
Shared Patterns of Brain Functional Connectivity for the Comorbidity between Migraine and Insomnia

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
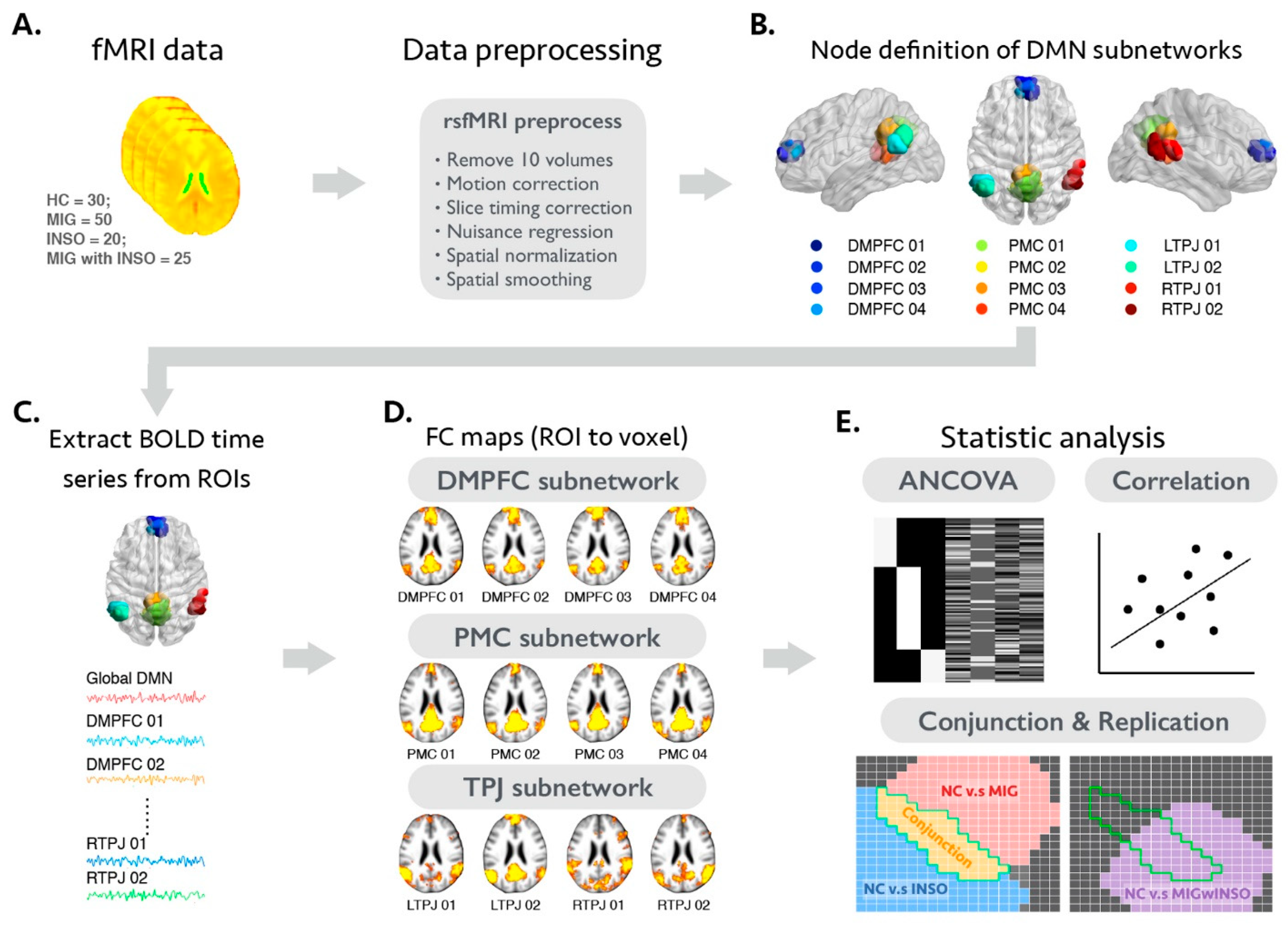
2.1. Participant Recruitment and Assessments
2.2. MRI Data Acquisitions
2.3. The Analytical Framework of the Present Study
2.4. Image Preprocessing of Individual rsfMRI Dataset
2.5. Selection Criteria and Analysis of Motion-Related Issues of the rsfMRI Dataset
2.6. Functional Connectivity Analysis and Statistical Criteria of Voxel-Wise Statistical Analyses
2.7. Statistical Analyses
2.7.1. Demographic Data, Clinical Evaluations, and Motion Profiles of rsfMRI Dataset
2.7.2. Analysis of FC Differences between Patients with Migraine, Patients with Insomnia, and Healthy Controls to Identify Shared and Distinct Functional Alterations
2.7.3. Validation Analysis of Shared Functional Alterations in an Additional Comorbid Migraine and Insomnia Group
2.7.4. Relationship between Regional FC Changes and Clinical Evaluations
3. Results
3.1. Demographic and Clinical Characteristics of Study Participants
3.2. Differences in the FC Magnitude of the Global DMN and Subnodal DMN between Healthy Controls and Patients with Migraine or Insomnia
3.2.1. FC Changes of the Global DMN in Patients with Migraine or Insomnia
3.2.2. FC Changes of the Subnodal DMN in Patients with Migraine or Insomnia
3.2.3. FC Changes of the Global and Subnodal DMN between Patients with Migraine or Insomnia
3.3. FC Alterations of the Global and Subnodal DMN Common to Migraine and Insomnia
3.4. Altered FC Patterns of the Global and Subnodal DMN in Patients with Comorbid Migraine and Insomnia
3.5. Potential Clinical Significance of Global and Subnodal DMN FC Changes in the Comorbidity Group
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Morin, C.M.; Benca, R. Chronic insomnia. Lancet 2012, 379, 1129–1141. [Google Scholar] [CrossRef]
- Ohayon, M.M. Epidemiology of insomnia: What we know and what we still need to learn. Sleep Med. Rev. 2002, 6, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Léger, D.; Partinen, M.; Hirshkowitz, M.; Chokroverty, S.; Touchette, E.; Hedner, J. Daytime consequences of insomnia symptoms among outpatients in primary care practice: EQUINOX international survey. Sleep Med. 2010, 11, 999–1009. [Google Scholar] [CrossRef]
- Kyle, S.D.; Beattie, L.; Spiegelhalder, K.; Rogers, Z.; Espie, C.A. Altered emotion perception in insomnia disorder. Sleep 2014, 37, 775–783. [Google Scholar] [CrossRef]
- Spiegelhalder, K.; Kyle, S.; Feige, B.; Prem, M.; Nissen, C.; Espie, C.A.; Riemann, D. The impact of sleep-related attentional bias on polysomnographically measured sleep in primary insomnia. Sleep 2010, 33, 107–112. [Google Scholar] [CrossRef]
- Lin, Y.-K.; Lin, G.-Y.; Lee, J.-T.; Lee, M.-S.; Tsai, C.-K.; Hsu, Y.-W.; Tsai, Y.-C.; Yang, F.-C. Associations between sleep quality and migraine frequency. Medicine 2016, 95, e3554. [Google Scholar] [CrossRef]
