
Epigenetic Dysregulation of Trophoblastic Gene Expression in Gestational Trophoblastic Disease

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Samples
2.2. Histopathologic Evaluation of Tissues
2.3. Tissue Microarray (TMA) Construction, Immunohistochemistry, Immunoscoring
2.4. High-Dimensional Molecular Data
2.5. Methylation Data Analysis
2.6. Analysis of Imprinted Sites
2.7. mRNA Microarray Analysis
2.8. RNAseq Analysis
2.9. Comparison of Differential DNA Methylation and Differential mRNA Expression
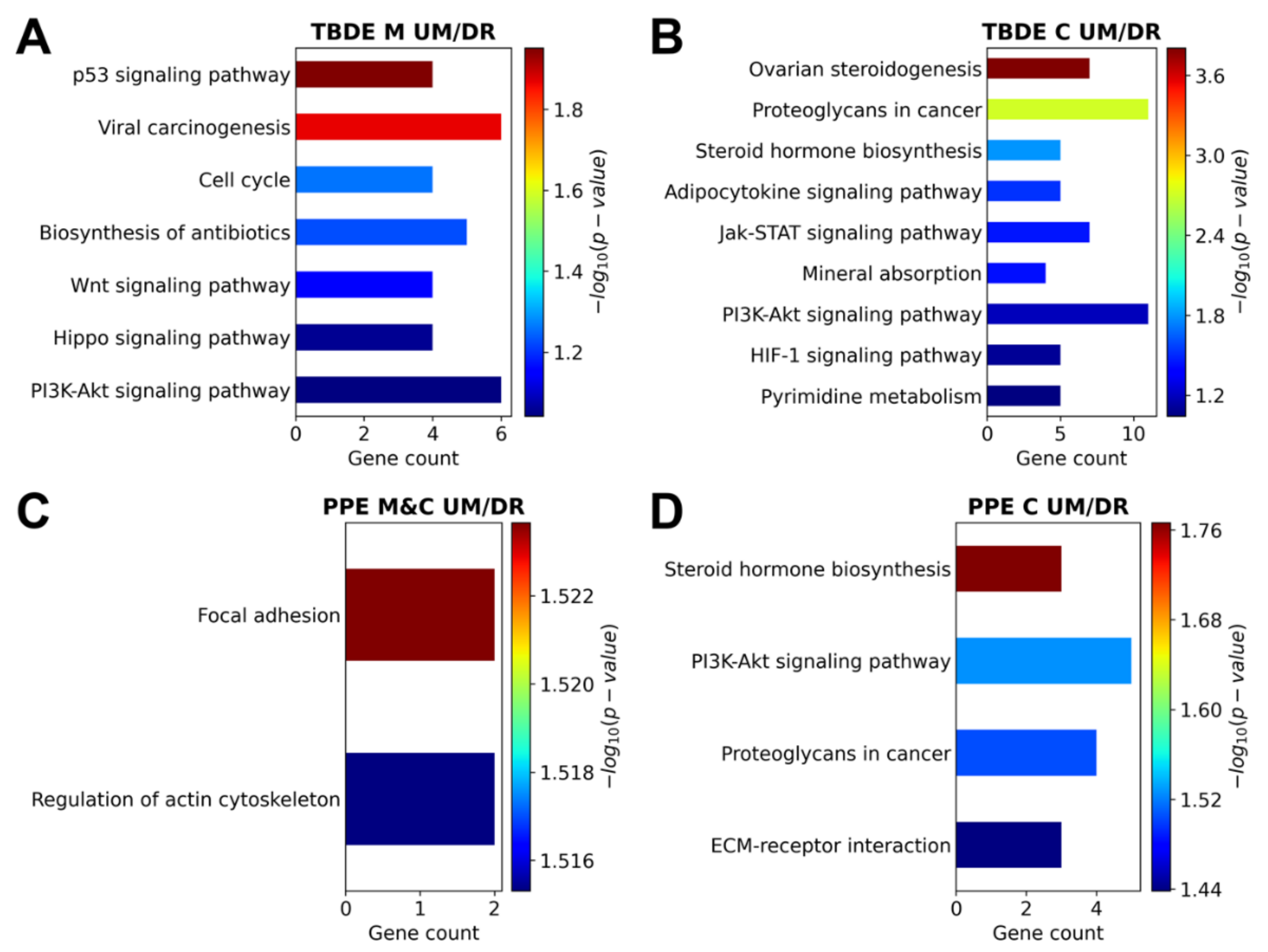
2.10. Pathway Analysis
2.11. TMA Immunoscoring Analysis
3. Results
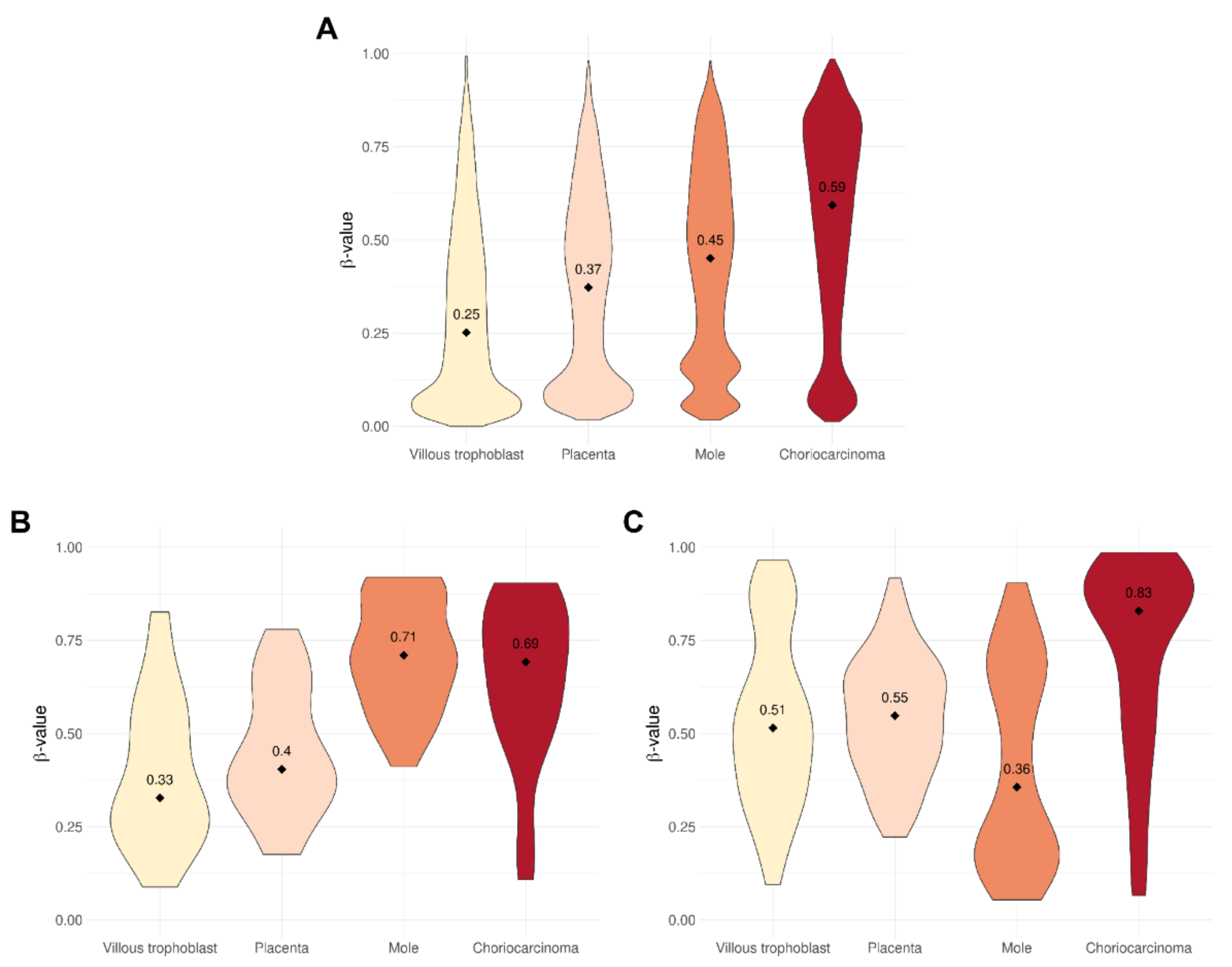
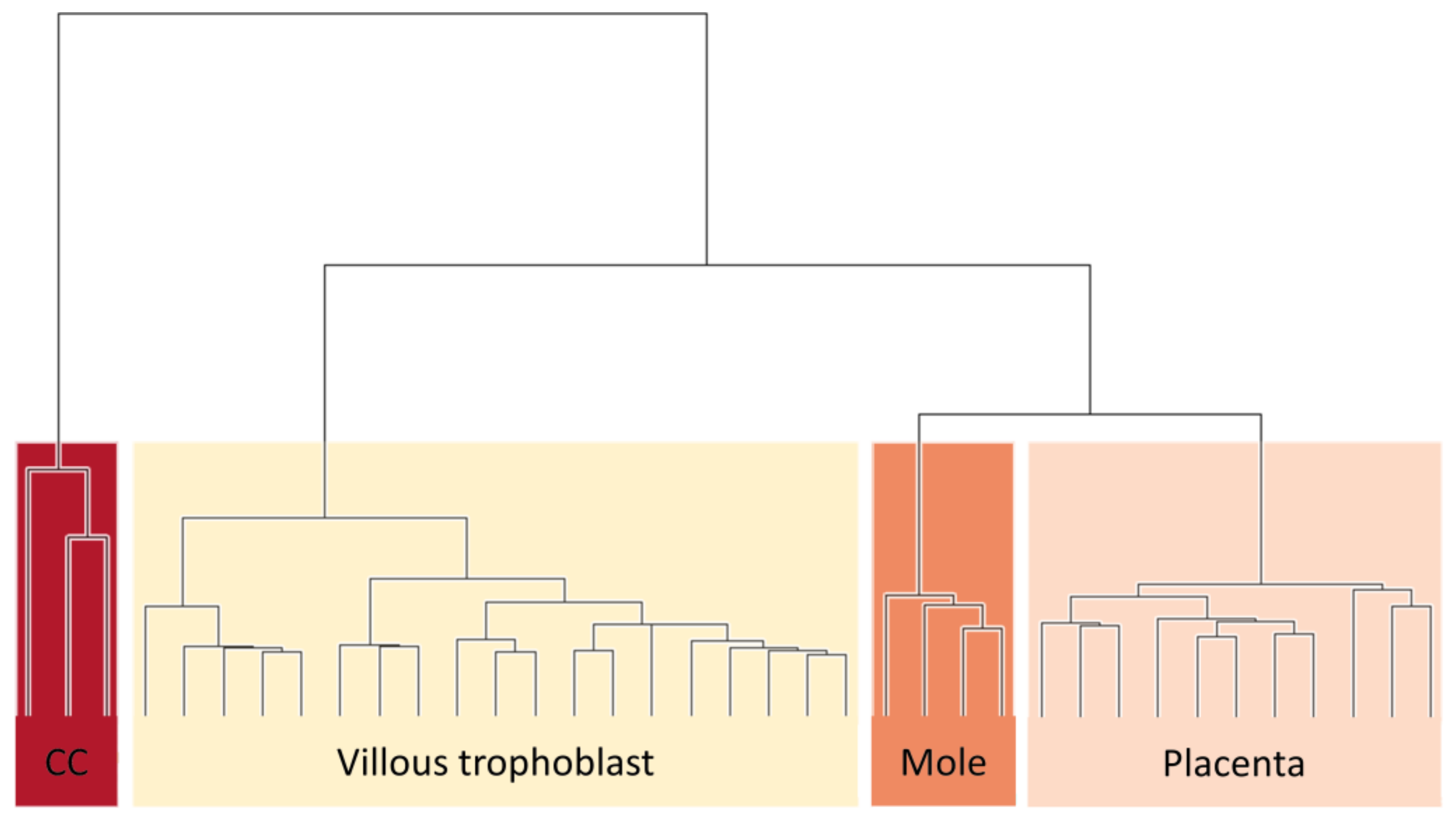
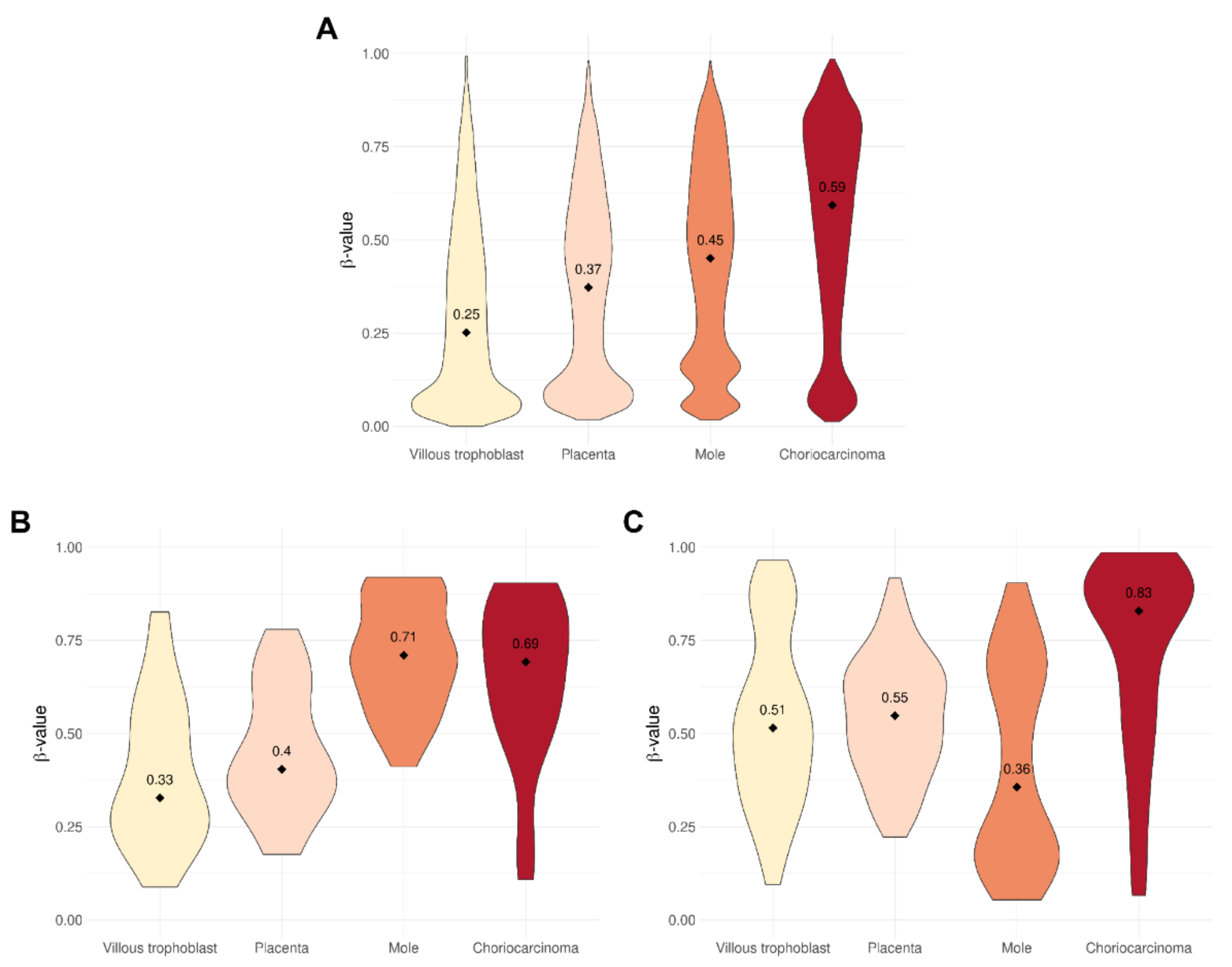
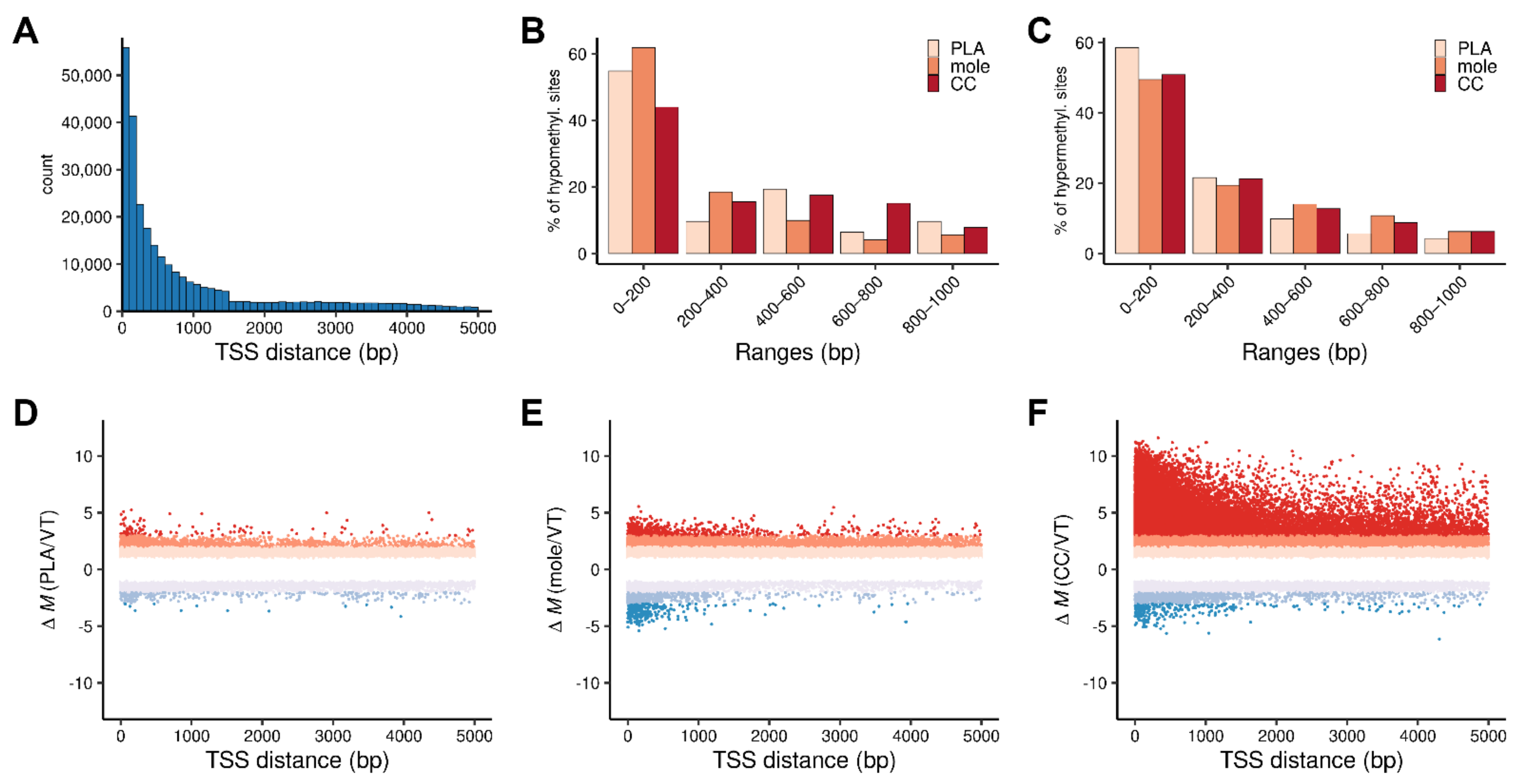
3.1. Level of DNA Methylation Is Related to Severity of Gestational Trophoblastic Disease
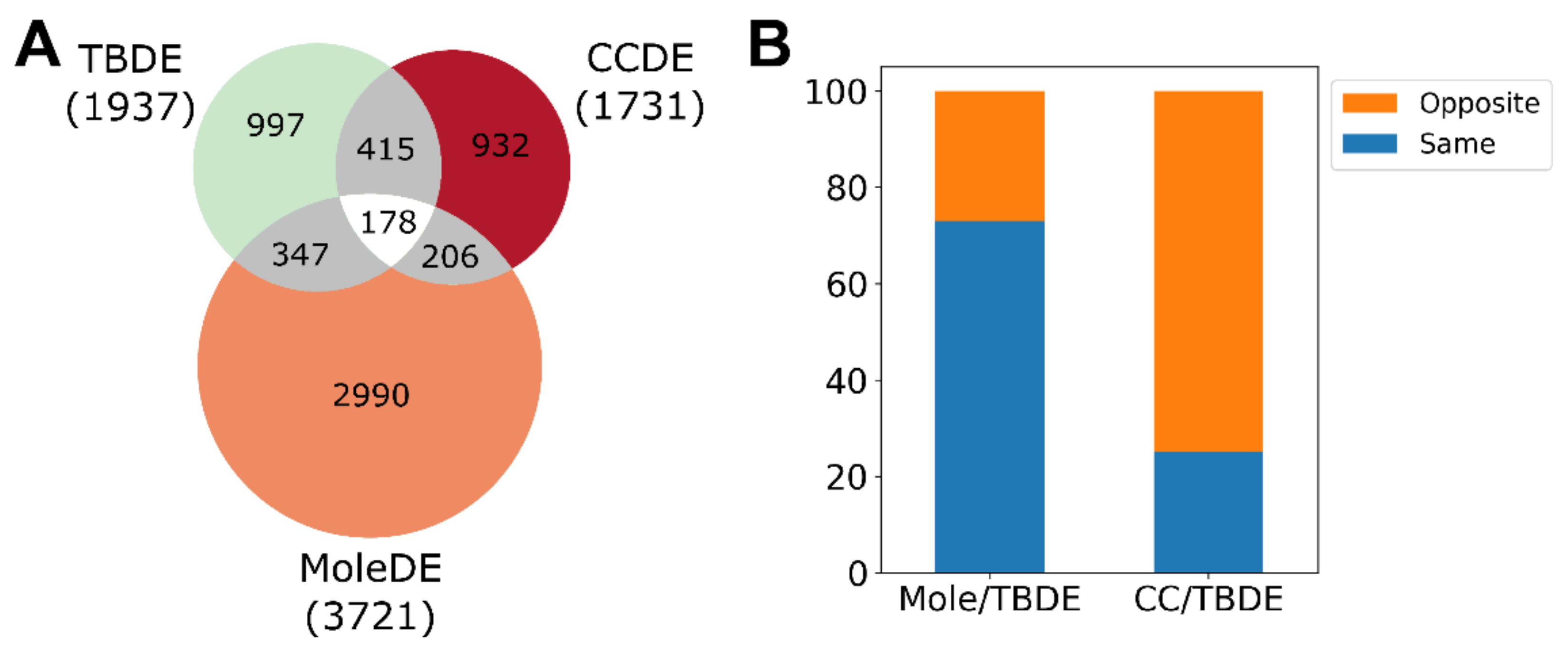
3.2. Differentially Expressed Genes Are Mainly Upregulated in Complete Moles While Predominantly Downregulated in Choriocarcinoma
3.3. DNA Methylation Influences Gene Expression of Villous Trophoblast Differentiation-Related or Predominantly Placenta-Expressed Genes
3.4. Genesis of Complete Moles and Choriocarcinoma Involve Shared and Distinct Pathways of Disease
4. Discussion
4.1. Principal Findings of this Study
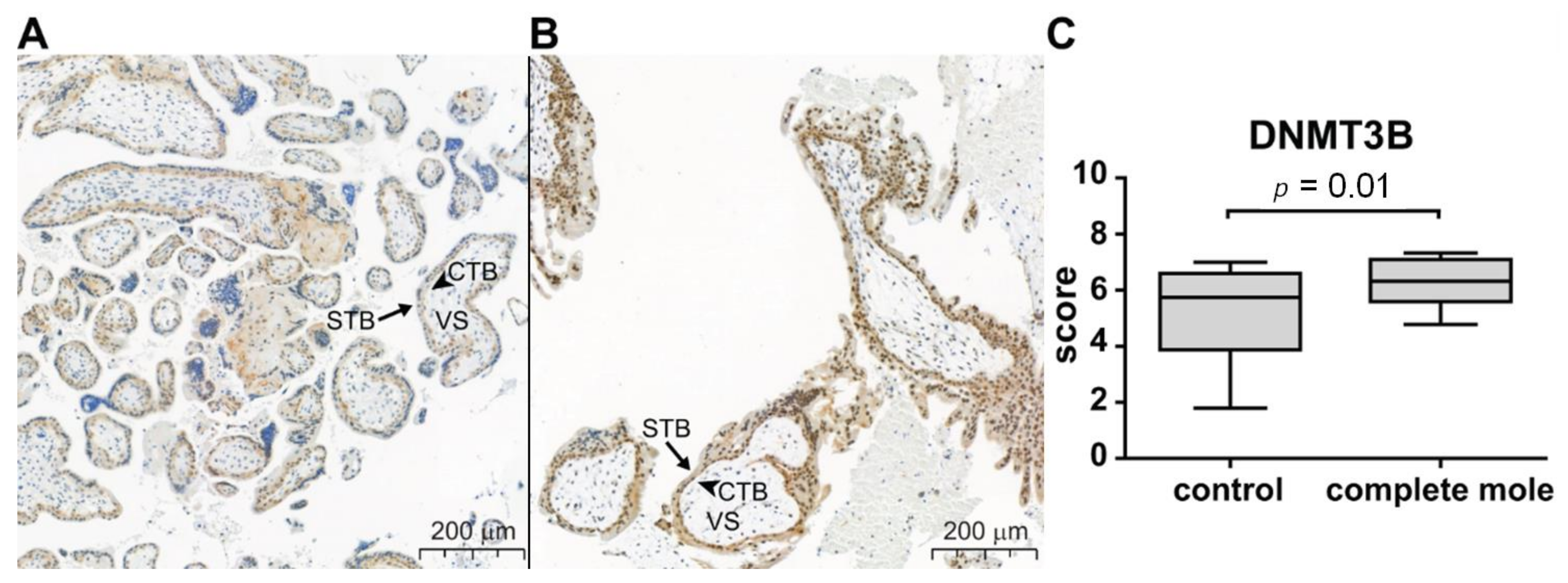
4.2. Trophoblastic Gene Expression and Functions Are Severely Affected by Epigenetic Changes in GTDs
4.3. Shared and Distinct Trophoblastic/Placental Disease Pathways in GTDs
4.4. Strengths and Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GTD Tissue Samples (n = 17) | Placenta Tissue Samples (n = 24) | |
|---|---|---|
| Race/ethnicity a | ||
| Hispanic | 18 (78%) | |
| Non-Hispanic white | 2 (9%) | 24 (100%) |
| Asian | 0 | |
| African-American | 0 | |
| Unknown/other | 3 (13%) | |
| Age (years) at procedure b | 31.9 ± 9.5 | 28.8 ± 6.2 |
| Comorbidities | ||
| Hypertension a | 1 (4%) | |
| Diabetes a | 1 (4%) | |
| β-hCG (mIU/mL) at procedure c | 185,277 (70,349–387,812) | |
| Histologic diagnosis a | ||
| Complete hydatidiform mole (CHM) | 17 (100%) | 0 (0%) |
| p57 immunostaining confirmation of CHM | 17 (100%) | |
| Gestational age at the procedure (days) b | 61.65 ± 16.97 | 62.79 ± 12.30 |
| Primary Antibody (Concentration/Dilution, Distributor) | Detection Antibody (Distributor) | Detection System (Distributor) |
|---|---|---|
| mouse monoclonal anti-human p57 antibody (1:3000) (code: MA5-11309, Thermo Fisher Scientific, Waltham, MA, USA) | Novolink detections system (Leica-Novocastra, Wetzlar, Germany) | Novolink DAB/substrate kit (Leica-Novocastra, Wetzlar, Germany) |
| rabbit polyclonal anti-human DNMT3B antibody (1:100) (code: LS-C352124, LSBio, Seattle, WA, USA) |
References
- Garcia-Sayre, J.; Castaneda, A.V.; Roman, L.D.; Matsuo, K. Diagnosis and Management of Gestational Trophoblastic Disease. In Handbook of Gynecology; Shoupe, D., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–15. ISBN 978-3-319-17002-2. [Google Scholar]
