
Sex-Dependent Effects of Eicosapentaenoic Acid on Hepatic Steatosis in UCP1 Knockout Mice

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Studies
2.2. Liver Fatty Acid Composition
2.3. Liver Histology
2.4. Liver Triglycerides
2.5. RNA Isolation and Quantitative Real-Time PCR
2.6. Immunoblotting
2.7. Statistical Analyses
3. Results
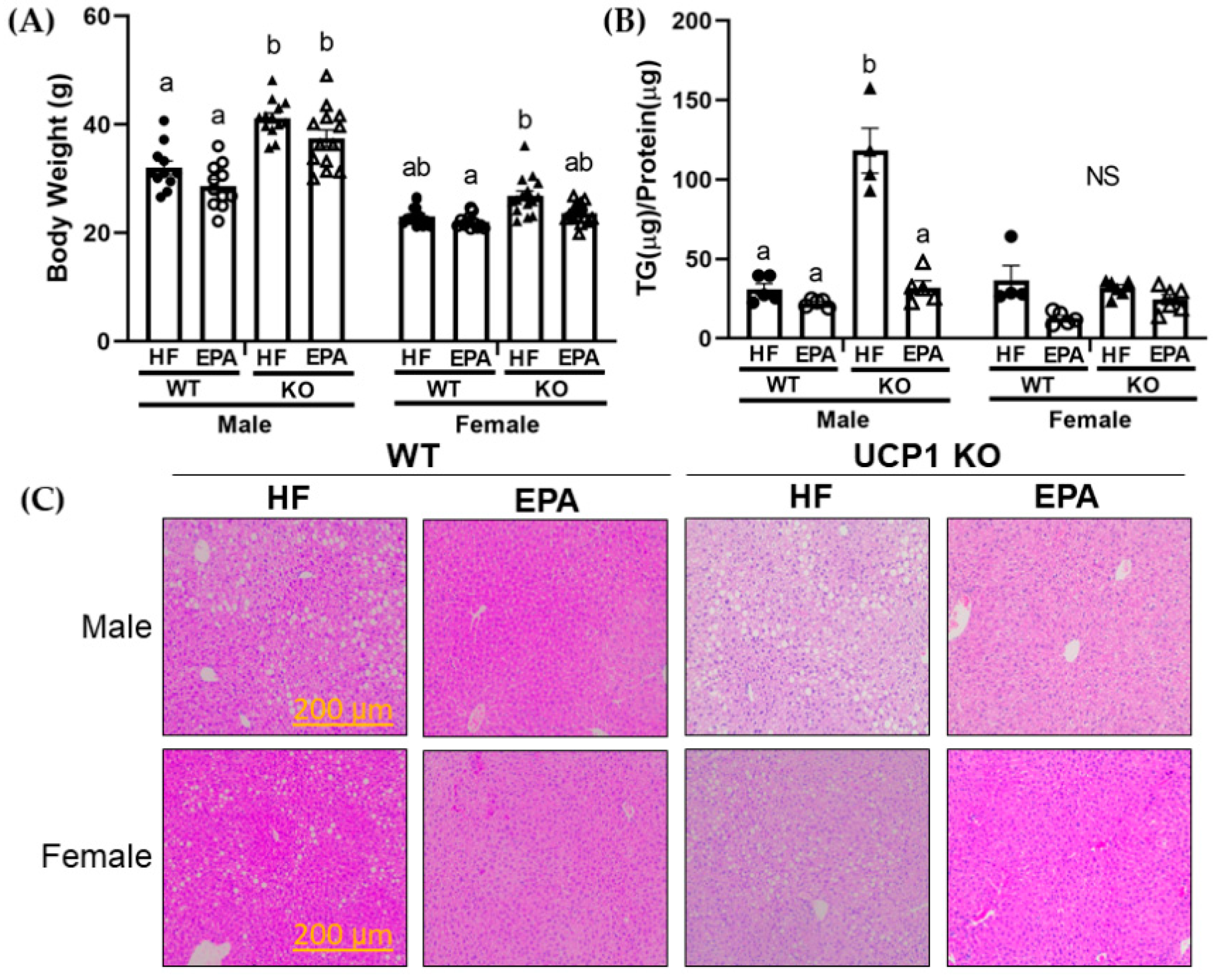
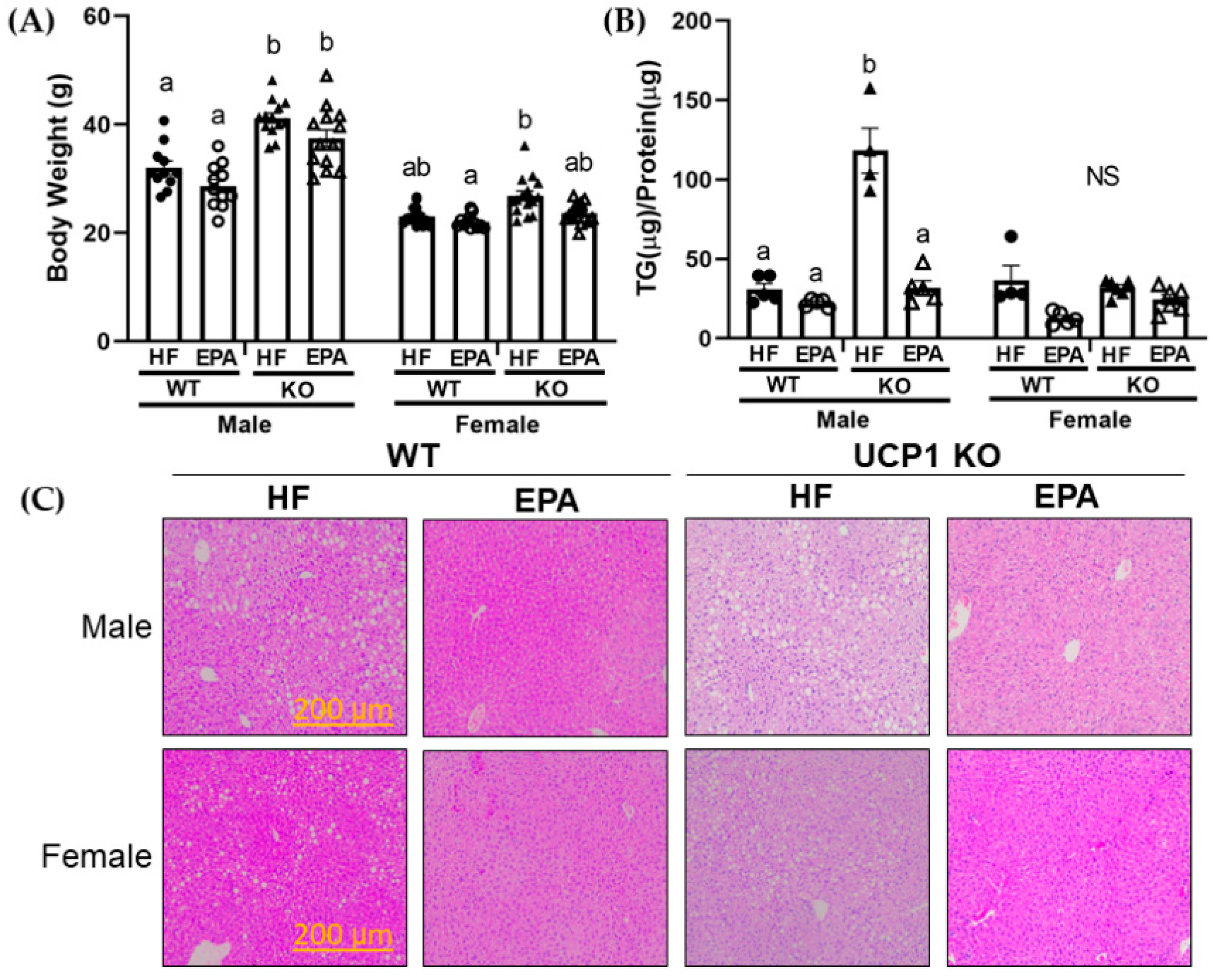
3.1. UCP1 Deficiency Sex-dependently Affects Body Weight
3.2. Dietary EPA Increases Hepatic EPA Content
3.3. EPA Reduces Hepatic Triglycerides
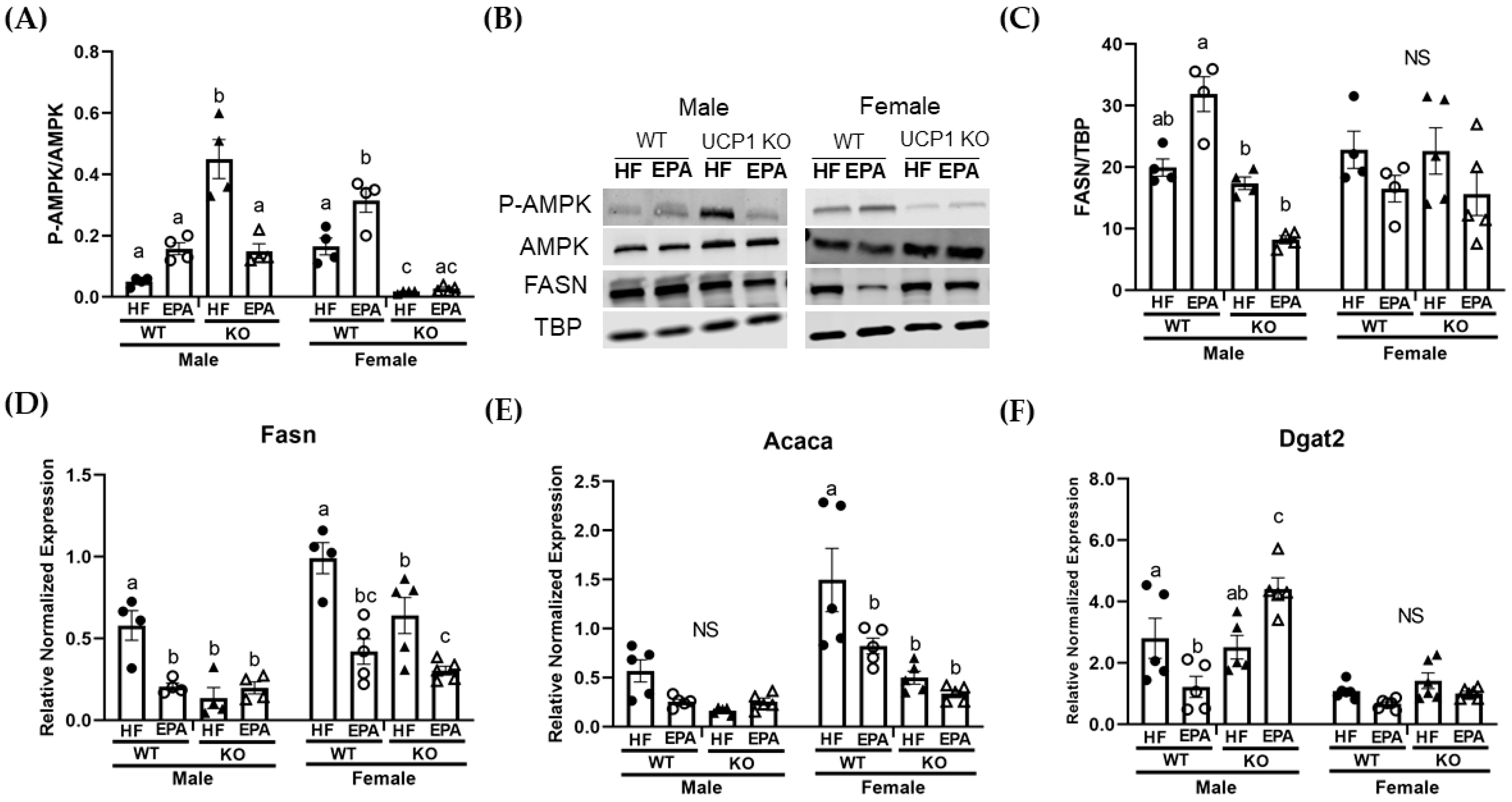
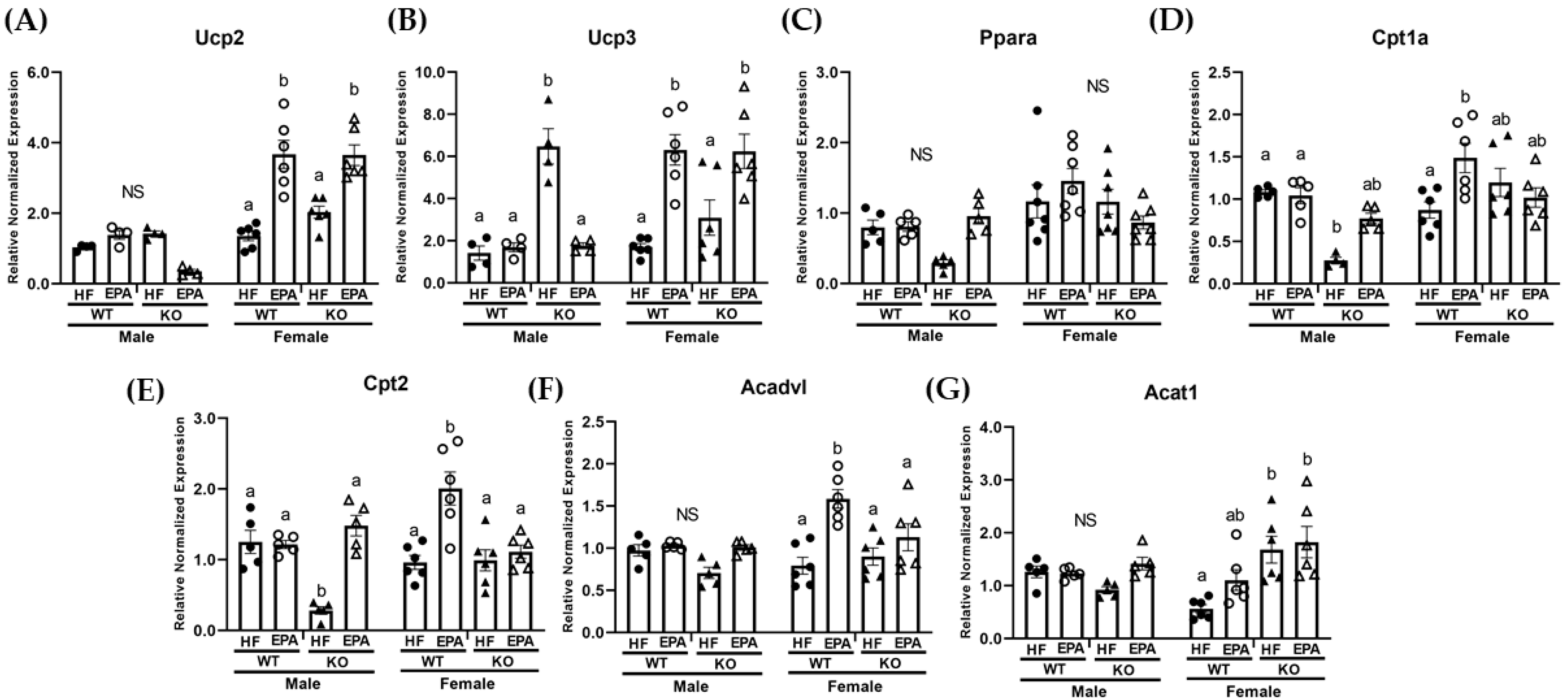
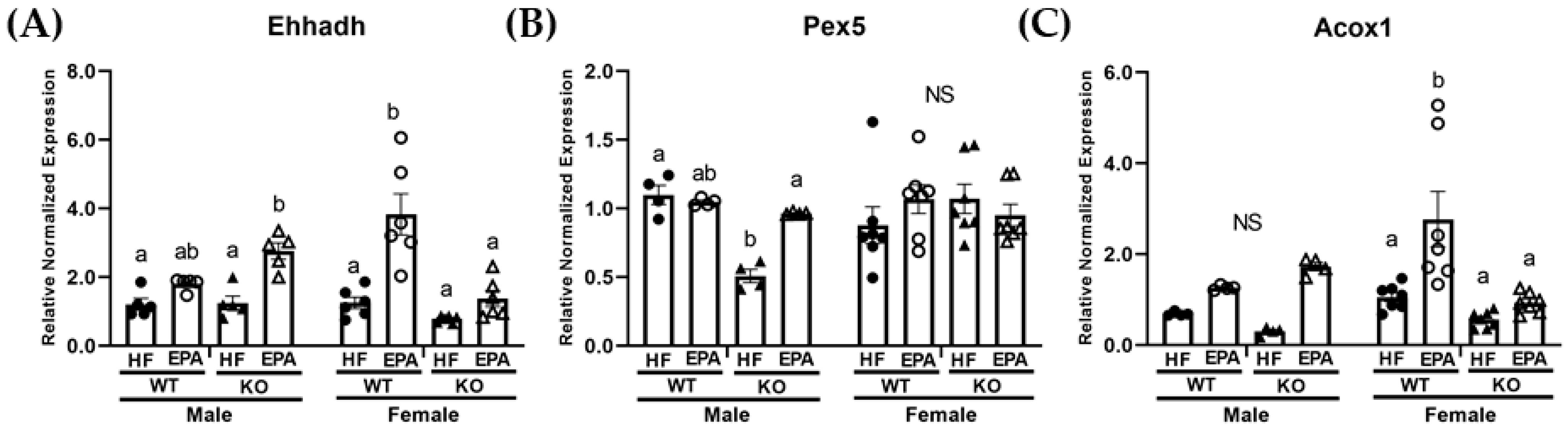
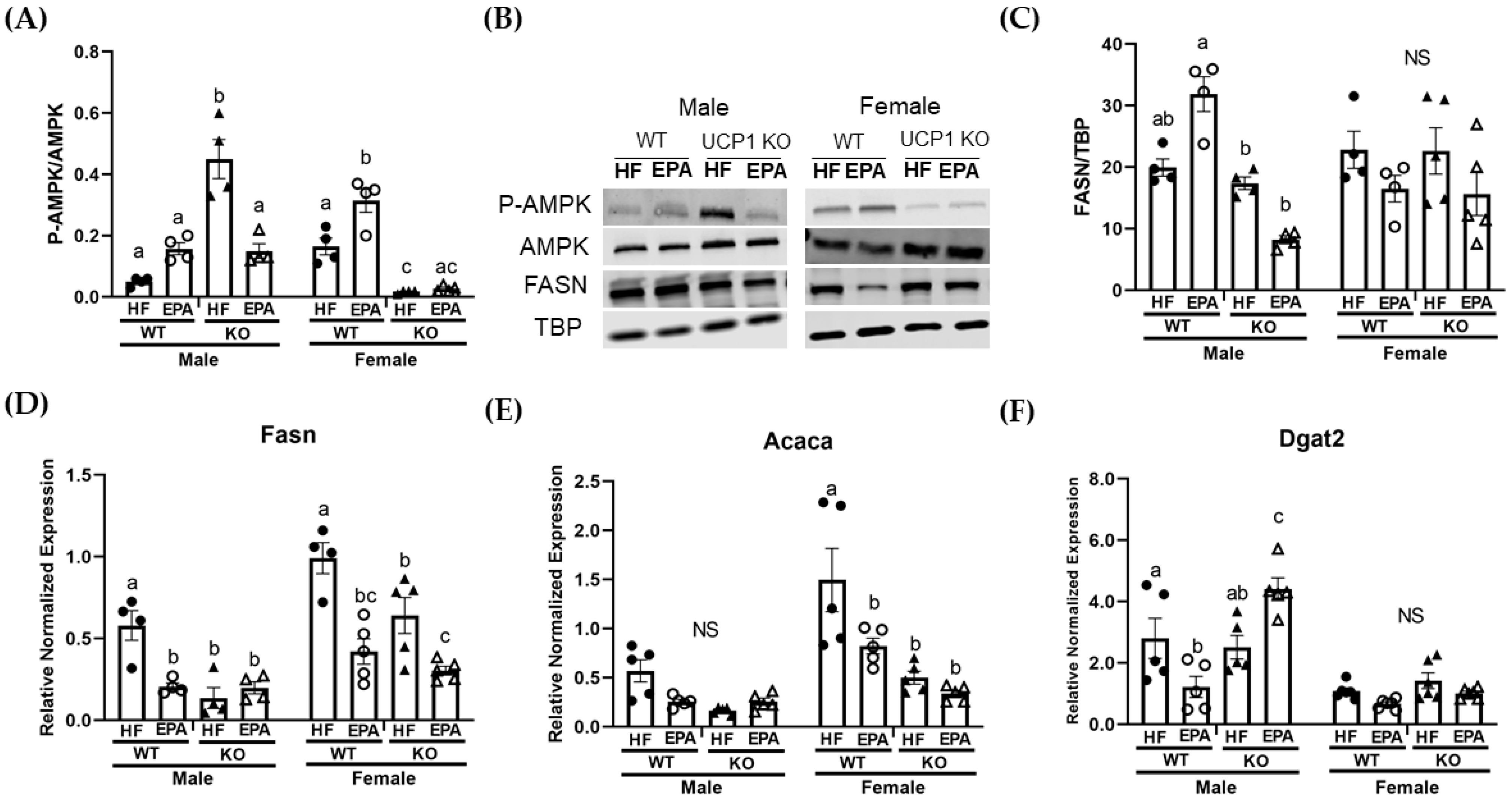
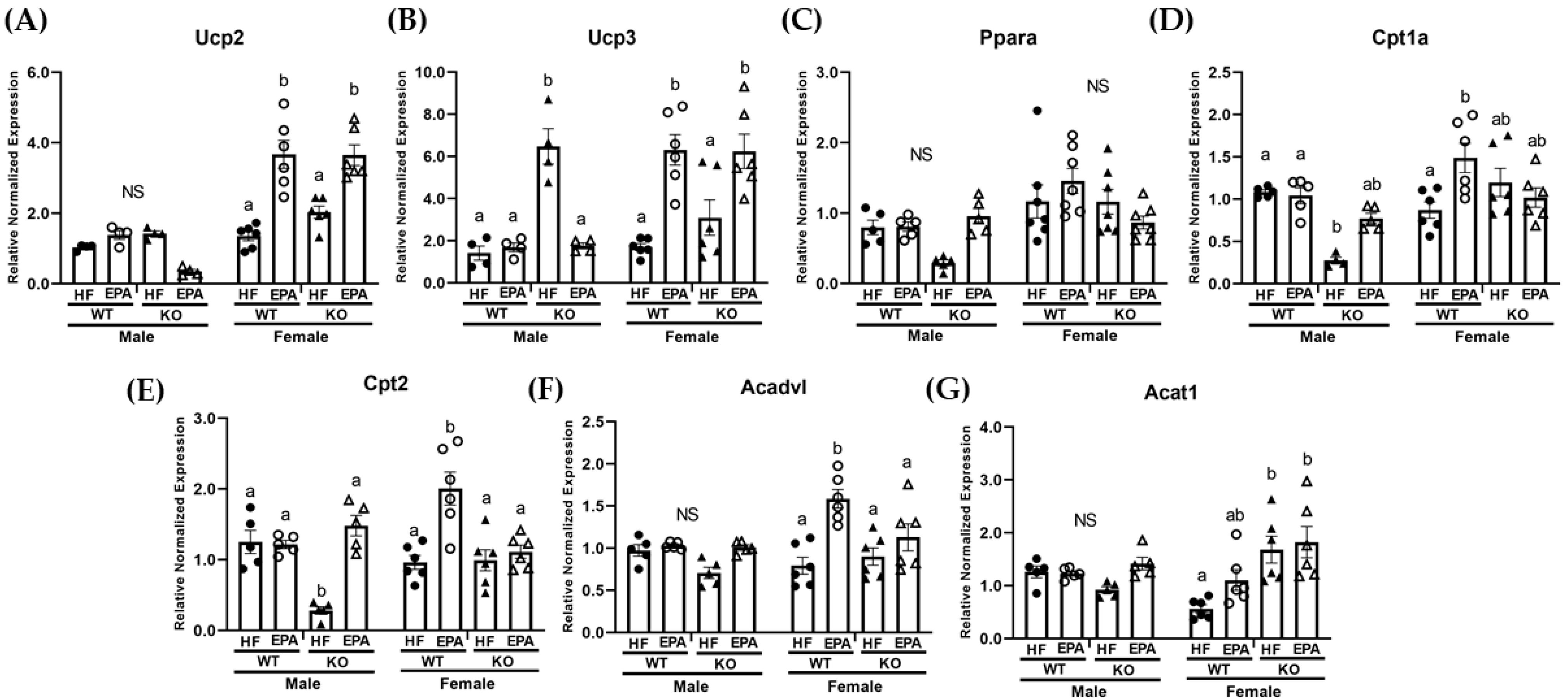
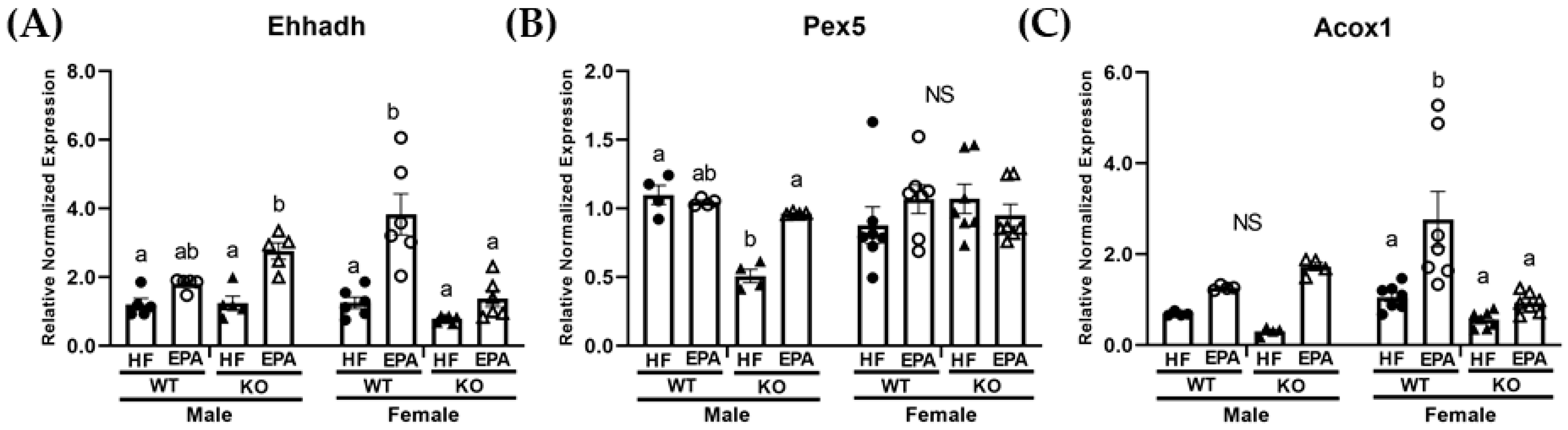
3.4. EPA Rescues Hepatic Triglyceride Load by Upregulating Fat Oxidation
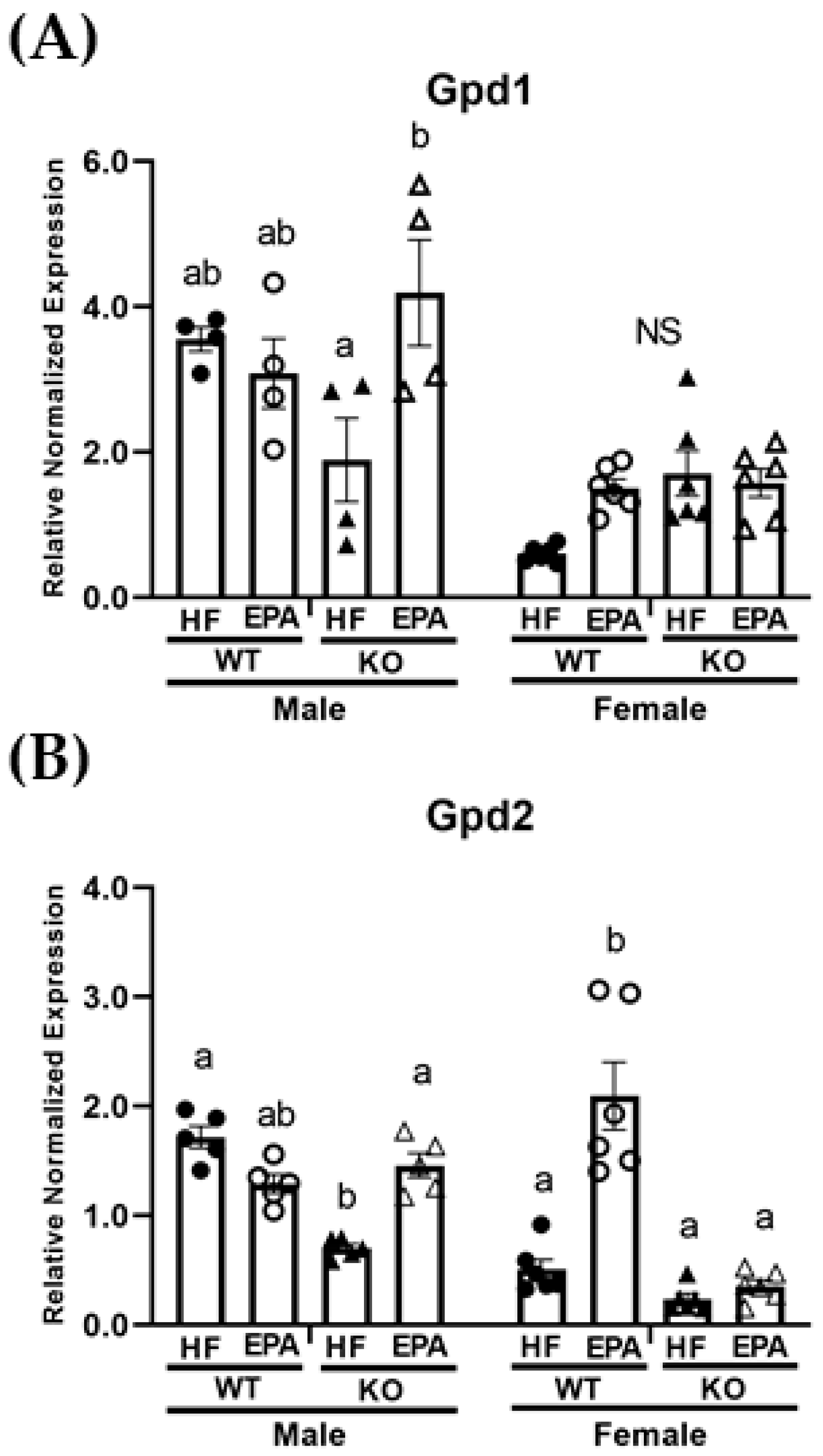
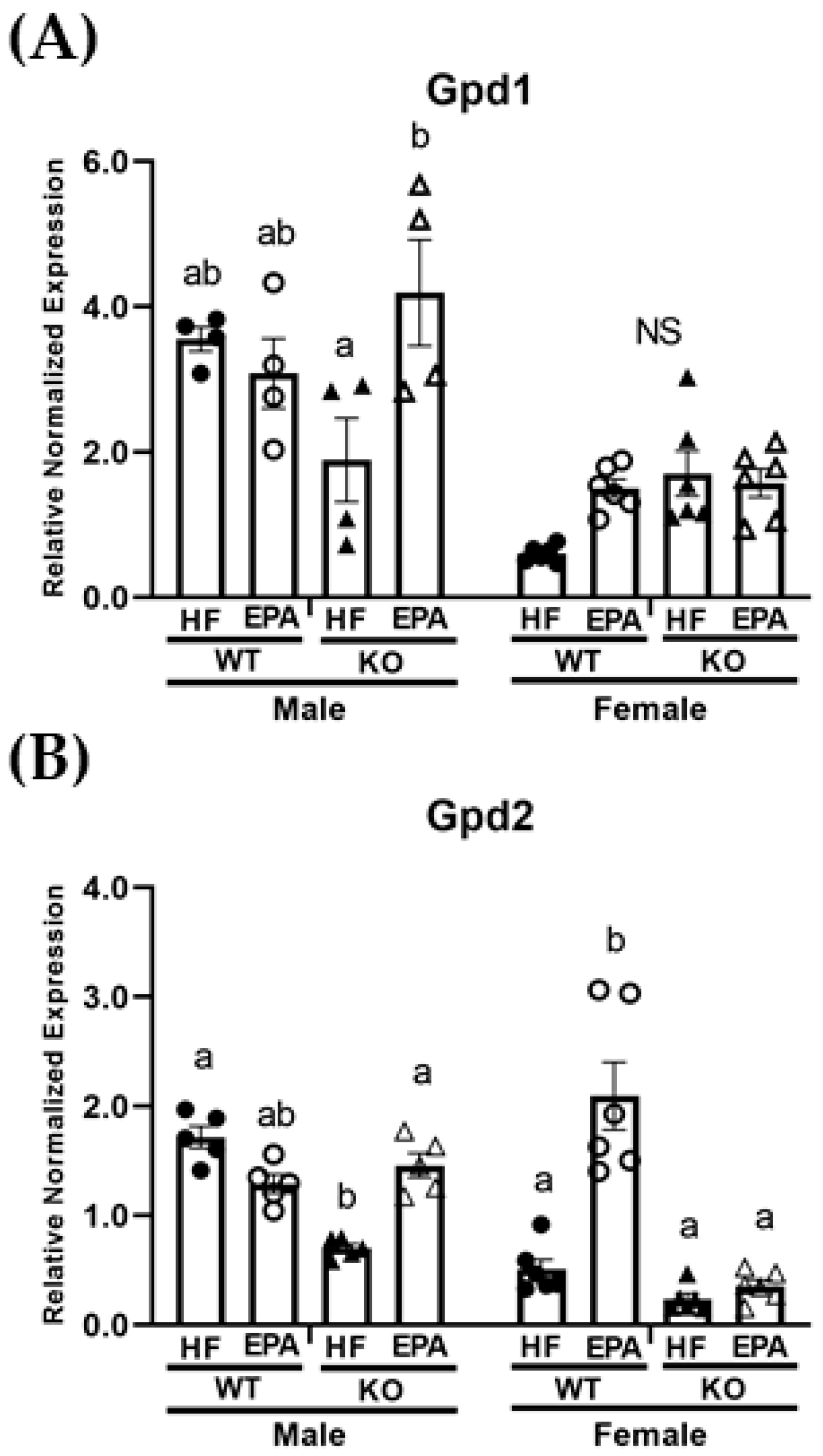
3.5. EPA Alters Metabolic Futile Cycles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Obesity and nonalcoholic fatty liver disease: From pathophysiology to therapeutics. Metabolism 2018, 92, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, J.; Viggiano, T.R.; McGill, D.B.; Oh, B.J. Nonalcoholic steatohepatitis: Mayo Clinic experiences with a hitherto unnamed disease. Mayo Clin. Proc. 1980, 55, 434–438. [Google Scholar]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Koenig, A.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2015, 64, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Golabi, P.; Paik, J.; Hwang, J.; Wang, S.; Lee, H.M.; Younossi, Z.M. Prevalence and outcomes of non-alcoholic fatty liver disease (NAFLD) among Asian American adults in the United States. Liver Int. 2018, 39, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Cannon, B.; Nedergaard, J. Brown Adipose Tissue: Function and Physiological Significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Shen, H.; Jiang, L.; Lin, J.D.; Omary, M.B.; Rui, L. Brown fat activation mitigates alcohol-induced liver steatosis and injury in mice. J. Clin. Investig. 2019, 129, 2305–2317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholls, D.G.; Locke, R.M. Thermogenic mechanisms in brown fat. Physiol. Rev. 1984, 64, 1–64. [Google Scholar] [CrossRef] [Green Version]
- Kopecky, J.; Clarke, G.; Enerbäck, S.; Spiegelman, B.; Kozak, L.P. Expression of the mitochondrial uncoupling protein gene from the aP2 gene promoter prevents genetic obesity. J. Clin. Investig. 1995, 96, 2914–2923. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Nolte, L.A.; Ju, J.