- Seidel, S.; Hartl, T.; Weber, M.; Matterey, S.; Pauli, A.; Riederer, F.; Gharabaghi, M.; Wöber-Bingöl, C.; Wöber, C.; PAMINA Study Group. Quality of sleep, fatigue and daytime sleepiness in migraine—A controlled study. Cephalalgia 2009, 29, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Rains, J.C.; Poceta, J.S. Headache and sleep disorders: Review and clinical implications for headache management. Headache J. Head Face Pain 2006, 46, 1344–1363. [Google Scholar] [CrossRef]
- Stovner, L.J.; Andree, C. Prevalence of headache in Europe: A review for the Eurolight project. J. Headache Pain 2010, 11, 289–299. [Google Scholar] [CrossRef]
- Haut, S.R.; Bigal, M.; Lipton, R.B. Chronic disorders with episodic manifestations: Focus on epilepsy and migraine. Lancet Neurol. 2006, 5, 148–157. [Google Scholar] [CrossRef]
- Freedom, T.; Evans, R.W. Headache and sleep. Headache 2013, 53, 1358–1366. [Google Scholar] [CrossRef] [PubMed]
- Kelman, L.; Rains, J.C. Headache and sleep: Examination of sleep patterns and complaints in a large clinical sample of migraineurs. Headache J. Head Face Pain 2005, 45, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Jansson-Fröjmark, M.; Lindblom, K. A bidirectional relationship between anxiety and depression, and insomnia? A prospective study in the general population. J. Psychosom. Res. 2008, 64, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Liang, C.; Lee, J.; Yeh, T.; Lee, M.-S.; Sung, Y.; Yang, F. Associations between depression/anxiety and headache frequency in migraineurs: A cross-sectional study. Headache J. Head Face Pain 2018, 58, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, M.; Steiner, T.J.; Scher, A.T.; Lipton, R.B. The global burden of migraine: Measuring disability in headache disorders with WHO’s Classification of Functioning, Disability and Health (ICF). J. Headache Pain 2005, 6, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Kyle, S.D.; Morgan, K.; Espie, C.A. Insomnia and health-related quality of life. Sleep Med. Rev. 2010, 14, 69–82. [Google Scholar] [CrossRef]
- Stokes, M.; Becker, W.J.; Lipton, R.B.; Sullivan, S.D.; Wilcox, T.K.; Wells, L.; Manack, A.; Msc, I.P.; Gladstone, J.; Buse, D.C.; et al. Cost of health care among patients with chronic and episodic migraine in Canada and the USA: Results from the international burden of migraine study (IBMS). Headache J. Head Face Pain 2011, 51, 1058–1077. [Google Scholar] [CrossRef]
- Biswal, B.; Yetkin, F.Z.; Haughton, V.M.; Hyde, J.S. Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn. Reson. Med. 1995, 34, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Greicius, M.D.; Krasnow, B.; Reiss, A.L.; Menon, V. Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proc. Natl. Acad. Sci. USA 2003, 100, 253–258. [Google Scholar] [CrossRef]
- Andrews-Hanna, J.R.; Saxe, R.; Yarkoni, T. Contributions of episodic retrieval and mentalizing to autobiographical thought: Evidence from functional neuroimaging, resting-state connectivity, and fMRI meta-analyses. NeuroImage 2014, 91, 324–335. [Google Scholar] [CrossRef]
- Buckner, R.L.; Andrews-Hanna, E.J.R.; Schacter, D. The brain’s default network. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef]
