
Keratinocyte and Fibroblast Wound Healing In Vitro Is Repressed by Non-Optimal Conditions but the Reparative Potential Can Be Improved by Water-Filtered Infrared A


Abstract
1. Introduction
2. Materials and Methods
3. Results
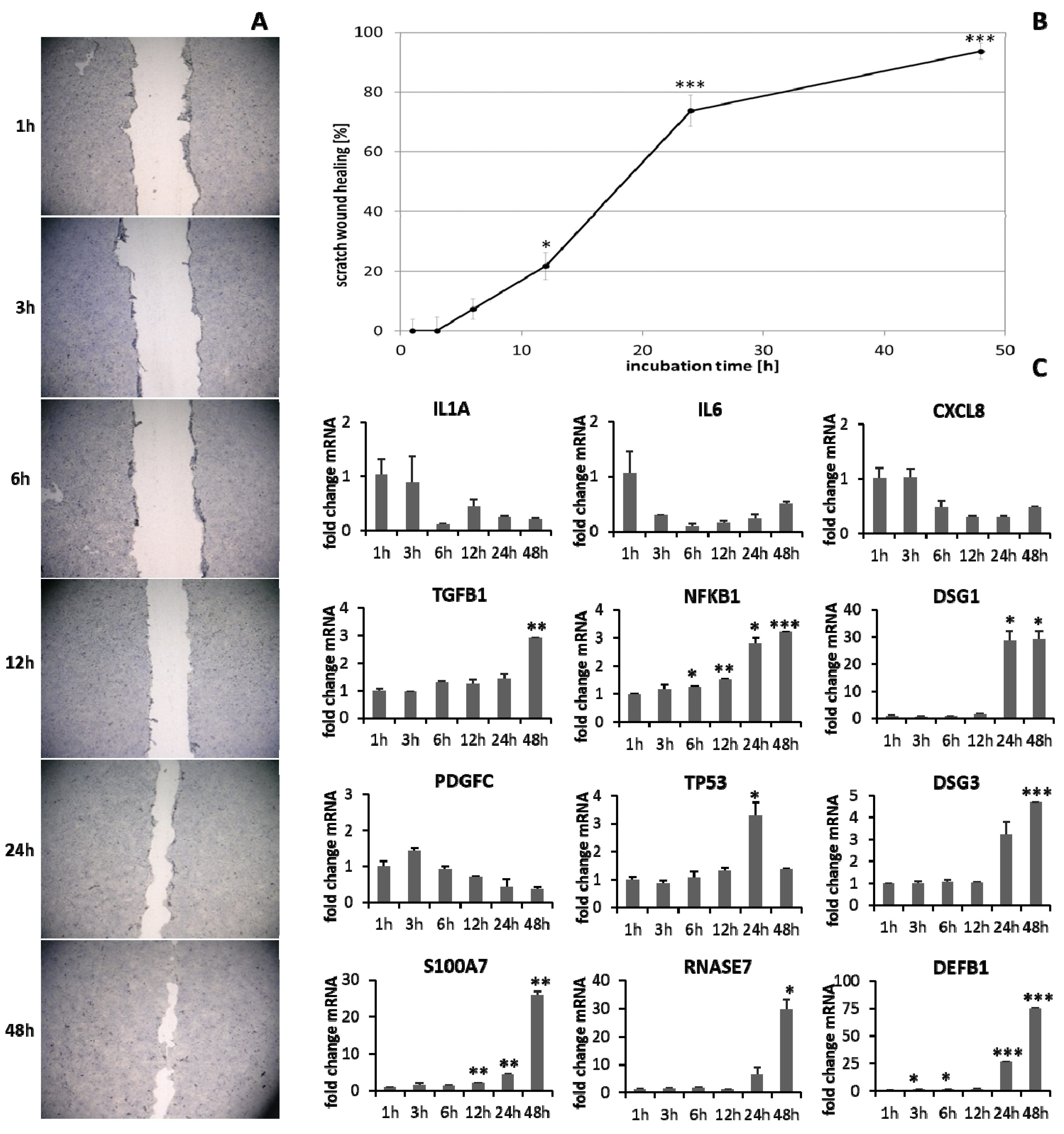
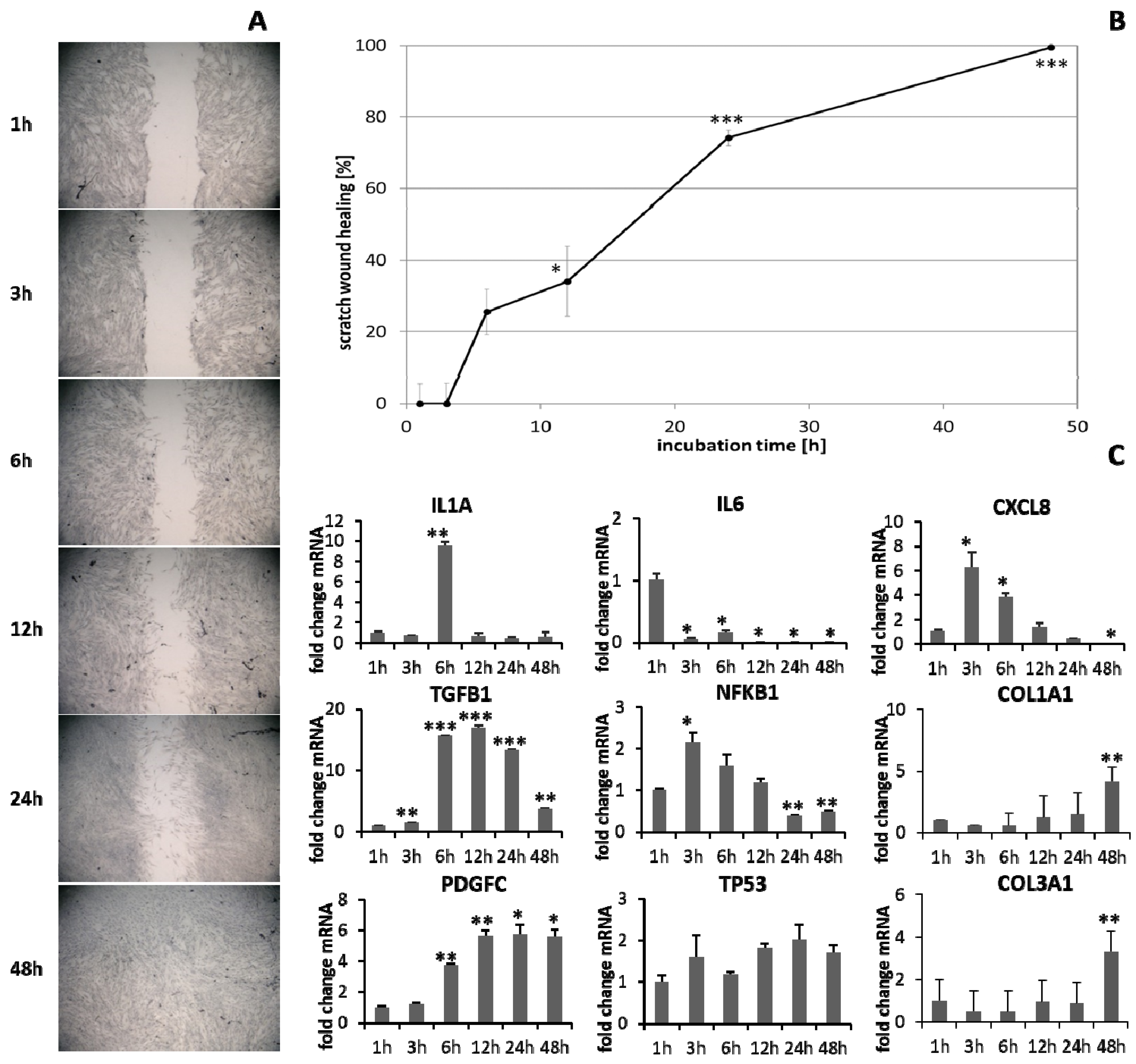
3.1. Scratch Wound Healing and Gene Expression Analysis over Time
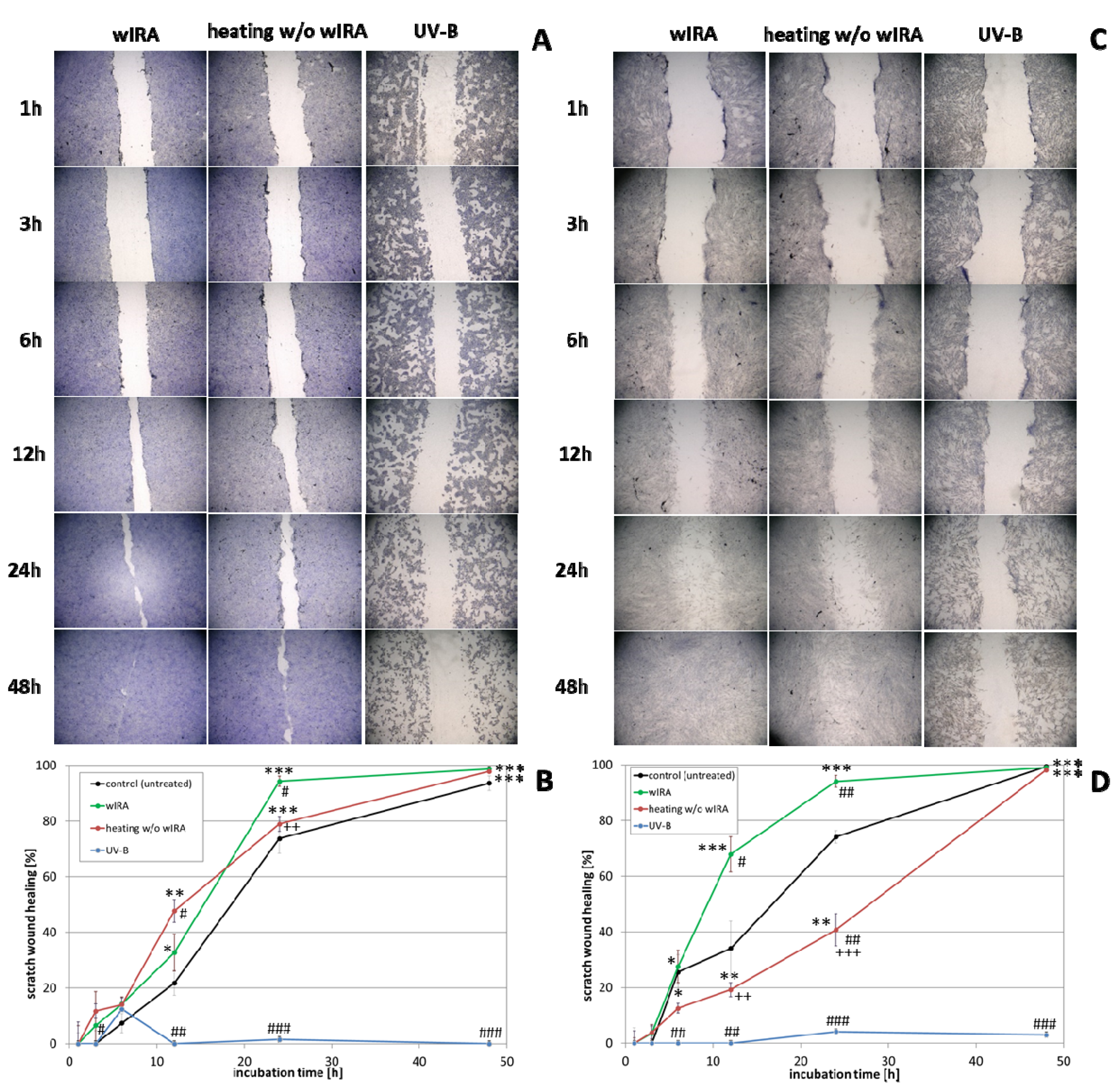
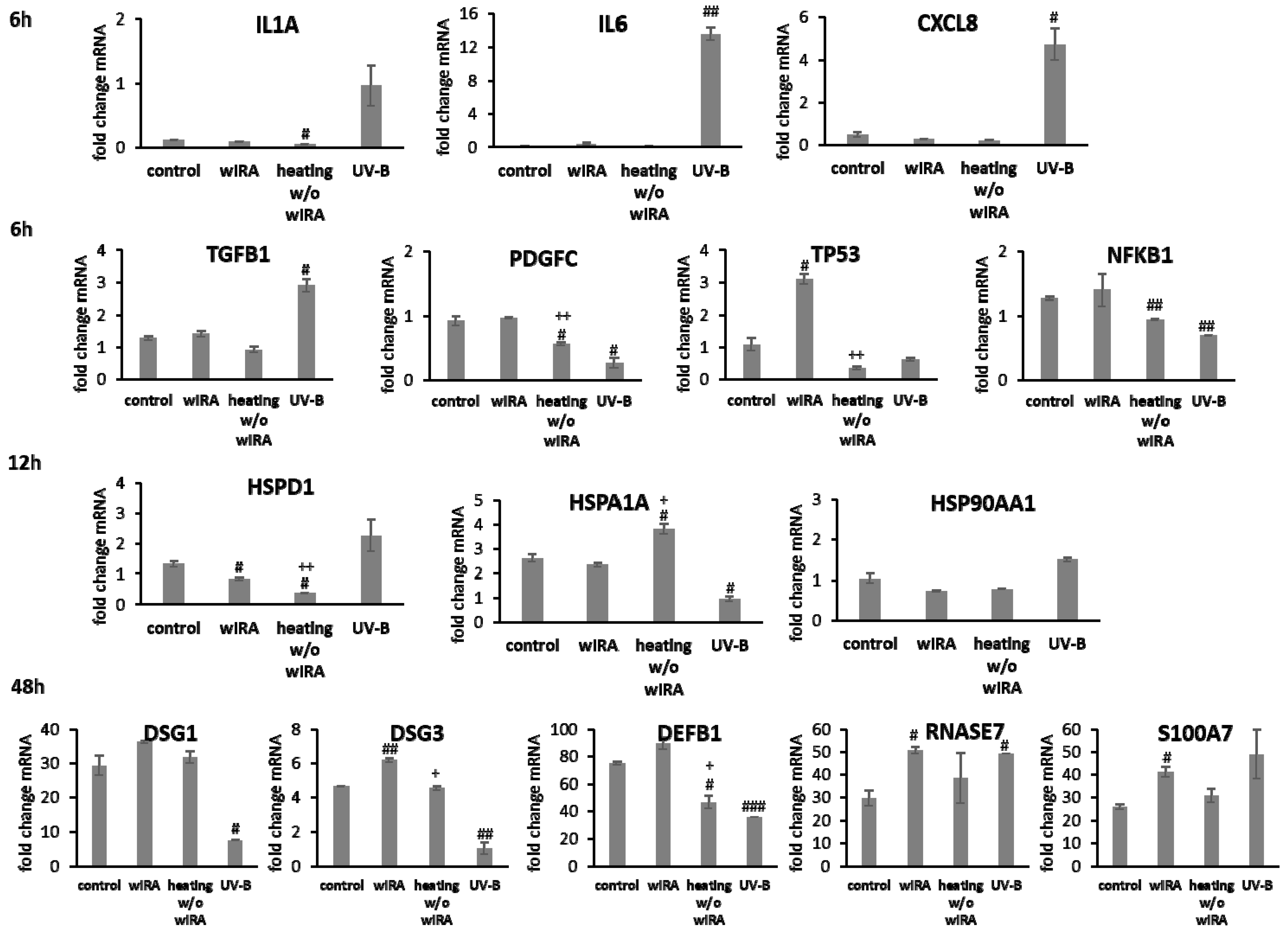
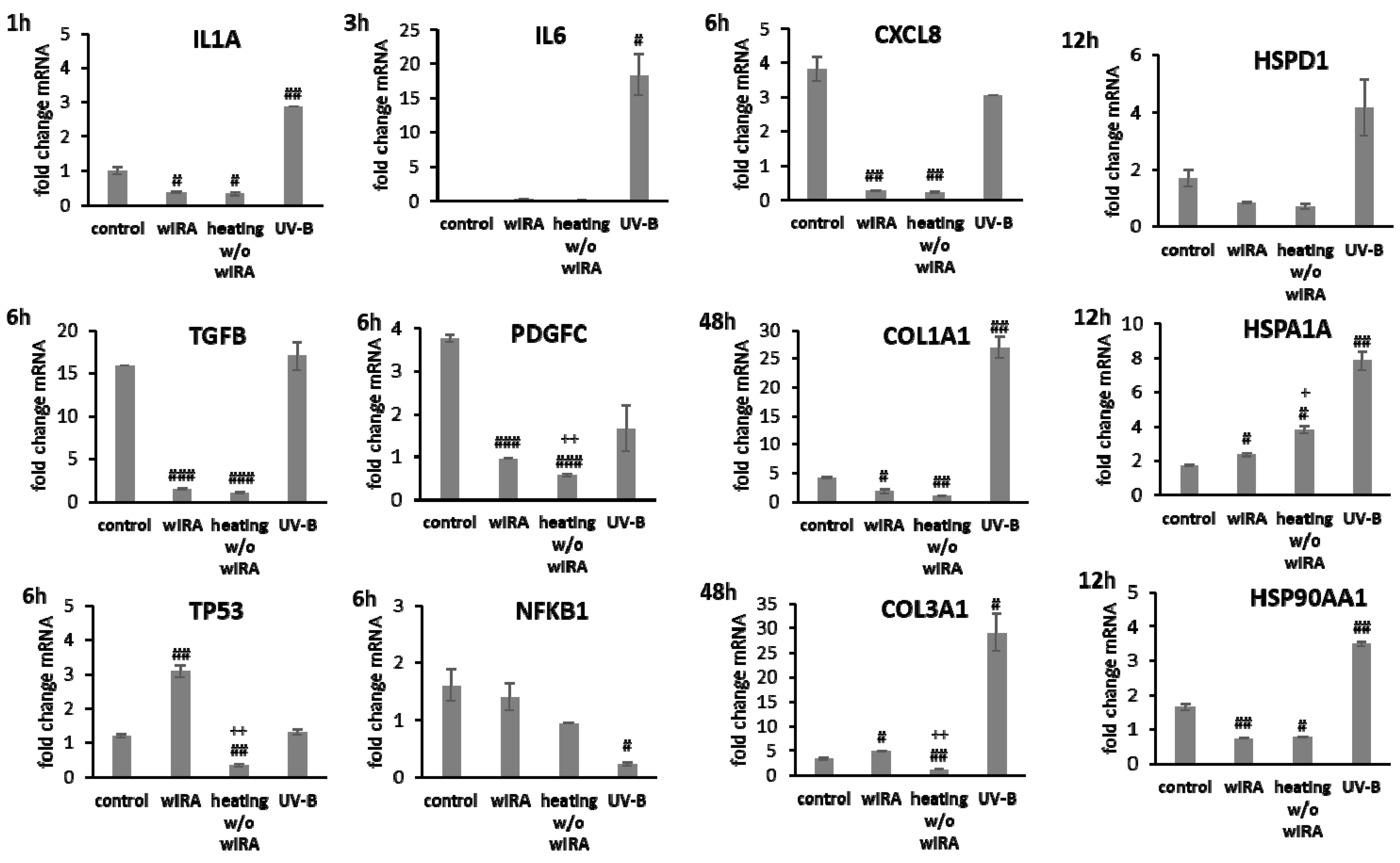
3.2. Scratch Treatment with wIRA and UV-B Radiation
3.3. Scratch Wound Healing under Non-Optimal Conditions
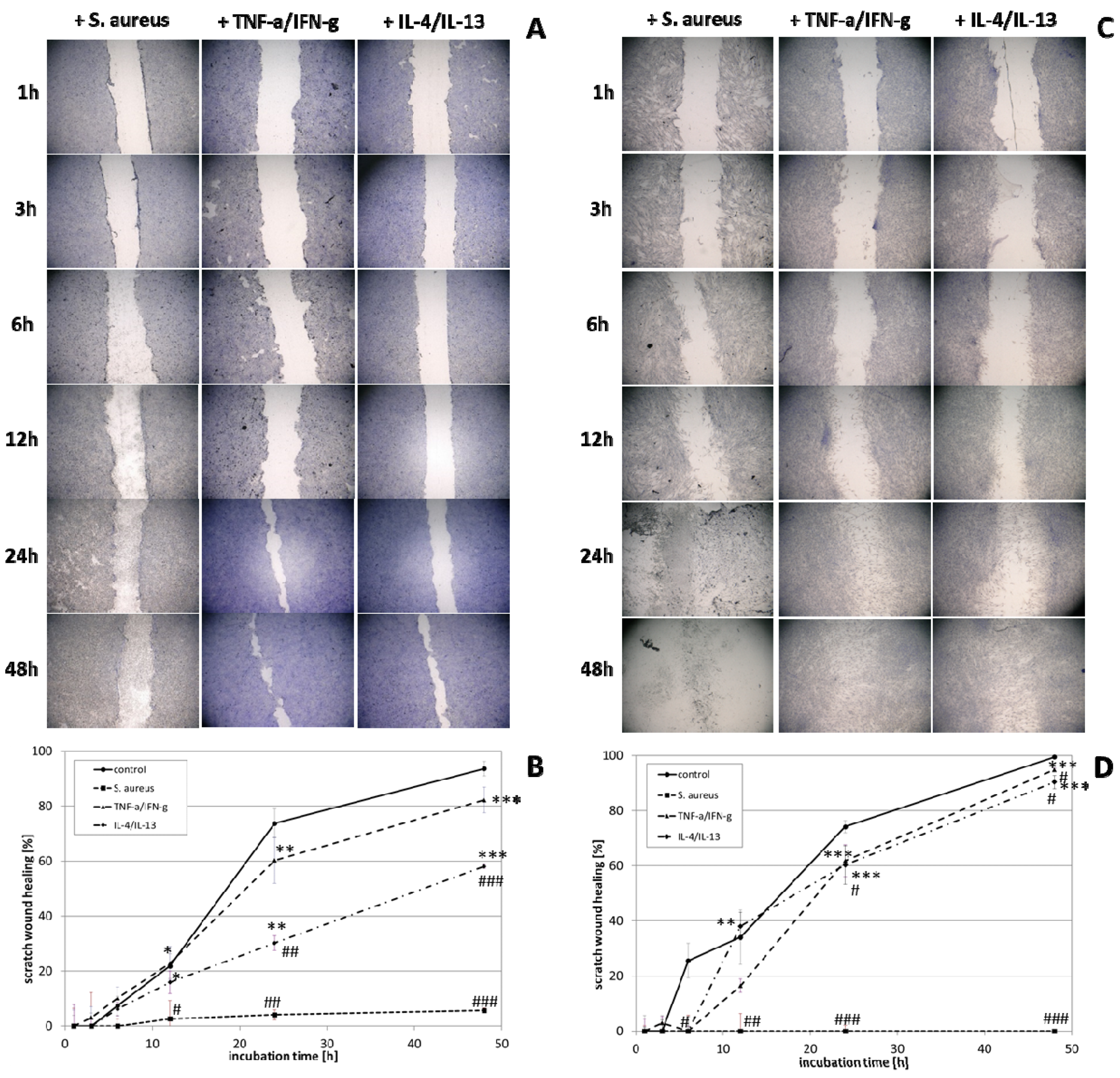
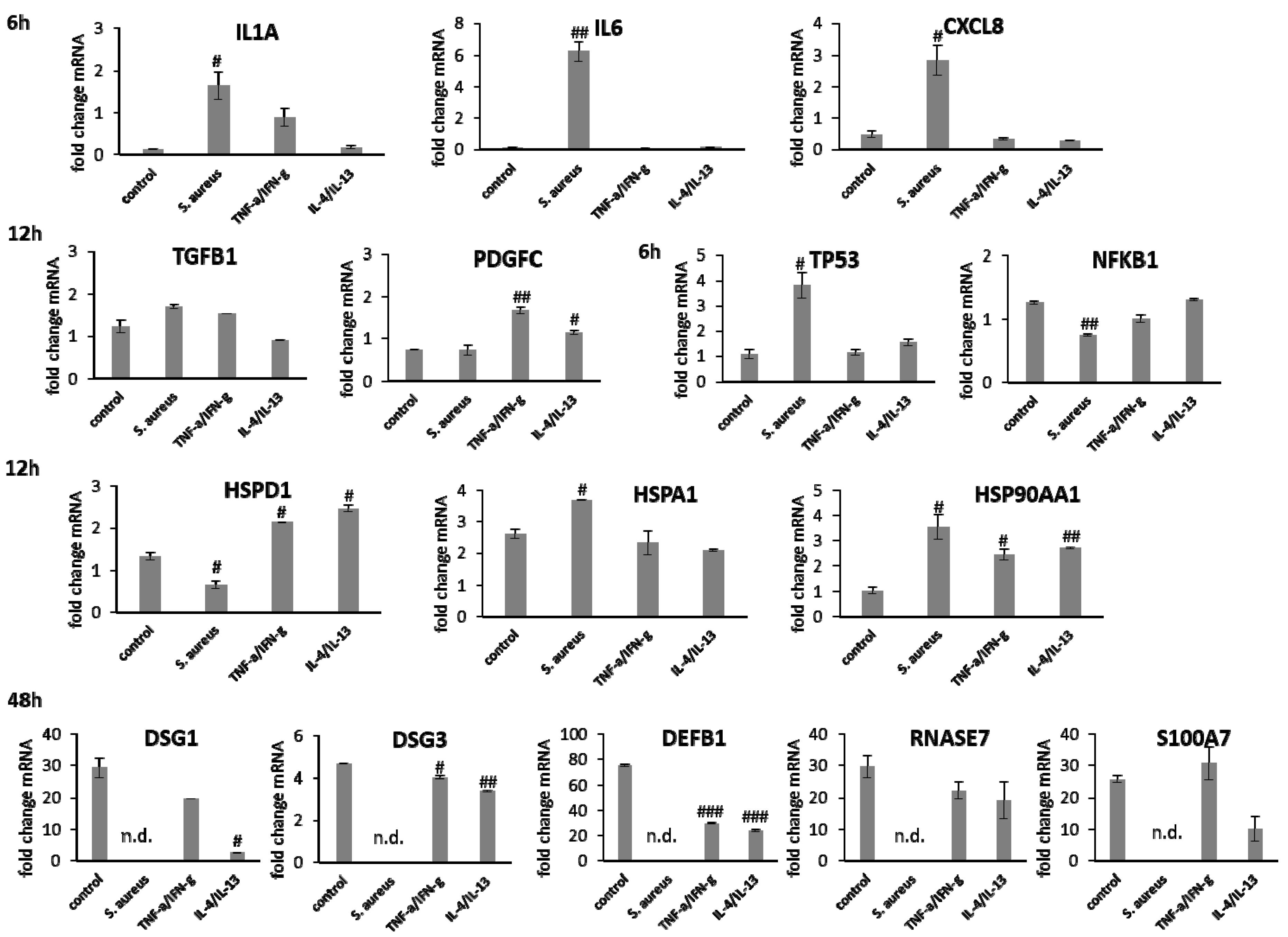
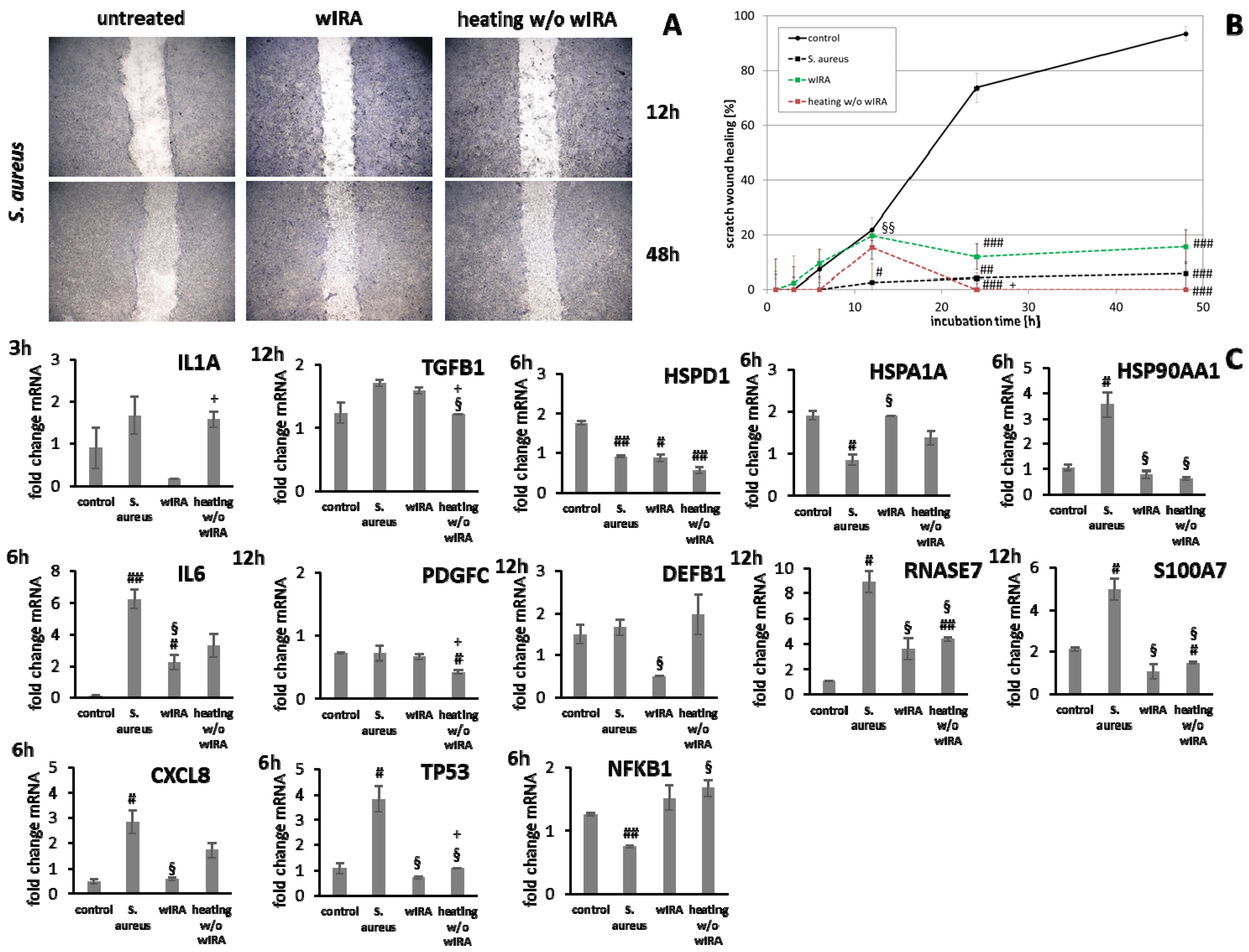
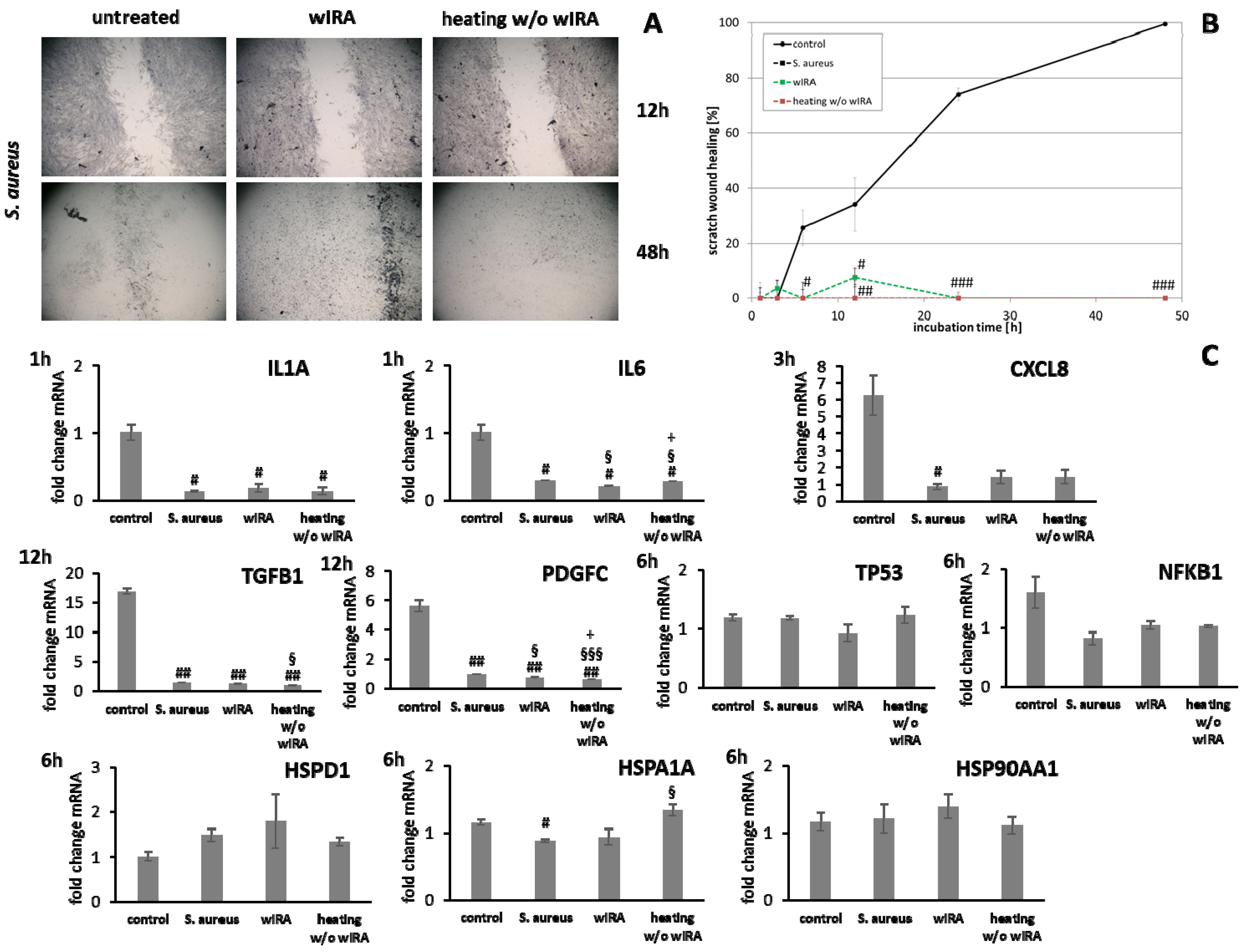
3.4. wIRA Treatment of Cell Scratches during S. aureus Infection
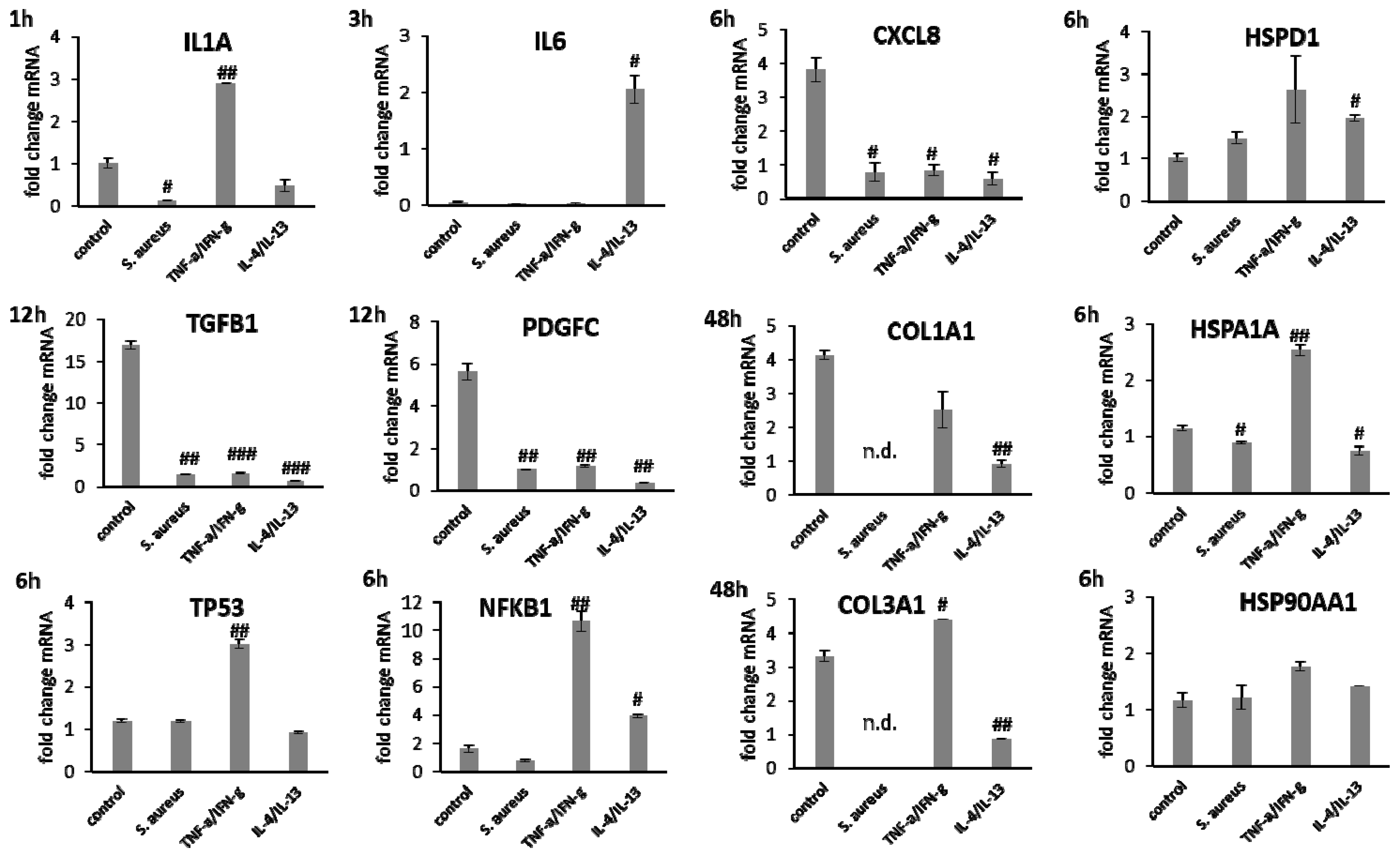
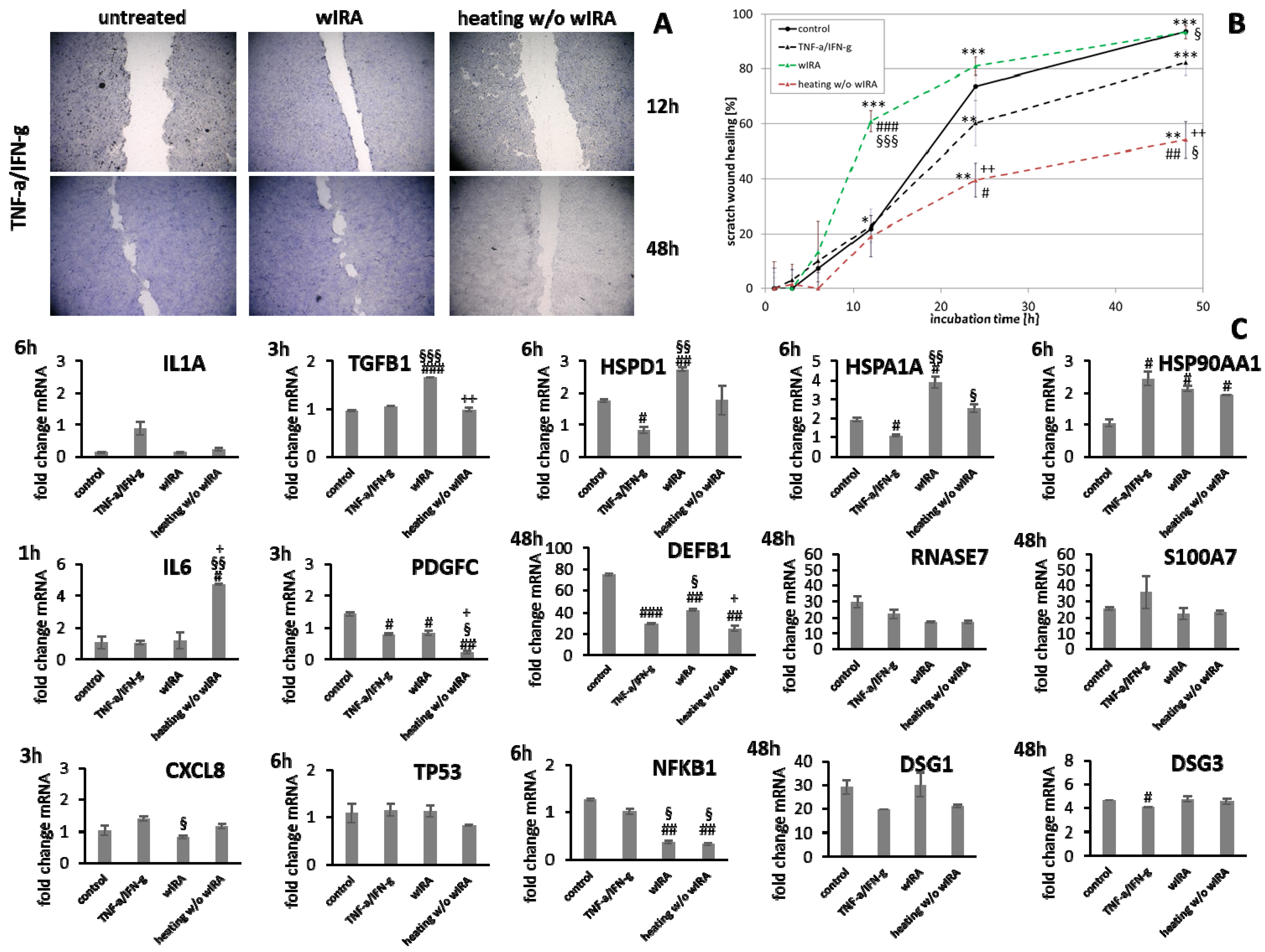
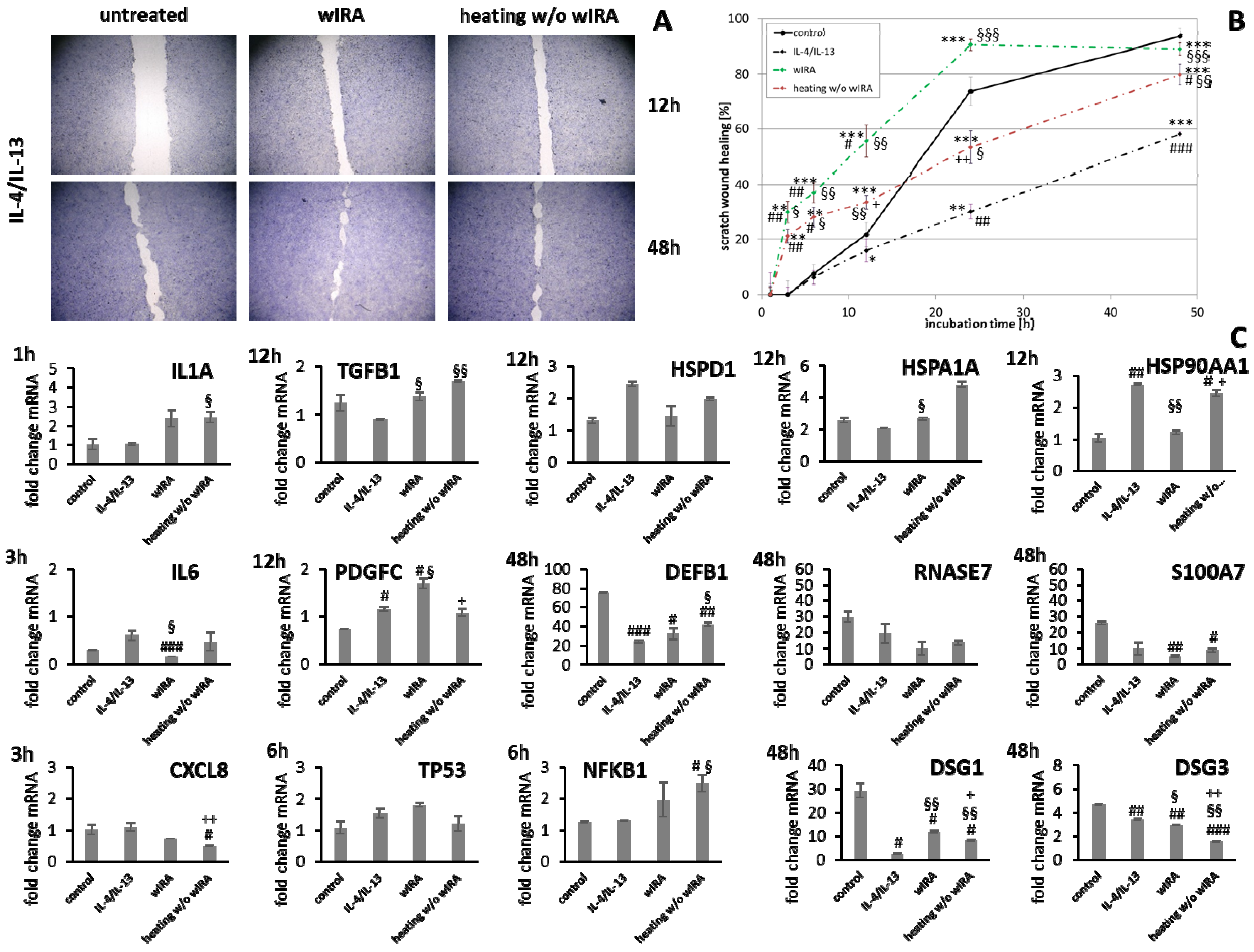
3.5. Stimulation of Scratch Wound Healing under Chronic Inflammatory Conditions by wIRA
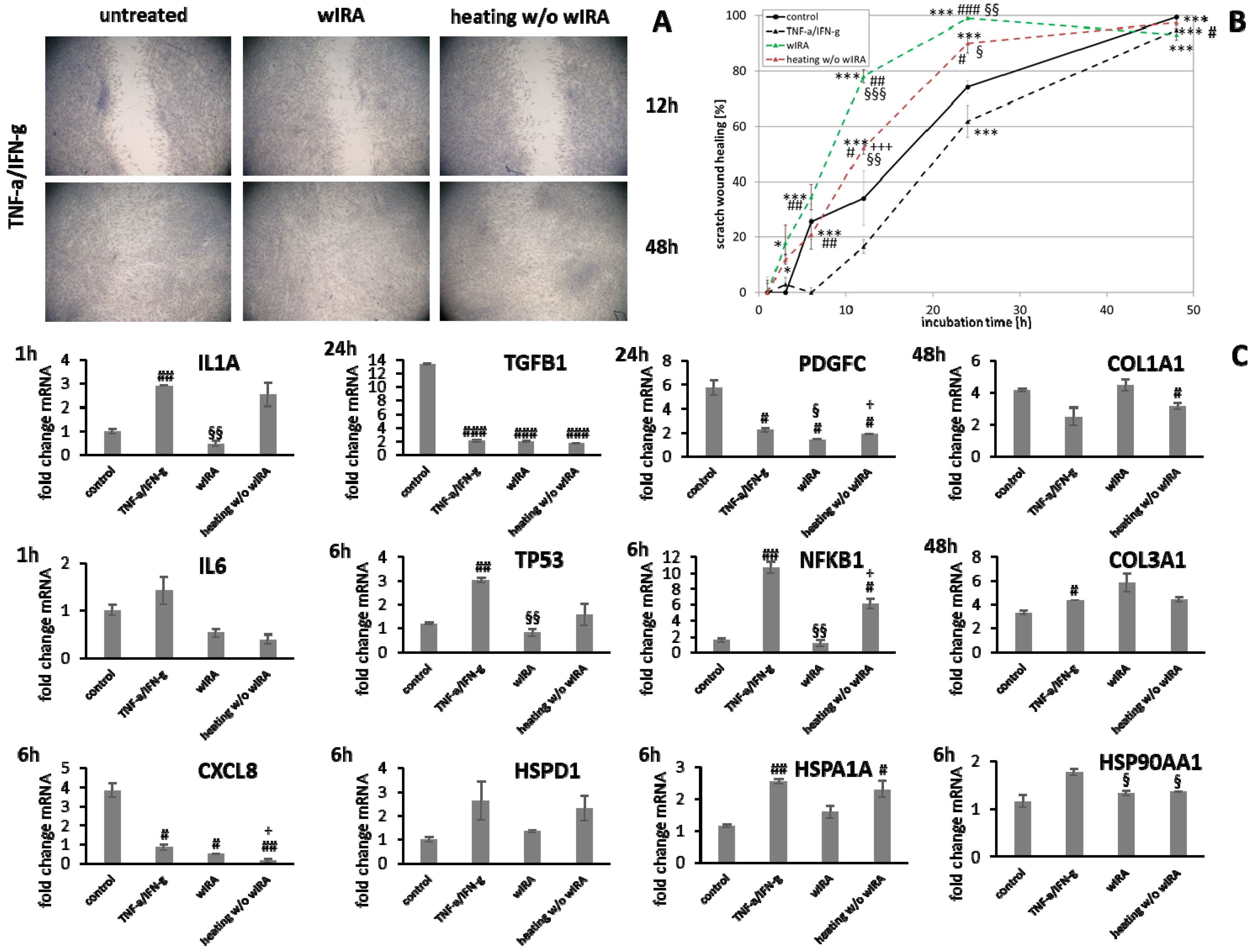
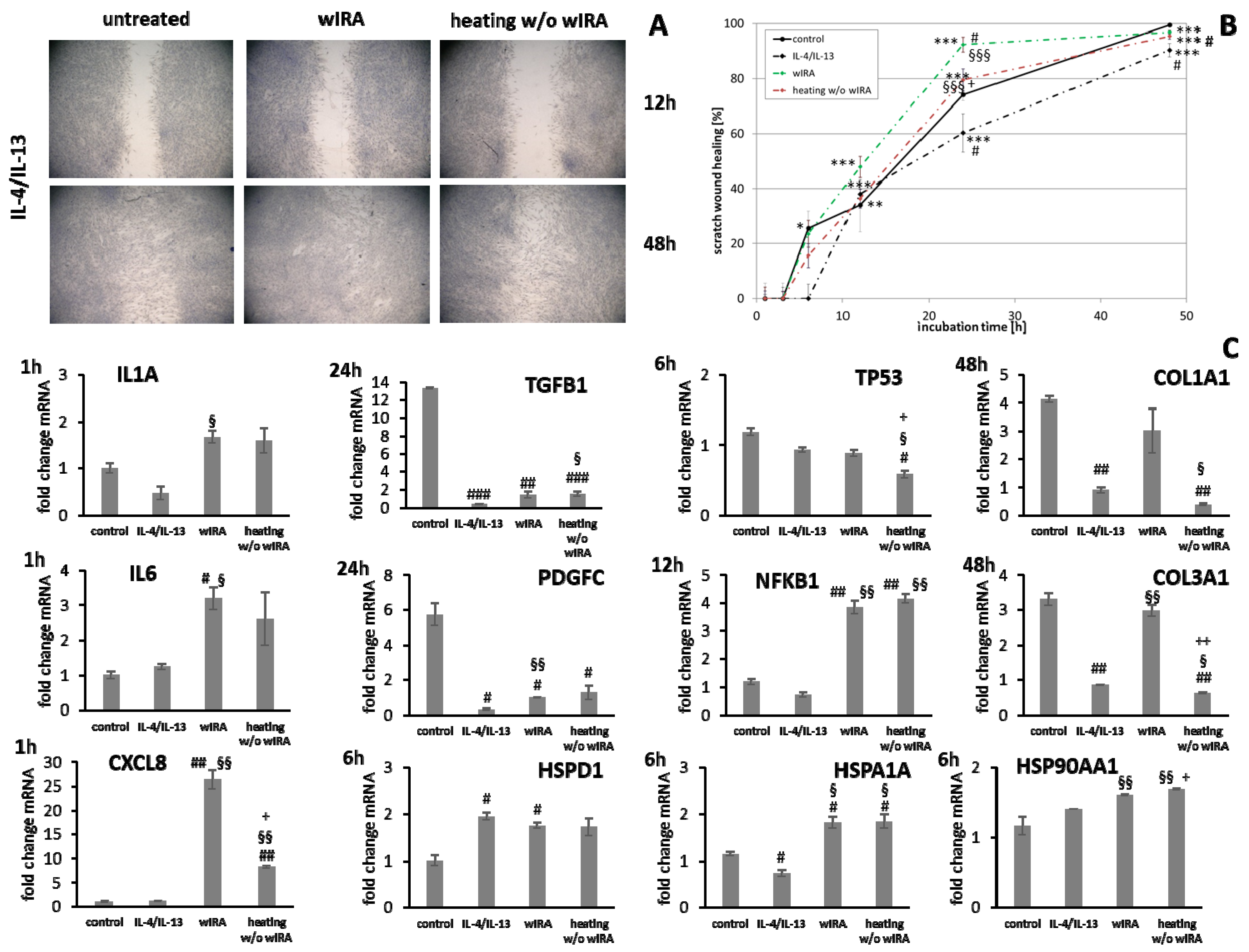
3.6. wIRA Treatment Accelerates Scratch Wound Healing under Anti-Inflammatory Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMP | antimicrobial peptide |
| COL | collagen |
| CXCL | C-X-C motif ligand |
| DSG | desmoglein |
| ELISA | enzyme linked immunosorbent assay |
| HSP | heat shock protein |
| IL | interleukin |
| IFN | interferon |
| MMP | matrix metalloproteinase |
| n.d. | not determined |
| NFKB | nuclear factor ‘kappa-light-chain-enhancer’ of activated B-cells |
| PDGF | platelet derived growth factor |
| qPCR | quantitative polymerase chain reaction |
| ROS | reactive oxygen species |
| TGF | transforming growth factor |
| TNF | tumor necrosis factor |
| TP | tumor suppressor gene |
| UV-B | ultraviolet light B |
| w/o | without |
| wIRA | water-filtered infrared A |
References
- Velnar, T.; Bailey, T.; Smrkolj, V. The wound healing process: An overview of the cellular and molecular mechanisms. J. Int. Med. Res. 2009, 37, 1528–1542. [Google Scholar] [CrossRef] [PubMed]
- Diegelmann, R.F.; Evans, M.C. Wound healing: An overview of acute; fibrotic and delayed healing. Front. Biosci. 2004, 9, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Suter, M.M.; Schulze, K.; Bergman, W.; Welle, M.; Roosje, P.; Müller, E.J. The keratinocyte in epidermal renewal and defence. Vet. Dermatol. 2009, 20, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Choudhury, D. Insulin Promotes Wound Healing by Inactivating NFkβP50/P65 and Activating Protein and Lipid Biosynthesis and alternating Pro/Anti-inflammatory Cytokines Dynamics. Biomol. Concepts 2019, 10, 11–24. [Google Scholar] [CrossRef]
- Rahim, K.; Saleha, S.; Zhu, X.; Huo, L.; Basit, A.; Franco, O.L. Bacterial Contribution in Chronicity of Wounds. Microb. Ecol. 2017, 73, 710–721. [Google Scholar] [CrossRef]
- Jockenhöfer, F.; Chapot, V.; Stoffels-Weindorf, M.; Körber, A.; Klode, J.; Buer, J.; Küpper, B.; Roesch, A.; Dissemond, J. Bacterial spectrum colonizing chronic leg ulcers: A 10-year comparison from a German wound care center. J. Dtsch. Dermatol. Ges. 2014, 12, 1121–1127. [Google Scholar] [CrossRef]
- Raziyeva, K.; Kim, Y.; Zharkinbekov, Z.; Kassymbek, K.; Jimi, S.; Saparov, A. Immunology of Acute and Chronic Wound Healing. Biomolecules 2021, 11, 700. [Google Scholar] [CrossRef]
- Nguyen, J.K.; Austin, E.; Huang, A.; Mamalis, A.; Jagdeo, J. The IL-4/IL-13 axis in skin fibrosis and scarring: Mechanistic concepts and therapeutic targets. Arch. Dermatol. Res. 2020, 312, 81–92. [Google Scholar] [CrossRef]
- Serezani, A.P.M.; Bozdogan, G.; Sehra, S.; Walsh, D.; Krishnamurthy, P.; Sierra Potchanant, E.A.; Nalepa, G.; Goenka, S.; Turner, M.J.; Spandau, D.F.; et al. IL-4 impairs wound healing potential in the skin by repressing fibronectin expression. J. Allergy Clin. Immunol. 2017, 139, 142–151. [Google Scholar] [CrossRef]
- Trautmann, A.; Toksoy, A.; Engelhardt, E.; Bröcker, E.B.; Gillitzer, R. Mast cell involvement in normal human skin wound healing: Expression of monocyte chemoattractant protein-1 is correlated with recruitment of mast cells which synthesize interleukin-4 in vivo. J. Pathol. 2000, 190, 100–106. [Google Scholar] [CrossRef]