-S.; Han, D.H.; Coleman, T.; Holloszy, J.O.; Semenkovich, C.F. Skeletal muscle respiratory uncoupling prevents diet-induced obesity and insulin resistance in mice. Nat. Med. 2000, 6, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Winn, N.; Vieira-Potter, V.J.; Gastecki, M.L.; Welly, R.J.; Scroggins, R.J.; Zidon, T.M.; Gaines, T.L.; Woodford, M.L.; Karasseva, N.G.; Kanaley, J.A.; et al. Loss of UCP1 exacerbates Western diet-induced glycemic dysregulation independent of changes in body weight in female mice. Am. J. Physiol. Integr. Comp. Physiol. 2017, 312, R74–R84. [Google Scholar] [CrossRef] [PubMed]
- Ishigaki, Y.; Katagiri, H.; Yamada, T.; Ogihara, T.; Imai, J.; Uno, K.; Hasegawa, Y.; Gao, J.; Ishihara, H.; Shimosegawa, T.; et al. Dissipating Excess Energy Stored in the Liver Is a Potential Treatment Strategy for Diabetes Associated with Obesity. Diabetes 2005, 54, 322–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couillard, C.; Bergeron, N.; Prud’homme, D.; Bergeron, J.; Tremblay, A.; Bouchard, C.; Mauriege, P.; Després, J.-P. Gender difference in postprandial lipemia: Importance of visceral adipose tissue accumulation. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 2448–2455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittendorfer, B.; Magkos, F.; Fabbrini, E.; Mohammed, B.S.; Klein, S. Relationship Between Body Fat Mass and Free Fatty Acid Kinetics in Men and Women. Obesity 2009, 17, 1872–1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalupahana, N.S.; Claycombe, K.; Newman, S.; Stewart, T.; Siriwardhana, N.; Matthan, N.; Lichtenstein, A.H.; Moustaid-Moussa, N. Eicosapentaenoic Acid Prevents and Reverses Insulin Resistance in High-Fat Diet-Induced Obese Mice via Modulation of Adipose Tissue Inflammation. J. Nutr. 2010, 140, 1915–1922. [Google Scholar] [CrossRef] [Green Version]
- Pahlavani, M.; Ramalingam, L.; Miller, E.K.; Scoggin, S.; Menikdiwela, K.R.; Kalupahana, N.S.; Festuccia, W.; Moustaid-Moussa, N. Eicosapentaenoic Acid Reduces Adiposity, Glucose Intolerance and Increases Oxygen Consumption Independently of Uncoupling Protein 1. Mol. Nutr. Food Res. 2019, 63, e1800821. [Google Scholar] [CrossRef]
- Dongiovanni, P.; Lanti, C.; Riso, P.; Valenti, L. Nutritional therapy for nonalcoholic fatty liver disease. J. Nutr. Biochem. 2015, 29, 1–11. [Google Scholar] [CrossRef]
- Albracht-Schulte, K.; Kalupahana, N.S.; Ramalingam, L.; Wang, S.; Rahman, S.M.; Robert-McComb, J.; Moustaid-Moussa, N. Omega-3 fatty acids in obesity and metabolic syndrome: A mechanistic update. J. Nutr. Biochem. 2018, 58, 1–16. [Google Scholar] [CrossRef]
- Albracht-Schulte, K.; Gonzalez, S.; Jackson, A.; Wilson, S.; Ramalingam, L.; Kalupahana, N.S.; Moustaid-Moussa, N. Eicosapentaenoic Acid Improves Hepatic Metabolism and Reduces Inflammation Independent of Obesity in High-Fat-Fed Mice and in HepG2 Cells. Nutrients 2019, 11, 599. [Google Scholar] [CrossRef] [Green Version]