- Heuvel, M.V.D.; Sporns, O. Network hubs in the human brain. Trends Cogn. Sci. 2013, 17, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.L.; Sepulcre, J.; Talukdar, T.; Krienen, F.M.; Liu, H.; Hedden, T.; Andrews-Hanna, J.R.; Sperling, R.A.; Johnson, K.A. Cortical hubs revealed by intrinsic functional connectivity: Mapping, assessment of stability, and relation to Alzheimer’s disease. J. Neurosci. 2009, 29, 1860–1873. [Google Scholar] [CrossRef]
- Padmanabhan, A.; Lynch, C.; Schaer, M.; Menon, V. The default mode network in autism. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Broyd, S.J.; Demanuele, C.; Debener, S.; Helps, S.K.; James, C.J.; Sonuga-Barke, E.J. Default-mode brain dysfunction in mental disorders: A systematic review. Neurosci. Biobehav. Rev. 2009, 33, 279–296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Su, J.; Wang, M.; Zhao, Y.; Yao, Q.; Zhang, Q.; Lu, H.; Zhang, H.; Wang, S.; Li, G.-F.; et al. Increased default mode network connectivity and increased regional homogeneity in migraineurs without aura. J. Headache Pain 2016, 17, 98. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Huang, S.; Gao, L.; Zhuang, Y.; Ding, S.; Gong, H. Temporal regularity of intrinsic cerebral activity in patients with chronic primary insomnia: A brain entropy study using resting-state fMRI. Brain Behav. 2016, 6, e00529. [Google Scholar] [CrossRef]
- Laird, A.R.; Fox, P.M.; Eickhoff, S.B.; Turner, J.A.; Ray, K.L.; McKay, D.R.; Glahn, D.C.; Beckmann, C.F.; Smith, S.M. Behavioral Interpretations of Intrinsic Connectivity Networks. J. Cogn. Neurosci. 2011, 23, 4022–4037. [Google Scholar] [CrossRef]
- Andrews-Hanna, J.R.; Reidler, J.; Sepulcre, J.; Poulin, R.; Buckner, R.L. Functional-anatomic fractionation of the brain’s default network. Neuron 2010, 65, 550–562. [Google Scholar] [CrossRef]
- Damoiseaux, J.S.; Prater, K.; Miller, B.L.; Greicius, M.D. Functional connectivity tracks clinical deterioration in Alzheimer’s disease. Neurobiol. Aging 2012, 33, 828.e19–828.e30. [Google Scholar] [CrossRef]
- Li, B.; Liu, L.; Friston, K.; Shen, H.; Wang, L.; Zeng, L.-L.; Hu, D. A treatment-resistant default mode subnetwork in major depression. Biol. Psychiatry 2013, 74, 48–54. [Google Scholar] [CrossRef]
- Davey, C.G.; Pujol, J.; Harrison, B. Mapping the self in the brain’s default mode network. NeuroImage 2016, 132, 390–397. [Google Scholar] [CrossRef]
- Cabeza, R.; Dolcos, F.; Graham, R.; Nyberg, L. Similarities and differences in the neural correlates of episodic memory retrieval and working memory. NeuroImage 2002, 16, 317–330. [Google Scholar] [CrossRef]
- Small, D.; Gitelman, D.; Gregory, M.; Nobre, A.; Parrish, T.; Mesulam, M.-M. The posterior cingulate and medial prefrontal cortex mediate the anticipatory allocation of spatial attention. NeuroImage 2003, 18, 633–641. [Google Scholar] [CrossRef]
- Simpson, J.R.; Drevets, W.C.; Snyder, A.Z.; Gusnard, D.A.; Raichle, M.E. Emotion-induced changes in human medial prefrontal cortex: II. During anticipatory anxiety. Proc. Natl. Acad. Sci. USA 2001, 98, 688–693. [Google Scholar] [CrossRef]
