cominetti, O.; Corthésy, J.; Kussmann, M. A highly automated shotgun proteomic workflow: Clinical scale and robustness for biomarker discovery in blood. Methods Mol. Biol. 2017, 1619, 433–449. [Google Scholar] [PubMed]
- Xue, L.; Lin, L.; Zhou, W.; Chen, W.; Tang, J.; Sun, X.; Huang, P.; Tian, R. Mixed-mode ion exchange-based integrated proteomics technology for fast and deep plasma proteome profiling. J. Chromatogr. A 2018, 1564, 76–84. [Google Scholar] [CrossRef]
- Huang, P.; Kong, Q.; Gao, W.; Chu, B.; Li, H.; Mao, Y.; Cai, Z.; Xu, R.; Tian, R. Spatial proteome profiling by immunohistochemistry-based laser capture microdissection and data-dependent acquisition proteomics. Anal. Chim. Acta 2020, 1127, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Tang, J.; Deng, Q.; He, W.; Sun, X.; Xia, L.; Cheng, Z.; He, L.; You, S.; Hu, J.; et al. Spatial-resolution cell type proteome profiling of cancer tissue by fully integrated proteomics technology. Anal. Chem. 2018, 90, 5879–5886. [Google Scholar] [CrossRef]
- Hu, Y.; An, Q.; Sheu, K.; Trejo, B.; Fan, S.; Guo, Y. Single cell multi-omics technology: Methodology and application. Front. Cell Dev. Biol. 2018, 6, 28. [Google Scholar] [CrossRef]
- Mustafa, G.M.; Larry, D.; Petersen, J.R.; Elferink, C.J. Targeted proteomics for biomarker discovery and validation of hepatocellular carcinoma in hepatitis C infected patients. World J. Hepatol. 2015, 7, 1312–1324. [Google Scholar] [CrossRef]
- Rafalko, A.; Dai, S.; Hancock, W.S.; Karger, B.L.; Hincapie, M. Development of a Chip/Chip/SRM platform using digital chip isoelectric focusing and LC-Chip mass spectrometry for enrichment and quantitation of low abundance protein biomarkers in human plasma. J. Proteome Res. 2012, 11, 808–817. [Google Scholar] [CrossRef]
- Huang, X.; Zeng, Y.; Xing, X.; Zeng, J.; Cao, Y.; Cai, Z.; Xu, B.; Liu, X.; Huang, A.; Liu, J. Quantitative proteomics analysis of early recurrence/metastasis of huge hepatocellular carcinoma following radical resection. Proteome Sci. 2014, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Xue, R.; Huang, X.; Zhang, D.; Dong, L.; Wu, H.; Shen, X. Proteomic profiling of hepatitis B virus-related hepatocellular carcinoma with magnetic bead-based matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Acta Biochim. Biophys. Sinica 2011, 43, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, M.G.; Petersen, J.R.; Ju, H.; Cicalese, L.; Snyder, N.; Haidacher, S.J.; Denner, L.; Elferink, C. Biomarker discovery for early detection of hepatocellular carcinoma in hepatitis C-infected patients. Mol. Cell. Proteom. 2013, 12, 3640–3652. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, Y.; Zhang, W.; Li, H.; Luo, R.; Zhou, Y.; Liao, C.L.; Huang, H.; Li, X.; Xie, Z.; et al. Screening differential expression of serum proteins in AFP-negative HBV-related hepatocellular carcinoma using iTRAQ-MALDI-MS/MS. Neoplasma 2014, 61, 17–26. [Google Scholar] [CrossRef]
- Chang, T.T.; Ho, C.H. Plasma proteome atlas for differentiating tumor stage and post-surgical prognosis of hepatocellular carcinoma and cholangiocarcinoma. PLoS ONE 2020, 15, e0238251. [Google Scholar] [CrossRef]
- Urban, S.K.; Sänger, H.; Krawczyk, M.; Julich-Haertel, H.; Willms, A.; Ligocka, J.; Azkargorta, M.; Mocan, T.; Kahlert, C.; Kruk, B.; et al. Synergistic effects of extracellular vesicle phenotyping and AFP in hepatobiliary cancer differentiation. Liver Int. 2020. [Google Scholar] [CrossRef]
- Wu, P.-Y.; Cheng, C.-W.; Kaddi, C.D.; Venugopalan, J.; Hoffman, R.; Wang, M.D. -Omic and electronic health record big data analytics for precision medicine. IEEE Transact. BioMed Engin. 2016, 64, 263–273. [Google Scholar]
- Craig, R.; Cortens, J.P.; Beavis, R.C. Open source system for analyzing, validating, and storing protein identification data. J. Proteome Res. 2004, 3, 1234–1242. [Google Scholar] [CrossRef]
- Farrah, T.; Deutsch, E.W.; Kreisberg, R.; Sun, Z.; Campbell, D.S.; Mendoza, L.; Kusebauch, U.; Brusniak, M.-Y.; Hüttenhain, R.; Schiess, R.; et al. PASSEL: The PeptideAtlas SRMexperiment library. Proteomics 2012, 12, 1170–1175. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, D.; Chen, X.; Jia, H. Computational data mining in cancer bioinformatics and cancer epidemiology. J. Biomed. Biotechnol. 2009, 2009, 582697. [Google Scholar] [CrossRef]
- Frey, L.J. Artificial intelligence and integrated genotype-phenotype identification. Genes 2018, 10, 18. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Qi, L.; Zou, Z.; Du, J.; Kong, W.; Zhao, L.; Wei, J.; Lin, L.; Ren, M.; Liu, B. Identification of a novel gene signature for the prediction of recurrence in HCC patients by machine learning of genome-wide databases. Sci. Rep. 2020, 10, 4435. [Google Scholar] [CrossRef]
- Giordano, S.; Takeda, S.; Donadon, M.; Saiki, H.; Brunelli, L.; Pastorelli, R.; Cimino, M.; Soldani, C.; Franceschini, B.; Di Tommaso, L.; et al. Rapid automated diagnosis of primary hepatic tumour by mass spectrometry and artificial intelligence. Liver Int. 2020. [Google Scholar] [CrossRef]
- LeCun, Y.; Bengio, Y.; Hinton, G. Deep learning. Nature 2015, 521, 436. [Google Scholar] [CrossRef] [PubMed]
- Bengio, Y.; Courville, A.; Vinvent, P. Representation learning: A review and new perspectives. IEEE Trans. Pattern Anal. Mach. Intell. 2013, 35, 1798–1828. [Google Scholar] [CrossRef] [PubMed]
- Munir, K.; Elahi, H.; Ayub, A.; Frezza, F.; Rizzi, A. Cancer diagnosis using deep learning: A bibliographic review. Cancers 2019, 11, 1235. [Google Scholar] [CrossRef]
- Singh, S.P.; Wang, L.; Gupta, S.; Goli, H.; Padmanabhan, P.; Gulyas, B. 3D deep learning on medical images: A review. Sensors 2020, 20, 5097. [Google Scholar] [CrossRef] [PubMed]