- Tada, K.; Ikeda, K.; Tomita, K. Effect of polarized light emitting diode irradiation on wound healing. J. Trauma 2009, 67, 1073–1079. [Google Scholar] [CrossRef]
- Mussttaf, R.A.; Jenkins, D.F.L.; Jha, A.N. Assessing the impact of low level laser therapy (LLLT) on biological systems: A review. Int. J. Radiat. Biol. 2019, 95, 120–143. [Google Scholar] [CrossRef]
- Liao, X.; Xie, G.H.; Liu, H.W.; Cheng, B.; Li, S.H.; Xie, S.; Xiao, L.L.; Fu, X.B. Helium-neon laser irradiation promotes the proliferation and migration of human epidermal stem cells in vitro: Proposed mechanism for enhanced wound re-epithelialization. Photomed. Laser Surg. 2014, 32, 219–225. [Google Scholar] [CrossRef]
- Haas, A.F.; Isseroff, R.R.; Wheeland, R.G.; Rood, P.A.; Graves, P.J. Low-energy helium-neon laser irradiation increases the motility of cultured human keratinocytes. J. Investig. Dermatol. 1990, 94, 822–826. [Google Scholar] [CrossRef]
- Houreld, N.; Abrahamse, H. Low-intensity laser irradiation stimulates wound healing in diabetic wounded fibroblast cells (WS1). Diabetes Technol. 2010, 12, 971–978. [Google Scholar] [CrossRef]
- Leite, S.N.; Andrade, T.A.; Masson-Meyers, D.S.; Leite, M.N.; Enwemeka, C.S.; Frade, M.A. Phototherapy promotes healing of cutaneous wounds in undernourished rats. Bras. Dermatol. 2014, 89, 899–904. [Google Scholar] [CrossRef]
- Caetano, K.S.; Frade, M.A.; Minatel, D.G.; Santana, L.A.; Enwemeka, C.S. Phototherapy improves healing of chronic venous ulcers. Photomed. Laser Surg. 2009, 27, 111–118. [Google Scholar] [CrossRef]
- Minatel, D.G.; Frade, M.A.; França, S.C.; Enwemeka, C.S. Phototherapy promotes healing of chronic diabetic leg ulcers that failed to respond to other therapies. Lasers Surg. Med. 2009, 41, 433–441. [Google Scholar] [CrossRef]
- Powell, M.W.; Carnegie, D.E.; Burke, T.J. Reversal of diabetic peripheral neuropathy and new wound incidence: The role of MIRE. Adv. Ski. Wound Care 2004, 17, 295–300. [Google Scholar] [CrossRef]
- Reddy, G.K.; Stehno-Bittel, L.; Enwemeka, C.S. Laser photostimulation accelerates wound healing in diabetic rats. Wound Repair Regen. 2001, 9, 248–255. [Google Scholar] [CrossRef]
- Reddy, G.K.; Stehno-Bittel, L.; Enwemeka, C.S. Matrix remodeling in healing rabbit Achilles tendon. Wound Repair Regen. 1999, 7, 518–527. [Google Scholar] [CrossRef]
- Hawkins, D.; Abrahamse, H. Effect of multiple exposures of low-level laser therapy on the cellular responses of wounded human skin fibroblasts. Photomed. Laser Surg. 2006, 24, 705–714. [Google Scholar] [CrossRef]
- von Felbert, V.; Schumann, H.; Mercer, J.B.; Strasser, W.; Daeschlein, G.; Hoffmann, G. Therapy of chronic wounds with water-filtered infrared-A (wIRA). GMS Krankenhhyg. Interdiszip. 2008, 2, Doc52. [Google Scholar]
- Hartel, M.; Illing, P.; Mercer, J.B.; Lademann, J.; Daeschlein, G.; Hoffmann, G. Therapy of acute wounds with water-filtered infrared-A (wIRA). GMS Krankenhhyg. Interdiszip. 2007, 2, Doc53. [Google Scholar]
- Winkel, R.; Hoffmann, G.; Hoffmann, R. Wassergefiltertes Infrarot A (wIRA) hilft Wunden heilen [Water-filtered infrared-A (wIRA) promotes wound healing]. Chirurg 2014, 85, 980–992. [Google Scholar] [CrossRef][Green Version]
- Mercer, J.B.; Nielsen, S.P.; Hoffmann, G. Improvement of wound healing by water-filtered infrared-A (wIRA) in patients with chronic venous stasis ulcers of the lower legs including evaluation using infrared thermography. Ger. Med. Sci. 2008, 6, Doc11. [Google Scholar]
- Hartel, M.; Hoffmann, G.; Wente, M.N.; Martignoni, M.E.; Büchler, M.W.; Friess, H. Randomized clinical trial of the influence of local water-filtered infrared A irradiation on wound healing after abdominal surgery. Br. J. Surg. 2006, 93, 952–960. [Google Scholar] [CrossRef]
- Schreml, S.; Szeimies, R.M.; Prantl, L.; Karrer, S.; Landthaler, M.; Babilas, P. Oxygen in acute and chronic wound healing. Br. J. Dermatol. 2010, 163, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Danno, K.; Mori, N.; Toda, K.; Kobayashi, T.; Utani, A. Near-infrared irradiation stimulates cutaneous wound repair: Laboratory experiments on possible mechanisms. Photodermatol. Photoimmunol. Photomed. 2001, 17, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Applegate, L.A.; Scaletta, C.; Panizzon, R.; Frenk, E.; Hohlfeld, P.; Schwarzkopf, S. Induction of the putative protective protein ferritin by infrared radiation: Implications in skin repair. Int. J. Mol. Med. 2000, 5, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Ehrlicher, A.; Betz, T.; Stuhrmann, B.; Koch, D.; Milner, V.; Raizen, M.G.; Kas, J. Guiding neuronal growth with light. Proc. Natl. Acad. Sci. USA 2002, 99, 16024–16028. [Google Scholar] [CrossRef]