- Wilcox, S.L.; Veggeberg, R.; Lemme, J.; Hodkinson, D.J.; Scrivani, S.; Burstein, R.; Becerra, L.; Borsook, D. Increased functional activation of limbic brain regions during negative emotional processing in migraine. Front. Hum. Neurosci. 2016, 10, 366. [Google Scholar] [CrossRef]
- Yan, C.-Q.; Wang, X.; Huo, J.-W.; Zhou, P.; Li, J.-L.; Wang, Z.-Y.; Zhang, J.; Fu, Q.-N.; Wang, X.-R.; Liu, C.-Z.; et al. Abnormal global brain functional connectivity in primary insomnia patients: A resting-state functional MRI study. Front. Neurol. 2018, 9, 856. [Google Scholar] [CrossRef]
- Regier, D.A.; Kuhl, E.A.; Kupfer, D.J. The DSM-5: Classification and criteria changes. World Psychiatry 2013, 12, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Bastien, C.H.; Vallières, A.; Morin, C.M. Validation of the Insomnia Severity Index as an outcome measure for insomnia research. Sleep Med. 2001, 2, 297–307. [Google Scholar] [CrossRef]
- Buysse, D.J.; Reynolds, C.F.; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh sleep quality index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef]
- Headache Classification Committee of the International Headache Society (IHS). The international classification of headache disorders, 3rd edition (beta version). Cephalalgia 2013, 33, 629–808. [Google Scholar] [CrossRef]
- Zigmond, A.S.; Snaith, R.P. The hospital anxiety and depression scale. Acta Psychiatr. Scand. 1983, 67, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. NeuroImage 2012, 59, 2142–2154. [Google Scholar] [CrossRef] [PubMed]
- Lefort-Besnard, J.; Bassett, D.S.; Smallwood, J.; Margulies, D.; Derntl, B.; Gruber, O.; Aleman, A.; Jardri, R.; Varoquaux, G.; Thirion, B.; et al. Different shades of default mode disturbance in schizophrenia: Subnodal covariance estimation in structure and function. Hum. Brain Mapp. 2017, 39, 644–661. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J.; Williams, S.; Howard, R.; Frackowiak, R.S.J.; Turner, R. Movement-related effects in fMRI time-series. Magn. Reson. Med. 1996, 35, 346–355. [Google Scholar] [CrossRef]
- Murphy, K.; Fox, M.D. Towards a consensus regarding global signal regression for resting state functional connectivity MRI. NeuroImage 2017, 154, 169–173. [Google Scholar] [CrossRef]
- Yang, F.-C.; Chou, K.-H.; Fuh, J.-L.; Lee, P.-L.; Lirng, J.-F.; Lin, Y.-Y.; Lin, C.-P.; Wang, S.-J. Altered hypothalamic functional connectivity in cluster headache: A longitudinal resting-state functional MRI study. J. Neurol. Neurosurg. Psychiatry 2015, 86, 437–445. [Google Scholar] [CrossRef]
- Power, J.D.; Mitra, A.; Laumann, T.O.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Methods to detect, characterize, and remove motion artifact in resting state fMRI. NeuroImage 2014, 84, 320–341. [Google Scholar] [CrossRef]
- Chou, K.-H.; Lee, P.-L.; Liang, C.-S.; Lee, J.-T.; Kao, H.-W.; Tsai, C.-L.; Lin, G.-Y.; Lin, Y.-K.; Lin, C.-P.; Yang, F.-C. Identifying neuroanatomical signatures in insomnia and migraine comorbidity. Sleep 2020, 44, zsaa202. [Google Scholar] [CrossRef]
- Ursin, H.; Eriksen, H. Cognitive activation theory of stress, sensitization, and common health complaints. Ann. N. Y. Acad. Sci. 2007, 1113, 304–310. [Google Scholar] [CrossRef]