- Azer, S.A. Deep learning with convolutional neural networks for identification of liver masses and hepatocellular carcinoma: A systematic review. World J. Gastrointest. Oncol. 2019, 11, 1218–1230. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Jiang, H.; Pang, W. Joint multiple fully connected convolutional neural network with extreme learning machine for hepatocellular carcinoma nuclei grading. Comput. Biol. Med. 2017, 84, 156–167. [Google Scholar] [CrossRef]
- Peng, J.; Kang, S.; Ning, Z.; Deng, H.; Shen, J.; Xu, Y.; Zhang, J.; Zhao, W.; Li, X.; Gong, W.; et al. Residual convolutional neural network for predicting response of transarterial chemoembolization in hepatocellular carcinoma from CT imaging. Eur. Radiol. 2020, 30, 413–424. [Google Scholar] [CrossRef]
- Liu, D.; Liu, F.; Xie, X.; Su, L.; Liu, M.; Xie, X.; Kuang, M.; Huang, G.; Wang, Y.; Zhou, H.; et al. Accurate prediction of responses to transarterial chemoembolization for patients with hepatocellular carcinoma by using artificial intelligence in contrast-enhanced ultrasound. Eur. Radiol. 2020, 30, 2365–2376. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Kuang, S.; Cao, S.; Hu, B.; Xie, S.; Chen, S.; Chen, Y.; Gao, D.; Chen, Y.; Zhu, Y.; et al. Deep learning assisted differentiation of hepatocellular carcinoma from focal liver lesions: Choice of four-phase and three-phase CT imaging protocol. Abdom. Radiol. (N. Y.) 2020, 45, 2688–2697. [Google Scholar] [CrossRef] [PubMed]
- Mokrane, F.Z.; Lu, L.; Vavasseur, A.; Otal, P.; Peron, J.M.; Luk, L.; Yang, H.; Ammari, S.; Saenger, Y.; Rousseau, H.; et al. Radiomics machine-learning signature for diagnosis of hepatocellular carcinoma in cirrhotic patients with indeterminate liver nodules. Eur. Radiol. 2020, 30, 558–570. [Google Scholar] [CrossRef]
- Wang, W.; Chen, Q.; Iwamoto, Y.; Han, X.; Zhang, Q.; Hu, H.; Lin, L.; Chen, Y.W. Deep learning-based radiomics models early recurrence prediction of hepatocellular carcinoma with multi-phase CT images and clinical data. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2019, 2019, 4881–4884. [Google Scholar] [PubMed]
- Chaudhary, K.; Poirion, O.B.; Lu, L.; Garmire, L.X. Deep learning-based multi-omics integration robustly predicts survival in liver cancer. Clin. Cancer Res. 2018, 24, 1248–1259. [Google Scholar] [CrossRef]
- Nam, J.Y.; Lee, J.H.; Bae, J.; Chang, Y.; Cho, Y.; Sinn, D.H.; Kim, B.H.; Kim, S.H.; Yi, N.J.; Lee, K.W.; et al. Novel model to predict HCC recurrence after liver transplantation obtained using deep learning: A multicenter study. Cancers 2020, 12, E2791. [Google Scholar] [CrossRef]
- Ji, G.W.; Zhu, F.P.; Xu, Q.; Wang, K.; Wu, M.Y.; Tang, W.W.; Li, X.C.; Wang, X.H. Machine-learning analysis of contrast-enhanced CT radiomics predicts recurrence of hepatocellular carcinoma after resection: A multi-institutional study. EBioMedicine 2019, 50, 156–165. [Google Scholar] [CrossRef]
- Shi, J.Y.; Wang, X.; Ding, G.Y.; Dong, Z.; Han, J.; Guan, Z.; Ma, L.J.; Zheng, Y.; Zhang, L.; Yu, G.Z.; et al. Exploring prognostic indicators in the pathological images of hepatocellular carcinoma based on deep learning. Gut 2020, 30. [Google Scholar] [CrossRef]
- Debruyne, E.N.; Delange, J.R. Diagnosing and monitoring hepatocellular carcinoma with alpha-fetoprotein: New aspects and applications. Clin. Chim. Acta 2008, 395, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Terentiev, A.A.; Moldogazieva, N.T. Alpha-fetoprotein: A renaissance. Tumor Biol. 2013, 34, 2075–2091. [Google Scholar] [CrossRef] [PubMed]
- Bergstrand, C.G.; Czar, B. Demonstration of a new protein fraction in serum from the human fetus. Scand. J. Clin. Lab. Investig. 1956, 8, 174–176. [Google Scholar] [CrossRef]
- Abelev, G.I.; Perova, S.D.; Kharkov, N.I.; Postnikova, Z.A.; Irlin, I.S. Production of embryonic alpha-globulin by the transplantable mouse hepatomas. Transplantation 1963, 1, 174–180. [Google Scholar] [CrossRef]
- Tatarinov, Y.S. Detection of embryo-specific alpha-globulin in the blood sera of patients with primary liver tumour. Vorprosy Meditsinskoi Khimii 1964, 10, 90–91. [Google Scholar]
- Jurišić, V.; Nikulina, D. The history of alpha-fetoprotein discovery. In Alpha-Fetoprotein: Functions and Clinical Applications; Lakhi, N., Moretti, M., Eds.; Nova Science Publishers: New York, NY, USA, 2016; pp. 3–20. [Google Scholar]
- Mizejewski, G.J. Alpha-fetoprotein structure and function: Relevance to isoforms, epitopes and conformational variants. Exp. Biol. Med. 2001, 226, 377–408. [Google Scholar] [CrossRef]
- Mizejewski, G.J. Biological role of alpha-fetoprotein in cancer: Prospects for anticancer therapy. Exp. Rev. Anticancer Ther. 2002, 2, 709–735. [Google Scholar] [CrossRef]
- Mizejewski, G.J. Alpha-fetoprotein and inflammation: Is AFP an acute and/or chronic phase reactant? J. Hematol. Thromboembol. Dis. 2015, 3, 1000191. [Google Scholar]
- Moldogazieva, N.T.; Terentiev, A.A. Alpha-fetoprotein in a protein interaction network: Systems biology approach in discovery biomarkers for hepatocellular carcinoma. In Alpha-Fetoprotein: Functions and Clinical Applications; Lakhi, N., Moretti, M., Eds.; Nova Science Publishers: New-York, NY, USA, 2016; pp. 83–102. [Google Scholar]
- Moldogazieva, N.T.; Terentiev, A.A.; Shaitan, K.V. Relationship between structure and function of alpha-fetoprotein: Conformational status and biological activity. Biomed. Khim. 2005, 51, 127–151. [Google Scholar] [PubMed]
- Moldogazieva, N.T.; Terentiev, A.A. Alpha-fetoprotein and growth factors. Structure/function relationships and analogies. Usp. Biol. Chim. 2006, 46, 99–148. [Google Scholar]
- Terentiev, A.A.; Moldogazieva, N.T. Cell adhesion proteins and alpha-fetoprotein. Similar structural motifs as prerequisites for common functions. Biochemistry 2007, 72, 920–935. [Google Scholar]
