
Effect Mechanism of Electroacupuncture at ST36 on the Injured Extensor Digitorum Longus in the Jumping Rat Model Based on mRNA-Seq Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Grouping
2.2. Animal Training and Acupuncture Methods
2.3. Sampling
2.4. Histomorphological Observation
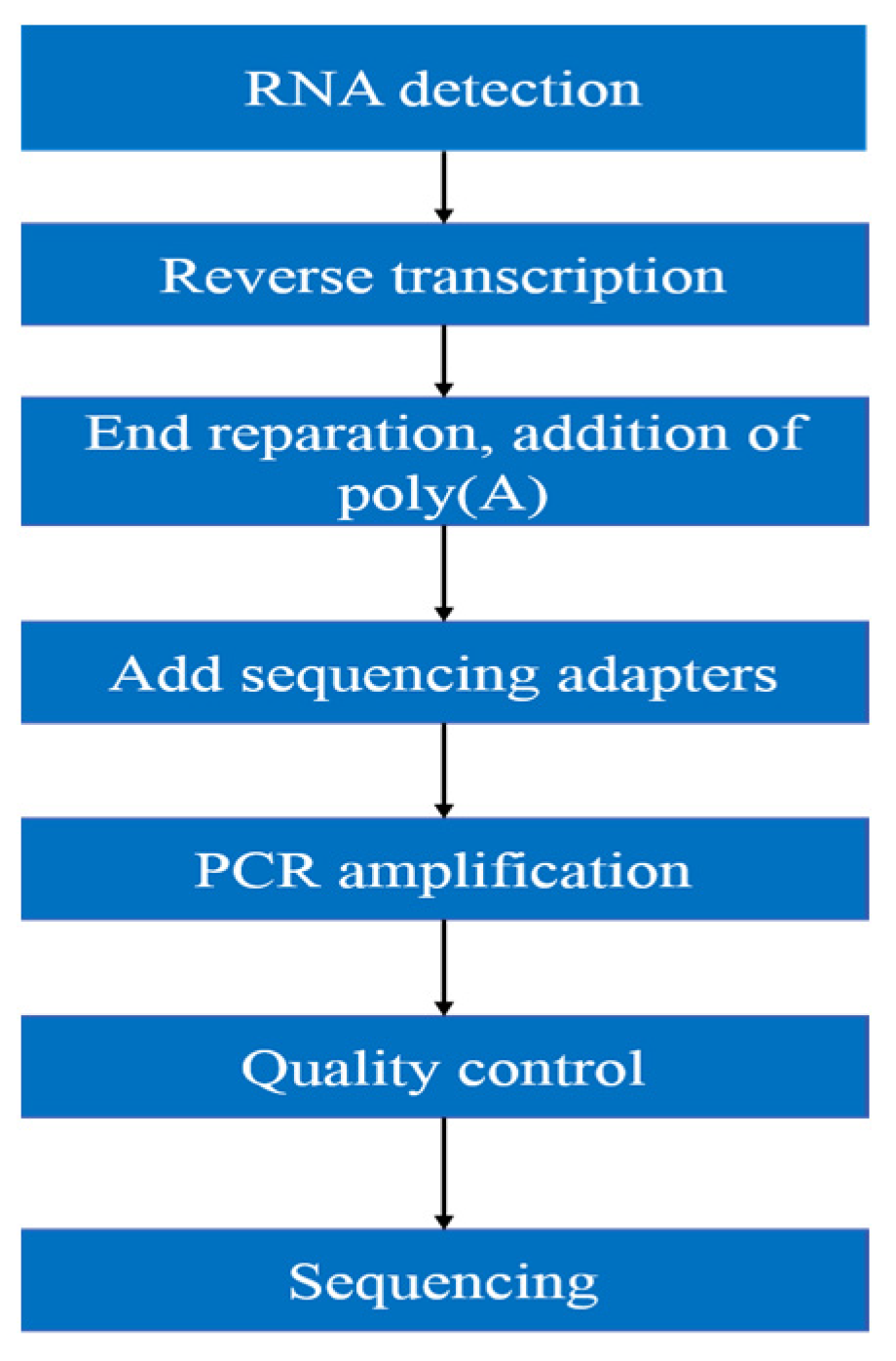
2.5. Transcriptomic Analysis
2.6. Western Blotting
2.7. Statistical Analysis
3. Results
3.1. Ultrastructural Analysis
3.2. Transcriptome Data Processing and Quality Control Analysis
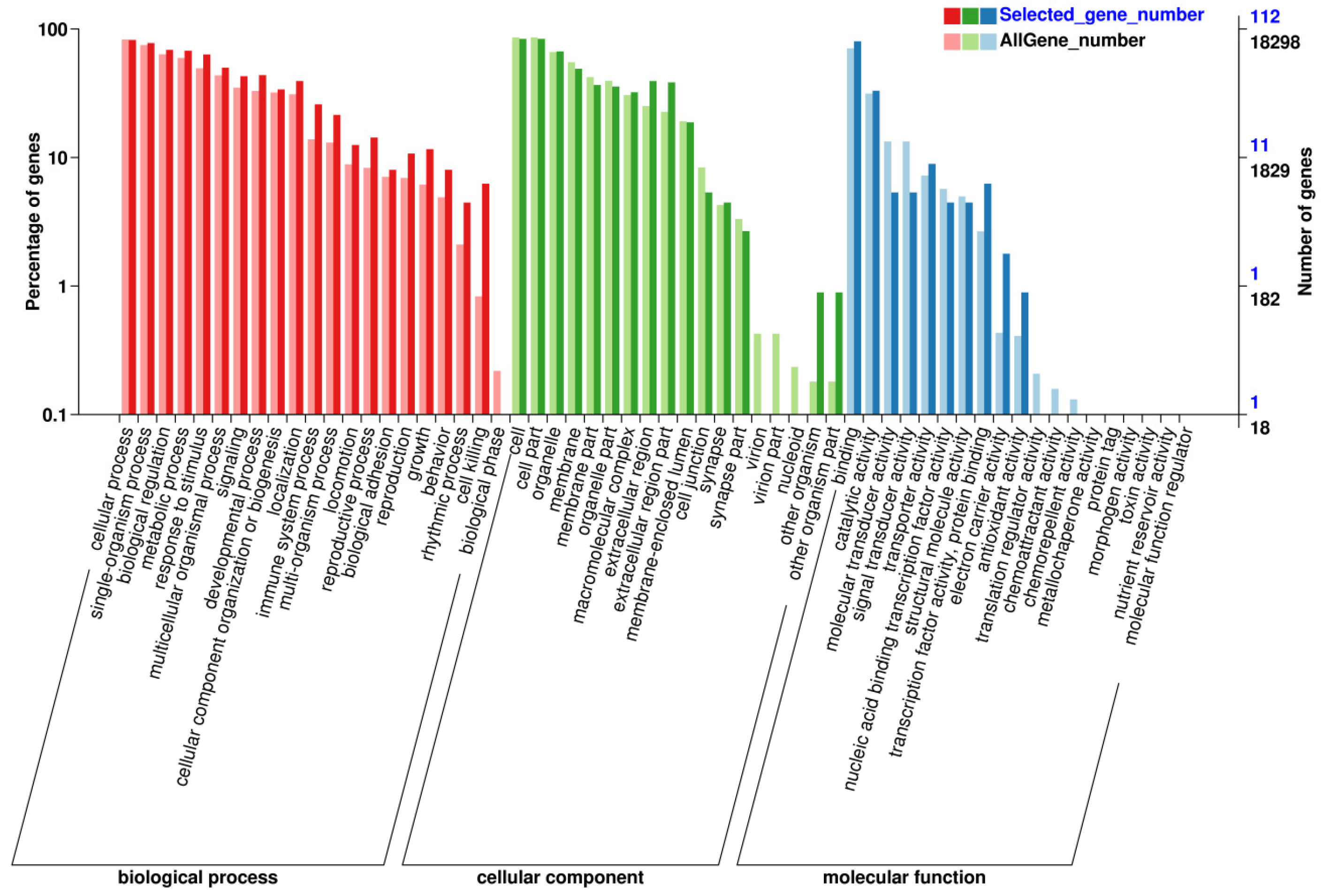
3.3. Screening Genes and Pathways through GO/KEGG
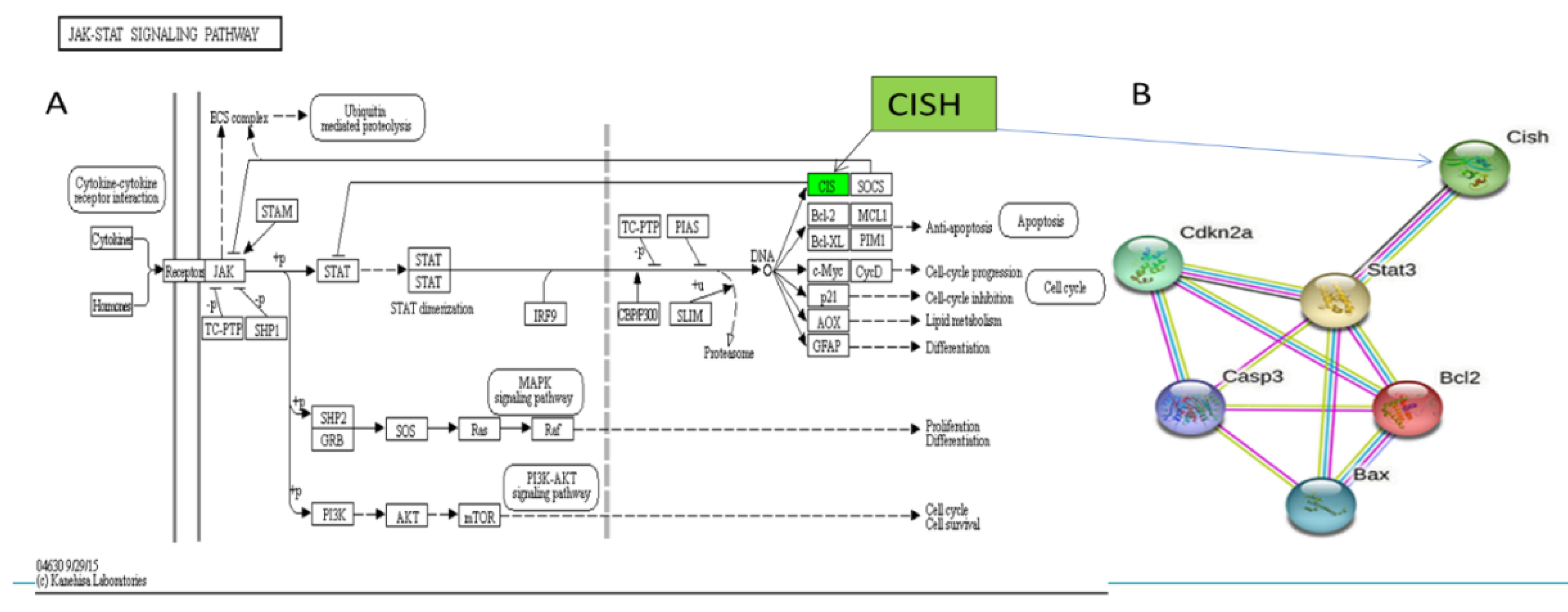
3.4. Screening Major Target Pathways and Key Target Genes
3.5. Changes in Key Target Proteins Expression from Each Group
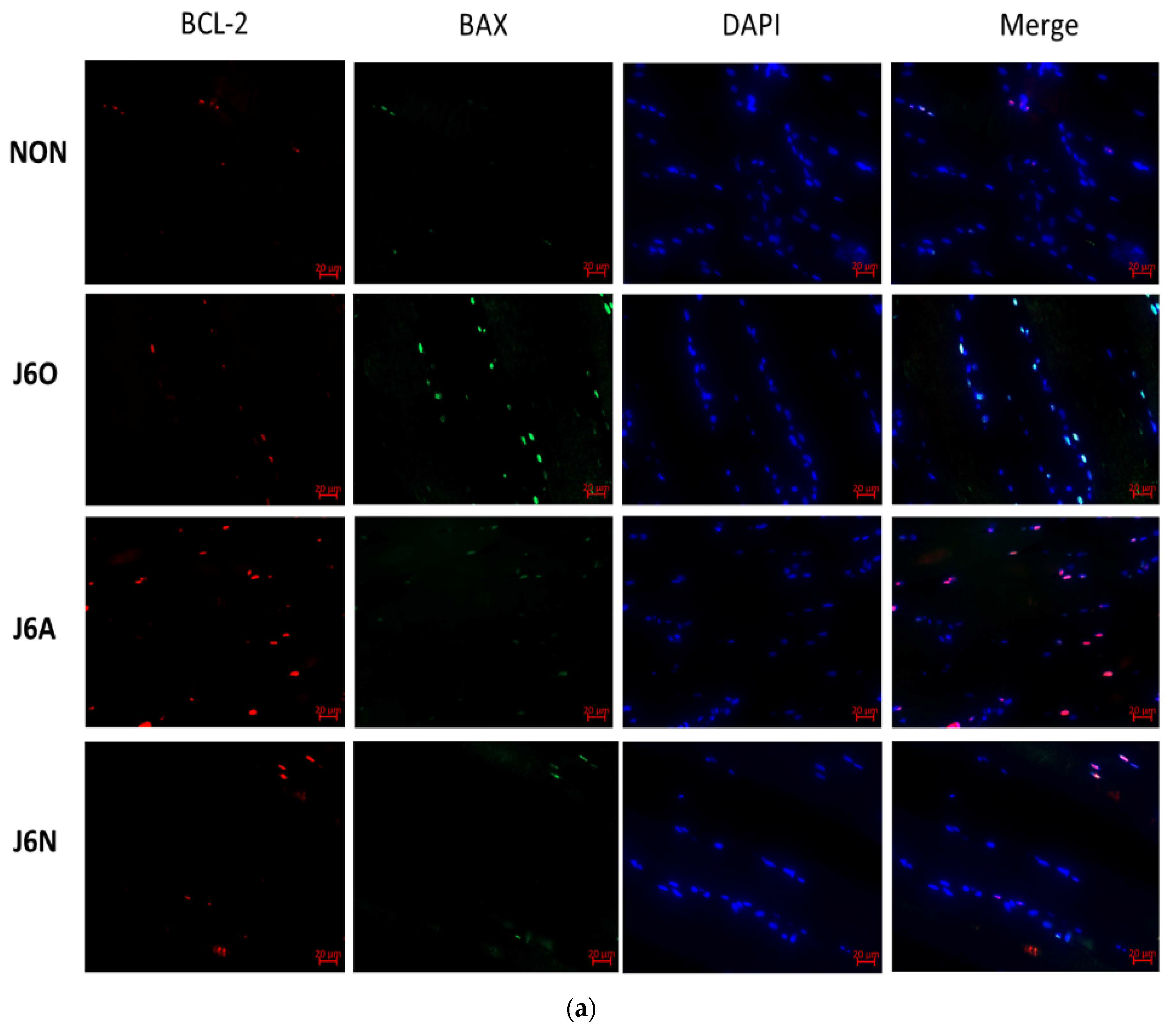
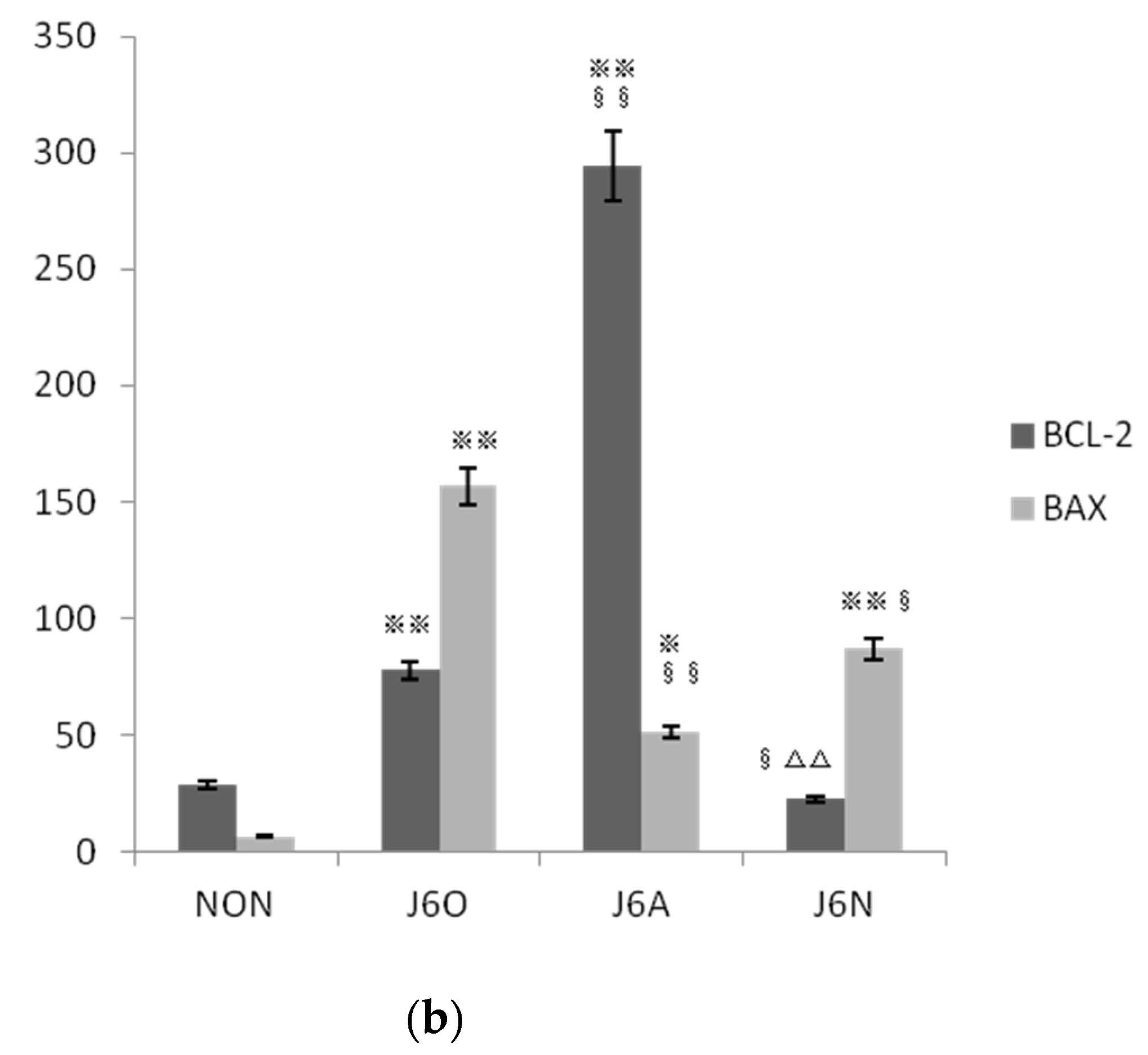
3.6. Immunofluorescence Detection of Changes in BCL-2/BAX Expression in Each Group
4. Discussion
4.1. Effects of Electroacupuncture at ST36 on the Morphological Structure of an Overused Extensor Digitorum Longus
4.2. Molecular Mechanism of Electroacupuncture Stimulation on the Repair of Damaged Muscle in Overused Jumping Rats
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Gent, R.N.; Siem, D.; van Middelkoop, M.; van Os, A.G.; Bierma-Zeinstra, S.M.; Koes, B.W. Incidence and determinants of lower extremity running injuries in long distance runners: A systematic review. Br. J. Sports Med. 2007, 41, 469–480. [Google Scholar] [CrossRef] [Green Version]
- Mizrahi, J.; Verbitsky, O.; Isakov, E. Fatigue-related loading imbalance on the shank in running: A possible factor in stress fractures. Ann. Biomed. Eng. 2000, 28, 463–469. [Google Scholar] [CrossRef]
