
A Hyaluronic Acid Demilune Scaffold and Polypyrrole-Coated Fibers Carrying Embedded Human Neural Precursor Cells and Curcumin for Surface Capping of Spinal Cord Injuries

 ,
,  , , , ,
, , , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Embedding iNPCs and CURC within PM
2.2. Cell Viability and Apoptosis Assays
2.3. Phenotypic Characterization of Embedded iNPCs
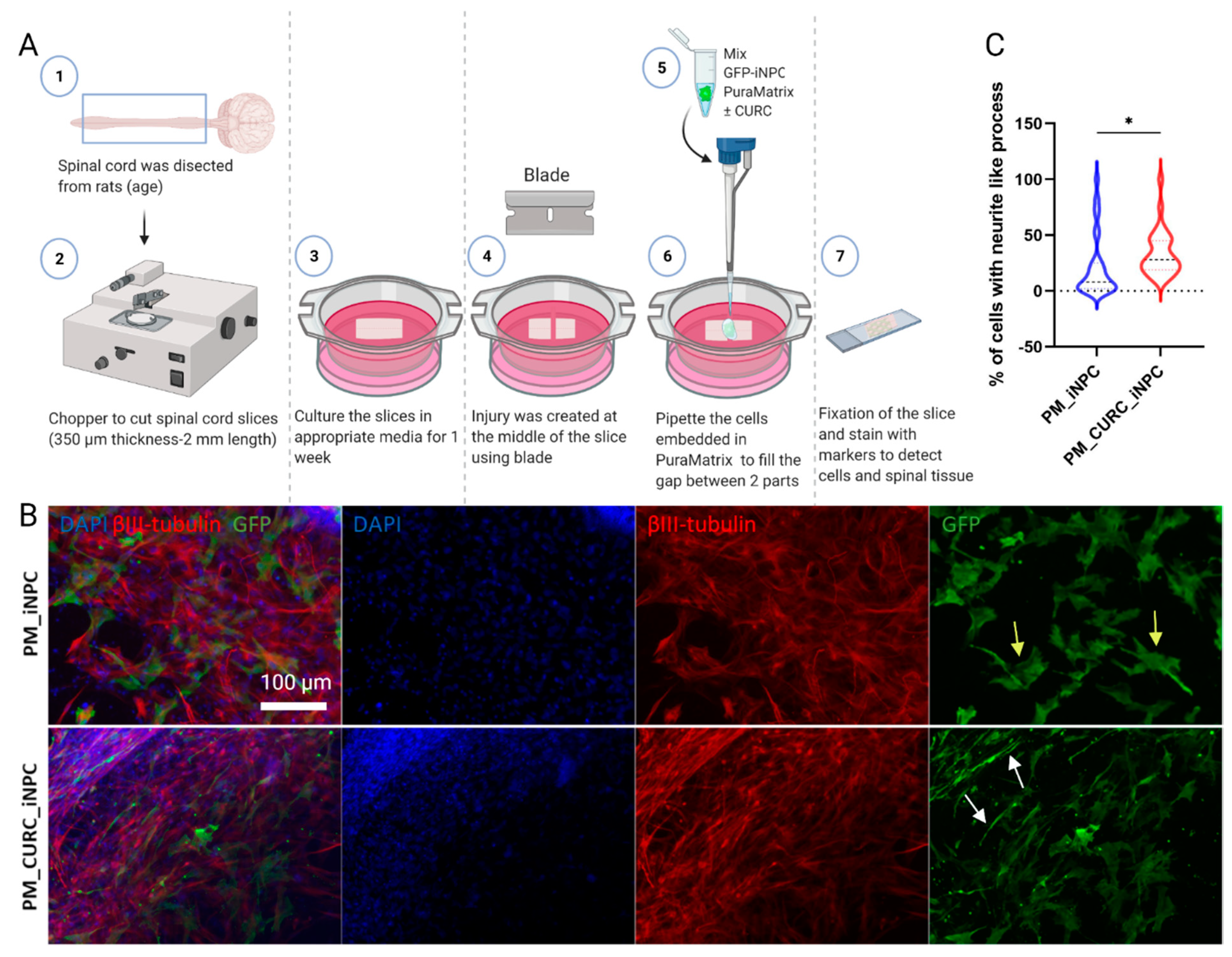
2.4. Organotypic Longitudinal Spinal Cord Slice Culture
2.5. Preparation of HA Demilune with PLA Fibers for In Vitro Experiments
2.6. Preparation, Combination with HA Demilune Scaffolds, and Sanitization of PPY-Coated Microfibers
2.7. Scanning Electron Microscopy
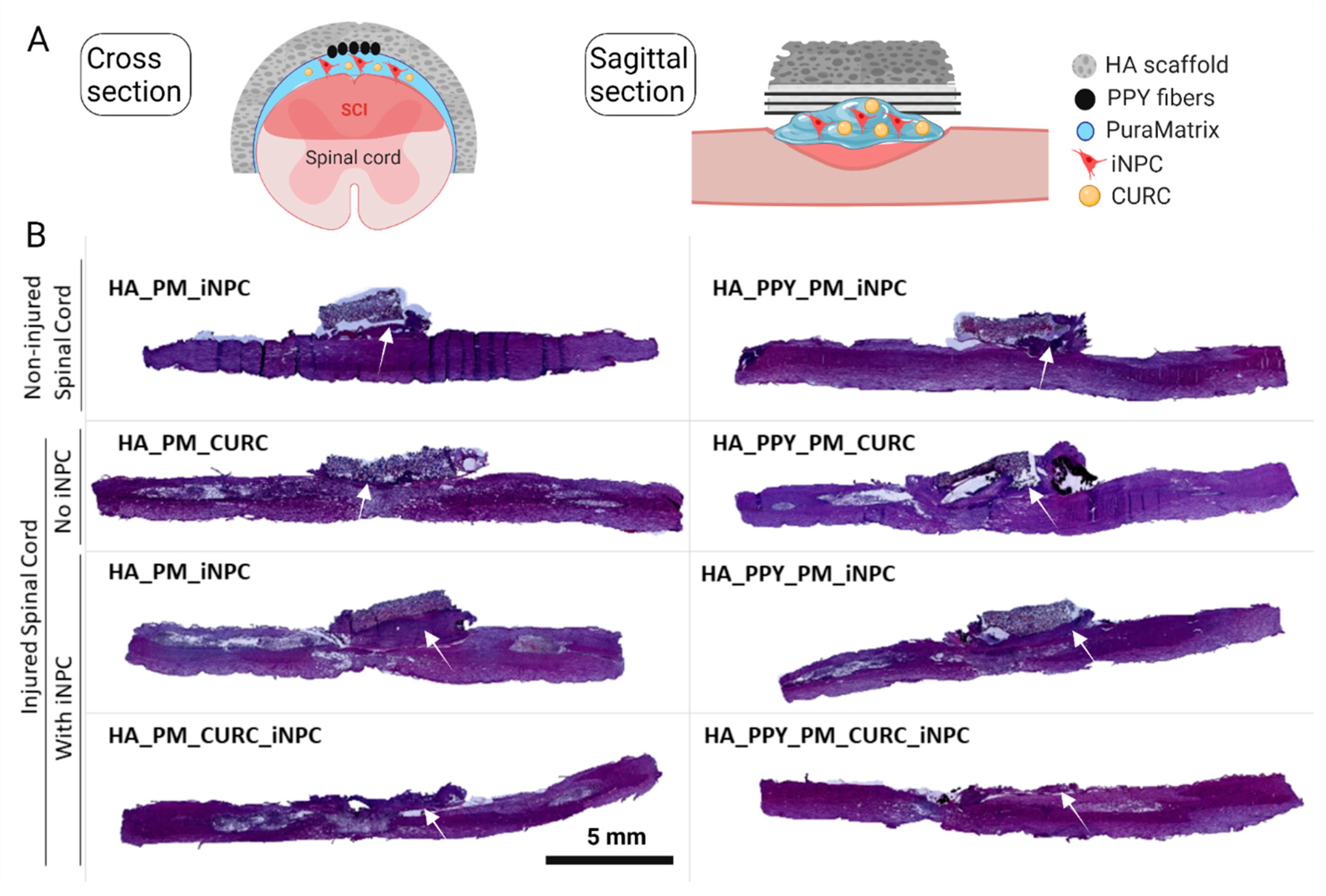
2.8. In Vivo SCI Model and Biomaterial Implantation
2.9. Histological Studies
2.10. Ethical Statement
2.11. Statistical Analysis
3. Results
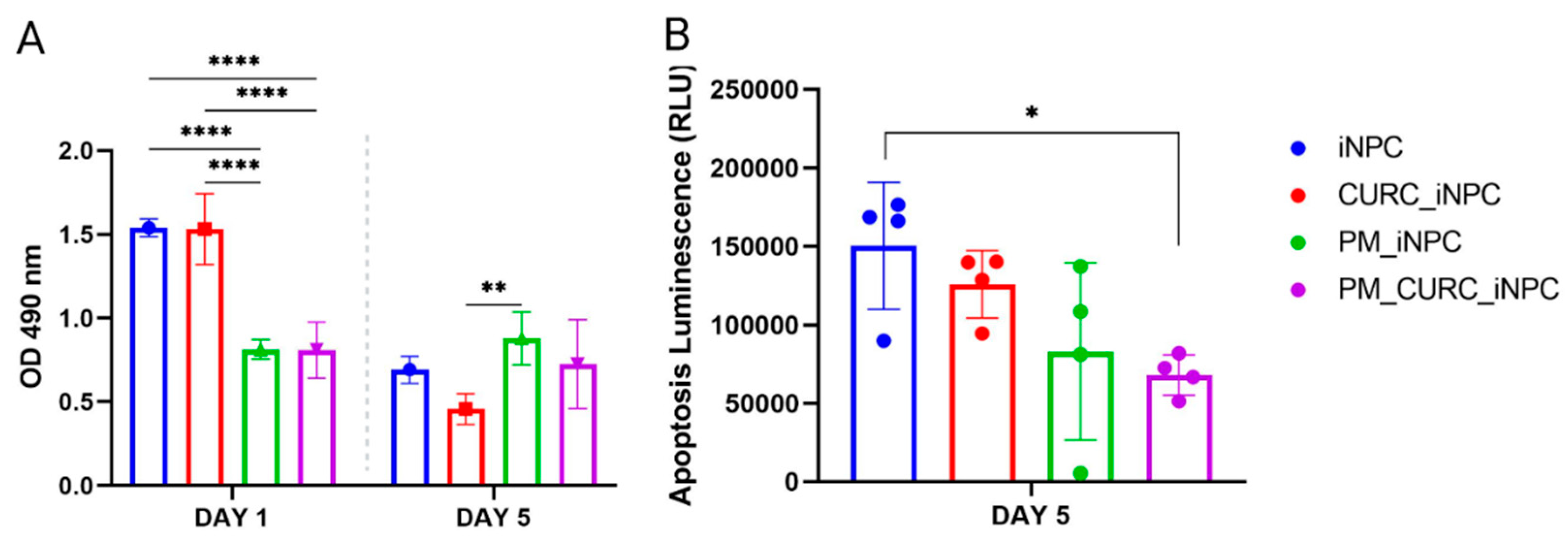
3.1. PuraMatrix-Embedded iNPCs Increase Long-Term Survival
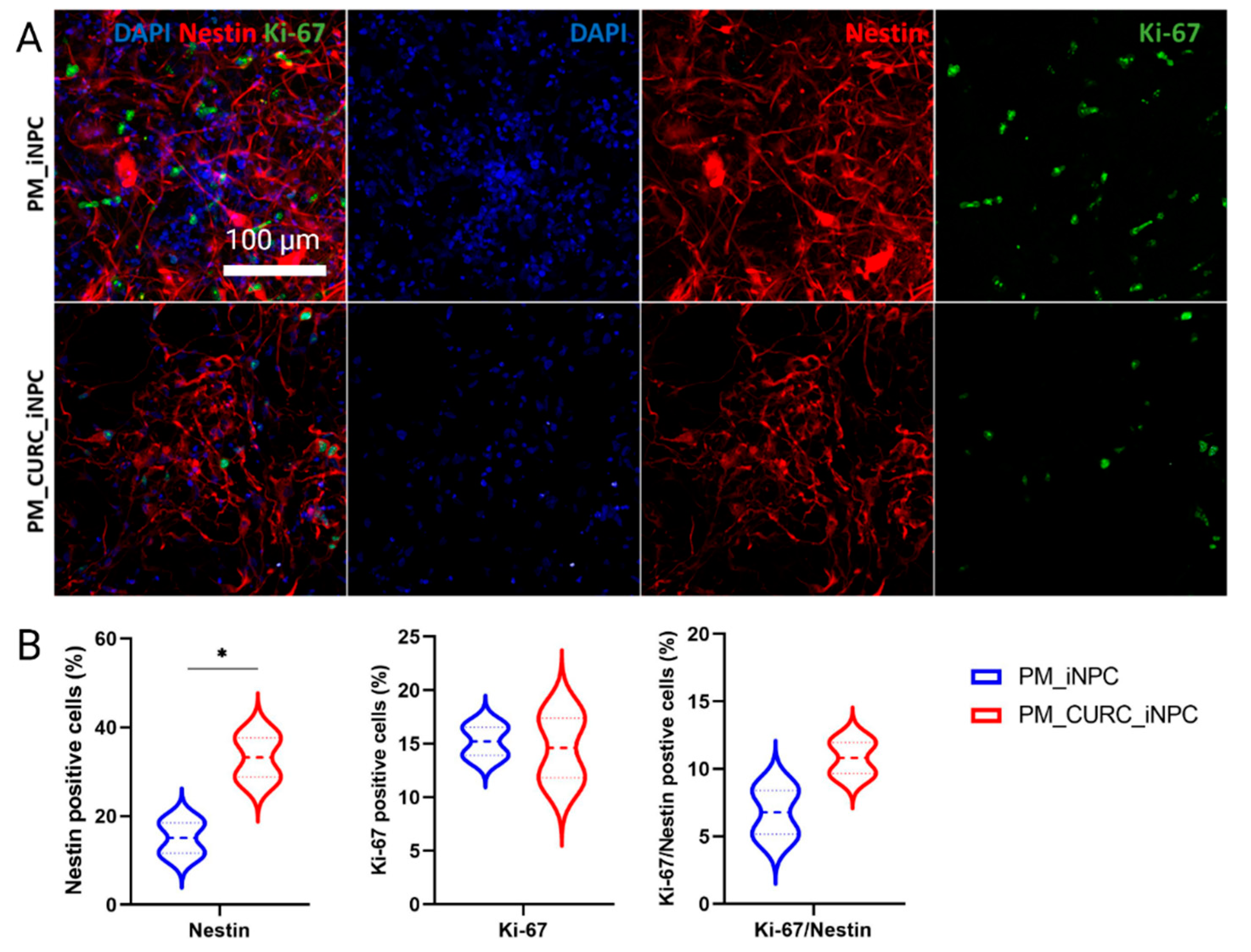
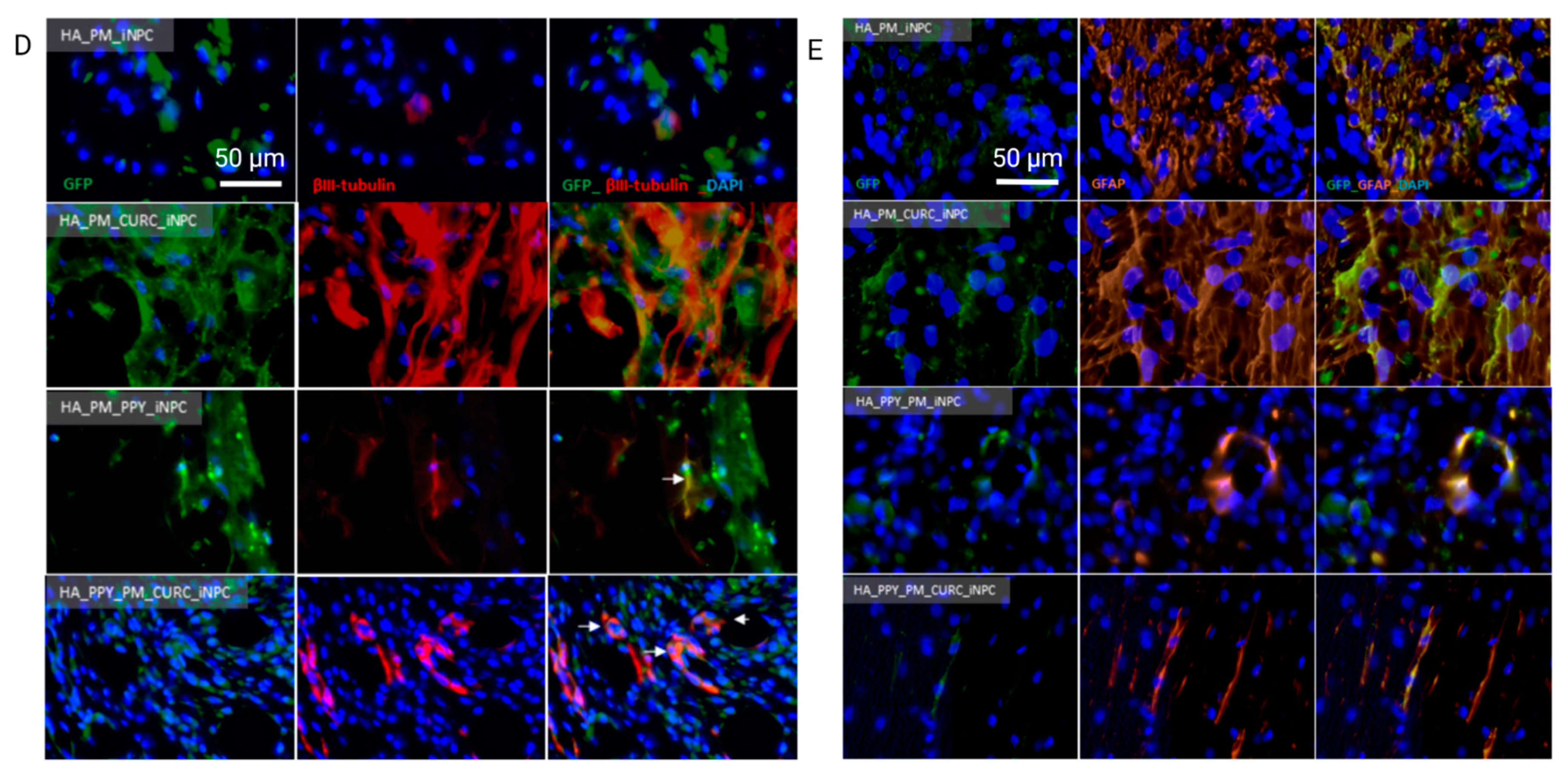
3.2. CURC Induces a Neurogenic-like Phenotype of PM-Embedded iNPCs
3.3. HA Demilune Scaffolds Containing PPY-Coated Fibers Guide PM-Embedded iNPCs for Spinal Cord Implantation
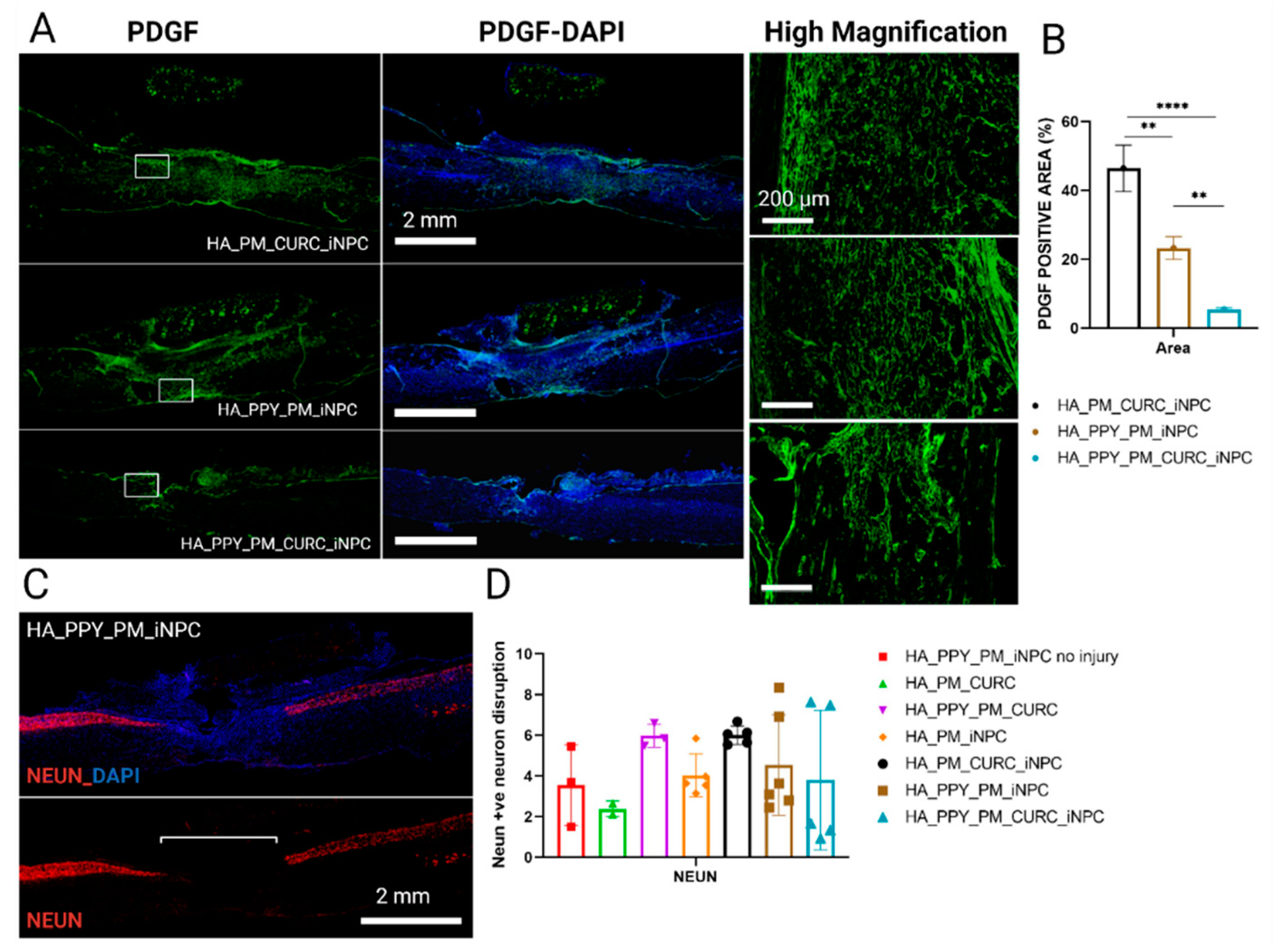
3.4. CURC Supports iNPC Migration within the Injured Spinal Cord
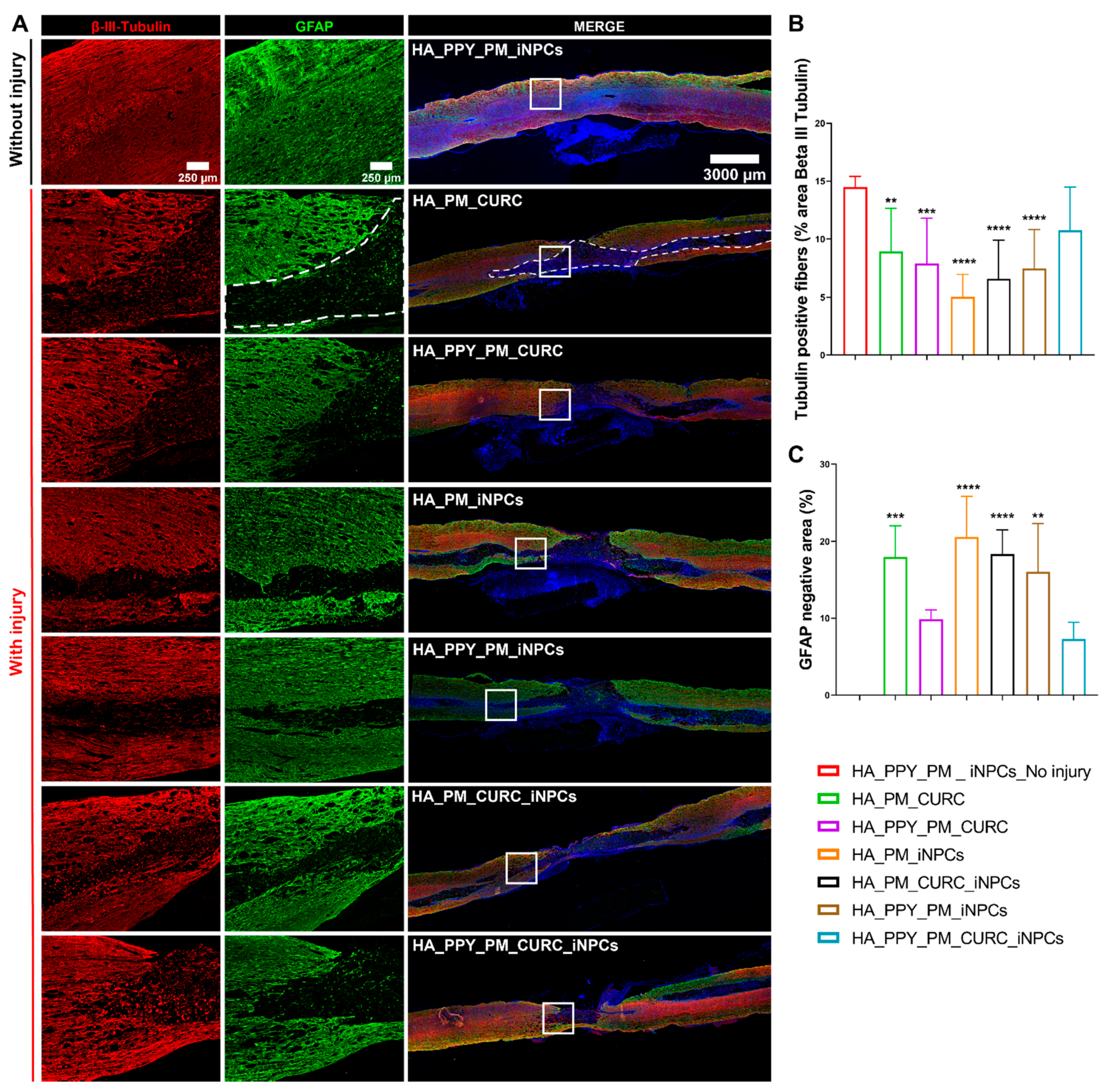
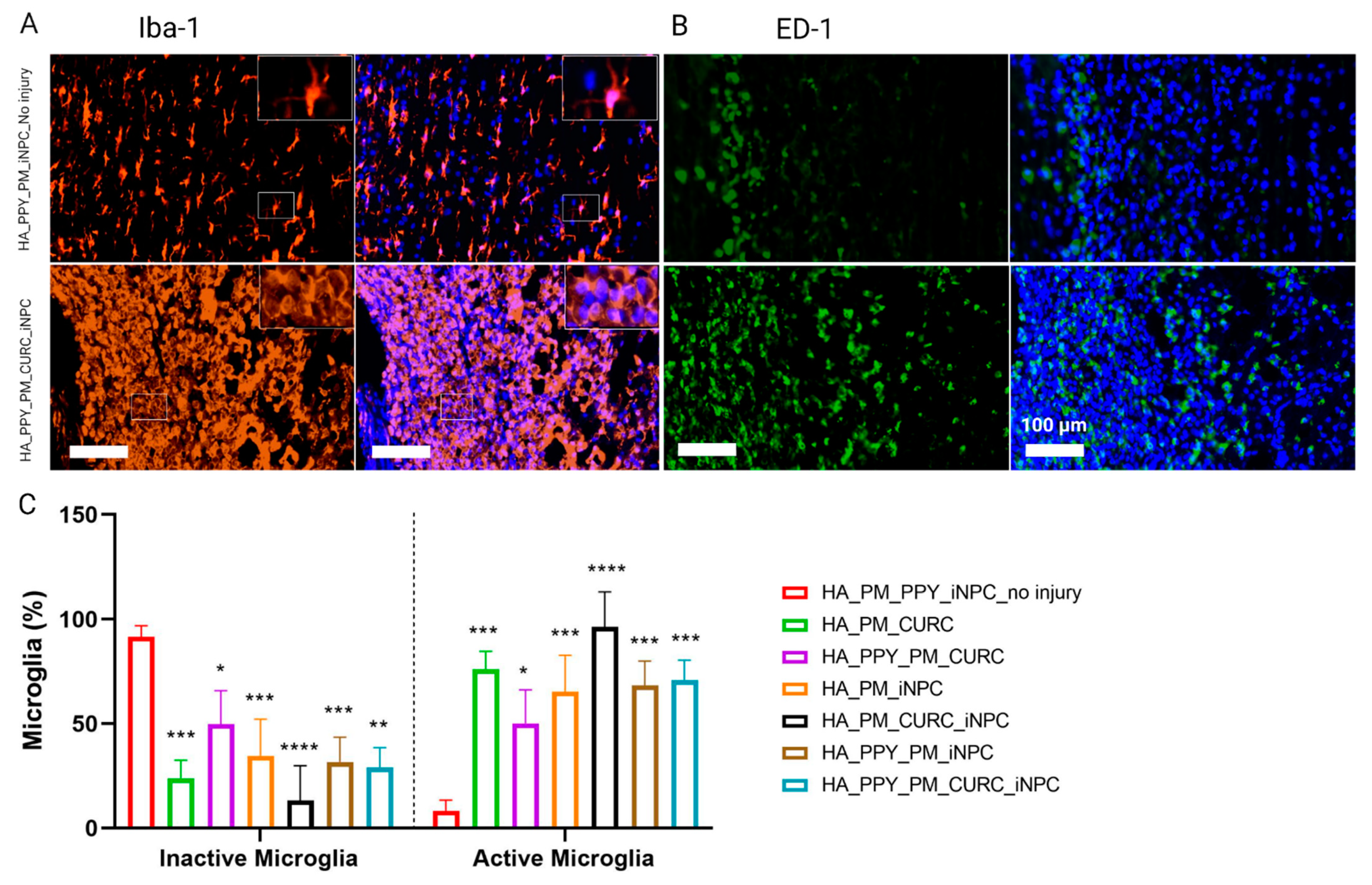
3.5. A Fully Functionalized HA Demilune Scaffold Preserves Neuronal Fibers and Reduces Glial Scarring but Does Not Reduce Neuroinflammation Early after SCI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ahuja, C.S.; Fehlings, M. Concise Review: Bridging the Gap: Novel Neuroregenerative and Neuroprotective Strategies in Spinal Cord Injury. Stem. Cells Transl. Med. 2016, 5, 914–924. [Google Scholar] [CrossRef]
- Kumamaru, H.; Kadoya, K.; Adler, A.F.; Takashima, Y.; Graham, L.; Coppola, G.; Tuszynski, M.H. Generation and post-injury integration of human spinal cord neural stem cells. Nat. Methods 2018, 15, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Assinck, P.; Duncan, G.J.; Hilton, B.J.; Plemel, J.R.; Tetzlaff, W. Cell transplantation therapy for spinal cord injury. Nat. Neurosci. 2017, 20, 637–647. [Google Scholar] [CrossRef]