- Balogun, K.; Randunu, R.; Cheema, S. The effect of dietary omega-3 polyunsaturated fatty acids on plasma lipids and lipoproteins of C57BL/6 mice is age and sex specific. Prostaglandins, Leukot. Essent. Fat. Acids 2014, 91, 39–47. [Google Scholar] [CrossRef]
- Pawlosky, R.J.; Hibbeln, J.R.; Novotny, J.A.; Salem, N. Physiological compartmental analysis of α-linolenic acid metabolism in adult humans. J. Lipid Res. 2001, 42, 1257–1265. [Google Scholar] [CrossRef]
- Pahlavani, M.; Razafimanjato, F.; Ramalingam, L.; Kalupahana, N.S.; Moussa, H.; Scoggin, S.; Moustaid-Moussa, N. Eicosapentaenoic acid regulates brown adipose tissue metabolism in high-fat-fed mice and in clonal brown adipocytes. J. Nutr. Biochem. 2017, 39, 101–109. [Google Scholar] [CrossRef]
- Ravn-Haren, G.; Kopp, T.I.; Outzen, M.; Olsen, A.; Voge, U. Nutrient-Gene Interactions. Curr. Dev. Nutr. 2018, 2, nzy044. [Google Scholar] [CrossRef]
- Miller, E.K.; Pahlavani, M.; Ramalingam, L.; Scoggin, S.; Moustaid-Moussa, N. Uncoupling Protein 1-Independent Effects of Eicosapentaenoic Acid in Brown Adipose Tissue of Diet-Induced Obese Female Mice. J. Nutr. Biochem. 2021, 98, 108819. [Google Scholar] [CrossRef] [PubMed]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A direct method for fatty acid methyl ester synthesis: Application to wet meat tissues, oils, and feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef] [Green Version]
- Wijayatunga, N.N.; Sams, V.G.; Dawson, J.A.; Mancini, M.L.; Mancini, G.J.; Moustaid-Moussa, N. Roux-en-Y gastric bypass surgery alters serum metabolites and fatty acids in patients with morbid obesity. Diabetes/Metab. Res. Rev. 2018, 34, e3045. [Google Scholar] [CrossRef] [Green Version]
- Serviddio, G.; Bellanti, F.; Tamborra, R.; Rollo, T.; Capitanio, N.; Romano, A.D.; Sastre, J.; Vendemiale, G.; Altomare, E. Uncoupling protein-2 (UCP2) induces mitochondrial proton leak and increases susceptibility of non-alcoholic steatohepatitis (NASH) liver to ischaemia-reperfusion injury. Gut 2008, 57, 957–965. [Google Scholar] [CrossRef]
- Feldmann, H.M.; Golozoubova, V.; Cannon, B.; Nedergaard, J. UCP1 Ablation Induces Obesity and Abolishes Diet-Induced Thermogenesis in Mice Exempt from Thermal Stress by Living at Thermoneutrality. Cell Metab. 2009, 9, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, T.E.; Castro, É.; Belchior, T.; Andrade, M.L.; Filho, A.D.B.C.; Peixoto, S.; Moreno, M.F.; Ortiz-Silva, M.; Moreira, R.; Inague, A.; et al. Fish Oil Protects Wild Type and Uncoupling Protein 1-Deficient Mice from Obesity and Glucose Intolerance by Increasing Energy Expenditure. Mol. Nutr. Food Res. 2019, 63, e1800813. [Google Scholar] [CrossRef]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and nonalcoholic fatty liver disease: Biochemical, metabolic, and clinical implications. Hepatology 2009, 51, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Serviddio, G.; Sastre, J.; Bellanti, F.; Viña, J.; Vendemiale, G.; Altomare, E. Mitochondrial involvement in non-alcoholic steatohepatitis. Mol. Asp. Med. 2008, 29, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-P.; Liang, L.; Wang, C.