- Lurain, J.R. Gestational trophoblastic disease I: Epidemiology, pathology, clinical presentation and diagnosis of gestational trophoblastic disease, and management of hydatidiform mole. Am. J. Obstet. Gynecol. 2010, 203, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Seckl, M.J.; Sebire, N.J.; Fisher, R.A.; Golfier, F.; Massuger, L.; Sessa, C. ESMO Guidelines Working Group Gestational trophoblastic disease: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2013, 24 (Suppl. 6), vi39–vi50. [Google Scholar] [CrossRef] [PubMed]
- Ning, F.; Hou, H.; Morse, A.N.; Lash, G.E. Understanding and management of gestational trophoblastic disease. F1000Research 2019, 8, F1000 Faculty Rev-428. [Google Scholar] [CrossRef] [PubMed]
- Atrash, H.K.; Hogue, C.J.; Grimes, D.A. Epidemiology of hydatidiform mole during early gestation. Am. J. Obstet. Gynecol. 1986, 154, 906–909. [Google Scholar] [CrossRef]
- Smith, H.O. Gestational trophoblastic disease epidemiology and trends. Clin. Obstet. Gynecol. 2003, 46, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, S. Incidence of gestational trophoblastic disease by regional registration in Japan. Hum. Reprod. 1987, 2, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Seckl, M.J.; Sebire, N.J.; Berkowitz, R.S. Gestational trophoblastic disease. Lancet 2010, 376, 717–729. [Google Scholar] [CrossRef]
- Gockley, A.A.; Joseph, N.T.; Melamed, A.; Sun, S.Y.; Goodwin, B.; Bernstein, M.; Goldstein, D.P.; Berkowitz, R.S.; Horowitz, N.S. Effect of race/ethnicity on clinical presentation and risk of gestational trophoblastic neoplasia in patients with complete and partial molar pregnancy at a tertiary care referral center. Am. J. Obstet. Gynecol. 2016, 215, 334.e1-6. [Google Scholar] [CrossRef]
- Carey, L.; Nash, B.M.; Wright, D.C. Molecular genetic studies of complete hydatidiform moles. Transl. Pediatr. 2015, 4, 181–188. [Google Scholar]
- Lindor, N.M.; Ney, J.A.; Gaffey, T.A.; Jenkins, R.B.; Thibodeau, S.N.; Dewald, G.W. A genetic review of complete and partial hydatidiform moles and nonmolar triploidy. Mayo. Clin. Proc. 1992, 67, 791–799. [Google Scholar] [CrossRef] [Green Version]
- Lurain, J.R. Gestational trophoblastic disease II: Classification and management of gestational trophoblastic neoplasia. Am. J. Obstet. Gynecol. 2011, 204, 11–18. [Google Scholar] [CrossRef]
- Tomizawa, S.; Sasaki, H. Genomic imprinting and its relevance to congenital disease, infertility, molar pregnancy and induced pluripotent stem cell. J. Hum. Genet. 2012, 57, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Savage, P.; Monk, D.; Mora, J.R.H.; van der Westhuizen, N.; Rauw, J.; Tinker, A.; Robinson, W.; Song, Q.; Seckl, M.J.; Fisher, R.A. A case of intraplacental gestational choriocarcinoma; characterised by the methylation pattern of the early placenta and an absence of driver mutations. BMC Cancer 2019, 19, 744. [Google Scholar] [CrossRef]