- Tanasescu, R.; Cottam, W.; Condon, L.; Tench, C.R.; Auer, D.P. Functional reorganisation in chronic pain and neural correlates of pain sensitisation: A coordinate based meta-analysis of 266 cutaneous pain fMRI studies. Neurosci. Biobehav. Rev. 2016, 68, 120–133. [Google Scholar] [CrossRef]
- Yang, S.; Chang, M.C. Chronic pain: Structural and functional changes in brain structures and associated negative affective states. Int. J. Mol. Sci. 2019, 20, 3130. [Google Scholar] [CrossRef]
- Vatthauer, K.; Craggs, J.G.; Robinson, M.; Staud, R.; Berry, R.B.; Perlstein, W.M.; McCrae, C.S. Sleep is associated with task-negative brain activity in fibromyalgia participants with comorbid chronic insomnia. J. Pain Res. 2015, 8, 819–827. [Google Scholar] [CrossRef][Green Version]
- Dai, X.-J.; Nie, X.; Shao, Y.; Liu, S.-Y.; Li, H.-J.; Wan, A.-L.; Nie, S.; Peng, D.-C. Functional connectivity of paired default mode network subregions in primary insomnia. Neuropsychiatr. Dis. Treat. 2015, 11, 3085–3093. [Google Scholar] [CrossRef]
- Regen, W.; Kyle, S.D.; Nissen, C.; Feige, B.; Baglioni, C.; Hennig, J.; Riemann, D.; Spiegelhalder, K. Objective sleep disturbances are associated with greater waking resting-state connectivity between the retrosplenial cortex/ hippocampus and various nodes of the default mode network. J. Psychiatry Neurosci. 2016, 41, 295–303. [Google Scholar] [CrossRef]
- Xue, T.; Yuan, K.; Zhao, L.; Yu, D.; Zhao, L.; Dong, T.; Cheng, P.; Von Deneen, K.M.; Qin, W.; Tian, J. Intrinsic brain network abnormalities in migraines without aura revealed in resting-state fMRI. PLoS ONE 2012, 7, e52927. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Yuan, K.; Zhao, L.; Zhao, L.; Dong, M.; Liu, P.; Wang, G.; Liu, J.; Sun, J.; Zhou, G.; et al. Regional homogeneity abnormalities in patients with interictal migraine without aura: A resting-state study. NMR Biomed. 2012, 25, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef]
- Petrovic, P. Placebo and opioid analgesia—Imaging a shared neuronal network. Science 2002, 295, 1737–1740. [Google Scholar] [CrossRef]
- Dai, X.-J.; Nie, X.; Liu, X.; Pei, L.; Jiang, J.; Peng, D.-C.; Gong, H.-H.; Zeng, X.-J.; Wáng, Y.-X.J.; Zhan, Y. Gender differences in regional brain activity in patients with chronic primary insomnia: Evidence from a resting-state fMRI study. J. Clin. Sleep Med. 2016, 12, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.-J.; Peng, D.-C.; Gong, H.-H.; Wan, A.-L.; Nie, X.; Li, H.-J.; Wang, Y.-X. Altered intrinsic regional brain spontaneous activity and subjective sleep quality in patients with chronic primary insomnia: A resting-state fMRI study. Neuropsychiatr. Dis. Treat. 2014, 10, 2163–2175. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, E.; Zhang, H.; Dou, S.; Liu, L.; Tong, L.; Lei, Y.; Wang, M.; Xu, J.; Shi, D.; et al. Functional connectivity changes between parietal and prefrontal cortices in primary insomnia patients: Evidence from resting-state fMRI. Eur. J. Med. Res. 2014, 19, 32. [Google Scholar] [CrossRef]