-L.; Fu, J.-F.; Liu, P.-N.; Lv, L.-Q.; Zhu, Y.-M. Association between UCP3 gene polymorphisms and nonalcoholic fatty liver disease in Chinese children. World J. Gastroenterol. 2013, 19, 5897–5903. [Google Scholar] [CrossRef]
- Aller, R.; De Luis, D.A.; Izaola, O.; Sagrado, M.G.; Conde, R.; Alvarez, T.; Pacheco, D.; Velasco, M.C. Role of -55CT polymorphism of UCP3 gene on non alcoholic fatty liver disease and insulin resistance in patients with obesity. 2010, 25. [Google Scholar]
- Dawson, A. Oxidation of cytosolic NADH formed during aerobic metabolism in mammalian cells. Trends Biochem. Sci. 1979, 4, 171–176. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. National Center for Chronic Disease Prevention and Health Promotion, D.o.N., Physical Activity, and Obesity. Data, Trend and Maps. Available online: https://www.cdc.gov/nccdphp/dnpao/data-trends-maps/index.html. (accessed on 16 February 2021).
- Chalasani, N.; Younossi, Z.; LaVine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases. Hepatology 2017, 67, 328–357. [Google Scholar] [CrossRef]
- Ray, K. NAFLD—the next global epidemic. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torno, C.; Staats, S.; Michl, S.C.; De Pascual-Teresa, S.; Izquierdo, M.; Rimbach, G.; Schulz, C. Fatty Acid Composition and Fatty Acid Associated Gene-Expression in Gilthead Sea Bream (Sparus aurata) are Affected by Low-Fish Oil Diets, Dietary Resveratrol, and Holding Temperature. Mar. Drugs 2018, 16, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, M.D.; Esteves, T.C. Physiological functions of the mitochondrial uncoupling proteins UCP2 and UCP3. Cell Metab. 2005, 2, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cioffi, F.; Senese, R.; De Lange, P.; Goglia, F.; Lanni, A.; Lombardi, A. Uncoupling proteins: A complex journey to function discovery. BioFactors 2009, 35, 417–428. [Google Scholar] [CrossRef]
- Léveillé, M.; Estall, J.L. Mitochondrial Dysfunction in the Transition from NASH to HCC. Metabolites 2019, 9, 233. [Google Scholar] [CrossRef] [Green Version]
- Camara, Y.; Mampel, T.; Armengol, J.; Villarroya, F.; Dejean, L. UCP3 Expression in Liver Modulates Gene Expression and Oxidative Metabolism in Response to Fatty Acids, and Sensitizes Mitochondria to Permeability Transition. Cell. Physiol. Biochem. 2009, 24, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Okada, L.S.D.R.R.; Oliveira, C.P.; Stefano, J.T.; Nogueira, M.A.; da Silva, I.D.C.G.; Cordeiro, F.; Alves, V.A.F.; Torrinhas, R.S.; Carrilho, F.J.; Puri, P.; et al. Omega-3 PUFA modulate lipogenesis, ER stress, and mitochondrial dysfunction markers in NASH—Proteomic and lipidomic insight. Clin. Nutr. 2018, 37, 1474–1484. [Google Scholar] [CrossRef] [PubMed]