- Hogg, K.; Robinson, W.P.; Beristain, A.G. Activation of endocrine-related gene expression in placental choriocarcinoma cell lines following DNA methylation knock-down. Mol. Hum. Reprod. 2014, 20, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Than, N.G.; Romero, R.; Xu, Y.; Erez, O.; Xu, Z.; Bhatti, G.; Leavitt, R.; Chung, T.H.; El-Azzamy, H.; LaJeunesse, C.; et al. Evolutionary origins of the placental expression of chromosome 19 cluster galectins and their complex dysregulation in preeclampsia. Placenta 2014, 35, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Wong, N.C.; Novakovic, B.; Weinrich, B.; Dewi, C.; Andronikos, R.; Sibson, M.; Macrae, F.; Morley, R.; Pertile, M.D.; Craig, J.M.; et al. Methylation of the adenomatous polyposis coli (APC) gene in human placenta and hypermethylation in choriocarcinoma cells. Cancer Lett. 2008, 268, 56–62. [Google Scholar] [CrossRef]
- King, J.R.; Wilson, M.L.; Hetey, S.; Kiraly, P.; Matsuo, K.; Castaneda, A.V.; Toth, E.; Krenacs, T.; Hupuczi, P.; Mhawech-Fauceglia, P.; et al. Dysregulation of Placental Functions and Immune Pathways in Complete Hydatidiform Moles. Int. J. Mol. Sci. 2019, 20, E4999. [Google Scholar] [CrossRef] [Green Version]
- Hargitai, B.; Marton, T.; Cox, P.M. Best practice no 178. Examination of the human placenta. J. Clin. Pathol. 2004, 57, 785–792. [Google Scholar] [CrossRef] [Green Version]
- Khong, T.Y.; Mooney, E.E.; Ariel, I.; Balmus, N.C.M.; Boyd, T.K.; Brundler, M.-A.; Derricott, H.; Evans, M.J.; Faye-Petersen, O.M.; Gillan, J.E.; et al. Sampling and Definitions of Placental Lesions: Amsterdam Placental Workshop Group Consensus Statement. Arch. Pathol. Lab. Med. 2016, 140, 698–713. [Google Scholar] [CrossRef] [Green Version]
- Redline, R.W. Placental pathology: A systematic approach with clinical correlations. Placenta 2008, 29, 86–91. [Google Scholar] [CrossRef]
- Than, N.G.; Abdul Rahman, O.; Magenheim, R.; Nagy, B.; Fule, T.; Hargitai, B.; Sammar, M.; Hupuczi, P.; Tarca, A.L.; Szabo, G.; et al. Placental protein 13 (galectin-13) has decreased placental expression but increased shedding and maternal serum concentrations in patients presenting with preterm pre-eclampsia and HELLP syndrome. Virchows Arch. 2008, 453, 387–400. [Google Scholar] [CrossRef] [Green Version]
- Aryee, M.J.; Jaffe, A.E.; Corrada-Bravo, H.; Ladd-Acosta, C.; Feinberg, A.P.; Hansen, K.D.; Irizarry, R.A. Minfi: A flexible and comprehensive Bioconductor package for the analysis of Infinium DNA methylation microarrays. Bioinformatics 2014, 30, 1363–1369. [Google Scholar] [CrossRef] [Green Version]
- Fortin, J.-P.; Labbe, A.; Lemire, M.; Zanke, B.W.; Hudson, T.J.; Fertig, E.J.; Greenwood, C.M.; Hansen, K.D. Functional normalization of 450k methylation array data improves replication in large cancer studies. Genome Biol. 2014, 15, 503. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Than, N.G.; Romero, R.; Tarca, A.L.; Kekesi, K.A.; Xu, Y.; Xu, Z.; Juhasz, K.; Bhatti, G.; Leavitt, R.J.; Gelencser, Z.; et al. Integrated Systems Biology Approach Identifies Novel Maternal and Placental Pathways of Preeclampsia. Front. Immunol. 2018, 9, 1661. [Google Scholar] [CrossRef]