- Li, M.; Li, H.; Li, C.; Zhou, S.; Guo, L.; Liu, H.; Jiang, W.; Liu, X.; Li, P.; McNutt, M.A.; et al. Alpha fetoprotein is a novel protein-binding partner for caspase-3 and blocks the apoptotic signaling pathway in human hepatoma cells. Int. J. Cancer 2009, 124, 2845–2854. [Google Scholar] [CrossRef]
- Li, M.; Li, H.; Li, C.; Wang, S.; Jiang, W.; Liu, Z.; Zhou, S.; Liu, X.; McNutt, M.A.; Li, G. Alpha-fetoprotein: A new member of intracellular signal molecules in regulation of the PI3K/AKT signaling in human hepatoma cell lines. Int. J. Cancer 2011, 128, 524–532. [Google Scholar] [CrossRef]
- Gao, R.; Cai, C.; Gan, J.; Yang, X.; Shuang, Z.; Liu, M.; Li, S.; Tang, H. Mir-1236 down-regulates alpha-fetoprotein, thus causing PTEN accumulation, which inhibits the PI3K/AKT pathway and malignant phenotype in hepatoma cells. Oncotarget 2015, 6, 6014–6028. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Guo, J.; Xia, H.; Li, W.; Lu, Y.; Dong, X.; Chen, Y.; Xie, X.; Fu, S.; Li, M. Alpha-fetoprotein activates AKT/mTOR signaling to promote CXCR4 expression and migration of hepatoma cells. Oncoscience 2015, 2, 59–70. [Google Scholar] [CrossRef]
- Sengupta, S.; Parikh, N.D. Biomarker development for hepatocellular carcinoma early detection: Current and future perspectives. Hepat. Oncol. 2017, 4, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Tyson, G.L.; Duan, Z.; Kramer, J.R.; Davila, J.A.; Richardson, P.A.; El-Serag, H.B. Level of α-fetoprotein predicts mortality among patients with hepatitis C-related hepatocellular carcinoma. Clin. Gastroenterol. Hepatol. 2011, 9, 989–994. [Google Scholar] [CrossRef]
- Peng, S.Y.; Chen, W.J.; Lai, P.L.; Jeng, Y.M.; Shen, J.C.; Hsu, H.C. High alpha-fetoprotein level correlates with high stage, early recurrence and poor prognosis of hepatocellular carcinoma: Significance of hepatitis virus infection, age, p53 and beta-catenin mutations. Int. J. Cancer 2004, 112, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tang, J.; Zhou, X.; Zhu, S.-L.; Li, L.-Q. Diagnostic accuracy of midkine for hepatocellular carcinoma: A meta-analysis. Mol. Genet. Genom. Med. 2020, 8, e1071. [Google Scholar] [CrossRef]
- Lee, E.; Edward, S.; Singal, A.G.; Lavieri, M.S.; Volk, M. Improving screening for hepatocellular carcinoma by incorporating data on levels of alpha-fetoprotein over time. Clin. Gastroenterol. Hepatol. 2013, 11, 437–440. [Google Scholar] [CrossRef]
- Yokoo, T.; Patel, A.D.; Lev-Cohain, N.; Singal, A.G.; Yopp, A.C.; Pedrosa, I. Extrahepatic metastasis risk of hepatocellular carcinoma based on α-fetoprotein and tumor staging parameters at cross-sectional imaging. Cancer Manag. Res. 2017, 9, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Bent, S.; Kohlwes, J. Test characteristics of alpha-fetoprotein for detecting hepatocellular carcinoma in patients with hepatitis C. A systematic review and critical analysis. Ann. Intern. Med. 2003, 139, 46–50. [Google Scholar] [CrossRef]
- Xu, J.; Liu, C.; Zhou, L.; Tian, F.; Tai, M.H.; Wei, J.C.; Qu, K.; Meng, F.D.; Zhang, L.Q.; Wang, Z.X.; et al. Distinctions between clinicopathological factors and prognosis of alpha-fetoprotein negative and positive hepatocellular carcinoma patients. Asian Pacific J. Cancer Prev. 2012, 13, 559–562. [Google Scholar] [CrossRef]
- Yang, J.D.; Dai, J.; Singal, A.G.; Gopal, P.; Addissie, B.D.; Nguyen, M.H.; Befeler, A.S.; Reddy, K.R.; Schwartz, M.; Harnois, D.M.; et al. Improved performance of serum alpha-fetoprotein for hepatocellular carcinoma diagnosis in HCV cirrhosis with normal alanine transaminase. Cancer Epidemiol. Biomarkers Prev. 2017, 26, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Zhang, R.; Qian, H.; Cao, L.; Zhang, Y.; Xu, W.; Li, J.; Wu, M.; Yin, Z. Serum profiling based on fucosylated glycoproteins for differentiating between chronic hepatitis B and hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2012, 240, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Wang, W.; Zhang, Y.; Liu, X.; Li, M.; Wu, Z.; Liu, Z.; Lv, Y.; Wang, B. Prognostic role of pre-treatment serum AFP-L3% in hepatocellular carcinoma: Systemic review and meta-analysis. PLoS ONE 2014, 9, e87011. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.J.; Ahmed, A.; Gish, R.G. Elevated alpha-fetoprotein: Differential diagnosis-hepatocellular carcinoma and other disorders. Clin. Liver Dis. 2015, 19, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, Y.; Tang, W.; Makuuchi, M.; Hasegawa, K.; Sugawara, Y.; Kokudo, N. Clinical and molecular insights into the hepatocellular carcinoma tumour marker des-γ-carboxyprothrombin. Liver Int. 2011, 31, 22–35. [Google Scholar] [CrossRef]
- Fujikawa, T.; Shiraha, H.; Yanamoto, K. Significance of des-gamma-carboxy prothrombin production in hepatocellular carcinoma. Acta Med. Okayama 2009, 63, 299–304. [Google Scholar]
- Matsubara, M.; Shiraha, M.; Kataoka, J.; Iwamuro, M.; Horiguchi, S.; Nishina, S.; Takaoka, N.; Uemura, M.; Takaki, A.; Nakamura, S.; et al. Des-γ-carboxyl prothrombin is associated with tumor angiogenesis in hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2012, 27, 1602–1608. [Google Scholar] [CrossRef] [PubMed]
- Yue, P.; Gao, Z.-H.; Xue, X.; Cui, S.-X.; Zhao, C.-R.; Yuan, Y.; Yin, Z.; Inagaki, Y.; Kokudo, N.; Tang, W.; et al. Des-γ-carboxyl prothrombin induces matrix metalloproteinase activity in hepatocellular carcinoma cells by involving the ERK1/2 MAPK signalling pathway. Eur. J. Cancer 2011, 47, 1115–1124. [Google Scholar] [CrossRef]
- Beale, G.; Chattopadhyay, D.; Gray, J.; Stewart, S.; Hudson, M.; Day, C.; Trerotoli, P.; Giannelli, G.; Manas, D.; Reeves, H. AFP, PIVKAII, GP3, SCCA-1 and follisatin as surveillance biomarkers for hepatocellular cancer in non-alcoholic and alcoholic fatty liver disease. BMC Cancer 2008, 8, 200. [Google Scholar] [CrossRef]