- Žáčková, M.; Škobisová, E.; Urbánková, E.; Ježek, P. Activating ω-6 Polyunsaturated Fatty Acids and Inhibitory Purine Nucleotides Are High Affinity Ligands for Novel Mitochondrial Uncoupling Proteins UCP2 and UCP3. J. Biol. Chem. 2003, 278, 20761–20769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrauwen, P.; Hardie, G.; Roorda, B.; Clapham, J.C.; Abuin, A.; Thomason-Hughes, M.; Green, K.; Frederik, P.M.; Hesselink, M.K.C. Improved glucose homeostasis in mice overexpressing human UCP3: A role for AMP-kinase? Int. J. Obes. 2004, 28, 824–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, X.; He, X.; Shi, Z.; Li, C.; Guo, F.; Li, S.; Li, Y.; Na, L.; Sun, C. Ursolic acid increases energy expenditure through enhancing free fatty acid uptake and β-oxidation via an UCP3/AMPK-dependent pathway in skeletal muscle. Mol. Nutr. Food Res. 2015, 59, 1491–1503. [Google Scholar] [CrossRef]
- Gupta, A.P.; Singh, P.; Garg, R.; Valicherla, G.R.; Riyazuddin, M.; Syed, A.A.; Hossain, Z.; Gayen, J.R. Pancreastatin inhibitor activates AMPK pathway via GRP78 and ameliorates dexamethasone induced fatty liver disease in C57BL/6 mice. Biomed. Pharmacother. 2019, 116, 108959. [Google Scholar] [CrossRef]
- Winder, W.W.; Wilson, H.A.; Hardie, D.G.; Rasmussen, B.B.; Hutber, C.A.; Call, G.B.; Clayton, R.D.; Conley, L.M.; Yoon, S.; Zhou, B. Phosphorylation of rat muscle acetyl-CoA carboxylase by AMP-activated protein kinase and protein kinase A. J. Appl. Physiol. 1997, 82, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Rashid, A.; Wu, T.-C.; Huang, C.-C.; Chen, C.-H.; Lin, H.Z.; Yang, S.Q.; Lee, F.Y.; Diehl, A.M. Mitochondrial proteins that regulate apoptosis and necrosis are induced in mouse fatty liver. Hepatology 1999, 29, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Echtay, K.S.; Roussel, D.; St-Pierre, J.; Jekabsons, M.B.; Cadenas, S.; Stuart, J.A.; Harper, J.A.; Roebuck, S.J.; Morrison, A.; Pickering, S.; et al. Superoxide activates mitochondrial uncoupling proteins. Nature 2002, 415, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Baffy, G.; Zhang, C.-Y.; Glickman, J.N.; Lowell, B.B. Obesity-related fatty liver is unchanged in mice deficient for mitochondrial uncoupling protein 2. Hepatology 2002, 35, 753–761. [Google Scholar] [CrossRef]
- Fülöp, P.; Derdak, Z.; Sheets, A.; Sabo, E.; Berthiaume, E.P.; Resnick, M.B.; Wands, J.R.; Paragh, G.; Baffy, G. Lack of UCP2 reduces fas-mediated liver injury inob/ob mice and reveals importance of cell-specific UCP2 expression. Hepatology 2006, 44, 592–601. [Google Scholar] [CrossRef] [Green Version]