- Monk, D.; Morales, J.; den Dunnen, J.T.; Russo, S.; Court, F.; Prawitt, D.; Eggermann, T.; Beygo, J.; Buiting, K.; Tümer, Z.; et al. Recommendations for a nomenclature system for reporting methylation aberrations in imprinted domains. Epigenetics 2018, 13, 117–121. [Google Scholar] [CrossRef]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. affy-analysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deaton, A.M.; Bird, A. CpG islands and the regulation of transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krinner, S.; Heitzer, A.P.; Diermeier, S.D.; Obermeier, I.; Längst, G.; Wagner, R. CpG domains downstream of TSSs promote high levels of gene expression. Nucleic Acids Res. 2014, 42, 3551–3564. [Google Scholar] [CrossRef]
- Szilagyi, A.; Gelencser, Z.; Romero, R.; Xu, Y.; Kiraly, P.; Demeter, A.; Palhalmi, J.; Gyorffy, B.A.; Juhasz, K.; Hupuczi, P.; et al. Placenta-Specific Genes, Their Regulation During Villous Trophoblast Differentiation and Dysregulation in Preterm Preeclampsia. Int. J. Mol. Sci. 2020, 21, E628. [Google Scholar] [CrossRef] [Green Version]
- Bilban, M.; Haslinger, P.; Prast, J.; Klinglmüller, F.; Woelfel, T.; Haider, S.; Sachs, A.; Otterbein, L.E.; Desoye, G.; Hiden, U.; et al. Identification of novel trophoblast invasion-related genes: Heme oxygenase-1 controls motility via peroxisome proliferator-activated receptor gamma. Endocrinology 2009, 150, 1000–1013. [Google Scholar] [CrossRef] [Green Version]
- Bilban, M.; Tauber, S.; Haslinger, P.; Pollheimer, J.; Saleh, L.; Pehamberger, H.; Wagner, O.; Knöfler, M. Trophoblast invasion: Assessment of cellular models using gene expression signatures. Placenta 2010, 31, 989–996. [Google Scholar] [CrossRef]
- Jia, D.; Jurkowska, R.Z.; Zhang, X.; Jeltsch, A.; Cheng, X. Structure of Dnmt3a bound to Dnmt3L suggests a model for de novo DNA methylation. Nature 2007, 449, 248–251. [Google Scholar] [CrossRef] [Green Version]
- Gu, T.-P.; Guo, F.; Yang, H.; Wu, H.-P.; Xu, G.-F.; Liu, W.; Xie, Z.-G.; Shi, L.; He, X.; Jin, S.; et al. The role of Tet3 DNA dioxygenase in epigenetic reprogramming by oocytes. Nature 2011, 477, 606–610. [Google Scholar] [CrossRef]
- Nottke, A.; Colaiácovo, M.P.; Shi, Y. Developmental roles of the histone lysine demethylases. Development 2009, 136, 879–889. [Google Scholar] [CrossRef] [Green Version]
- Gagliardi, M.; Strazzullo, M.; Matarazzo, M.R. DNMT3B Functions: Novel Insights From Human Disease. Front. Cell Dev. Biol. 2018, 6, 140. [Google Scholar] [CrossRef]
- Reik, W.; Dean, W.; Walter, J. Epigenetic reprogramming in mammalian development. Science 2001, 293, 1089–1093. [Google Scholar] [CrossRef] [Green Version]
- Hervouet, E.; Peixoto, P.; Delage-Mourroux, R.; Boyer-Guittaut, M.; Cartron, P.-F. Specific or not specific recruitment of DNMTs for DNA methylation, an epigenetic dilemma. Clin. Epigenet. 2018, 10, 17. [Google Scholar] [CrossRef]
- Kawakami, E.; Tokunaga, A.; Ozawa, M.; Sakamoto, R.; Yoshida, N. The histone demethylase Fbxl11/Kdm2a plays an essential role in embryonic development by repressing cell-cycle regulators. Mech. Dev. 2015, 135, 31–42. [Google Scholar] [CrossRef]
- Herzog, M.; Josseaux, E.; Dedeurwaerder, S.; Calonne, E.; Volkmar, M.; Fuks, F. The histone demethylase Kdm3a is essential to progression through differentiation. Nucleic Acids Res. 2012, 40, 7219–7232. [Google Scholar] [CrossRef]