- Cavanna, A.E.; Trimble, M.R. The precuneus: A review of its functional anatomy and behavioural correlates. Brain 2006, 129, 564–583. [Google Scholar] [CrossRef] [PubMed]
- Goffaux, P.; Girard-Tremblay, L.; Marchand, S.; Daigle, K.; Whittingstall, K. Individual differences in pain sensitivity vary as a function of precuneus reactivity. Brain Topogr. 2014, 27, 366–374. [Google Scholar] [CrossRef]
- Seifert, F.; Maihöfner, C. Functional and structural imaging of pain-induced neuroplasticity. Curr. Opin. Anaesthesiol. 2011, 24, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Georgopoulos, A.P. Neural aspects of cognitive motor control. Curr. Opin. Neurobiol. 2000, 10, 238–241. [Google Scholar] [CrossRef]
- Valfrè, W.; Rainero, I.; Bergui, M.; Pinessi, L. Voxel-based morphometry reveals gray matter abnormalities in migraine. Headache J. Head Face Pain 2007, 48, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Tamura, Y.; Okabe, S.; Ohnishi, T.; Saito, D.N.; Arai, N.; Mochio, S.; Inoue, K.; Ugawa, Y. Effects of 1-Hz repetitive transcranial magnetic stimulation on acute pain induced by capsaicin. Pain 2004, 107, 107–115. [Google Scholar] [CrossRef]
- Wang, T.; Li, S.; Jiang, G.; Lin, C.; Li, M.; Ma, X.; Zhan, W.; Fang, J.; Li, L.; Li, C.; et al. Regional homogeneity changes in patients with primary insomnia. Eur. Radiol. 2016, 26, 1292–1300. [Google Scholar] [CrossRef]
- Huang, Z.; Liang, P.; Jia, X.; Zhan, S.; Li, N.; Ding, Y.; Lu, J.; Wang, Y.; Li, K. Abnormal amygdala connectivity in patients with primary insomnia: Evidence from resting state fMRI. Eur. J. Radiol. 2012, 81, 1288–1295. [Google Scholar] [CrossRef]
- Mehnert, J.; Schulte, L.; Timmann, D.; May, A. Activity and connectivity of the cerebellum in trigeminal nociception. NeuroImage 2017, 150, 112–118. [Google Scholar] [CrossRef]
- Goadsby, P.; Holland, P.; Martins-Oliveira, M.; Hoffmann, J.; Schankin, C.; Akerman, S. Pathophysiology of migraine: A disorder of sensory processing. Physiol. Rev. 2017, 97, 553–622. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Li, C.; Ma, X.; Dong, M.; Yin, Y.; Hua, K.; Li, M.; Li, C.; Zhan, W.; Li, C. Abnormal spontaneous regional brain activity in primary insomnia: A resting-state functional magnetic resonance imaging study. Neuropsychiatr. Dis. Treat. 2016, 12, 1371–1378. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Dong, M.; Yin, Y.; Hua, K.; Fu, S.; Jiang, G. Aberrant effective connectivity of the right anterior insula in primary insomnia. Front. Neurol. 2018, 9, 317. [Google Scholar] [CrossRef]
- Zhang, J.; Su, J.; Wang, M.; Zhao, Y.; Zhang, Q.-T.; Yao, Q.; Lu, H.; Zhang, H.; Li, G.-F.; Wu, Y.-L.; et al. The sensorimotor network dysfunction in migraineurs without aura: A resting-state fMRI study. J. Neurol. 2017, 264, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.; et al. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci. Transl. Med. 2012, 4, 147. [Google Scholar] [CrossRef] [PubMed]
- Christensen, J.; Yamakawa, G.R.; Shultz, S.R.; Mychasiuk, R. Is the glymphatic system the missing link between sleep impairments and neurological disorders? Examining the implications and uncertainties. Prog. Neurobiol. 2021, 198, 101917. [Google Scholar] [CrossRef]