- Ishida, T.; Yamanaka, M.; Takeda, N.; Homan, K.; Koshino, Y.; Kobayashi, T.; Matsumoto, H.; Aoki, Y. The effect of changing toe direction on knee kinematics during drop vertical jump: A possible risk factor for anterior cruciate ligament injury. Knee Surg. Sports Traumatol. Arthrosc. 2015, 23, 1004–1009. [Google Scholar] [CrossRef] [PubMed]
- Lezak, B.; Summers, S. Anatomy, Bony Pelvis and Lower Limb, Extensor Hallucis Longus Muscle. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Brazier, J.; Maloney, S.; Bishop, C.; Read, P.J.; Turner, A.N. Lower Extremity Stiffness: Considerations for Testing, Performance Enhancement, and Injury Risk. J. Strength Cond. Res. 2019, 33, 1156–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobara, H.; Inoue, K.; Muraoka, T.; Omuro, K.; Sakamoto, M.; Kanosue, K. Leg stiffness adjustment for a range of hopping frequencies in humans. J. Biomech. 2010, 43, 506–511. [Google Scholar]
- Maloney, S.J.; Richards, J.; Nixon, D.G.D.; Harvey, L.J.; Fletcher, I.M. Do stiffness and asymmetries predict change of direction performance? J. Sports Sci. 2017, 35, 547–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazevich, A. The stretch shortening cycle (SSC). In Strength and Conditioning Biological Principles and Practical Applications; Cardinale, M., Newton, R., Nosaka, K., Eds.; Wiley-Blackwell: West Sussex, UK, 2011; pp. 209–218. [Google Scholar]
- Litchwark, G.A.; Wilson, A.M. In vivo mechanical properties of the human achillies tendon during one legged hopping. J. Exp. Biol. 2005, 208, 4715–4725. [Google Scholar] [CrossRef] [Green Version]
- Best, T.M.; McElhaney, J.; Garrett, W.E.; Myers, B.S. Characterisation of the passive responses of live skeletal muscle using quasi-linear theory of viscoelasticity. J. Biomech. 1994, 27, 413–419. [Google Scholar] [CrossRef]
- Liu, H.; Gao, F.; Liang, X.; Chen, X.; Qu, Y.; Wang, L. Pathogenesis and Development of Patellar Tendon Fibrosis in a Rabbit Overuse Model. SAGA J. 2020, 19, 1141–1150. [Google Scholar] [CrossRef]