- Kajikawa, K.; Imaizumi, K.; Shinozaki, M.; Shibata, S.; Shindo, T.; Kitagawa, T.; Shibata, R.; Kamata, Y.; Kojima, K.; Nagoshi, N.; et al. Cell therapy for spinal cord injury by using human iPSC-derived region-specific neural progenitor cells. Mol. Brain 2020, 13, 120. [Google Scholar] [CrossRef]
- Iwai, H.; Shimada, H.; Nishimura, S.; Kobayashi, Y.; Itakura, G.; Hori, K.; Hikishima, K.; Ebise, H.; Negishi, N.; Shibata, S.; et al. Allogeneic Neural Stem/Progenitor Cells Derived From Embryonic Stem Cells Promote Functional Recovery After Transplantation Into Injured Spinal Cord of Nonhuman Primates. Stem. Cells Transl. Med. 2015, 4, 708–719. [Google Scholar] [CrossRef] [PubMed]
- Tanimoto, Y.; Yamasaki, T.; Nagoshi, N.; Nishiyama, Y.; Nori, S.; Nishimura, S.; Iida, T.; Ozaki, M.; Tsuji, O.; Ji, B.; et al. In vivo monitoring of remnant undifferentiated neural cells following human induced pluripotent stem cell-derived neural stem/progenitor cells transplantation. Stem. Cells Transl. Med. 2020, 9, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.; Kalalinia, F. Application of encapsulation technology in stem cell therapy. Life Sci. 2015, 143, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Gelain, F.; Bottai, D.; Vescovi, A.; Zhang, S. Designer self-assembling peptide nanofiber scaffolds for adult mouse neural stem cell 3-dimensional cultures. PLoS ONE 2006, 1, e119. [Google Scholar] [CrossRef]
- Ortinau, S.; Schmich, J.; Block, S.; Liedmann, A.; Jonas, L.; Weiss, D.G.; Helm, C.A.; Rolfs, A.; Frech, M.J. Effect of 3D-scaffold formation on differentiation and survival in human neural progenitor cells. Biomed. Eng. Online 2010, 9, 70. [Google Scholar] [CrossRef]
- Thonhoff, J.R.; Lou, D.I.; Jordan, P.M.; Zhao, X.; Wu, P. Compatibility of human fetal neural stem cells with hydrogel biomaterials in vitro. Brain Res. 2008, 1187, 42–51. [Google Scholar] [CrossRef]
- Mahoney, M.J.; Anseth, K.S. Three-dimensional growth and function of neural tissue in degradable polyethylene glycol hydrogels. Biomaterials 2006, 27, 2265–2274. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, W.; Wu, X.; Zhang, N.; Zhang, Y.; Ouyang, S.; Song, X.; Fang, X.; Seeram, R.; Xue, W. Functional self-assembling peptide nanofiber hydrogels designed for nerve degeneration. ACS Appl. Mater. Interfaces 2016, 8, 2348–2359. [Google Scholar] [CrossRef] [PubMed]