- Liu, Y.; Li, H.; Dong, J.; Ma, L.; Liao, A.; Rong, Z.; Zhou, Z.; Cao, L.; Wang, F.; Wang, J.; et al. mTOR and ERK regulate VKORC1 expression in both hepatoma cells and hepatocytes which influence blood coagulation. Clin. Exp. Med. 2019, 19, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.I.; Kim, H.S.; Kim, W.J.; Shin, W.G.; Kim, D.J.; Kim, K.H.; Jang, M.K.; Lee, J.H.; Kim, J.S.; Kim, H.Y.; et al. Diagnostic value of PIVKA-II and alpha-fetoprotein in hepatitis B virus-associated hepatocellular carcinoma. World J. Gastroenterol. 2015, 21, 3928–3935. [Google Scholar] [CrossRef] [PubMed]
- Lok, A.S.; Sterling, R.K.; Everhart, J.E.; Wright, E.C.; Hoefs, J.C.; Di Bisceglie, A.M.; Morgan, T.R.; Kim, H.-Y.; Lee, W.M.; Bonkovsky, H.L.; et al. Des-γ-carboxy prothrombin and α-fetoprotein as biomarkers for the early detection of hepatocellular carcinoma. Gastroenterology 2010, 138, 493–502. [Google Scholar] [CrossRef]
- Toyoda, H.; Kumada, T.; Kiriyama, S.; Sone, Y.; Tanikawa, M.; Hisanaga, Y.; Yamaguchi, A.; Isogai, M.; Kaneoka, Y.; Washizu, J. Prognostic significance of simultaneous measurement of three tumor markers in patients with hepatocellular carcinoma. Clin. Gastroenterol. Hepatol. 2006, 4, 111–117. [Google Scholar] [CrossRef]
- Park, S.J.; Jang, J.Y.; Jeong, S.W.; Cho, Y.K.; Lee, S.H.; Kim, S.G.; Cha, S.-W.; Kim, Y.S.; Cho, Y.D.; Kim, H.S.; et al. Usefulness of AFP, AFP-L3, and PIVKA-II, and their combinations in diagnosing hepatocellular carcinoma. Medicine 2017, 96, e5811. [Google Scholar] [CrossRef]
- Lim, T.S.; Kim, D.Y.; Han, K.H.; Kim, H.S.; Shin, S.H.; Jung, K.S.; Kim, B.K.; Kim, S.U.; Park, J.Y.; Ahn, S.H. Combined use of AFP, PIVKA-II, and AFP-L3 as tumor markers enhances diagnostic accuracy for hepatocellular carcinoma in cirrhotic patients. Scand. J. Gastroenterol. 2016, 51, 344–353. [Google Scholar] [CrossRef]
- Takeji, S.; Hirooka, M.; Koizumi, Y.; Tokumoto, Y.; Abe, M.; Ikeda, Y.; Nadano, S.; Hiasa, Y.; Onji, M. Des-gamma-carboxy prothrombin identified by P-11 and P-16 antibodies reflects prognosis for patients with hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2013, 28, 671–677. [Google Scholar] [CrossRef]
- Sumi, A.; Akiba, J.; Ogasawara, S.; Nakayama, M.; Nomura, Y.; Yasumoto, M.; Sanada, S.; Nakashima, O.; Abe, T.; Yano, H. Des-γ-carboxyprothrombin (DCP) and NX-DCP expressions and their relationship with clinicopathological features in hepatocellular carcinoma. PLoS ONE 2015, 10, e0118452. [Google Scholar] [CrossRef] [PubMed]
- Nakatsura, T.; Nishimura, Y. Usefulness of the novel oncofetal antigen for diagnosis of hepatocellular carcinoma and melanoma. BioDrug 2005, 19, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Y.; Wang, S.Y.; Lin, Y.; Yi, H.C.; Niu, J.J. The diagnostic performance of lncRNAs from blood specimens in patients with hepatocellular carcinoma: A meta-analysis. Lab. Med. 2021, 52, 64–73. [Google Scholar] [CrossRef]
- Gan, Y.; Liang, Q.; Song, X. Diagnostic value of alpha-L-fucosidase for hepatocellular carcinoma: A meta-analysis. Tumor Biol. 2014, 35, 3953–3960. [Google Scholar] [CrossRef]
- Chen, I.P.; Ariizumi, S.; Nakano, M.; Yamamoto, M. Positive glypican-3 expression in early hepatocellular carcinoma predicts recurrence after hepatectomy. J. Gastroenterol. 2014, 49, 117–125. [Google Scholar] [CrossRef]
- Schizas, D.; Mastoraki, A.; Routsi, E.; Papapanou, M.; Tsapralis, D.; Vassiliu, P.; Toutouzas, K.; Felekouras, E. Combined hepatocellular-cholangiocarcinoma: An update on epidemiology, classification, diagnosis and management. Hepatobiliary Pancreat Dis Int. 2020, 19, 515–523. [Google Scholar] [CrossRef]
- Mao, Y.; Yang, H.; Xu, H.; Lu, X.; Sang, X.; Du, S.; Zhao, H.; Chen, W.; Xu, Y.; Chi, T.; et al. Golgi protein 73 (GOLPH2) is a valuable serum marker for hepatocellular carcinoma. Gut 2010, 59, 1687–1693. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Wang, J.; Katayama, H.; Sen, S.; Liu, S.-M. Circulating microRNAs (cmiRNAs) as novel potential biomarkers for hepatocellular carcinoma. Neoplasma 2013, 60, 135. [Google Scholar] [CrossRef]
- Ye, Q.; Ling, S.; Zheng, S.; Xu, X. Liquid biopsy in hepatocellular carcinoma: Circulating tumor cells and circulating tumor DNA. Mol. Cancer 2019, 18, 114. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, N.; Sawada, Y.; Endo, I.; Saito, K.; Uemura, Y.; Nakatsura, T. Biomarkers for the early diagnosis of hepatocellular carcinoma. World J. Gastroenterol. 2015, 21, 10573–10583. [Google Scholar] [CrossRef] [PubMed]
- Shang, S.; Plymoth, A.; Feng, Z.; Rosen, H.R.; Sangrajrang, S.; Hainaut, P.; Marrero, J.A.; Beretta, L. Identification of osteopontin as a novel marker for early hepatocellular carcinoma. Hepatology 2012, 55, 483–490. [Google Scholar] [CrossRef]
- Qin, L. Osteopontin is a promoter for hepatocellular carcinoma metastasis: A summary of 10 years of studies. Front. Med. 2014, 8, 24–32. [Google Scholar] [CrossRef]
- Zhu, M.; Zheng, J.; Wu, F.; Kang, B.; Liang, J.; Heskia, F.; Zhang, X.; Shan, Y. OPN is a promising serological biomarker for hepatocellular carcinoma diagnosis. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Denhardt, D.T.; Guo, X. Osteopontin: A protein with diverse functions. FASEB J. 1993, 7, 1475–1482. [Google Scholar] [CrossRef]
- El-Tanani, M.K. Role of osteopontin in cellular signaling and metastatic phenotype. Front. Biosci. 2008, 13, 4276–4284. [Google Scholar] [CrossRef]
- Anborgh, P.H.; Mutrie, J.C.; Tuck, A.B.; Chambers, A.F. Role of the metastasis-promoting protein osteopontin in the tumour microenvironment. J. Cell. Mol. Med. 2010, 14, 2037–2044. [Google Scholar] [CrossRef] [PubMed]