- Wei, J.A.; Hu, X.; Zhang, B.; Liu, L.; Chen, K.; So, K.F.; Li, M.; Zhang, L. Electroacupuncture activates inhibitory neural circuits in the somatosensory cortex to relieve neuropathic pain. iScience 2021, 24, 102066. [Google Scholar] [CrossRef]
- Casagrande, S.M.; de Lourdes Pessole Biondo-Simões, M.; Ioshii, S.; Robes, R.R.; Biondo-Simões, R.; de Oliveira Boeno , B.R. Histological evaluation of the effect of low-frequency electric stimulation on healing Achilles tendons in rats. Acta Cir. Bras. 2021, 36, e360103. [Google Scholar] [CrossRef]
- Benrick, A.; Pillon, N.J.; Nilsson, E.; Lindgren, E.; Krook, A.; Ling, C.; Stener-Victorin, E. Electroacupuncture Mimics Exercise-Induced Changes in Skeletal Muscle Gene Expression in Women with Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2020, 105, 2027–2041. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liu, Y.; Bian, J.; Li, Q.; Zhang, Y. The preemptive analgesia of pre-electroacupuncture in rats with formalin-induced acute inflammatory pain. Mol. Pain. 2019, 15, 1744806919866529. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Lao, L.; Ren, K.; Berman, B.M. Mechanisms of Acupuncture-Electroacupuncture on Persistent Pain. Anesthesiology 2014, 120, 482–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.H.; Li, C.W.; Wang, J.Y.; Kan, Y.; Tan, L.H.; Jing, X.H.; Liu, J.L. Activation of hippocampal MEK1 contributes to the cumulative antinociceptive effect of electroacupuncture in neuropathic pain rats. BMC Complement Altern. Med. 2016, 16, 517. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.Y.; Liang, S.; Yu, M.L.; Fu, S.P.; Chen, X.; Lu, S.F. A systematic review and meta-analysis of acupuncture for improving learning and memory ability in animals. BMC Complement Altern. Med. 2016, 16, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Li, S.; Wang, Y.; Lei, Y.; Foreman, R.D.; Yin, J.; Chen, J.D. Effects and mechanisms of auricular electroacupuncture on gastric hypersensitivity in a rodent model of functional dyspepsia. PLoS ONE 2017, 12, e0174568. [Google Scholar] [CrossRef]