- Hsu, H.-C.; Chen, C.-Y.; Chen, M.-F. N-3 polyunsaturated fatty acids decrease levels of doxorubicin-induced reactive oxygen species in cardiomyocytes—Involvement of uncoupling protein UCP2. J. Biomed. Sci. 2014, 21, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Borsonelo, E.C.; Vieira, L.; Galduróz, J.C.F. The influence of the polyunsaturated fatty acids on body weight and anxiolytic-like behavior in female rats. Nutr. Neurosci. 2013, 16, 2–5. [Google Scholar] [CrossRef]
- Tsuboyama-Kasaoka, N.; Sano, K.; Shozawa, C.; Osaka, T.; Ezaki, O. Studies of UCP2 transgenic and knockout mice reveal that liver UCP2 is not essential for the antiobesity effects of fish oil. Am. J. Physiol. Metab. 2008, 294, E600–E606. [Google Scholar] [CrossRef]
- Fischer, A.; Cannon, B.; Nedergaard, J. Optimal housing temperatures for mice to mimic the thermal environment of humans: An experimental study. Mol. Metab. 2017, 7, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.R.; Keijer, J. Not so hot: Optimal housing temperatures for mice to mimic the thermal environment of humans. Mol. Metab. 2013, 2, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Eto, K.; Tsubamoto, Y.; Terauchi, Y.; Sugiyama, T.; Kishimoto, T.; Takahashi, N.; Yamauchi, N.; Kubota, N.; Murayama, S.; Aizawa, T.; et al. Role of NADH Shuttle System in Glucose-Induced Activation of Mitochondrial Metabolism and Insulin Secretion. Science 1999, 283, 981–985. [Google Scholar] [CrossRef]
- Zheng, Y.; Qu, H.; Xiong, X.; Wang, Y.; Liu, X.; Zhang, L.; Liao, X.; Liao, Q.; Sun, Z.; Ouyang, Q.; et al. Deficiency of Mitochondrial Glycerol 3-Phosphate Dehydrogenase Contributes to Hepatic Steatosis. Hepatology 2019, 70, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Pahlavani, M.; Ramalingam, L.; Miller, E.K.; Davis, H.; Scoggin, S.; Moustaid-Moussa, N. Discordant Dose-Dependent Metabolic Effects of Eicosapentanoic Acid in Diet-Induced Obese Mice. Nutrients 2020, 12, 1342. [Google Scholar] [CrossRef]
- Dieckmann, S.; Strohmeyer, A.; Willershäuser, M.; Maurer, S.; Wurst, W.; Marschall, S.; de Angelis, M.H.; Kühn, R.; Worthmann, A.; Fuh, M.M.; et al. Susceptibility to diet induced obesity at thermoneutral conditions is independent of UCP1. bioRxiv 2021. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Effects | Interactions | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Statistic | Sex (S) | Genotype (G) | Diet (D) | S × G | S × D | G × D | S × G × D |
| Final Body Weight | p F (1, 96) | <0.0001 259.3 | <0.0001 73.57 | <0.0001 17.14 | <0.0001 21.58 | 0.30 1.09 | 0.35 0.89 | 0.51 0.44 |
| Liver Triglycerides | p F (1, 32) | <0.0001 40.96 | <0.0001 46.23 | <0.0001 68.59 | <0.0001 35.23 | 0.0002 17.27 | 0.0003 16.01 | <0.0001 38.00 |
| P-AMPK/AMPK | p F (1, 24) | 0.003 10.90 | 0.62 0.25 | 0.71 0.14 | <0.0001 93.56 | 0.0004 16.98 | <0.0001 40.96 | 0.005 9.78 |