- Zöller, N.; König, A.; Butting, M.; Kaufmann, R.; Bernd, A.; Valesky, E.; Kippenberger, S. Water-filtered near-infrared influences collagen synthesis of keloid-fibroblasts in contrast to normal foreskin fibroblasts. J. Photochem. Photobiol. B 2016, 163, 194–202. [Google Scholar] [CrossRef]
- Knels, L.; Valtink, M.; Piazena, H.; de la Vega Marin, J.; Gommel, K.; Lupp, A.; Roehlecke, C.; Mehner, M.; Funk, R.H. Effects of narrow-band IR-A and of water-filtered infrared A on fibroblasts. Photochem. Photobiol. 2016, 92, 475–487. [Google Scholar] [CrossRef]
- Mann, E.R.; Smith, K.M.; Bernardo, D.; Al-Hassi, H.O.; Knight, S.C.; Hart, A.L. Review: Skin and the Immune System. J. Clin. Exp. Dermatol. Res. 2012, S2, 3. [Google Scholar] [CrossRef]
- Shiraki, Y.; Ishibashi, Y.; Hiruma, M.; Nishikawa, A.; Ikeda, S. Cytokine secretion profiles of human keratinocytes during Trichophyton tonsurans and Arthroderma benhamiae infections. J. Med. Microbiol. 2006, 55, 1175–1185. [Google Scholar] [CrossRef]
- Büth, H.; Luigi Buttigieg, P.; Ostafe, R.; Rehders, M.; Dannenmann, S.R.; Schaschke, N.; Stark, H.J.; Boukamp, P.; Brix, K. Cathepsin B is essential for regeneration of scratch-wounded normal human epidermal keratinocytes. Eur. J. Cell Biol. 2007, 86, 747–761. [Google Scholar] [CrossRef]
- Wiegand, C.; Abel, M.; Hipler, U.C.; Elsner, P. Effect of non-adhering dressings on promotion of fibroblast proliferation and wound healing in vitro. Sci. Rep. 2019, 9, 4320. [Google Scholar] [CrossRef]
- Wiegand, C.; Tittelbach, J.; Hipler, U.-C.; Elsner, P. Water-filtered infrared A (wIRA): From observations in clinical studies to complex in vitro models. In Water-Filtered Infrared-A Radiation: From Principle to Practice, 1st ed.; Vaupel, P., Ed.; UNI-MED: Bremen, Germany, accepted.
- Wiederholt, T.; Heise, R.; Skazik, C.; Marquardt, Y.; Joussen, S.; Erdmann, K.; Schröder, H.; Merk, H.F.; Baron, J.M. Calcium pantothenate modulates gene expression in proliferating human dermal fibroblasts. Exp. Dermatol. 2009, 18, 969–978. [Google Scholar] [CrossRef]
- Rose, M.T. Effect of growth factors on the migration of equine oral and limb fibroblasts using an in vitro scratch assay. Vet. J. 2012, 193, 539–544. [Google Scholar] [CrossRef]
- Maruyama, M.; Rhee, C.; Utsunomiya, T.; Zhang, N.; Ueno, M.; Yao, Z.; Goodman, S.B. Modulation of the Inflammatory Response and Bone Healing. Front. Endocrinol. 2020, 11, 386. [Google Scholar] [CrossRef]
- Ricci, G.; Patrizi, A.; Baldi, E.; Menna, G.; Tabanelli, M.; Masi, M. Long-term follow-up of atopic dermatitis: Retrospective analysis of related risk factors and association with concomitant allergic diseases. J. Am. Acad. Dermatol. 2006, 55, 765–771. [Google Scholar] [CrossRef]
- Jung, T.; Grune, T. Experimental basis for discriminating between thermal and athermal effects of water-filtered infrared A irradiation. Ann. N. Y. Acad. Sci. 2012, 1259, 33–38. [Google Scholar] [CrossRef]
- Jung, T.; Höhn, A.; Lau, A.M.; Piazena, H.; Grune, T. An experimental setup for the measurement of nonthermal effects during water-filtered infrared A-irradiation of mammalian cell cultures. Photochem. Photobiol. 2012, 88, 371–380. [Google Scholar] [CrossRef]
- Sims, J.; Towne, J.; Blumberg, H. 11 IL-1 family members in inflammatory skin disease. Ernst. Scher. Res. Found Workshop 2006, 56, 187–191. [Google Scholar]
- Sauder, D.N. The role of epidermal cytokines in inflammatory skin diseases. J. Investig. Dermatol. 1990, 95 (Suppl. S5), 27S–28S. [Google Scholar] [CrossRef]
- Choi, J.H.; Jun, J.H.; Kim, J.H.; Sung, H.J.; Lee, J.H. Synergistic effect of interleukin-6 and hyaluronic acid on cell migration and ERK activation in human keratinocytes. J. Korean Med. Sci. 2014, 29 (Suppl. S3), S210–S216. [Google Scholar] [CrossRef]
- Ramezanpour, M.; Smith, J.L.P.; Ooi, M.L.; Gouzos, M.; Psaltis, A.J.; Wormald, P.J.; Vreugde, S. Deferiprone has anti-inflammatory properties and reduces fibroblast migration in vitro. Sci. Rep. 2019, 9, 2378. [Google Scholar] [CrossRef]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef]