- Abdolahi, S.; Aligholi, H.; Khodakaram-Tafti, A.; Ghadiri, M.K.; Stummer, W.; Gorji, A. Improvement of Rat Spinal Cord Injury Following Lentiviral Vector-Transduced Neural Stem/Progenitor Cells Derived from Human Epileptic Brain Tissue Transplantation with a Self-assembling Peptide Scaffold. Mol. Neurobiol. 2021, 58, 2481–2493. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, P.; Hernandez, J.; Giraldo, E.; González-Pérez, M.A.; Alastrue-Agudo, A.; Elkhenany, H.; Vicent, M.J.; Navarro, X.; Edel, M.; Moreno-Manzano, V. Human-Induced Neural and Mesenchymal Stem Cell Therapy Combined with a Curcumin Nanoconjugate as a Spinal Cord Injury Treatment. Int. J. Mol. Sci. 2021, 22, 5966. [Google Scholar] [CrossRef]
- Mortazavi, M.M.; Jaber, M.; Adeeb, N.; Deep, A.; Hose, N.; Rezaei, M.; Fard, S.A.; Kateb, B.; Yashar, P.; Liker, M.A.; et al. Engraftment of neural stem cells in the treatment of spinal cord injury. Transl. Res. Anat. 2015, 1, 11–16. [Google Scholar] [CrossRef][Green Version]
- Bakshi, A.; Barshinger, A.L.; Swanger, S.A.; Madhvani, V.; Shumsky, J.S.; Neuhuber, B.; Fischer, I. Lumbar puncture delivery of bone marrow stromal cells in spinal cord contusion: A novel method for minimally invasive cell transplantation. J. Neurotrauma 2006, 23, 55–65. [Google Scholar] [CrossRef]
- Martínez-Ramos, C.; Doblado, L.R.; Mocholi, E.L.; Alastrue-Agudo, A.; Petidier, M.S.; Giraldo, E.; Pradas, M.M.; Moreno-Manzano, V. Biohybrids for spinal cord injury repair. J. Tissue Eng. Regen. Med. 2019, 13, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Horn, E.M.; Beaumont, M.; Shu, X.Z.; Harvey, A.; Prestwich, G.D.; Horn, K.M.; Gibson, A.R.; Preul, M.C.; Panitch, A. Influence of cross-linked hyaluronic acid hydrogels on neurite outgrowth and recovery from spinal cord injury. J. Neurosurg. Spine 2007, 6, 133–140. [Google Scholar] [CrossRef]
- Kushchayev, S.V.; Giers, M.B.; Eng, D.H.; Martirosyan, N.L.; Eschbacher, J.M.; Mortazavi, M.M.; Theodore, N.; Panitch, A.; Preul, M.C. Hyaluronic acid scaffold has a neuroprotective effect in hemisection spinal cord injury. J. Neurosurg. Spine 2016, 25, 114–124. [Google Scholar] [CrossRef]
- Wen, Y.; Yu, S.; Wu, Y.; Ju, R.; Wang, H.; Liu, Y.; Wang, Y.; Xu, Q. Spinal cord injury repair by implantation of structured hyaluronic acid scaffold with PLGA microspheres in the rat. Cell Tissue Res. 2016, 364, 17–28. [Google Scholar] [CrossRef]
- Doblado, L.R.; Martínez-Ramos, C.; García-Verdugo, J.M.; Moreno-Manzano, V.; Pradas, M.M. Engineered axon tracts within tubular biohybrid scaffolds. J. Neural Eng. 2021, 18, 0460c0465. [Google Scholar] [CrossRef]