- Protasi, F.; Pietrangelo, L.; Boncompagni, S. Calcium entry units (CEUs): Perspectives in skeletal muscle function and disease. J. Muscle Res. Cell Motil. 2021, 42, 233–249. [Google Scholar] [CrossRef]
- Matsumoto, A.; Masuhara, M.; Mitsui, K.; Yokouchi, M.; Ohtsubo, M.; Misawa, H. CIS, a cytokine inducible SH2 protein, is a target of the JAK-STAT5 pathway and modulates STAT5 activation. Blood 1997, 89, 3148–3154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.O.; Zhang, H.; Kim, B.S.; Niu, X.; Peng, J.; Chen, Y. The signaling suppressor CIS controls proallergic T cell development and allergic airway inflammation. Nat. Immunol. 2013, 14, 732–740. [Google Scholar] [CrossRef] [Green Version]
- Takeshima, H.; Horie, M.; Mikami, Y.; Makita, K.; Miyashita, N.; Matsuzaki, H. CISH is a negative regulator of IL-13-induced CCL26 production in lung fibroblasts. Allergol. Int. 2019, 68, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.Y.; Hsiao, J.R.; Chou, S.T.; Hsu, Y.M.; Wu, G.H.; Shieh, Y.S.; Shiah, S.G. MiR-944/CISH mediated inflammation via STAT3 is involved in oral cancer malignance by cigarette smoking. Neoplasia 2020, 22, 554–565. [Google Scholar] [CrossRef]
- Ahuja, A.; Kim, E.; Sung, G.H.; Cho, J.Y. STAT3 Differentially Regulates TLR4-Mediated Inflammatory Responses in Early or Late Phases. Int. J. Mol. Sci. 2020, 21, 7675. [Google Scholar] [CrossRef] [PubMed]
- Welte, T.; Zhang, S.S.; Wang, T.; Zhang, Z.; Hesslein, D.G.; Yin, Z.; Kano, A.; Iwamoto, Y.; Li, E.; Craft, J.E.; et al. STAT3 deletion during hematopoiesis causes Crohn’s disease-like pathogenesis and lethality: A critical role of STAT3 in innate immunity. Proc. Natl. Acad. Sci. USA 2003, 100, 1879–1884. [Google Scholar] [CrossRef] [Green Version]