- Chakraborty, D.; Cui, W.; Rosario, G.X.; Scott, R.L.; Dhakal, P.; Renaud, S.J.; Tachibana, M.; Rumi, M.A.K.; Mason, C.W.; Krieg, A.J.; et al. HIF-KDM3A-MMP12 regulatory circuit ensures trophoblast plasticity and placental adaptations to hypoxia. Proc. Natl. Acad. Sci. USA 2016, 113, E7212–E7221. [Google Scholar] [CrossRef] [Green Version]
- Mak, K.H.-M.; Lam, Y.M.; Ng, R.K. Histone demethylase JMJD2B/KDM4B regulates transcriptional program via distinctive epigenetic targets and protein interactors for the maintenance of trophoblast stem cells. Sci. Rep. 2021, 11, 884. [Google Scholar] [CrossRef]
- Dahl, J.A.; Jung, I.; Aanes, H.; Greggains, G.D.; Manaf, A.; Lerdrup, M.; Li, G.; Kuan, S.; Li, B.; Lee, A.Y.; et al. Broad histone H3K4me3 domains in mouse oocytes modulate maternal-to-zygotic transition. Nature 2016, 537, 548–552. [Google Scholar] [CrossRef]
- Bai, G.-Y.; Song, S.-H.; Zhang, Y.-W.; Huang, X.; Huang, X.-W.; Sun, R.-Z.; Lei, L. Kdm6a overexpression improves the development of cloned mouse embryos. Zygote 2018, 26, 24–32. [Google Scholar] [CrossRef]
- Shen, H.; Shih, J.; Hollern, D.P.; Wang, L.; Bowlby, R.; Tickoo, S.K.; Thorsson, V.; Mungall, A.J.; Newton, Y.; Hegde, A.M.; et al. Integrated Molecular Characterization of Testicular Germ Cell Tumors. Cell Rep. 2018, 23, 3392–3406. [Google Scholar] [CrossRef]
- Than, G.N.; Tatra, G.; Szabó, D.G.; Csaba, I.F.; Bohn, H. Beta lactoglobulin homologue placental protein 14 (PP14) in serum of patients with trophoblastic disease and non-trophoblastic gynecologic malignancy. Arch. Gynecol. Obstet. 1988, 243, 131–137. [Google Scholar] [CrossRef]
- Lee, J.N.; Salem, H.T.; Chard, T.; Huang, S.C.; Ouyang, P.C. Circulating placental proteins (hCG, SP1 and PP5) in trophoblastic disease. Br. J. Obstet. Gynaecol. 1982, 89, 69–72. [Google Scholar] [CrossRef]
- Than, G.N.; Csaba, I.F.; Szabó, D.G.; Bognár, Z.J.; Arany, A.; Bohn, H. Levels of placenta-specific tissue protein 12 (PP12) in serum during normal pregnancy and in patients with trophoblastic tumour. Arch. Gynecol. 1983, 234, 39–46. [Google Scholar] [CrossRef]
- Lee, J.N.; Salem, H.T.; Al-Ani, A.T.; Chard, T.; Huang, S.C.; Ouyang, P.C.; Wei, P.Y.; Seppälä, M. Circulating concentrations of specific placental proteins (human chorionic gonadotropin, pregnancy-specific beta-1 glycoprotein, and placental protein 5) in untreated gestational trophoblastic tumors. Am. J. Obstet. Gynecol. 1981, 139, 702–704. [Google Scholar] [CrossRef]
- Candelier, J.-J.; Frappart, L.; Diatta, A.L.; Yadaden, T.; Cissé, M.-L.; Afoutou, J.-M.; Picard, J.-Y.; Mennesson, B.; Slim, R.; Si-Tayeb, K.; et al. Differential expression of E-cadherin, β-catenin, and Lewis x between invasive hydatidiform moles and post-molar choriocarcinomas. Virchows Arch. 2013, 462, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Nagymanyoki, Z.; Callahan, M.J.; Parast, M.M.; Berkowitz, R.S.; Mok, S.C.; Fulop, V. Overexpression of laminin receptor 1 on decidual cells in partial and complete mole. Gynecol. Oncol. 2008, 108, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Crescimanno, C.; Marzioni, D.; Paradinas, F.J.; Schrurs, B.; Mühlhauser, J.; Todros, T.; Newlands, E.; David, G.; Castellucci, M. Expression pattern alterations of syndecans and glypican-1 in normal and pathological trophoblast. J. Pathol. 1999, 189, 600–608. [Google Scholar] [CrossRef]