- Yang, L.; Chan, T.; Demare, J.; Iwashina, T.; Ghahary, A.; Scott, P.G.; Tredget, E.E. Healing of burn wounds in transgenic mice overexpressing transforming growth factor-beta 1 in the epidermis. Am. J. Pathol. 2001, 159, 2147–2157. [Google Scholar] [CrossRef]
- Desmoulière, A.; Geinoz, A.; Gabbiani, F.; Gabbiani, G. Transforming growth factor-beta 1 induces alpha-smooth muscle actin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. J. Cell Biol. 1993, 122, 103–111. [Google Scholar] [CrossRef]
- Schmid, P.; Kunz, S.; Cerletti, N.; McMaster, G.; Cox, D. Injury induced expression of TGF-beta 1 mRNA is enhanced by exogenously applied TGF-beta S. Biochem. Biophys. Res. Commun. 1993, 194, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Jiang, L.; Li, H.; Shi, H.; Luo, H.; Zhang, Y.; Yu, C.; Jin, Y. Mesenchymal stem cells prevent hypertrophic scar formation via inflammatory regulation when undergoing apoptosis. J. Investig. Dermatol. 2014, 134, 2648–2657. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhao, X.; Ji, S.; He, C.; Wang, G.; Tang, C.; Gu, S.; Yin, C. The use of gene activated matrix to mediate effective SMAD2 gene silencing against hypertrophic scar. Biomaterials 2014, 35, 2488–2498. [Google Scholar] [CrossRef] [PubMed]
- Jester, J.V.; Ho-Chang, J. Modulation of cultured corneal keratocyte phenotype by growth factors/cytokines control in vitro contractility and extracellular matrix contraction. Exp. Eye Res. 2003, 77, 581–592. [Google Scholar] [CrossRef]
- Peng, Y.; Wu, S.; Tang, Q.; Li, S.; Peng, C. KGF-1 accelerates wound contraction through the TGF-β1/Smad signaling pathway in a double-paracrine manner. J. Biol. Chem. 2019, 294, 8361–8370. [Google Scholar] [CrossRef]
- Merlo, S.; Frasca, G.; Canonico, P.L.; Sortino, M.A. Differential involvement of estrogen receptor alpha and estrogen receptor beta in the healing promoting effect of estrogen in human keratinocytes. J. Endocrinol. 2009, 200, 189–197. [Google Scholar] [CrossRef]
- Pinto, D.; Marzani, B.; Minervini, F.; Calasso, M.; Giuliani, G.; Gobbetti, M.; De Angelis, M. Plantaricin A synthesized by Lactobacillus plantarum induces in vitro proliferation and migration of human keratinocytes and increases the expression of TGF-β1; FGF7; VEGF-A and IL-8 genes. Peptides 2011, 32, 1815–1824. [Google Scholar] [CrossRef]
- Glim, J.E.; Niessen, F.B.; Everts, V.; van Egmond, M.; Beelen, R.H. Platelet derived growth factor-CC secreted by M2 macrophages induces alpha-smooth muscle actin expression by dermal and gingival fibroblasts. Immunobiology 2013, 218, 924–929. [Google Scholar] [CrossRef]
- Li, Q.; Verma, I.M. NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Qin, J.Z.; Chaturvedi, V.; Denning, M.F.; Choubey, D.; Diaz, M.O.; Nickoloff, B.J. Role of NF-kappaB in the apoptotic-resistant phenotype of keratinocytes. J. Biol. Chem. 1999, 274, 37957–37964. [Google Scholar] [CrossRef]
- Muresan, X.M.; Sticozzi, C.; Belmonte, G.; Cervellati, F.; Ferrara, F.; Lila, M.A.; Valacchi, G. SR-B1 involvement in keratinocytes in vitro wound closure. Arch. Biochem. Biophys. 2018, 658, 1–6. [Google Scholar] [CrossRef]
- Na, J.; Lee, K.; Na, W.; Shin, J.Y.; Lee, M.J.; Yune, T.Y.; Lee, H.K.; Jung, H.S.; Kim, W.S.; Ju, B.G. Histone H3K27 Demethylase JMJD3 in Cooperation with NF-κB Regulates Keratinocyte Wound Healing. J. Investig. Dermatol. 2016, 136, 847–858. [Google Scholar] [CrossRef]
- Parveen, N.; Varman, R.; Nair, S.; Das, G.; Ghosh, S.; Mukhopadhyay, S. Endocytosis of Mycobacterium tuberculosis heat shock protein 60 is required to induce interleukin-10 production in macrophages. J. Biol. Chem. 2013, 288, 24956–24971. [Google Scholar] [CrossRef]
- Pockley, A.G.; Muthana, M.; Calderwood, S.K. The dual immunoregulatory roles of stress proteins. Trends Biochem. Sci. 2008, 33, 71–79. [Google Scholar] [CrossRef]
- Da Costa, C.U.; Wantia, N.; Kirschning, C.J.; Busch, D.H.; Rodriguez, N.; Wagner, H.; Miethke, T. Heat shock protein 60 from Chlamydia pneumoniae elicits an unusual set of inflammatory responses via Toll-like receptor 2 and 4 in vivo. Eur. J. Immunol. 2004, 34, 2874–2884. [Google Scholar] [CrossRef]
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009, 14, 105–111. [Google Scholar] [CrossRef]