- Schain, A.J.; Melo-Carrillo, A.; Strassman, A.M.; Burstein, R. Cortical spreading depression closes paravascular space and impairs glymphatic flow: Implications for migraine headache. J. Neurosci. 2017, 37, 2904–2915. [Google Scholar] [CrossRef]
- Roh, J.H.; Huang, Y.; Bero, A.W.; Kasten, T.; Stewart, F.R.; Bateman, R.; Holtzman, D.M. Disruption of the sleep-wake cycle and diurnal fluctuation of -amyloid in mice with alzheimer’s disease pathology. Sci. Transl. Med. 2012, 4, 150. [Google Scholar] [CrossRef]
- Ringstad, G.; Vatnehol, S.A.S.; Eide, P.K. Glymphatic MRI in idiopathic normal pressure hydrocephalus. Brain 2017, 140, 2691–2705. [Google Scholar] [CrossRef]
- Ringstad, G.; Valnes, L.M.; Dale, A.M.; Pripp, A.H.; Vatnehol, S.A.S.; Emblem, K.; Mardal, K.-A.; Eide, P.K. Brain-wide glymphatic enhancement and clearance in humans assessed with MRI. JCI Insight 2018, 3, e121537. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.; Wang, D.; Fox, M.D.; Yeo, B.T.; Sepulcre, J.; Sabuncu, M.; Shafee, R.; Lu, J.; Liu, H. Individual variability in functional connectivity architecture of the human brain. Neuron 2013, 77, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Buckner, R.L.; Fox, M.D.; Holt, D.J.; Holmes, A.J.; Stoecklein, S.; Langs, G.; Pan, R.; Qian, T.; Li, K.; et al. Parcellating cortical functional networks in individuals. Nat. Neurosci. 2015, 18, 1853–1860. [Google Scholar] [CrossRef] [PubMed]
- Kushida, C.A.; Littner, M.R.; Morgenthaler, T.; Alessi, C.A.; Bailey, D.; Coleman, J.; Friedman, L.; Hirshkowitz, M.; Kapen, S.; Kramer, M.; et al. Practice parameters for the indications for polysomnography and related procedures: An update for 2005. Sleep 2005, 28, 499–523. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Demographic Variables | HC (n = 30) | MIG (n = 50) | INSO (n = 20) | MIG with INSO (n = 25) | p Value |
|---|---|---|---|---|---|
| Age (years) | 41.47 ± 10.41 | 40.14 ± 8.83 | 40.15 ± 10.25 | 41.9 ± 11.63 | 0.863 a |
| Sex (male/female) | 7/23 | 6/44 | 7/13 | 3/22 | 0.102 b |
| Framewise Displacement | 0.073 ± 0.02 | 0.074 ± 0.02 | 0.072 ± 0.02 | 0.076 ± 0.02 | 0.512 e |
| Aura /no Aura | - | 6/44 | - | 7/18 | 0.084 c |
| Migraine duration (years) | - | 15.63 ± 10.55 | - | 15.00 ± 10.68 | 0.622 d |
| Migraine freq. (days/month) | - | 7.38 ± 5.58 | - | 12.70 ± 9.38 | 0.003 d |
| Insomnia duration (month) | - | - | 21.32 ± 22.88 | 15.54 ± 10.31 | 0.313 d |
| ISI total score (0–28) | 7.03 ± 4.45 | 6.80 ± 4.79 | 15.95 ± 3.65 | 16.36 ± 4.33 | <0.001 e |
| PSQI total score (0–21) | 7.19 ± 3.43 | 8.00 ± 3.61 | 11.05 ± 3.52 | 11.00 ± 3.51 | <0.001 e |
| HADS score (0–42) | 9.83 ± 4.98 | 12.18 ± 7.19 | 13.45 ± 8.66 | 15.97 ± 7.74 | 0.018 e |
| MNI Coordinates | Cluster Size | Anatomical Region | Regional FC (Mean ± SD) | |||||
|---|---|---|---|---|---|---|---|---|
| x | y | z | HC | MIG | INSO | |||
| Global DMN | −24 | −10 | 68 | 68 | Lt. Precentral Gyrus | 0.025 ± 0.185 | 0.175 ± 0.152 | 0.215 ± 0.174 |
| 6 | −79 | −6 | 100 | Rt. Cerebellum V | 0.251 ± 0.147 | 0.403 ± 0.139 | 0.451 ± 0.134 | |