- Ockleford, C.; Barker, C.; Griffiths, J.; McTurk, G.; Fisher, R.; Lawler, S. Hydatidiform mole: An ultrastructural analysis of syncytiotrophoblast surface organization. Placenta 1989, 10, 195–212. [Google Scholar] [CrossRef]
- Sonderegger, S.; Pollheimer, J.; Knöfler, M. Wnt signalling in implantation, decidualisation and placental differentiation-review. Placenta 2010, 31, 839–847. [Google Scholar] [CrossRef] [Green Version]
- Knöfler, M.; Pollheimer, J. Human placental trophoblast invasion and differentiation: A particular focus on Wnt signaling. Front. Genet. 2013, 4, 190. [Google Scholar] [CrossRef] [Green Version]
- Meinhardt, G.; Haider, S.; Kunihs, V.; Saleh, L.; Pollheimer, J.; Fiala, C.; Hetey, S.; Feher, Z.; Szilagyi, A.; Than, N.G.; et al. Pivotal role of the transcriptional co-activator YAP in trophoblast stemness of the developing human placenta. Proc. Natl. Acad. Sci. USA 2020, 117, 13562–13570. [Google Scholar] [CrossRef]
- Saha, B.; Ganguly, A.; Home, P.; Bhattacharya, B.; Ray, S.; Ghosh, A.; Rumi, M.A.K.; Marsh, C.; French, V.A.; Gunewardena, S.; et al. TEAD4 ensures postimplantation development by promoting trophoblast self-renewal: An implication in early human pregnancy loss. Proc. Natl. Acad. Sci. USA 2020, 117, 17864–17875. [Google Scholar] [CrossRef]
- Crocker, I. Gabor Than Award Lecture 2006: Pre-eclampsia and villous trophoblast turnover: Perspectives and possibilities. Placenta 2007, 28, S4–S13. [Google Scholar] [CrossRef]
- Heazell, A.E.P.; Crocker, I.P. Live and let die—Regulation of villous trophoblast apoptosis in normal and abnormal pregnancies. Placenta 2008, 29, 772–783. [Google Scholar] [CrossRef]
- Pollheimer, J.; Knöfler, M. Signalling pathways regulating the invasive differentiation of human trophoblasts: A review. Placenta 2005, 26, S21–S30. [Google Scholar] [CrossRef]
- Ferretti, C.; Bruni, L.; Dangles-Marie, V.; Pecking, A.P.; Bellet, D. Molecular circuits shared by placental and cancer cells, and their implications in the proliferative, invasive and migratory capacities of trophoblasts. Hum. Reprod. Update 2007, 13, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Mascaró, M.; Pibuel, M.A.; Lompardía, S.L.; Díaz, M.; Zotta, E.; Bianconi, M.I.; Lago, N.; Otero, S.; Jankilevich, G.; Alvarez, E.; et al. Low molecular weight hyaluronan induces migration of human choriocarcinoma JEG-3 cells mediated by RHAMM as well as by PI3K and MAPK pathways. Histochem. Cell Biol. 2017, 148, 173–187. [Google Scholar] [CrossRef]
- Fulop, V.; Mok, S.C.; Berkowitz, R.S. Molecular biology of gestational trophoblastic neoplasia: A review. J. Reprod. Med. 2004, 49, 415–422. [Google Scholar]
- Lash, G.E.; Quenby, S.; Burton, G.J.; Nakashima, A.; Kamat, B.R.; Ray, J.; Bulmer, J.N. Gestational diseases—A workshop report. Placenta 2008, 29, S92–S94. [Google Scholar] [CrossRef]