- Welch, W.J.; Feramisco, J.R. Purification of the major mammalian heat shock proteins. J. Biol. Chem. 1982, 257, 14949–14959. [Google Scholar] [CrossRef]
- Welch, W.J. The role of heat-shock proteins as molecular chaperones. Curr. Opin. Cell Biol. 1991, 3, 1033–1038. [Google Scholar] [CrossRef]
- Makhnevych, T.; Houry, W.A. The role of Hsp90 in protein complex assembly. Biochim. Biophys. Acta 2012, 1823, 674–682. [Google Scholar] [CrossRef]
- Arya, R.; Mallik, M.; Lakhotia, S.C. Heat shock genes—integrating cell survival and death. J. Biosci. 2007, 32, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Pei, Q.; Ni, W.; Fu, X.; Zhang, W.; Song, C.; Peng, Y.; Guo, Q.; Dong, J.; Yao, M. HSPA1A Protects Cells from Thermal Stress by Impeding ESCRT-0-Mediated Autophagic Flux in Epidermal Thermoresistance. J. Investig. Dermatol. 2021, 141, 48–58. [Google Scholar] [CrossRef]
- Voos, W.; Röttgers, K. Molecular chaperones as essential mediators of mitochondrial biogenesis. Biochim. Biophys. Acta 2002, 1592, 51–62. [Google Scholar] [CrossRef]
- Pei, W.; Tanaka, K.; Huang, S.C.; Xu, L.; Liu, B.; Sinclair, J.; Idol, J.; Varshney, G.K.; Huang, H.; Lin, S.; et al. Extracellular HSP60 triggers tissue regeneration and wound healing by regulating inflammation and cell proliferation. NPJ Regen. Med. 2016, 1, 16013. [Google Scholar] [CrossRef] [PubMed]
- Niyonsaba, F.; Kiatsurayanon, C.; Chieosilapatham, P.; Ogawa, H. Friends or Foes? Host defense (antimicrobial) peptides and proteins in human skin diseases. Exp. Dermatol. 2017, 26, 989–998. [Google Scholar] [CrossRef]
- Fritz, P.; Beck-Jendroschek, V.; Brasch, J. Inhibition of dermatophytes by the antimicrobial peptides human β-defensin-2; ribonuclease 7 and psoriasin. Med. Mycol. 2012, 50, 579–584. [Google Scholar] [CrossRef]
- Gläser, R.; Harder, J.; Lange, H.; Bartels, J.; Christophers, E.; Schröder, J.M. Antimicrobial psoriasin (S100A7) protects human skin from Escherichia coli infection. Nat. Immunol. 2005, 6, 57–64. [Google Scholar] [CrossRef]
- Harder, J.; Bartels, J.; Christophers, E.; Schroder, J.M. Isolation and characterization of human beta -defensin-3; a novel human inducible peptide antibiotic. J. Biol. Chem. 2001, 276, 5707–5713. [Google Scholar] [CrossRef]
- Harder, J.; Schroder, J.M. RNase 7; a novel innate immune defense antimicrobial protein of healthy human skin. J. Biol. Chem. 2002, 277, 46779–46784. [Google Scholar] [CrossRef]
- Yasin, B.; Pang, M.; Turner, J.S.; Cho, Y.; Dinh, N.N.; Waring, A.J.; Lehrer, R.I.; Wagar, E.A. Evaluation of the inactivation of infectious Herpes simplex virus by host-defense peptides. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 187–194. [Google Scholar] [CrossRef]
- Pfalzgraff, A.; Bárcena-Varela, S.; Heinbockel, L.; Gutsmann, T.; Brandenburg, K.; Martinez-de-Tejada, G.; Weindl, G. Antimicrobial endotoxin-neutralizing peptides promote keratinocyte migration via P2X7 receptor activation and accelerate wound healing in vivo. Br. J. Pharm. 2018, 175, 3581–3593. [Google Scholar] [CrossRef] [PubMed]
- Pfalzgraff, A.; Heinbockel, L.; Su, Q.; Gutsmann, T.; Brandenburg, K.; Weindl, G. Synthetic antimicrobial and LPS-neutralising peptides suppress inflammatory and immune responses in skin cells and promote keratinocyte migration. Sci. Rep. 2016, 6, 31577. [Google Scholar] [CrossRef] [PubMed]
- Galkowska, H.; Olszewski, W.L.; Wojewodzka, U. Expression of natural antimicrobial peptide beta-defensin-2 and Langerhans cell accumulation in epidermis from human non-healing leg ulcers. Folia Histochem. Cytobiol. 2005, 43, 133–136. [Google Scholar]
- Heilborn, J.D.; Nilsson, M.F.; Kratz, G.; Weber, G.; Sørensen, O.; Borregaard, N.; Ståhle-Bäckdahl, M. The cathelicidin anti-microbial peptide LL-37 is involved in re-epithelialization of human skin wounds and is lacking in chronic ulcer epithelium. J. Investig. Dermatol. 2003, 120, 379–389. [Google Scholar] [CrossRef]
- Howell, M.D.; Fairchild, H.R.; Kim, B.E.; Bin, L.; Boguniewicz, M.; Redzic, J.S.; Hansen, K.C.; Leung, D.Y. Th2 cytokines act on S100/A11 to downregulate keratinocyte differentiation. J. Investig. Dermatol. 2008, 128, 2248–2258. [Google Scholar] [CrossRef]