| DMPFC 01 | −23 | −9 | 67 | 59 | Lt. Precentral Gyrus | −0.031 ± 0.154 | 0.111 ± 0.151 | 0.151 ± 0.167 |
| DMPFC 03 | −4 | −42 | 65 | 37 | Lt. Precentral Gyrus | 0.072 ± 0.141 | 0.208 ± 0.146 | 0.264 ± 0.159 |
| 7 | −39 | 71 | 24 | Rt. Precentral Gyrus | 0.063 ± 0.140 | 0.215 ± 0.151 | 0.236 ± 0.205 | |
| DMPFC 04 | 8 | −46 | 12 | 30 | Rt. Cingulate Gyrus, posterior division | 0.248 ± 0.144 | 0.374 ± 0.165 | 0.424 ± 0.148 |
| 55 | −62 | 32 | 62 | Rt. Lateral Occipital Cortex, superior division | 0.218 ± 0.160 | 0.365 ± 0.180 | 0.415 ± 0.135 | |
| PMC 03 | −27 | −10 | 69 | 40 | Lt. Precentral Gyrus | −0.033 ± 0.152 | 0.109 ± 0.187 | 0.166 ± 0.193 |
| 6 | −77 | −6 | 333 | Rt. Cerebellum V | 0.177 ± 0.167 | 0.336 ± 0.135 | 0.391 ± 0.149 | |
| PMC 04 | −45 | −22 | 57 | 85 | Lt. Postcentral Gyrus | 0.153 ± 0.181 | 0.294 ± 0.177 | 0.327 ± 0.221 |
| MNI Coordinates | Cluster Size | Maximum T Value | Anatomical Region | Regional FC (Mean ± SD) | ||||
|---|---|---|---|---|---|---|---|---|
| x | y | z | HC | MIGwINSO | ||||
| Global DMN | −22 | −16 | 76 | 391 | 5.61 | Lt. Precentral Gyrus | 0.063 ± 0.141 | 0.253 ± 0.134 |
| DMPFC 01 | −22 | −14 | 76 | 1371 | 5.96 | Lt. Precentral Gyrus | 0.039 ± 0.111 | 0.179 ± 0.117 |
| DMPFC 03 | 4 | −14 | 68 | 363 | 4.18 | Rt. Precentral Gyrus | 0.040 ± 0.106 | 0.197 ± 0.134 |
| DMPFC 04 | 9 | −41 | 7 | 1664 | 4.67 | Rt. Cingulate Gyrus, posterior division | 0.161 ± 0.102 | 0.324 ± 0.099 |
| 38 | −61 | 39 | 465 | 5.05 | Rt. Lateral Occipital Cortex, superior division | 0.079 ± 0.086 | 0.234 ± 0.118 | |
| PMC 03 | −25 | −14 | 74 | 340 | 4.88 | Lt. Precentral Gyrus | −0.015 ± 0.104 | 0.154 ± 0.086 |
| PMC 04 | −57 | −22 | 44 | 484 | 4.83 | Lt. Postcentral Gyrus | 0.106 ± 0.147 | 0.291 ± 0.186 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, K.-H.; Kuo, C.-Y.; Liang, C.-S.; Lee, P.-L.; Tsai, C.-K.; Tsai, C.-L.; Huang, M.-H.; Hsu, Y.-C.; Lin, G.-Y.; Lin, Y.-K.; et al. Shared Patterns of Brain Functional Connectivity for the Comorbidity between Migraine and Insomnia. Biomedicines 2021, 9, 1420. https://doi.org/10.3390/biomedicines9101420
Chou K-H, Kuo C-Y, Liang C-S, Lee P-L, Tsai C-K, Tsai C-L, Huang M-H, Hsu Y-C, Lin G-Y, Lin Y-K, et al. Shared Patterns of Brain Functional Connectivity for the Comorbidity between Migraine and Insomnia. Biomedicines. 2021; 9(10):1420. https://doi.org/10.3390/biomedicines9101420
Chicago/Turabian StyleChou, Kun-Hsien, Chen-Yuan Kuo, Chih-Sung Liang, Pei-Lin Lee, Chia-Kuang Tsai, Chia-Lin Tsai, Ming-Hao Huang, Yi-Chih Hsu, Guan-Yu Lin, Yu-Kai Lin, and et al. 2021. "Shared Patterns of Brain Functional Connectivity for the Comorbidity between Migraine and Insomnia" Biomedicines 9, no. 10: 1420. https://doi.org/10.3390/biomedicines9101420
APA StyleChou, K.-H., Kuo, C.-Y., Liang, C.-S., Lee, P.-L., Tsai, C.-K., Tsai, C.-L., Huang, M.-H., Hsu, Y.-C., Lin, G.-Y., Lin, Y.-K., Lin, C.-P., & Yang, F.-C. (2021). Shared Patterns of Brain Functional Connectivity for the Comorbidity between Migraine and Insomnia. Biomedicines, 9(10), 1420. https://doi.org/10.3390/biomedicines9101420