- Rangaswami, H.; Bulbule, A.; Kundu, G.C. Osteopontin: Role in cell signaling and cancer progression. Trends Cell Biol. 2006, 16, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Fan, X.; Tang, M.; Chen, R.; Wang, H.; Jia, R.; Zhou, X.; Jing, W.; Wang, H.; Yang, Y.; et al. Osteopontin induces autophagy to promote chemo-resistance in human hepatocellular carcinoma cells. Cancer Lett. 2016, 383, 171–182. [Google Scholar] [CrossRef]
- Cao, L.; Fan, X.; Jing, W.; Liang, Y.; Chen, R.; Liu, Y.; Zhu, M.; Jia, R.; Wang, H.; Zhang, X.; et al. Osteopontin promotes a cancer stem cell-like phenotype in hepatocellular carcinoma cells via integrin-κB-HIF-1α pathway. Oncotarget 2015, 6, 6627–6640. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Fang, S.; Weng, Q.; Lv, X.; Meng, M.; Zhu, J.; Zheng, L.; Hu, Y.; Gao, Y.; Wu, X.; et al. Integrated analysis reveals critical regulators in hepatocellular carcinoma. Cell Commun. Signal. 2020, 18, 97. [Google Scholar] [CrossRef]
- Yu, X.; Zheng, Y.; Zhu, X.; Gao, X.; Wang, C.; Sheng, Y.; Cheng, W.; Qin, L.; Ren, N.; Jia, H.; et al. Osteopontin promotes hepatocellular carcinoma progression via the PI3K/AKT/Twist signaling pathway. Oncol. Lett. 2018, 16, 5299–5308. [Google Scholar] [CrossRef]
- Zhu, Y.; Gao, X.M.; Yang, J.; Xu, D.; Zhang, Z.; Sheng, Y.Y.; Li, J.H.; Yu, X.X.; Zheng, Y.; Dong, Q.Z.; et al. C-C chemokine receptor type 1 mediates osteopontin-promoted metastasis in hepatocellular carcinoma. Cancer Sci. 2018, 109, 710–723. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, J.; Xu, D.; Gao, X.-M.; Zhang, Z.; Hsu, J.L.; Li, C.-W.; Lim, S.-O.; Sheng, Y.-Y.; Zhang, Y.; et al. Disruption of tumour-associated macrophage trafficking by the osteopontin-induced colony-stimulating factor-1 signalling sensitises hepatocellular carcinoma to anti-PD-L1 blockade. Gut 2019, 68, 1653–1666. [Google Scholar] [CrossRef]
- Wan, H.G.; Xu, H.; Gu, Y.M.; Wang, H.; Xu, W.; Zu, M.H. Comparison osteopontin vs AFP for the diagnosis of HCC: A meta-analysis. Clin. Res. Hepatol. Gastroenterol. 2014, 38, 706–714. [Google Scholar] [CrossRef]
- Fouad, S.A.; Mohamed, N.A.G.; Fawzy, M.W.; Moustafa, D.A. Plasma osteopontin level in chronic liver disease and hepatocellular carcinoma. Hepat. Mon. 2015, 15, e30753. [Google Scholar] [CrossRef]
- Li, J.; Chen, X.; Dai, M.; Huang, S.; Chen, J.; Dai, S. Diagnostic accuracy of osteopontin plus alpha-fetoprotein in the hepatocellular carcinoma: A meta-analysis. Clin. Res. Hepatol. Gastroenterol. 2017, 41, 543–553. [Google Scholar] [CrossRef]
- Sun, T.; Tang, Y.; Sun, D.; Bu, Q.; Li, P. Osteopontin versus alpha-fetoprotein as a diagnostic marker for hepatocellular carcinoma: A meta-analysis. Onco Targets Ther. 2018, 11, 8925–8935. [Google Scholar] [CrossRef]
- Ronald, J.; Nixon, A.B.; Marin, D.; Gupta, R.T.; Janas, G.; Chen, W.; Suhocki, P.V.; Pabon-Ramos, W.; Sopko, D.R.; Starr, M.D.; et al. Pilot evaluation of angiogenesis signaling factor response after transcatheter arterial embolization for hepatocellular carcinoma. Radiology 2017, 285, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Rong, W.; Xiao, T.; Zhang, Y.; Xu, B.; Liu, Y.; Wang, L.; Wu, F.; Qi, J.; Zhao, X.; et al. Secretory/releasing proteome-based identification of plasma biomarkers in HBV-associated hepatocellular carcinoma. Sci. China Life Sci. 2013, 56, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, A.N.; Plymoth, A.; Santos-Silva, D.; Ortiz-Cuaran, S.; Camey, S.; Guilloreau, P.; Sangrajrang, S.; Khuhaprema, T.; Mendy, M.; Lesi, O.A.; et al. Osteopontin and latent-TGF β binding-protein 2 as potential diagnostic markers for HBV-related hepatocellular carcinoma. Int. J. Cancer 2015, 136, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Yang, L.; Rong, W.; Feng, L.; Han, N.; Zhang, K.; Cheng, S.; Wu, J.; Xiao, T.; Gao, Y. Latent transforming growth factor-beta binding protein-1 in circulating plasma as a novel biomarker for early detection of hepatocellular carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 16046–16054. [Google Scholar]
- Capurro, M.; Wanless, I.R.; Sherman, M.; Deboer, G.; Shi, W.; Miyoshi, E.; Filmus, J. Glypican-3: A novel serum and histochemical marker for hepatocellular carcinoma. Gastroenterology 2003, 125, 89–97. [Google Scholar] [CrossRef]
- Morford, L.A.; Davis, C.; Jin, L.; Dobierzewska, A.; Peterson, M.L.; Spear, B.T. The oncofetal gene glypican-3 is regulated in the postnatal liver by zinc fingers and homeoboxes 2 and in the regenerating liver by alpha-fetoprotein regulator 2. Hepatology 2007, 46, 1541–1547. [Google Scholar] [CrossRef] [PubMed]
- Capurro, M.I.; Xiang, Y.Y.; Lobe, C.; Filmus, J. Glypican-3 promotes the growth of hepatocellular carcinoma by stimulating canonical Wnt signaling. Cancer Res. 2005, 65, 6245–6254. [Google Scholar] [CrossRef]
- Jia, X.; Gao, Y.; Zhai, D.; Liu, J.; Cai, J.; Wang, Y.; Jing, L.; Du, Z. Assessment of the clinical utility of glypican 3 as a serum marker for the diagnosis of hepatocellular carcinoma. Technol. Cancer Res. Treat. 2016, 15, 780–786. [Google Scholar] [CrossRef]
- Jeon, Y.; Jang, E.S.; Choi, Y.S.; Kim, J.-W.; Jeong, S.-H. Glypican-3 level assessed by the enzyme-linked immunosorbent assay is inferior to alpha-fetoprotein level for hepatocellular carcinoma diagnosis. Clin. Mol. Hepatol. 2016, 22, 359–365. [Google Scholar] [CrossRef]
- Gu, D.; Xie, Y.; Wei, J.; Li, W.; Ye, Z.; Zhu, Z.; Tian, J.; Li, X. MRI-based radiomics signature: A potential biomarker for identifying glypican 3-positive hepatocellular carcinoma. J. Magn. Reson. Imaging 2020, 52, 1679–1687. [Google Scholar] [CrossRef]
- Wang, L.; Yao, M.; Pan, L.H.; Qian, Q.; Yao, D.F. Glypican-3 is a biomarker and a therapeutic target of hepatocellular carcinoma. Hepatobiliary Pancreat. Dis. Int. 2015, 14, 361–366. [Google Scholar] [CrossRef]