- Pires, K.S.N.; Sun, S.Y.; Gonçalves, C.M.; Santos, J.C.; Tenorio, L.P.G.; Botelho, R.M.; Silva, A.L.M.; Oliveira, H.G.S.; Pendeloski, K.P.T.; Daher, S.; et al. Versican silencing in BeWo cells and its implication in gestational trophoblastic diseases. Histochem. Cell. Biol. 2019, 151, 305–313. [Google Scholar] [CrossRef]
- Kang, J.; Wang, J.; Tian, J.; Shi, R.; Jia, H.; Wang, Y. The emerging role of EGFL6 in angiogenesis and tumor progression. Int. J. Med. Sci. 2020, 17, 1320–1326. [Google Scholar] [CrossRef]
- Holbro, T.; Beerli, R.R.; Maurer, F.; Koziczak, M.; Barbas, C.F.; Hynes, N.E. The ErbB2/ErbB3 heterodimer functions as an oncogenic unit: ErbB2 requires ErbB3 to drive breast tumor cell proliferation. Proc. Natl. Acad. Sci. USA 2003, 100, 8933–8938. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Zhang, Z.; Jia, C.; Li, J.; Yin, L.; Jiang, S. The relationship between expression of c-ras, c-erbB-2, nm23, and p53 gene products and development of trophoblastic tumor and their predictive significance for the malignant transformation of complete hydatidiform mole. Gynecol. Oncol. 2002, 85, 438–444. [Google Scholar] [CrossRef]
- Fülöp, V.; Végh, G.; Doszpod, J. The c-erbB-related oncoproteins in normal placenta and in gestational trophoblastic diseases (in vitro study). Orv. Hetil. 2001, 142, 1147–1154. [Google Scholar]
- Fulop, V.; Mok, S.C.; Genest, D.R.; Szigetvari, I.; Cseh, I.; Berkowitz, R.S. c-myc, c-erbB-2, c-fms and bcl-2 oncoproteins. Expression in normal placenta, partial and complete mole, and choriocarcinoma. J. Reprod. Med. 1998, 43, 101–110. [Google Scholar]
- Kumagai, S.; Koyama, S.; Nishikawa, H. Antitumour immunity regulated by aberrant ERBB family signalling. Nat. Rev. Cancer 2021, 21, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Altmäe, S.; Segura, M.T.; Esteban, F.J.; Bartel, S.; Brandi, P.; Irmler, M.; Beckers, J.; Demmelmair, H.; López-Sabater, C.; Koletzko, B.; et al. Maternal Pre-Pregnancy Obesity Is Associated with Altered Placental Transcriptome. PLoS ONE 2017, 12, e0169223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuan, Y.H.; Choi, Y.L.; Shin, Y.K.; Ahn, G.H.; Kim, K.H.; Kim, W.J.; Lee, H.C.; Kim, S.H. Expression of TGF-beta signaling proteins in normal placenta and gestational trophoblastic disease. Histol. Histopathol. 2007, 22, 227–234. [Google Scholar] [PubMed]
- Collet, C.; Lopez, J.; Battail, C.; Allias, F.; Devouassoux-Shisheboran, M.; Patrier, S.; Lemaitre, N.; Hajri, T.; Massardier, J.; You, B.; et al. Transcriptomic Characterization of Postmolar Gestational Choriocarcinoma. Biomedicines 2021, 9, 1474. [Google Scholar] [CrossRef]
- Fülöp, V.; Vermes, G.; Demeter, J. The relationship between inflammatory and immunological processes during pregnancy. Practical aspects. Orv. Hetil. 2019, 160, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Reynaud, D.; Abi Nahed, R.; Lemaitre, N.; Bolze, P.-A.; Traboulsi, W.; Sergent, F.; Battail, C.; Filhol, O.; Sapin, V.; Boufettal, H.; et al. NLRP7 Promotes Choriocarcinoma Growth and Progression through the Establishment of an Immunosuppressive Microenvironment. Cancers 2021, 13, 2999. [Google Scholar] [CrossRef]