- Linossi, E.M.; Nicholson, S.E. Kinase inhibition, competitive binding and proteasomal degradation: Resolving the molecular function of the suppressor of cytokine signaling (SOCS) proteins. Immunol Rev. 2015, 266, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, W.W.; Liu, P.; Yu, W.; Liu, T.; Yu, J. Dysregulation of SOCS-mediated negative feedback of cytokine signalling in carcinogenesis and its significance in cancer treatment. Front. Immunol. 2017, 8, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buettner, R.; Mora, L.B.; Jove, R. Activated STAT signaling in human tumors provides novel molecular targets for therapeutic intervention. Clin. Cancer Res. 2002, 8, 945–954. [Google Scholar]
- Liu, Y.; Liu, L.; Zhou, Y.; Zhou, P.; Yan, Q.; Chen, X.; Ding, S.; Zhu, F. CKLF1 Enhances Inflammation-Mediated Carcinogenesis and Prevents Doxorubicin-Induced Apoptosis via IL6/STAT3 Signaling in HCC. Clin. Cancer Res. 2019, 25, 4141–4154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laumonier, T.; Menetrey, J. Muscle injuries and strategies for improving their repair. J. Exp. Orthop. 2016, 3, 15. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Pardoll, D.; Jove, R. STATs in cancer inflammation and immunity: A leading role for STAT3. Nat. Rev. Cancer 2009, 9, 798–809. [Google Scholar] [CrossRef]
- Pan, Y.; Song, D.; Zhou, W.; Lu, X.; Wang, H.; Li, Z. Baicalin inhibits C2C12 myoblast apoptosis and prevents against skeletal muscle injury. Mol. Med. Rep. 2019, 20, 709–718. [Google Scholar] [CrossRef]
- Labi, V.; Erlacher, M. How cell death shapes cancer. Cell. Death Dis. 2015, 6, e1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.T.; Liu, J.P.; Wei, W.L. The effects of tanshinone IIA on hypoxia/reoxygenation-induced myocardial microvascular endothelial cell apoptosis in rats via the JAK2/STAT3 signaling pathway. Biomed. Pharmacother. 