- Stupack, D.G.; Cheresh, D.A. Get a ligand, get a life: Integrins, signaling and cell survival. J. Cell Sci. 2002, 115, 3729–3738. [Google Scholar] [CrossRef] [PubMed]
- Smeal, R.M.; Rabbitt, R.; Biran, R.; Tresco, P.A. Substrate curvature influences the direction of nerve outgrowth. Ann. Biomed. Eng. 2005, 33, 376–382. [Google Scholar] [CrossRef]
- Wang, X.; Gu, X.; Yuan, C.; Chen, S.; Zhang, P.; Zhang, T.; Yao, J.; Chen, F.; Chen, G. Evaluation of biocompatibility of polypyrrole in vitro and in vivo. J. Biomed. Mater. Res. Part A 2004, 68, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-X.; Li, X.-G.; Yang, Y.-L. Preparation, properties and applications of polypyrroles. React. Funct. Polym. 2001, 47, 125–139. [Google Scholar] [CrossRef]
- Sadki, S.; Schottland, P.; Brodie, N.; Sabouraud, G. The mechanisms of pyrrole electropolymerization. Chem. Soc. Rev. 2000, 29, 283–293. [Google Scholar]
- George, P.M.; Lyckman, A.W.; LaVan, D.A.; Hegde, A.; Leung, Y.; Avasare, R.; Testa, C.; Alexander, P.M.; Langer, R.; Sur, M. Fabrication and biocompatibility of polypyrrole implants suitable for neural prosthetics. Biomaterials 2005, 26, 3511–3519. [Google Scholar] [CrossRef]
- Alvarez-Mejia, L.; Morales, J.; Cruz, G.J.; Olayo, M.-G.; Olayo, R.; Díaz-Ruíz, A.; Ríos, C.; Mondragón-Lozano, R.; Sánchez-Torres, S.; Morales-Guadarrama, A. Functional recovery in spinal cord injured rats using polypyrrole/iodine implants and treadmill training. J. Mater. Sci. Mater. Med. 2015, 26, 1–11. [Google Scholar] [CrossRef]
- Shu, B.; Liu, X.B.; Zhou, J.F.; Huang, H.; Wang, J.Y.; Sun, X.D.; Qin, C.; An, Y.H. Polypyrrole/polylactic acid nanofibrous scaffold cotransplanted with bone marrow stromal cells promotes the functional recovery of spinal cord injury in rats. CNS Neurosci. Ther. 2019, 25, 951–964. [Google Scholar]
- Roca, F.G.; Estellés, J.M.; Pradas, M.M.; Martínez-Ramos, C. Axonal extension from dorsal root ganglia on fibrillar and highly aligned poly (lactic acid)-polypyrrole substrates obtained by two different techniques: Electrospun nanofibres and extruded microfibres. Int. J. Biol. Macromol. 2020, 163, 1959–1969. [Google Scholar] [CrossRef] [PubMed]
- Gisbert Roca, F.; García-Bernabé, A.; Compañ Moreno, V.; Martínez-Ramos, C.; Monleón Pradas, M. Solid Polymer Electrolytes Based on Polylactic Acid Nanofiber Mats Coated with Polypyrrole. Macromol. Mater. Eng. 2021, 306, 2000584. [Google Scholar] [CrossRef]