- Kilic, M.; Flossmann, E.; Flossmann, O.; Vogelsang, H.; Junker, U.; Chaouat, G.; Markert, U.R. Jeg-3 human choriocarcinoma-induced immunosuppression: Downregulation of interleukin-2, interleukin-2 receptor alpha-chain, and its Jak/Stat signaling pathway. Am. J. Reprod. Immunol. 1999, 41, 61–69. [Google Scholar] [CrossRef]
- Mary Photini, S.; Chaiwangyen, W.; Weber, M.; Al-Kawlani, B.; Favaro, R.R.; Jeschke, U.; Schleussner, E.; Morales-Prieto, D.M.; Markert, U.R. PIM kinases 1, 2 and 3 in intracellular LIF signaling, proliferation and apoptosis in trophoblastic cells. Exp Cell Res 2017, 359, 275–283. [Google Scholar] [CrossRef]
- Borg, A.J.; Yong, H.E.J.; Lappas, M.; Degrelle, S.A.; Keogh, R.J.; Da Silva-Costa, F.; Fournier, T.; Abumaree, M.; Keelan, J.A.; Kalionis, B.; et al. Decreased STAT3 in human idiopathic fetal growth restriction contributes to trophoblast dysfunction. Reproduction 2015, 149, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Sousa, F.L.P.; Morales Prieto, D.M.; Ospina Prieto, S.; Chaiwangyen, W.; Daher, S.; Sass, N.; Markert, U.R. PP007. Effects of STAT1 suppression on ERK1/2 in trophoblastic cells. Pregnancy Hypertens 2012, 2, 243. [Google Scholar] [CrossRef]


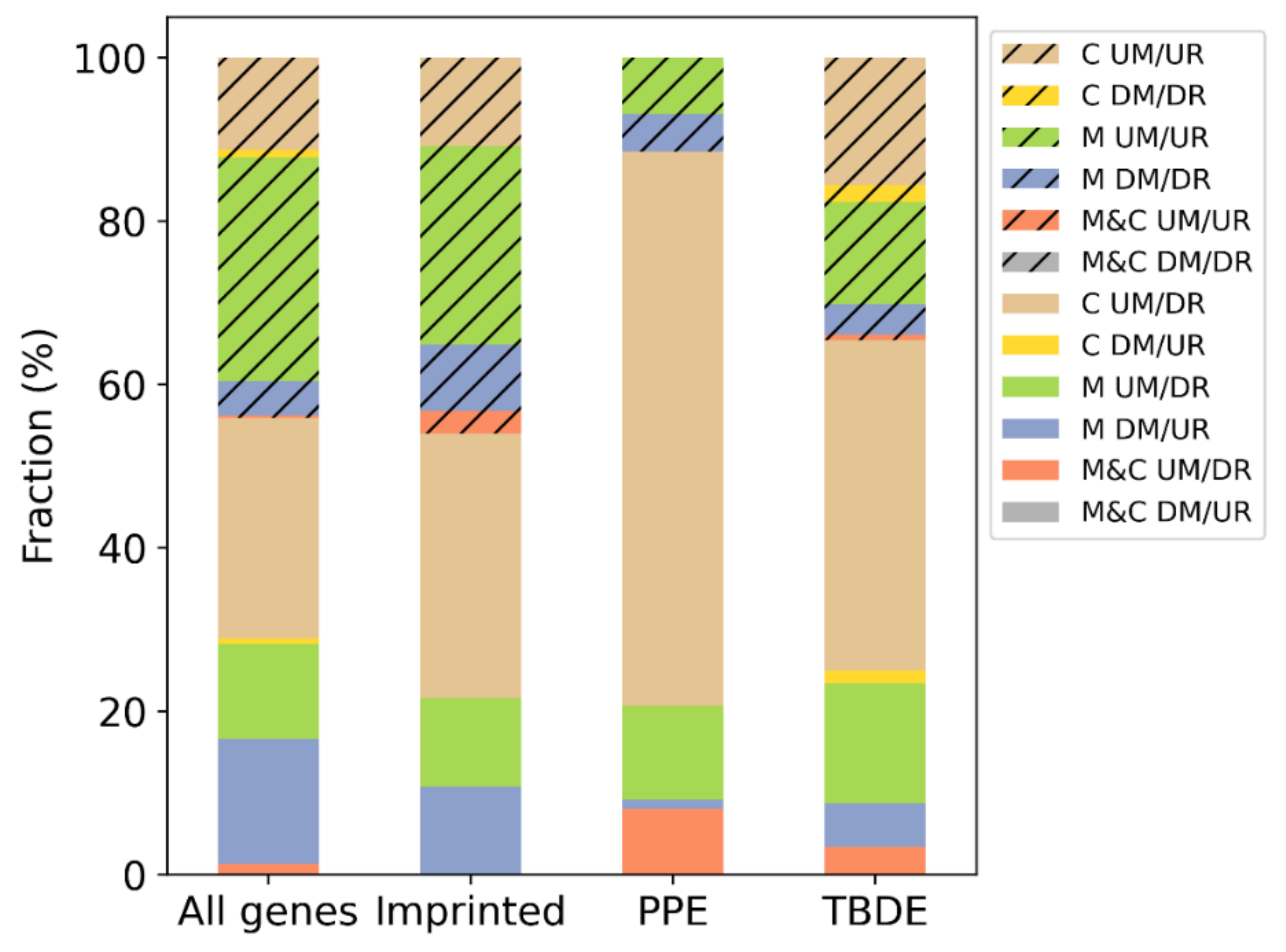
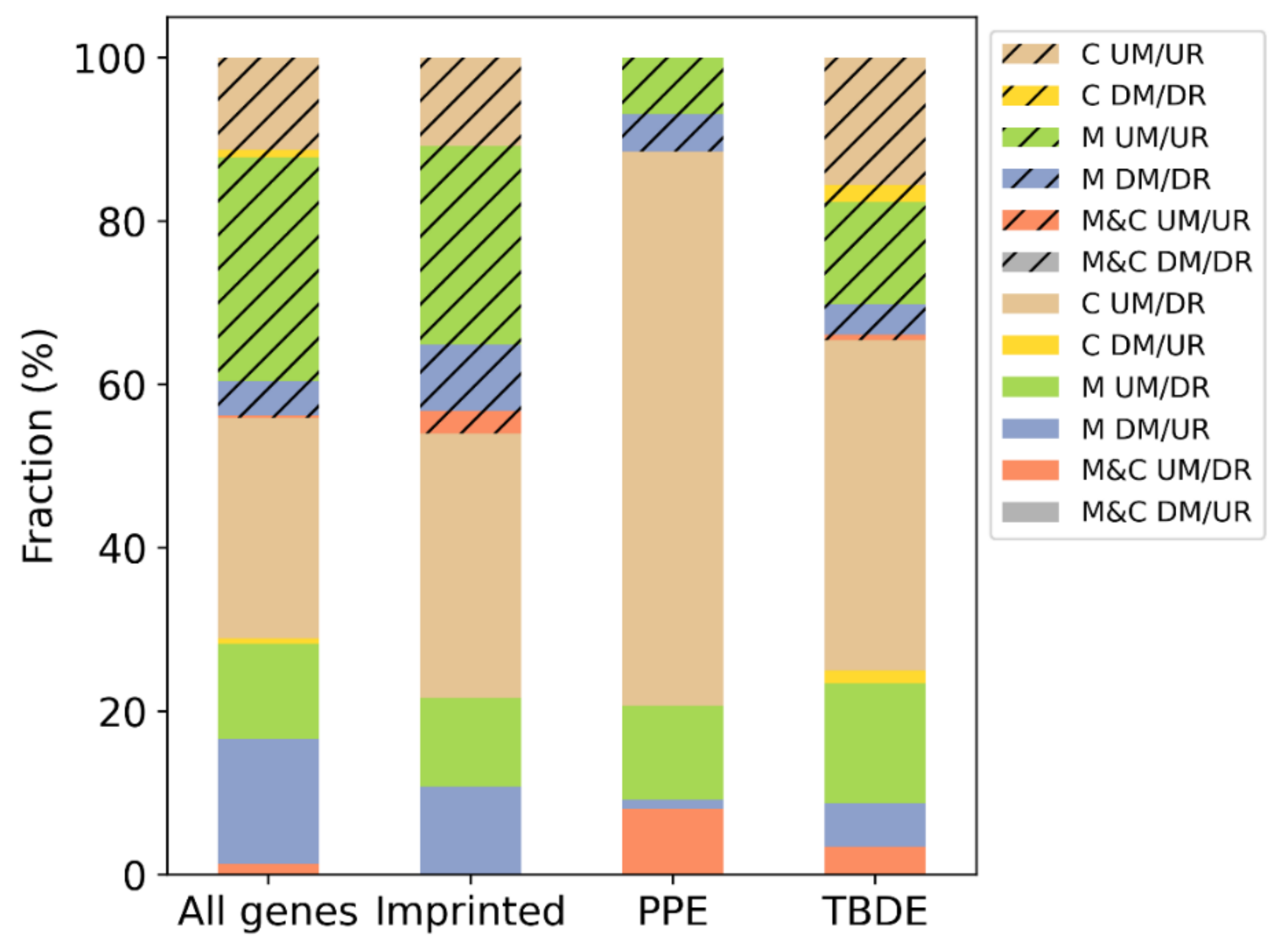
| Subset | Number of Genes | Imprinted (204) | PPE (164) | TBDE (1937) |
|---|---|---|---|---|
| M&C DM/UR | 1 | 0 | 0 | 0 |
| M&C UM/DR | 34 | 0 | 7 | 20 |
| M DM/UR | 405 | 4 | 1 | 31 |
| C DM/UR | 17 | 0 | 0 | 9 |
| M UM/DR | 307 | 4 | 10 | 86 |
| C UM/DR | 714 | 12 | 59 | 236 |
| M&C DM/DR | 0 | 0 | 0 | 0 |
| M&C UM/UR | 9 | 1 | 0 | 4 |
| M DM/DR | 111 | 3 | 4 | 22 |
| M UM/UR | 723 | 9 | 6 | 73 |
| C DM/DR | 25 | 0 | 0 | 12 |
| C UM/UR | 298 | 4 | 0 | 91 |
| Total | 2644 | 37 | 87 | 584 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szabolcsi, Z.; Demeter, A.; Kiraly, P.; Balogh, A.; Wilson, M.L.; King, J.R.; Hetey, S.; Gelencser, Z.; Matsuo, K.; Hargitai, B.; et al. Epigenetic Dysregulation of Trophoblastic Gene Expression in Gestational Trophoblastic Disease. Biomedicines 2021, 9, 1935. https://doi.org/10.3390/biomedicines9121935
Szabolcsi Z, Demeter A, Kiraly P, Balogh A, Wilson ML, King JR, Hetey S, Gelencser Z, Matsuo K, Hargitai B, et al. Epigenetic Dysregulation of Trophoblastic Gene Expression in Gestational Trophoblastic Disease. Biomedicines. 2021; 9(12):1935. https://doi.org/10.3390/biomedicines9121935
Chicago/Turabian StyleSzabolcsi, Zoltan, Amanda Demeter, Peter Kiraly, Andrea Balogh, Melissa L. Wilson, Jennifer R. King, Szabolcs Hetey, Zsolt Gelencser, Koji Matsuo, Beata Hargitai, and et al. 2021. "Epigenetic Dysregulation of Trophoblastic Gene Expression in Gestational Trophoblastic Disease" Biomedicines 9, no. 12: 1935. https://doi.org/10.3390/biomedicines9121935
APA StyleSzabolcsi, Z., Demeter, A., Kiraly, P., Balogh, A., Wilson, M. L., King, J. R., Hetey, S., Gelencser, Z., Matsuo, K., Hargitai, B., Mhawech-Fauceglia, P., Hupuczi, P., Szilagyi, A., Papp, Z., Roman, L. D., Cortessis, V. K., & Than, N. G. (2021). Epigenetic Dysregulation of Trophoblastic Gene Expression in Gestational Trophoblastic Disease. Biomedicines, 9(12), 1935. https://doi.org/10.3390/biomedicines9121935