| FASN/TBP | p F (1, 26) | 0.98 0.0006 | 0.002 12.30 | 0.18 1.86 | 0.003 10.47 | 0.048 4.27 | 0.01 7.85 | 0.01 6.88 |
| Mitochondrial Uncoupling | ||||||||
| Ucp2 | p F (1, 32) | <0.0001 89.90 | 0.98 0.0006 | <0.0001 21.99 | 0.07 3.58 | <0.0001 45.87 | 0.004 9.57 | 0.30 1.12 |
| Ucp3 | p F (1, 32) | 0.003 10.19 | 0.002 11.77 | 0.09 3.11 | 0.05 4.08 | <0.0001 42.02 | 0.002 11.79 | 0.07 3.52 |
| FA Synthesis | ||||||||
| Acaca | p F (1, 32) | <0.0001 27.9 | 0.0001 27.18 | 0.006 8.59 | 0.006 8.87 | 0.01 2.88 | 0.02 6.41 | 0.78 0.09 |
| Fasn | p F (1, 27) | <0.0001 34.25 | 0.0002 18.86 | <0.0001 33.21 | 0.94 0.01 | 0.009 7.94 | 0.004 10.02 | 0.35 0.91 |
| Dgat2 | p F (1, 36) | <0.0001 58.66 | 0.0003 16.41 | 0.56 0.34 | 0.02 6.30 | 0.21 1.65 | 0.0004 15.55 | 0.0004 15.41 |
| FA Oxidation | ||||||||
| Pparα | p F (1, 40) | 0.0003 15.99 | 0.04 4.48 | 0.14 2.22 | 0.60 0.28 | 0.14 2.29 | 0.88 0.02 | 0.009 7.56 |
| Cpt1a | p F (1, 36) | 0.0002 16.67 | 0.001 12.62 | 0.01 6.79 | 0.01 7.40 | 0.96 0.002 | 0.46 0.55 | 0.0004 15.10 |
| Cpt2 | p F (1, 36) | 0.04 4.41 | 0.0004 15.40 | <0.0001 33.86 | 0.69 0.16 | 0.98 0.0004 | 0.44 0.60 | <0.0001 29.17 |
| Acadvl | p F (1, 36) | 0.02 5.95 | 0.03 5.09 | <0.0001 23.89 | 0.84 0.04 | 0.03 5.50 | 0.27 1.28 | 0.007 8.17 |
| Acat1 | p F (1, 36) | 0.51 0.45 | 0.002 10.65 | 0.03 5.01 | 0.0005 14.90 | 0.67 0.18 | 0.80 0.06 | 0.08 3.21 |
| Ehhadh | p F (1, 36) | 0.79 0.07 | 0.03 5.68 | <0.0001 42.84 | <0.0001 23.72 | 0.20 1.73 | 0.19 1.77 | 0.0009 12.99 |
| Pex5 | p F (1, 36) | 0.24 1.38 | 0.05 4.00 | 0.12 2.52 | 0.02 6.58 | 0.27 1.25 | 0.51 0.44 | 0.01 7.52 |
| Acox1 | p F (1, 36) | 0.13 2.44 | 0.01 6.87 | <0.0001 22.65 | 0.008 7.95 | 0.92 0.009 | 0.57 0.32 | 0.02 6.52 |
| Futile Cycle | ||||||||
| Gpd1 | p F (1, 36) | <0.0001 57.29 | 0.52 0.42 | 0.01 7.22 | 0.08 3.19 | 0.28 1.18 | 0.08 3.23 | 0.0004 15.42 |
| Gpd2 | p F (1, 36) | <0.0001 24.40 | <0.0001 51.07 | <0.0001 25.93 | 0.006 8.44 | 0.002 11.88 | 0.47 0.52 | <0.0001 44.04 |
| Group | Main Effects | Interactions | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fatty acid | Sex | WT-HF | WT-EPA | KO-HF | KO-EPA | S | G | D | S × G | S × D | G × D | S × G × D | |
| Palmitic acid 16:0 | M | 19.65 ± 0.91 | 19.14 ± 0.28 | 21.04 ± 0.34 | 20.08 ± 0.20 | p | <0.0001 76.78 | 0.24 1.46 | 0.008 8.71 | 0.06 4.03 | <0.0001 24.89 | 0.64 0.23 | 0.90 0.02 |
| F | 21.80 ± 0.58 ac | 24.81 ± 0.42 b | 21.63 ± 0.52 c | 24.39 ± 0.30 ab | F (1, 20) | ||||||||
| Palmitoleic acid 16:1 (n-7) | M | 1.59 ± 0.41 a | 0.21 ± 0.21 b | 1.53 ± 0.21 a | 1.17 ± 0.22 ab | p | 0.04 4.85 | 0.40 0.73 | 0.003 11.79 | 0.25 1.41 | 0.60 0.28 | 0.13 2.43 | 0.46 0.57 |
| F | 2.05 ± 0.45 | 1.23 ± 0.07 | 1.80 ± 0.26 | 1.34 ± 0.46 | F (1, 20) | ||||||||
| n-6 PUFAs | |||||||||||||