- Ramot, Y.; Haim-Zada, M.; Domb, A.J.; Nyska, A. Biocompatibility and safety of PLA and its copolymers. Adv. Drug Deliv. Rev. 2016, 107, 153–162. [Google Scholar] [CrossRef]
- da Silva, D.; Kaduri, M.; Poley, M.; Adir, O.; Krinsky, N.; Shainsky-Roitman, J.; Schroeder, A. Biocompatibility, biodegradation and excretion of polylactic acid (PLA) in medical implants and theranostic systems. Chem. Eng. J. 2018, 340, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Palomo, A.B.; Requena-Osete, J.; Delgado-Morales, R.; Moreno-Manzano, V.; Grau-Bove, C.; Tejera, A.M.; Otero, M.J.; Barrot, C.; Santos-Barriopedro, I.; Vaquero, A. A synthetic mRNA cell reprogramming method using CYCLIN D1 promotes DNA repair generating improved genetically stable human induced pluripotent stem cells. Stem. Cells 2021, 39, 866–881. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, E.; Nebot, V.J.; Đorđević, S.; Requejo-Aguilar, R.; Alastrue-Agudo, A.; Zagorodko, O.; Armiñan, A.; Martinez-Rojas, B.; Vicent, M.J.; Moreno-Manzano, V. A rationally designed self-immolative linker enhances the synergism between a polymer-rock inhibitor conjugate and neural progenitor cells in the treatment of spinal cord injury. Biomaterials 2021, 276, 121052. [Google Scholar] [CrossRef]
- Gisbert Roca, F.; André, F.M.; Más Estellés, J.; Monleón Pradas, M.; Mir, L.M.; Martínez-Ramos, C. BDNF-Gene Transfected Schwann Cell-Assisted Axonal Extension and Sprouting on New PLA–PPy Microfiber Substrates. Macromol. Biosci. 2021, 21, 2000391. [Google Scholar] [CrossRef]
- Alastrue-Agudo, A.; Erceg, S.; Cases-Villar, M.; Bisbal-Velasco, V.; Griffeth, R.J.; Rodriguez-Jiménez, F.J.; Moreno-Manzano, V. Experimental cell transplantation for traumatic spinal cord injury regeneration: Intramedullar or intrathecal administration. In Stem Cells and Tissue Repair; Kioussi, C., Ed.; Springer: New York, NY, USA, 2014; pp. 23–35. [Google Scholar]
- Fan, Y.; Chen, Z.; Pathak, J.L.; Carneiro, A.M.D.; Chung, C.Y. Differential Regulation of Adhesion and Phagocytosis of Resting and Activated Microglia by Dopamine. Front. Cell Neurosci. 2018, 12, 309. [Google Scholar] [CrossRef]
- Bao, F.; Chen, Y.; Dekaban, G.A.; Weaver, L.C. Early anti-inflammatory treatment reduces lipid peroxidation and protein nitration after spinal cord injury in rats. J. Neurochem. 2004, 88, 1335–1344. [Google Scholar] [CrossRef]