2016, 83, 1116–1126. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhan, X.; Shen, X.; Li, M.; Yang, J.; Yu, W.; Chen, H.; Jin, B.; Mao, Z. P16 promotes the growth and mobility potential of breast cancer both in vitro and in vivo: The key role of the activation of IL-6/JAK2/STAT3 signaling. Mol. Cell Biochem. 2018, 446, 137–148. [Google Scholar] [CrossRef]
- Medina, C.B.; Mehrotra, P.; Arandjelovic, S.; Perry, J.S.A.; Guo, Y.; Morioka, S.; Barron, B.; Walk, S.F.; Ghesquière, B.; Krupnick, A.S.; et al. Metabolites released from apoptotic cells act as tissue messengers. Nature 2020, 580, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Li, F.; Liu, X.; Li, W.; Shi, W.; Liu, F.F.; O’Sullivan, B.; He, Z.; Peng, Y.; Tan, A.C.; et al. Caspase 3-mediated stimulation of tumor cell repopulation during cancer radiotherapy. Nat. Med. 2011, 17, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Guadagnin, E.; Mázala, D.; Chen, Y.W. STAT3 in Skeletal Muscle Function and Disorders. Int. J. Mol. Sci. 2018, 19, 2265. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody Name | Molecular Weight (kDa) | Classification | Antibody Source | Catalog Number | Dilution Ratio |
|---|---|---|---|---|---|
| Caspase3 | 30 | apoptosis | Servicebio | GB11009 | 1:1000 |
| CDKN2α | 17 | cell cycle | Servicebio | GB13513 | 1:1000 |
| STAT3 | 88 | transcription factors | Servicebio | GB12176 | 1:1000 |
| CISH | 29 | inflammatory | BOSTER | BA3758 | 1:1000 |
| GAPDH | 37 | Internal reference | Servicebio | GB11002 | 1:1000 |
| Sample | Total Reads | Mapped Reads | GC(%) | Q20(%) | Q30(%) | Uniq Map | Multiple Map |
|---|---|---|---|---|---|---|---|
| NON1 | 46501974 (100%) | 44400196 (95.48%) | 49.7 | 98.09 | 94.58 | 41206966 (88.61%) | 3193230 (6.87%) |
| NON2 | 49633854 (100%) | 47481735 (95.66%) | 50.4 | 98.05 | 94.4 | 43979527 (88.61%) | 3502208 (7.06%) |