- Ghousein, A.; Mosca, N.; Cartier, F.; Charpentier, J.; Dupuy, J.-W.; Raymond, A.-A.; Bioulac-Sage, P.; Grosset, C.F. miR-4510 blocks hepatocellular carcinoma development through RAF1 targeting and RAS/RAF/MEK/ERK signalling inactivation. Liver Int. 2020, 40, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Ning, X.; Deng, Z.; Liu, M.; Zhou, B.; Chen, X.; Huang, S.; Xu, Y.; Chen, Z.; Luo, R. Propofol-induced miR-219-5p inhibits growth and invasion of hepatocellular carcinoma through suppression of GPC3-mediated Wnt/β-catenin signalling activation. J. Cell Biochem. 2019, 120, 16934–16945. [Google Scholar] [CrossRef]
- Gao, J.; Dai, C.; Yu, X.; Yin, X.B.; Zhou, F. microRNA-485-5p inhibits the progression of hepatocellular carcinoma through blocking the WBP2/Wnt signaling pathway. Cell. Signal. 2020, 66, 09466. [Google Scholar] [CrossRef]
- Liu, X.; Gao, F.; Jiang, L.; Jia, M.; Ao, L.; Lu, M.; Gou, L.; Ho, M.; Jia, S.; Chen, F.; et al. 32A9, a novel human antibody for designing an immunotoxin and CAR-T cells against glypican-3 in hepatocellular carcinoma. J. Transl. Med. 2020, 18, 295. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Shi, Y.; Kaseb, A.O.; Qi, X.; Zhang, Y.; Chi, J.; Lu, Q.; Gao, H.; Jiang, H.; Wang, H.; et al. Chimeric antigen receptor-glypican-3 T-cell therapy for advanced hepatocellular carcinoma: Results of phase I trials. Clin. Cancer Res. 2020, 26, 3979–3989. [Google Scholar] [CrossRef]
- Koide, N.; Hada, H.; Shinji, T.; Ujike, K.; Hirasaki, S.; Yumoto, Y.; Hanafusa, T.; Kadomatsu, K.; Muramatsu, H.; Muramatsu, T.; et al. Expression of the midkine gene in human hepatocellular carcinomas. Hepatogastroenterology 1999, 46, 3189–3196. [Google Scholar] [PubMed]
- Sun, B.; Hu, C.; Yang, Z.; Zhang, X.; Zhao, L.; Xiong, J.; Ma, J.; Chen, L.; Qian, H.; Luo, X.; et al. Midkine promotes hepatocellular carcinoma metastasis by elevating anoikis resistance of circulating tumor cells. Oncotarget 2017, 8, 32523–32535. [Google Scholar] [CrossRef]
- Duan, R.; Li, C.; Wang, F.; Han, F.; Zhu, L. The long noncoding RNA ZFAS1 potentiates the development of hepatocellular carcinoma via the microRNA-624/MDK/ERK/JNK/P38 signaling pathway. Onco Targets Ther. 2020, 13, 4431–4444. [Google Scholar] [CrossRef]
- Zhu, W.-W.; Guo, J.-J.; Guo, L.; Jia, H.L.; Zhu, M.; Zhang, J.B.; Loffredo, C.A.; Forgues, M.; Huang, H.; Xing, X.J.; et al. Evaluation of midkine as a diagnostic serum biomarker in hepatocellular carcinoma. Clin. Cancer Res. 2013, 19, 3944–3954. [Google Scholar] [CrossRef]
- Hodeib, H.; Elshora, O.; Selim, A.; Sabry, N.M.; El-Ashry, H.M. Serum midkine and osteopontin levels as diagnostic biomarkers of hepatocellular carcinoma. Electron. Physician 2017, 9, 3492–3498. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, K.Y.A.; Abdel-Mageed, A.I.; Safwat, E.; Albreedy, A.M. The value of serum midkine level in diagnosis of hepatocellular carcinoma. Int. J. Hepatol. 2015, 2015, 146389. [Google Scholar] [CrossRef] [PubMed]
- Vongsuvanh, R.; van Der Poorten, D.; Iseli, T.; Strasser, S.I.; McCaughan, G.W.; George, J. Midkine increases diagnostic yield in AFP negative and NASH-related hepatocellular carcinoma. PLoS ONE 2016, 11, e0155800. [Google Scholar] [CrossRef]
- Lu, Q.; Li, J.; Cao, H.; Lv, C.; Wang, X.; Cao, S. Comparison of diagnostic accuracy of midkine and AFP for detecting hepatocellular carcinoma: A systematic review and meta-analysis. BioSci. Rep. 2020, 40, BSR20192424. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, G.; Zhang, P.; Zhang, J.; Li, X.; Gan, D.; Cao, X.; Han, M.; Du, H.; Ye, Y. The threshold of alpha-fetoprotein (AFP) for the diagnosis of hepatocellular carcinoma: A systematic review and meta-analysis. PLoS ONE 2020, 15, e0228857. [Google Scholar] [CrossRef]
- Glinka, A.; Wu, W.; Delius, H.; Monaghan, A.P.; Blumenstock, C.; Niehrs, C. Dickkopf-1 is a member of a new family of secreted proteins and functions in head induction. Nature 1998, 391, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.W.; Park, J.S.; Han, J.H.; Kim, J.-H.; Ahn, M.S.; Lee, H.W.; Kang, S.Y.; Choi, J.-H.; Jeong, S.H. Strong immunoexpression of dickkopf-1 is associated with response to bortezomib in multiple myeloma. Leuk. Lymphoma 2018, 59, 2670–2678. [Google Scholar] [CrossRef] [PubMed]
- Pamuk, G.E.; Uyanik, M.S.; Pamuk, O.N.; Maden, M.; Tapan, U. Decreased dickkopf-1 levels in chronic lymphocytic leukemia and increased osteopontin levels in non-Hodgkin’s lymphoma at initial diagnosis: Could they be playing roles in pathogenesis? Hematology 2015, 20, 267–271. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, B.; Qi, L.; Li, Y.; Zhao, X.; Zhang, D.; Zhang, Y. Dickkopf-1 expression is down-regulated during the colorectal adenoma-carcinoma sequence and correlates with reduced microvessel density and VEGF expression. Histopathology 2015, 67, 158–166. [Google Scholar] [CrossRef]
- Liang, L.; He, H.; Lv, R.; Zhang, M.; Huang, H.; An, Z.; Li, S. Preliminary mechanism on the methylation modification of Dkk-1 and Dkk-3 in hepatocellular carcinoma. Tumor Biol. 2014, 36, 1245–1250. [Google Scholar] [CrossRef]
- Sakabe, T.; Azumi, J.; Umekita, Y.; Toriguchi, K.; Hatano, E.; Hirooka, Y.; Shiota, G. Expression of cancer stem cell-associated DKK1 mRNA serves as prognostic marker for hepatocellular carcinoma. Anticancer Res. 2017, 37, 4881–4888. [Google Scholar]
- Kagey, M.H.; He, X. Rationale for targeting the Wnt signalling modulator Dickkopf-1 for oncology. Br. J. Pharmacol. 2017, 174, 4637–4650. [Google Scholar] [CrossRef] [PubMed]
- Fezza, M.; Moussa, M.; Aoun, R.; Haber, R.; Hilal, G. DKK1 promotes hepatocellular carcinoma inflammation, migration and invasion: Implication of TGF-β1. PLoS ONE 2019, 14, e0223252. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Fan, J.; Yang, X.R.; Tan, Y.; Zhao, W.; Xu, Y.; Wang, N.; Niu, Y.; Wu, Z.; Zhou, J.; et al. Serum DKK1 as a protein biomarker for the diagnosis of hepatocellular carcinoma: A large-scale, multicentre study. Lancet Oncol. 2012, 13, 817–826. [Google Scholar] [CrossRef]