- Riew, T.-R.; Choi, J.-H.; Kim, H.L.; Jin, X.; Lee, M.-Y. PDGFR-β-Positive Perivascular Adventitial Cells Expressing Nestin Contribute to Fibrotic Scar Formation in the Striatum of 3-NP Intoxicated Rats. Front. Mol. Neurosci. 2018, 11, 402. [Google Scholar] [CrossRef]
- Requejo-Aguilar, R.; Alastrue-Agudo, A.; Cases-Villar, M.; Lopez-Mocholi, E.; England, R.; Vicent, M.J.; Moreno-Manzano, V. Combined polymer-curcumin conjugate and ependymal progenitor/stem cell treatment enhances spinal cord injury functional recovery. Biomaterials 2017, 113, 18–30. [Google Scholar] [CrossRef]
- Huang, C.; Hu, Z.l.; Wu, W.N.; Yu, D.F.; Xiong, Q.J.; Song, J.R.; Shu, Q.; Fu, H.; Wang, F.; Chen, J.G. Existence and distinction of acid-evoked currents in rat astrocytes. Glia 2010, 58, 1415–1424. [Google Scholar] [CrossRef]
- Wang, Y.-F.; Zu, J.-N.; Li, J.; Chen, C.; Xi, C.-Y.; Yan, J.-L. Curcumin promotes the spinal cord repair via inhibition of glial scar formation and inflammation. Neurosci. Lett. 2014, 560, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Liu, W.; Zhu, H.; Chen, Y.; Zhang, X.; Li, L.; Chu, W.; Wen, Z.; Feng, H.; Lin, J. Curcumin inhibits glial scar formation by suppressing astrocyte-induced inflammation and fibrosis in vitro and in vivo. Brain Res. 2017, 1655, 90–103. [Google Scholar] [CrossRef]
- Attari, F.; Ghadiri, T.; Hashemi, M. Combination of curcumin with autologous transplantation of adult neural stem/progenitor cells leads to more efficient repair of damaged cerebral tissue of rat. Exp. Physiol. 2020, 105, 1610–1622. [Google Scholar]
- Jahanbazi Jahan-Abad, A.; Sahab Negah, S.; Hosseini Ravandi, H.; Ghasemi, S.; Borhani-Haghighi, M.; Stummer, W.; Gorji, A.; Khaleghi Ghadiri, M. Human Neural Stem/Progenitor Cells Derived From Epileptic Human Brain in a Self-Assembling Peptide Nanoscaffold Improve Traumatic Brain Injury in Rats. Mol. Neurobiol. 2018, 55, 9122–9138. [Google Scholar] [CrossRef] [PubMed]
- Demirel, C.; Turkoz, D.; Yazicioglu, I.M.; Cokluk, C. The Preventive Effect of Curcumin on the Experimental Rat Epidural Fibrosis Model. World Neurosurg. 2021, 145, e141–e148. [Google Scholar] [CrossRef]