| NON3 | 43148380 (100%) | 41288426 (95.69%) | 50.01 | 98.31 | 95.02 | 38262260 (88.68%) | 3026166 (7.01%) |
| J6O1 | 46373644 (100%) | 44334550 (95.60%) | 50.76 | 98.19 | 94.78 | 40978261 (88.37%) | 3356289 (7.24%) |
| J6O2 | 51941888 (100%) | 49654816 (95.60%) | 50.48 | 98.23 | 94.81 | 45850954 (88.27%) | 3803862 (7.32%) |
| J6O3 | 58064958 (100%) | 55558156 (95.68%) | 50.66 | 98.1 | 94.55 | 51541420 (88.77%) | 4016736 (6.92%) |
| J6A1 | 40451452 (100%) | 38442027 (95.03%) | 49.4 | 98.22 | 94.95 | 35434601 (87.60%) | 3007426 (7.43%) |
| J6A2 | 41286370 (100%) | 39154025 (94.84%) | 49.79 | 98.03 | 94.52 | 36119262 (87.48%) | 3034763 (7.35%) |
| J6A3 | 44047722 (100%) | 42032385 (95.42%) | 49.5 | 98.3 | 95.01 | 38909299 (88.33%) | 3123086 (7.09%) |
| J6N1 | 45749406 (100%) | 43614333 (95.33%) | 50.3 | 97.95 | 94.17 | 40441682 (88.40%) | 3172651 (6.93%) |
| J6N2 | 59023458 (100%) | 56409967 (95.57%) | 50.55 | 98.09 | 94.54 | 52548642 (89.03%) | 3861325 (6.54%) |
| J6N3 | 54380728 (100%) | 52011006 (95.64%) | 50.48 | 98.13 | 94.68 | 48213108 (88.66%) | 3797898 (6.98%) |
| KEGG Signaling Pathway | Genes | Percentage | TOP GO Signaling Pathway | Annotated | Significant | Expected | KS |
|---|---|---|---|---|---|---|---|
| Endocytosis | 6 | 8.57% | endocytosis | 613 | 6 | 3.78 | 0.47256 |
| Phagosome | 8 | 11.43% | early phagosome | 7 | 0 | 0.04 | 0.75058 |
| Jak-STAT signaling pathway | 5 | 7.14% | negative regulation of JAK-STAT cascade | 50 | 1 | 0.31 | 0.0209 |
| JAK-STAT cascade | 166 | 3 | 1.02 | 0.02164 | |||
| regulation of JAK-STAT cascade | 147 | 3 | 0.91 | 0.07301 | |||
| positive regulation of JAK-STAT cascade | 76 | 2 | 0.47 | 0.15578 | |||
| JAK-STAT cascade involved in growth hormone signaling pathway | 7 | 0 | 0.04 | 0.18095 | |||
| Calcium signaling pathway | 5 | 7.14% | none | ||||
| Cell adhesion molecules CAMs | 8 | 11.43% | none | ||||