- Jang, E.S.; Jeong, S.-H.; Kim, J.-W.; Choi, Y.S.; Leissner, P.; Brechot, C. Diagnostic performance of alpha-fetoprotein, protein Induced by vitamin K absence, osteopontin, dickkopf-1 and its combinations for hepatocellular carcinoma. PLoS ONE 2016, 11, e0151069. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Mou, L.; Gao, H.; Zeng, Y.; Tang, X.; Deng, X.; Pu, Z.; Ni, Y.; Zhan, Y. Diagnostic accuracy of serum dickkopf-1 protein in diagnosis hepatocellular carcinoma: An updated meta-analysis. Medicine 2019, 98, e16725. [Google Scholar] [CrossRef]
- Ge, T.; Shen, Q.; Wang, N.; Zhang, Y.; Ge, Z.; Chu, W.; Lv, X.; Zhao, F.; Zhao, W.; Fan, J.; et al. Diagnostic values of alpha-fetoprotein, dickkopf-1, and osteopontin for hepatocellular carcinoma. Med. Oncol. 2015, 32, 59. [Google Scholar] [CrossRef] [PubMed]
- Byeon, H.; Lee, S.D.; Hong, E.-K.; Lee, D.E.; Kim, B.H.; Seo, Y.; Joo, J.; Han, S.-S.; Kim, S.H.; Park, S.-J. Long-term prognostic impact of osteopontin and Dickkopf-related protein 1 in patients with hepatocellular carcinoma after hepatectomy. Pathol. Res. Pract. 2018, 214, 814–820. [Google Scholar] [CrossRef]
- Suminami, Y.; Kishi, F.; Sekiguchi, K.; Kato, H. Squamous cell carcinoma antigen is a new member of the serine protease inhibitors. Biochem. Biophys. Res. Commun. 1991, 181, 51–58. [Google Scholar] [CrossRef]
- Suminami, Y.; Kishi, F.; Murakami, A.; Sakaguchi, Y.; Nawata, S.; Numa, F.; Kato, H. Novel forms of squamous cell carcinoma antigen transcripts produced by alternative splicing. Biochim. Biophys. Acta 2001, 1519, 122–126. [Google Scholar] [CrossRef]
- Cataltepe, S.; Gornstein, E.R.; Schick, C.; Kamachi, Y.; Chatson, K.; Fries, J.; Silverman, G.A.; Upton, M.P. Co-expression of the squamous cell carcinoma antigens 1 and 2 in normal adult human tissues and squamous cell carcinomas. J. Histochem. Cytochem. 2000, 48, 113–122. [Google Scholar] [CrossRef]
- Turato, C.; Vitale, A.; Fasolato, S.; Ruvoletto, M.; Terrin, L.; Quarta, S.; Ramirez Morales, R.; Biasiolo, A.; Zanus, G.; Tan, P.S.; et al. SERPINB3 is associated with TGF-β1 and cytoplasmic β-catenin expression in hepatocellular carcinomas with poor prognosis. Br. J. Cancer 2014, 110, 2708–2715. [Google Scholar] [CrossRef] [PubMed]
- Pontisso, P. Role of SERPINB3 in hepatocellular carcinoma. Ann. Hepatol. 2014, 13, 722–727. [Google Scholar] [CrossRef]
- Lachenmayer, A.; Alsinet, C.; Savic, R.; Cabellos, L.; Toffanin, S.; Hoshida, Y.; Vilanueva, A.; Minquez, B.; Newell, P.; Tsai, H.W.; et al. Wnt-pathway activation in two molecular classes of hepatocellular carcinoma and experimental modulation by sorafenib. Clin. Cancer Res. 2012, 18, 4997–5007. [Google Scholar] [CrossRef]
- Hernanda, P.Y.; Chen, K.; Das, A.M.; Sideras, K.; Wang, W.; Li, J.; Cao, W.; Bots, S.J.; Kodach, L.L.; de Man, R.A.; et al. SMAD4 exerts a tumor-promoting role in hepatocellular carcinoma. Oncogene 2015, 34, 5055–5068. [Google Scholar] [CrossRef] [PubMed]
- Cannito, S.; Turato, C.; Paternostro, C.; Biasiolo, A.; Colombatto, S.; Cambieri, I.; Quarta, S.; Novo, E.; Morello, E.; Villano, G.; et al. Hypoxia up-regulates SERPINB3 through HIF-2α in human liver cancer cells. Oncotarget 2015, 6, 2206–2221. [Google Scholar] [CrossRef]
- Cannito, S.; Foglia, B.; Villano, G.; Turato, C.; Delgado, T.C.; Morello, E.; Pin, F.; Novo, E.; Napione, L.; Quarta, S.; et al. SerpinB3 differently up-regulates hypoxia inducible factors-1α and -2α in hepatocellular carcinoma: Mechanisms revealing novel potential therapeutic targets. Cancers 2019, 11, 1933. [Google Scholar] [CrossRef] [PubMed]
- Quarta, S.; Vidalino, L.; Turato, C.; Ruvoletto, M.; Calabrese, F.; Valente, M.; Cannito, S.; Fassina, G.; Parola, M.; Gatta, A.; et al. SERPINB3 induces epithelial-mesenchymal transition. J. Pathol. 2010, 221, 343–536. [Google Scholar] [CrossRef]
- Beneduce, L.; Castaldi, F.; Marino, M.; Quarta, S.; Ruvoletto, M.; Benvegnu, L.; Calabrese, F.; Gatta, A.; Pontisso, P.; Fassina, G. Squamous cell carcinoma antigen-immunoglobulin M complexes as novel biomarkers for hepatocellular carcinoma. Cancer 2005, 103, 2558–2565. [Google Scholar] [CrossRef] [PubMed]
- Mossad, N.A.; Mahmoud, E.H.; Osman, E.A.; Mahmoud, S.H.; Shousha, H.I. Evaluation of squamous cell carcinoma antigen-immunoglobulin M complex (SCCA-IGM) and alpha-L-fucosidase (AFU) as novel diagnostic biomarkers for hepatocellular carcinoma. Tumor Biol. 2014, 35, 11559–11564. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shao, C.; Zhou, Q.; Zhu, Y.; Zhu, J.; Tu, C. Diagnostic accuracy of serum squamous cell carcinoma antigen and squamous cell carcinoma antigen-immunoglobulin M for hepatocellular carcinoma: A meta-analysis. Mol. Clin. Oncol. 2015, 3, 1165–1171. [Google Scholar] [CrossRef]
- Yu, J.; Wang, Z.-J.; Chen, L.-H.; Dong, W.-Z. Diagnostic value of serum squamous cell carcinoma antigen for hepatocellular carcinoma: A systematic review and meta-analysis. Scand. J. Clin. Lab. Investig. 2017, 77, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-H.; Gil-Gomez, A.; Ampuero, J.; Romero-Gomez, M. Diagnostic accuracy of SCCA and SCCA-IgM for hepatocellular carcinoma: A meta-analysis. Liver Int. 2018, 38, 1820–1831. [Google Scholar] [CrossRef]
- Bertino, G.; Neri, S.; Bruno, C.M.; Ardiri, A.M.; Calvagno, G.S.; Malaguarnera, M.; Toro, A.; Malaguarnera, M.; Clementi, S.; Bertino, N.; et al. Diagnostic and prognostic value of alpha-fetoprotein, des-γ-carboxyprothrombin and squamous cell carcinoma antigen immunoglobulin M complexes in hepatocellular carcinoma. Minerva Med. 2011, 102, 363–371. [Google Scholar]