- Rieg, S.; Steffen, H.; Seeber, S.; Humeny, A.; Kalbacher, H.; Dietz, K.; Garbe, C.; Schittek, B. Deficiency of dermcidin-derived antimicrobial peptides in sweat of patients with atopic dermatitis correlates with an impaired innate defense of human skin in vivo. J. Immunol. 2005, 174, 8003–8010. [Google Scholar] [CrossRef]
- Brandner, J.; Haftek, M.; Niessen, C. Adherens Junctions; Desmosomes and Tight Junctions in Epidermal Barrier Function. Open Dermatol. J. 2010, 4, 14–20. [Google Scholar] [CrossRef]
- Niessen, C.M. Tight junctions/adherens junctions: Basic structure and function. J. Investig. Dermatol. 2007, 127, 2525–2532. [Google Scholar] [CrossRef]
- Ebrahiminaseri, A.; Sadeghizadeh, M.; Moshaii, A.; Asgaritarghi, G.; Safari, Z. Combination treatment of dendrosomal nanocurcumin and low-level laser therapy develops proliferation and migration of mouse embryonic fibroblasts and alter TGF-β, VEGF, TNF-α and IL-6 expressions involved in wound healing process. PLoS ONE 2021, 16, e0247098. [Google Scholar] [CrossRef]
- Komine, N.; Ikeda, K.; Tada, K.; Hashimoto, N.; Sugimoto, N.; Tomita, K. Activation of the extracellular signal-regulated kinase signal pathway by light emitting diode irradiation. Lasers Med. Sci. 2010, 25, 531–537. [Google Scholar] [CrossRef]
- Nie, Y.; Xu, X.; Wang, W.; Ma, N.; Lendlein, A. The effects of oscillatory temperature on HaCaT keratinocyte behaviors. Clin. Hemorheol. Microcirc. 2020, 76, 317–327. [Google Scholar] [CrossRef]
- Jung, T.; Höhn, A.; Piazena, H.; Grune, T. Effects of water-filtered infrared A irradiation on human fibroblasts. Free Radic Biol. Med. 2010, 48, 153–160. [Google Scholar] [CrossRef]
- Wiegand, C.; Abel, M.; Ruth, P.; Hipler, U.C. HaCaT keratinocytes in co-culture with Staphylococcus aureus can be protected from bacterial damage by polihexanide. Wound Repair Regen. 2009, 17, 730–738. [Google Scholar] [CrossRef]
- Gaspar, K.; Kukova, G.; Bunemann, E.; Buhren, B.A.; Sonkoly, E.; Szollosi, A.G.; Muller, A.; Savinko, T.; Lauerma, A.I.; Alenius, H.; et al. The chemokine receptor CCR3 participates in tissue remodeling during atopic skin inflammation. J. Dermatol. Sci. 2013, 71, 12–21. [Google Scholar] [CrossRef]
- Ghazi, K.; Deng-Pichon, U.; Warnet, J.M.; Rat, P. Hyaluronan fragments improve wound healing on in vitro cutaneous model through P2X7 purinoreceptor basal activation: Role of molecular weight. PLoS ONE 2012, 7, e48351. [Google Scholar] [CrossRef]
- Volksdorf, T.; Heilmann, J.; Eming, S.A.; Schawjinski, K.; Zorn-Kruppa, M.; Ueck, C.; Vidal-Y-Sy, S.; Windhorst, S.; Jücker, M.; Moll, I.; et al. Tight Junction Proteins Claudin-1 and Occludin Are Important for Cutaneous Wound Healing. Am. J. Pathol. 2017, 187, 1301–1312. [Google Scholar] [CrossRef]
- Rötzer, V.; Hartlieb, E.; Winkler, J.; Walter, E.; Schlipp, A.; Sardy, M.; Spindler, V.; Waschke, J. Desmoglein 3-Dependent Signaling Regulates Keratinocyte Migration and Wound Healing. J. Investig. Dermatol. 2016, 136, 301–310. [Google Scholar] [CrossRef]
- Lee, H.; Park, H.Y.; Jeong, T.S. Pheophorbide a Derivatives Exert Antiwrinkle Effects on UVB-Induced Skin Aging in Human Fibroblasts. Life 2021, 11, 147. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Kwon, T.R.; Kim, J.H.; Lee, B.C.; Oh, C.T.; Im, M.; Hwang, Y.K.; Paik, S.H.; Han, S.; Kim, J.Y.; et al. Anti photoaging and anti oxidative activities of natural killer cell conditioned medium following UV B irradiation of human dermal fibroblasts and a reconstructed skin model. Int. J. Mol. Med. 2019, 44, 1641–1652. [Google Scholar] [CrossRef]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-degrading metalloproteinases in photoaging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef]
- Wlaschek, M.; Thceva-Poor, I.; Naderi, L.; Ma, W.; Schneider, L.A.; Razi-Wolf, Z.; Schüller, J.; Scharfetter-Kochanek, K. Solar UV irradiation and dermal photoaging. J. Pthotochem. Photobiol. B 2001, 63, 41–51. [Google Scholar] [CrossRef]
- Cho, Y.H.; Bahuguna, A.; Kim, H.H.; Kim, D.I.; Kim, H.J.; Yu, J.M.; Jung, H.G.; Jang, J.Y.; Kwak, J.H.; Park, G.H.; et al. Potential effect of compounds isolated from Coffea arabica against UV-B induced skin damage by protecting fibroblast cells. J. Photochem. Photobiol. B 2017, 174, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Nagy, Z.Z.; Hiscott, P.; Seitz, B.; Shlötzer-Schrehardt, U.; Simon, M., Jr.; Süveges, I.; Naumann, G.O. Ultraviolet-B enhances corneal stromal response to 193-nm excimer laser treatment. Ophthalmology 1997, 104, 375–380. [Google Scholar] [CrossRef]