- Guo, S.; Meng, X.-W.; Yang, X.-S.; Liu, X.-F.; Ou-Yang, C.-H.; Liu, C. Curcumin administration suppresses collagen synthesis in the hearts of rats with experimental diabetes. Acta Pharmacol. Sin. 2018, 39, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; Gangireddy, S.R.; Narala, V.R.; Hogaboam, C.M.; Standiford, T.J.; Christensen, P.J.; Kondapi, A.K.; Reddy, R.C. Curcumin inhibits fibrosis-related effects in IPF fibroblasts and in mice following bleomycin-induced lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 298, L616–L625. [Google Scholar] [CrossRef]
- Dias, D.O.; Kim, H.; Holl, D.; Solnestam, B.W.; Lundeberg, J.; Carlén, M.; Göritz, C.; Frisén, J. Reducing pericyte-derived scarring promotes recovery after spinal cord injury. Cell 2018, 173, 153–165. [Google Scholar] [CrossRef]
- Birbrair, A.; Zhang, T.; Files, D.C.; Mannava, S.; Smith, T.; Wang, Z.-M.; Messi, M.L.; Mintz, A.; Delbono, O. Type-1 pericytes accumulate after tissue injury and produce collagen in an organ-dependent manner. Stem Cell Res. Ther. 2014, 5, 122. [Google Scholar] [CrossRef] [PubMed]
- Iwayama, T.; Olson, L.E. Involvement of PDGF in fibrosis and scleroderma: Recent insights from animal models and potential therapeutic opportunities. Curr. Rheumatol. Rep. 2013, 15, 304. [Google Scholar] [CrossRef] [PubMed]
- Heebkaew, N.; Rujanapun, N.; Kunhorm, P.; Jaroonwitchawan, T.; Chaicharoenaudomrung, N.; Promjantuek, W.; Noisa, P. Curcumin Induces Neural Differentiation of Human Pluripotent Embryonal Carcinoma Cells through the Activation of Autophagy. BioMed. Res. Int. 2019, 2019, 4378710. [Google Scholar] [CrossRef]
- Lu, P.; Woodruff, G.; Wang, Y.; Graham, L.; Hunt, M.; Wu, D.; Boehle, E.; Ahmad, R.; Poplawski, G.; Brock, J.; et al. Long-distance axonal growth from human induced pluripotent stem cells after spinal cord injury. Neuron 2014, 83, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yang, A.; Huang, Z.; Yin, G.; Pu, X.; Jin, J. Enhancement of neurite adhesion, alignment and elongation on conductive polypyrrole-poly(lactide acid) fibers with cell-derived extracellular matrix. Colloids Surf. B Biointerfaces 2017, 149, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-W.; Choi, C.-H.; Park, J.P.; Lee, S.-J. Nanospheres loaded with curcumin improve the bioactivity of umbilical cord BLOOD-mesenchymal stem cells via c-Src activation during the skin wound healing process. Cells 2020, 9, 1467. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Induction of SCI | Treatment |
|---|---|---|