- Yeo, I.; Kim, G.A.; Kim, H.; Lee, J.H.; Sohn, A.; Gwak, G.Y.; Lee, J.H.; Lim, Y.S.; Kim, Y. Proteome multimarker panel with multiple reaction monitoring-mass spectrometry for early detection of hepatocellular carcinoma. Hepatol. Commun. 2020, 4, 753–768. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Liu, X.; Wei, X.; Luo, H.; Li, P.; Shi, M.; Guo, B.; Cui, Y.; Su, Z.; Zeng, J.; et al. Quantitative proteomics identifies a plasma multi-protein model for detection of hepatocellular carcinoma. Sci. Rep. 2020, 10, 15552. [Google Scholar] [CrossRef]
- Jia, C.; Wang, G.; Wang, T.; Fu, B.; Zhang, Y.; Huang, L.; Deng, Y.; Chen, G.; Wu, X.; Chen, J.; et al. Cancer-associated fibroblasts induce epithelial-mesenchymal transition via the transglutaminase 2-dependent IL-6/IL6R/STAT3 axis in hepatocellular carcinoma. Int. J. Biol. Sci. 2020, 16, 2542–2558. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.H.; Yang, D.L. Identification of a protein signature for predicting overall survival of hepatocellular carcinoma: A study based on data mining. BMC Cancer 2020, 20, 720. [Google Scholar] [CrossRef] [PubMed]
- Hodge, K.; Makjaroen, J.; Robinson, J.; Khoomrung, S.; Pisitkun, T. Deep proteomic deconvolution of interferons and HBV transfection effects on a hepatoblastoma cell line. ACS Omega 2020, 5, 16796–16810. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zheng, X.; Yuan, H.; Chen, G.; Ouyang, J.; Liu, J.; Liu, X.; Xing, X.; Zhao, B. Proteomic analyses reveal divergent ubiquitylation patterns in hepatocellular carcinoma cell lines with different metastasis potential. J. Proteomics 2020, 225, 103834. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.; Kim, S.; Kim, K.; Yeom, J.; Park, S.; Kim, I. Euchromatin histone methyltransferase II (EHMT2) regulates the expression of Ras-related GTP binding C (RRAGG) protein. BMB Rep. 2020, 20, 4978. [Google Scholar]
- Wang, Y.; Tian, Y.; Liu, X.; Dong, J.; Wang, L.; Ye, M. A new workflow for the analysis of phosphosite occupancy in paired samples by integration of proteomics and phosphoproteomics data sets. J. Proteome Res. 2020, 19, 3807–3816. [Google Scholar] [CrossRef]
- Jen, H.W.; Gu, D.L.; Lang, Y.D.; Jou, Y.S. PSPC1 potentiates IGF1R expression to augment cell adhesion and motility. Cells 2020, 9, 1490. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Jia, L.; Li, J.; Ma, C.; Wu, J.; Shen, J.; Dang, L.; Zhu, B.; Li, P.; Zhi, Y.; et al. Heterogeneities of site-specific N-glycosylation in HCC tumors with low and high AFP concentrations. Front. Oncol. 2020, 10, 496. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Huang, J.; Zhang, J.; Chen, Z.; Lin, Y.; Grigorean, G.; Li, L.; Liu, S.; Singal, A.G.; Parikh, N.D.; et al. Glycopeptide biomarkers in serum haptoglobin for hepatocellular carcinoma. J. Proteome Res. 2020, 19, 3452–3466. [Google Scholar] [CrossRef]
- Li, H.; Rong, Z.; Wang, H.; Zhang, N.; Pu, C.; Zhao, Y.; Zheng, X.; Lei, C.; Liu, Y.; Luo, X.; et al. Proteomic analysis revealed common, unique and systemic signatures in gender-dependent hepatocarcinogenesis. Biol. Sex Dif. 2020, 11, 46. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomarker | Chemical Nature | Functions | Signaling Pathways | References |
|---|---|---|---|---|
| AFP and AFL-L3 | Embryo-specific and tumor-associated glycoprotein | Dual regulation of cell proliferation and survival | MAPK- an PI3K/Akt/mTOR signaling | [69,70,71,72,73] |
| DCP | Abnormal prothrombin without carboxylation of γ-carbon atom in Glu residues in γ-carboxyglutamic (Gla) domain | Growth factor activity and DNA synthesis | JAK/STAT3, Raf/MEK1/2/ERK1/2 (MAPK) signaling | [89,90,91] |
| OPN | Acidic chemokine-like secreted ECM-specific phosphoglycoprotein | Cell adhesion, migration, invasion, and survival, epithelial-to-mesenchymal transition | Integrin αvβ3/NF-κB/HIF-1α and PI3K/Akt/NF-κB and CD44-mediated signaling | [117,118,119,120,121] |
| GPC3 | Heparan sulfate proteoglycan | Cell proliferation and tumor growth | Co-receptor of canonical Wnt/β-catenin signaling; Ras/Raf/MEK/ERK signaling | [138,139,140] |
| MDK | Small heparin-binding growth factor | HCC progression and metastasis, resistance of CTCs to anoikis | PI3K/Akt/NF-kB/TrkB and ERK/JNK/p38-mediated signaling | [143,144,145] |
| DKK1 | Secreted glycoprotein | TME remodeling, promotion of inflammation, cell migration and invasion | TGF-β1-mediated pathway | [158,159] |
| SCCA and SCCA-IgM | Member of serine protease inhibitor (serpin) family | Inhibition of apoptosis and intra-tumor infiltration by NK cells; induction of epithelial-to-mesenchymal transition, cell proliferation and invasion | c-Myc and Ras/TGF-β/SMAD4 signaling | [169,170,171] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moldogazieva, N.T.; Mokhosoev, I.M.; Zavadskiy, S.P.; Terentiev, A.A. Proteomic Profiling and Artificial Intelligence for Hepatocellular Carcinoma Translational Medicine. Biomedicines 2021, 9, 159. https://doi.org/10.3390/biomedicines9020159
Moldogazieva NT, Mokhosoev IM, Zavadskiy SP, Terentiev AA. Proteomic Profiling and Artificial Intelligence for Hepatocellular Carcinoma Translational Medicine. Biomedicines. 2021; 9(2):159. https://doi.org/10.3390/biomedicines9020159
Chicago/Turabian StyleMoldogazieva, Nurbubu T., Innokenty M. Mokhosoev, Sergey P. Zavadskiy, and Alexander A. Terentiev. 2021. "Proteomic Profiling and Artificial Intelligence for Hepatocellular Carcinoma Translational Medicine" Biomedicines 9, no. 2: 159. https://doi.org/10.3390/biomedicines9020159
APA StyleMoldogazieva, N. T., Mokhosoev, I. M., Zavadskiy, S. P., & Terentiev, A. A. (2021). Proteomic Profiling and Artificial Intelligence for Hepatocellular Carcinoma Translational Medicine. Biomedicines, 9(2), 159. https://doi.org/10.3390/biomedicines9020159