- Borel, N.; Sauer-Durand, A.M.; Hartel, M.; Kuratli, J.; Vaupel, P.; Scherr, N.; Pluschke, G. wIRA: Hyperthermia as a treatment option for intracellular bacteria; with special focus on Chlamydiae and Mycobacteria. Int. J. Hyperth. 2020, 37, 373–383. [Google Scholar] [CrossRef]
- Al-Ahmad, A.; Tennert, C.; Karygianni, L.; Wrbas, K.T.; Hellwig, E.; Altenburger, M.J. Antimicrobial photodynamic therapy using visible light plus water-filtered infrared-A (wIRA). J. Med. Microbiol. 2013, 62, 467–473. [Google Scholar] [CrossRef]
- Al-Ahmad, A.; Walankiewicz, A.; Hellwig, E.; Follo, M.; Tennert, C.; Wittmer, A.; Karygianni, L. Photoinactivation Using Visible Light Plus Water-Filtered Infrared-A (vis+wIRA) and Chlorine e6 (Ce6) Eradicates Planktonic Periodontal Pathogens and Subgingival Biofilms. Front. Microbiol. 2016, 7, 1900. [Google Scholar] [CrossRef]
- Daeschlein, G.; Alborova, J.; Patzelt, A.; Kramer, A.; Lademann, J. Kinetics of physiological skin flora in a suction blister wound model on healthy subjects after treatment with water-filtered infrared-A radiation. Ski. Pharm. Physiol. 2012, 25, 73–77. [Google Scholar] [CrossRef]
- Zöller, N.N.; Kippenberger, S.; Thaçi, D.; Mewes, K.; Spiegel, M.; Sättler, A.; Schultz, M.; Bereiter-Hahn, J.; Kaufmann, R.; Bernd, A. Evaluation of beneficial and adverse effects of glucocorticoids on a newly developed full-thickness skin model. Toxicol. Vitr. 2008, 22, 747–759. [Google Scholar] [CrossRef]
- de Christo Scherer, M.M.; Marques, F.M.; Figueira, M.M.; Peisino, M.C.O.; Schmitt, E.F.P.; Kondratyuk, T.P.; Endringer, D.C.; Scherer, R.; Fronza, M. Wound healing activity of terpinolene and α-phellandrene by attenuating inflammation and oxidative stress in vitro. J. Tissue Viability 2019, 28, 94–99. [Google Scholar] [CrossRef]
- de Moura Sperotto, N.D.; Steffens, L.; Veríssimo, R.M.; Henn, J.G.; Péres, V.F.; Vianna, P.; Chies, J.A.B.; Roehe, A.; Saffi, J.; Moura, D.J. Wound healing and anti-inflammatory activities induced by a Plantago australis hydroethanolic extract standardized in verbascoside. J. Ethnopharmacol. 2018, 225, 178–188. [Google Scholar] [CrossRef]
- Wedler, J.; Daubitz, T.; Schlotterbeck, G.; Butterweck, V. In vitro anti-inflammatory and wound-healing potential of a Phyllostachys edulis leaf extract—identification of isoorientin as an active compound. Planta Med. 2014, 80, 1678–1684. [Google Scholar] [CrossRef]
- Arndt, S.; Unger, P.; Wacker, E.; Shimizu, T.; Heinlin, J.; Li, Y.F.; Thomas, H.M.; Morfill, G.E.; Zimmermann, J.L.; Bosserhoff, A.K.; et al. Cold atmospheric plasma (CAP) changes gene expression of key molecules of the wound healing machinery and improves wound healing in vitro and in vivo. PLoS ONE 2013, 8, e79325. [Google Scholar] [CrossRef]
- Magne, B.; Dedier, M.; Nivet, M.; Coulomb, B.; Banzet, S.; Lataillade, J.J.; Trouillas, M. IL-1β-Primed Mesenchymal Stromal Cells Improve Epidermal Substitute Engraftment and Wound Healing via Matrix Metalloproteinases and Transforming Growth Factor-β1. J. Investig. Dermatol. 2020, 140, 688–698. [Google Scholar] [CrossRef]
- Dinçer, T.; Boz Er, A.B.; Er, İ.; Toraman, B.; Yildiz, G.; Kalay, E. RIPK4 suppresses the TGF-β1 signaling pathway in HaCaT cells. Cell Biol. Int. 2020, 44, 848–860. [Google Scholar] [CrossRef]
- Premarathna, A.D.; Ranahewa, T.H.; Wijesekera, S.K.; Harishchandra, D.L.; Karunathilake, K.J.K.; Waduge, R.N.; Wijesundara, R.R.M.K.K.; Jayasooriya, A.P.; Wijewardana, V.; Rajapakse, R.P.V.J. Preliminary screening of the aqueous extracts of twenty-three different seaweed species in Sri Lanka with in-vitro and in-vivo assays. Heliyon 2020, 6, e03918. [Google Scholar] [CrossRef]
- Toraldo, G.; Bhasin, S.; Bakhit, M.; Guo, W.; Serra, C.; Safer, J.D.; Bhawan, J.; Jasuja, R. Topical androgen antagonism promotes cutaneous wound healing without systemic androgen deprivation by blocking β-catenin nuclear translocation and cross-talk with TGF-β signaling in keratinocytes. Wound Repair Regen. 2012, 20, 61–73. [Google Scholar] [CrossRef]
- Liu, K.; Gao, Z.; Zhou, G.; Zhang, W.; Wu, X.; Liu, W. Characterization of Smad3 knockout mouse derived skin cells. Vitr. Cell Dev. Biol. Anim. 2017, 53, 458–466. [Google Scholar] [CrossRef]