| HA_PM_iNPC (non-injured) | No | Hyaluronic acid scaffold with PM-embedded iNPCs |
| HA_PPY_PM_iNPC (non-injured) | No | Hyaluronic acid scaffold with PPY fibers and PM-embedded iNPCs |
| HA_PM_CURC | Yes | Hyaluronic acid scaffold and PM-embedded CURC |
| HA_PPY_PM_CURC | Yes | Hyaluronic acid scaffold with PPY fibers and PM-embedded CURC |
| HA_PM_iNPC | Yes | Hyaluronic acid scaffold with PM-embedded iNPCs |
| HA_PPY_PM_iNPC | Yes | Hyaluronic acid scaffold with PPY fibers and PM-embedded iNPCs |
| HA_PM_CURC_iNPC | Yes | Hyaluronic acid scaffold with PM-embedded iNPCs and CURC |
| HA_PPY_PM_CURC_iNPC | Yes | Hyaluronic acid scaffold with PPY fibers, PM-embedded iNPCs, and CURC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elkhenany, H.; Bonilla, P.; Giraldo, E.; Alastrue Agudo, A.; Edel, M.J.; Vicent, M.J.; Roca, F.G.; Ramos, C.M.; Doblado, L.R.; Pradas, M.M.; et al. A Hyaluronic Acid Demilune Scaffold and Polypyrrole-Coated Fibers Carrying Embedded Human Neural Precursor Cells and Curcumin for Surface Capping of Spinal Cord Injuries. Biomedicines 2021, 9, 1928. https://doi.org/10.3390/biomedicines9121928
Elkhenany H, Bonilla P, Giraldo E, Alastrue Agudo A, Edel MJ, Vicent MJ, Roca FG, Ramos CM, Doblado LR, Pradas MM, et al. A Hyaluronic Acid Demilune Scaffold and Polypyrrole-Coated Fibers Carrying Embedded Human Neural Precursor Cells and Curcumin for Surface Capping of Spinal Cord Injuries. Biomedicines. 2021; 9(12):1928. https://doi.org/10.3390/biomedicines9121928
Chicago/Turabian StyleElkhenany, Hoda, Pablo Bonilla, Esther Giraldo, Ana Alastrue Agudo, Michael J. Edel, María Jesus Vicent, Fernando Gisbert Roca, Cristina Martínez Ramos, Laura Rodríguez Doblado, Manuel Monleón Pradas, and et al. 2021. "A Hyaluronic Acid Demilune Scaffold and Polypyrrole-Coated Fibers Carrying Embedded Human Neural Precursor Cells and Curcumin for Surface Capping of Spinal Cord Injuries" Biomedicines 9, no. 12: 1928. https://doi.org/10.3390/biomedicines9121928
APA StyleElkhenany, H., Bonilla, P., Giraldo, E., Alastrue Agudo, A., Edel, M. J., Vicent, M. J., Roca, F. G., Ramos, C. M., Doblado, L. R., Pradas, M. M., & Manzano, V. M. (2021). A Hyaluronic Acid Demilune Scaffold and Polypyrrole-Coated Fibers Carrying Embedded Human Neural Precursor Cells and Curcumin for Surface Capping of Spinal Cord Injuries. Biomedicines, 9(12), 1928. https://doi.org/10.3390/biomedicines9121928