| Linoleic acid 18:2 (n-6) | M | 13.31 ± 0.76 | 13.36 ± 0.79 | 12.24 ± 0.59 | 12.52 ± 0.50 | p | <0.0001 1012 | 0.25 1.38 | 0.88 0.02 | 0.24 1.47 | 0.58 0.31 | 0.86 0.03 | 0.91 0.01 |
| F | 0.33 ± 0.09 | 0.02 ± 0.02 | 0.32 ± 0.08 | 0.06 ± 0.03 | F (1, 20) | ||||||||
| Arachidonic acid 20:4 (n-6) | M | 7.18 ± 1.21 a | 4.14 ± 0.39 ab | 4.18 ± 0.43 ab | 3.10 ± 0.21 b | p | 0.50 0.48 | 0.03 5.42 | 0.0001 22.77 | 0.23 1.51 | 0.27 1.30 | 0.20 1.78 | 0.70 0.16 |
| F | 7.29 ± 0.84 | 3.42 ± 0.41 | 6.14 ± 1.60 | 3.33 ± 0.42 | F (1, 20) | ||||||||
| n-3 PUFAs | |||||||||||||
| α-Linolenic acid 18:3 (n-3) | M | 0.06 ± 0.06 | 0.00 ± 0.00 | 0.13 ± 0.04 | 0.00 ± 0.00 | p | <0.0001 147.4 | 0.80 0.07 | 0.25 1.42 | 0.16 2.17 | 0.05 4.35 | 0.83 0.05 | 0.33 0.99 |
| F | 0.41 ± 0.05 | 0.41 ± 0.03 | 0.33 ± 0.05 | 0.38 ± 0.04 | F (1, 20) | ||||||||
| Eicosapentaenoic acid 20:5 (n-3) | M | 0.00 ± 0.00 a | 6.78 ± 0.73 b | 0.00 ± 0.00 a | 6.18 ± 0.35 b | p | 0.28 1.22 | 0.11 2.82 | <0.0001 329.5 | 0.39 0.79 | 0.28 1.22 | 0.11 2.82 | 0.39 0.79 |
| F | 0.00 ± 0.00 a | 8.29 ± 0.45 b | 0.00 ± 0.00 a | 6.34 ± 1.33 b | F (1, 20) | ||||||||
| Docosahexaenoic acid 22:5 (n-3) | M | 2.27 ± 0.60 a | 8.35 ± 0.50 b | 1.49 ± 0.18 a | 6.86 ± 0.28 b | p | 0.89 0.02 | 0.14 2.42 | <0.0001 116.6 | 0.36 0.089 | 0.08 3.30 | 0.97 0.001 | 0.43 0.65 |
| F | 3.10 ± 0.34 ab | 6.80 ± 0.69 b | 2.44 ± 0.58 a | 6.90 ± 1.54 b | F (1, 20) | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albracht-Schulte, K.; Wilson, S.; Johnson, P.; Pahlavani, M.; Ramalingam, L.; Goonapienuwala, B.; Kalupahana, N.S.; Festuccia, W.T.; Scoggin, S.; Kahathuduwa, C.N.; et al. Sex-Dependent Effects of Eicosapentaenoic Acid on Hepatic Steatosis in UCP1 Knockout Mice. Biomedicines 2021, 9, 1549. https://doi.org/10.3390/biomedicines9111549
Albracht-Schulte K, Wilson S, Johnson P, Pahlavani M, Ramalingam L, Goonapienuwala B, Kalupahana NS, Festuccia WT, Scoggin S, Kahathuduwa CN, et al. Sex-Dependent Effects of Eicosapentaenoic Acid on Hepatic Steatosis in UCP1 Knockout Mice. Biomedicines. 2021; 9(11):1549. https://doi.org/10.3390/biomedicines9111549
Chicago/Turabian StyleAlbracht-Schulte, Kembra, Savanna Wilson, Paige Johnson, Mandana Pahlavani, Latha Ramalingam, Bimba Goonapienuwala, Nishan S. Kalupahana, William T. Festuccia, Shane Scoggin, Chanaka N. Kahathuduwa, and et al. 2021. "Sex-Dependent Effects of Eicosapentaenoic Acid on Hepatic Steatosis in UCP1 Knockout Mice" Biomedicines 9, no. 11: 1549. https://doi.org/10.3390/biomedicines9111549
APA StyleAlbracht-Schulte, K., Wilson, S., Johnson, P., Pahlavani, M., Ramalingam, L., Goonapienuwala, B., Kalupahana, N. S., Festuccia, W. T., Scoggin, S., Kahathuduwa, C. N., & Moustaid-Moussa, N. (2021). Sex-Dependent Effects of Eicosapentaenoic Acid on Hepatic Steatosis in UCP1 Knockout Mice. Biomedicines, 9(11), 1549. https://doi.org/10.3390/biomedicines9111549