| Ribosome | 5 | 7.14% | regulation of ribosome biogenesis | 4 | 0 | 0.02 | 0.09929 |
| rescue of stalled ribosome | 2 | 0 | 0.01 | 0.26722 | |||
| negative regulation of ribosome biogenesis | 2 | 0 | 0.01 | 0.31515 | |||
| ribosome disassembly | 8 | 0 | 0.05 | 0.36302 | |||
| mature ribosome assembly | 6 | 0 | 0.04 | 0.56498 | |||
| ribosome localization | 17 | 0 | 0.1 | 0.62718 | |||
| establishment of ribosome localization | 17 | 0 | 0.1 | 0.62718 | |||
| assembly of large subunit precursor of preribosome | 4 | 0 | 0.02 | 0.71236 | |||
| ribosome assembly | 113 | 2 | 0.7 | 0.78913 | |||
| ribosome biogenesis | 417 | 6 | 2.57 | 0.91798 | |||
| 90S preribosome assembly | 11 | 0 | 0.07 | 0.99607 | |||
| HTLV-I infection | 10 | 14.29% | none | ||||
| PPAR signal pathway | 6 | 8.57% | none | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Guo, Y.; Han, R. Effect Mechanism of Electroacupuncture at ST36 on the Injured Extensor Digitorum Longus in the Jumping Rat Model Based on mRNA-Seq Analysis. Biomedicines 2021, 9, 1849. https://doi.org/10.3390/biomedicines9121849
Liu Q, Guo Y, Han R. Effect Mechanism of Electroacupuncture at ST36 on the Injured Extensor Digitorum Longus in the Jumping Rat Model Based on mRNA-Seq Analysis. Biomedicines. 2021; 9(12):1849. https://doi.org/10.3390/biomedicines9121849
Chicago/Turabian StyleLiu, Qianqian, Yanwei Guo, and Rui Han. 2021. "Effect Mechanism of Electroacupuncture at ST36 on the Injured Extensor Digitorum Longus in the Jumping Rat Model Based on mRNA-Seq Analysis" Biomedicines 9, no. 12: 1849. https://doi.org/10.3390/biomedicines9121849
APA StyleLiu, Q., Guo, Y., & Han, R. (2021). Effect Mechanism of Electroacupuncture at ST36 on the Injured Extensor Digitorum Longus in the Jumping Rat Model Based on mRNA-Seq Analysis. Biomedicines, 9(12), 1849. https://doi.org/10.3390/biomedicines9121849