- Varga, J.; Abraham, D. Systemic sclerosis: A prototypic multisystem fibrotic disorder. J. Clin. Investig. 2007, 117, 557–567. [Google Scholar] [CrossRef]
- D’Errico, M.; Lemma, T.; Calcagnile, A.; Proietti De Santis, L.; Dogliotti, E. Cell type and DNA damage specific response of human skin cells to environmental agents. Mutat. Res. 2007, 614, 37–47. [Google Scholar] [CrossRef]
- Tonello, S.; Rizzi, M.; Migliario, M.; Rocchetti, V.; Renò, F. Low concentrations of neutrophil extracellular traps induce proliferation in human keratinocytes via NF-kB activation. J. Derm. Sci. 2017, 88, 110–116. [Google Scholar] [CrossRef]
- Calderwood, S.K.; Ciocca, D.R. Heat shock proteins: Stress proteins with Janus-like properties in cancer. Int. J. Hyperth. 2008, 24, 31–39. [Google Scholar] [CrossRef]
- Favatier, F.; Bornman, L.; Hightower, L.E.; Günther, E.; Polla, B.S. Variation in hsp gene expression and Hsp polymorphism: Do they contribute to differential disease susceptibility and stress tolerance? Cell Stress Chaperones 1997, 2, 141–155. [Google Scholar] [CrossRef]
- Gally, F.; Minor, M.N.; Smith, S.K.; Case, S.R.; Chu, H.W. Heat shock factor 1 protects against lung mycoplasma pneumoniae infection in mice. J. Innate Immun. 2012, 4, 59–68. [Google Scholar] [CrossRef]
- Donnarumma, G.; Paoletti, I.; Buommino, E.; Tufano, M.A.; Baroni, A. Alpha-MSH reduces the internalization of Staphylococcus aureus and down-regulates HSP 70; integrins and cytokine expression in human keratinocyte cell lines. Exp. Dermatol. 2004, 13, 748–754. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grouping | Target Gene | Biological Function |
|---|---|---|
| Inflammatory cytokines | IL1A | |
| IL6 | ||
| CXCL8 | ||
| Growth factors | TGFB1 | |
| PDGFC | ||
| Transcription factors | NFKB1 |
|
| TP53 | ||
| Heat shock proteins | HSP90AA1 |
|
| HSPA1A | ||
| HSPD1 | ||
| Antimicrobial peptides | DEFB1 |
|
| RNASE7 | ||
| S100A7 | ||
| Structural components | DSG1 |
|
| DSG3 | ||
| COL1A1 |
| |
| COL1A3 |
| Target Gene | Ordering IDs/Primer Sequence | Manufacturer | |
|---|---|---|---|
| ACTB | Hs_ACTB_1_SG QuantiTect® Primer Assay | Qiagen, Hilden, Germany | |
| CXCL8 | Hs_CXCL8_1_SG QuantiTect® Primer Assay | ||
| TGFB1 | Hs_TGFB1_1_SG QuantiTect® Primer Assay | ||
| PDGFC | Hs_PDGFC_1_SG QuantiTect® Primer Assay | ||
| RNASE7 | Hs_RNASE7_1_SG QuantiTect® Primer Assay | ||
| COL1A1 | Hs_COL1A1_1_SG QuantiTect® Primer Assay | ||
| COL1A3 | Hs_COL1A3_1_SG QuantiTect® Primer Assay | ||
| HSP90AA1 | QT01002603 | ||
| HSPA1A | QT01002568 | ||
| HSPD1 | QT00018970 | ||
| NFKB1 | QT00063791 | ||
| TP53 | QT00060235 | ||
| DEFB1 | QT00008302 | ||
| IL1A | Fw Rev | 5′-CGCCAATGACTCAGAGGAAGA-3′ 5′-AGGGCGTCATTCAGGATGAA-3′ | Eurofins Genomics, Ebersberg, Germany |
| IL6 | Fw Rev | 5′-CCACCGGGAACGAAAGAGAA-3′ 5′-GAGAAGGCAACTGGACCGAA-3′ | |
| S100A7 | Fw Rev | 5′-GTCCAAACACACACATCTCACT-3′ 5′-TCATCATCGTCAGCAGGCTT-3′ | |
| DSG1 | Fw Rev | 5′-TCCCCACATTTCGGCACTAC-3′ 5′-GCCCAGAGGATCGAGAATAGG-3′ | |
| DSG3 | Fw Rev | 5′-GTCAGAACAATCGGTGTGAGATG-3′ 5′-TGCGGCCTGCCATACCT-3′ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiegand, C.; Hipler, U.-C.; Elsner, P.; Tittelbach, J. Keratinocyte and Fibroblast Wound Healing In Vitro Is Repressed by Non-Optimal Conditions but the Reparative Potential Can Be Improved by Water-Filtered Infrared A. Biomedicines 2021, 9, 1802. https://doi.org/10.3390/biomedicines9121802
Wiegand C, Hipler U-C, Elsner P, Tittelbach J. Keratinocyte and Fibroblast Wound Healing In Vitro Is Repressed by Non-Optimal Conditions but the Reparative Potential Can Be Improved by Water-Filtered Infrared A. Biomedicines. 2021; 9(12):1802. https://doi.org/10.3390/biomedicines9121802
Chicago/Turabian StyleWiegand, Cornelia, Uta-Christina Hipler, Peter Elsner, and Jörg Tittelbach. 2021. "Keratinocyte and Fibroblast Wound Healing In Vitro Is Repressed by Non-Optimal Conditions but the Reparative Potential Can Be Improved by Water-Filtered Infrared A" Biomedicines 9, no. 12: 1802. https://doi.org/10.3390/biomedicines9121802
APA StyleWiegand, C., Hipler, U.-C., Elsner, P., & Tittelbach, J. (2021). Keratinocyte and Fibroblast Wound Healing In Vitro Is Repressed by Non-Optimal Conditions but the Reparative Potential Can Be Improved by Water-Filtered Infrared A. Biomedicines, 9(12), 1802. https://doi.org/10.3390/biomedicines9121802