
Bile Duct Ligation Upregulates Expression and Function of L-Amino Acid Transporter 1 at Blood–Brain Barrier of Rats via Activation of Aryl Hydrocarbon Receptor by Bilirubin

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
2.3. Development of BDL Rats
2.4. Brain Distributions of Gabapentin and AAAs in Rats
2.5. HCMEC/D3 Culture and Drug Treatment
2.6. Uptakes of Gabapentin in HCMEC/D3
2.7. AhR Knockdown with siRNA in HCMEC/D3 Cells
2.8. Effects of Hyperbilirubinemia on LAT1 Function and Expression in the Brain of Rats
2.9. Western Blot
2.10. Statistical Analysis
3. Results
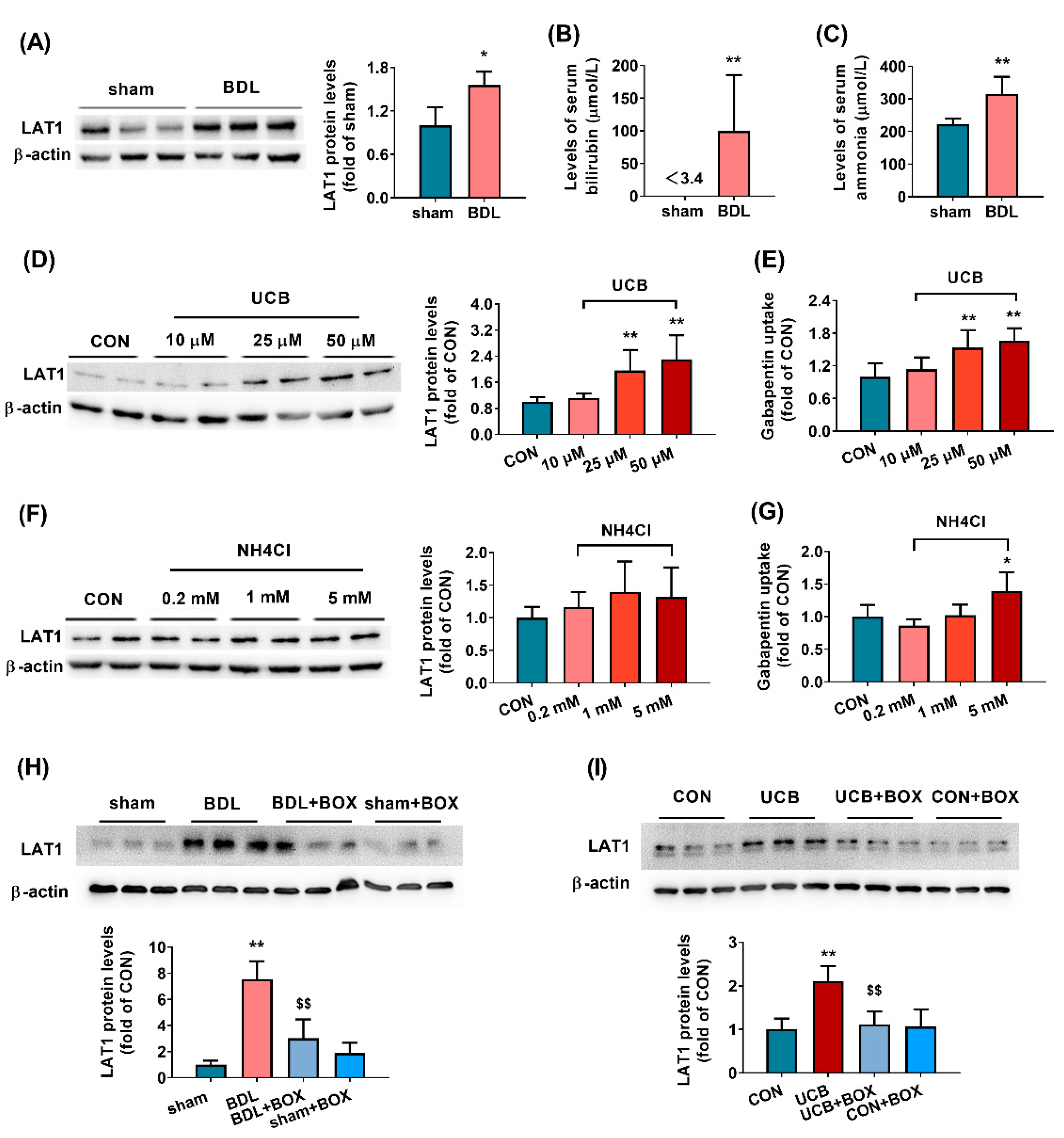
3.1. Biochemical and Physiological Parameters in BDL Rats
3.2. BDL Increases the Function and Expression of LAT1 in Brain Tissues of Rats
3.3. Effects of UCB and Ammonia on Expression and Functions of LAT1 in HCMEC/D3 Cells
3.4. Implications of AhR Pathway in Upregulation of LAT1 Expression by UCB in HCMEC/D3 Cells
3.5. Effects of Hyperbilirubinemia on Brain LAT1 Expression and Function
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wijdicks, E.F.M. Hepatic Encephalopathy. N. Engl. J. Med. 2016, 375, 1660–1670. [Google Scholar] [CrossRef]
- Weissenborn, K. Hepatic Encephalopathy: Definition, Clinical Grading and Diagnostic Principles. Drugs 2019, 79, 5–9. [Google Scholar] [CrossRef]
- Record, C.O.; Buxton, B.; Chase, R.A.; Curzon, G.; Murray-Lyon, I.M.; Williams, R. Plasma and brain amino acids in fulminant hepatic failure and their relationship to hepatic encephalopathy. Eur. J. Clin. Investig. 1976, 6, 387–394. [Google Scholar] [CrossRef]
- Ono, J.; Hutson, D.G.; Dombro, R.S.; Levi, J.U.; Livingstone, A.; Zeppa, R. Tryptophan and hepatic coma. Gastroenterology 1978, 74, 196–200. [Google Scholar] [CrossRef]
- Cascino, A.; Cangiano, C.; Fiaccadori, F.; Ghinelli, F.; Merli, M.; Pelosi, G.; Riggio, O.; Rossi Fanelli, F.; Sacchini, D.; Stortoni, M.; et al. Plasma and cerebrospinal fluid amino acid patterns in hepatic encephalopathy. Dig. Dis. Sci. 1982, 27, 828–832. [Google Scholar] [CrossRef]
- Smith, A.R.; Rossi-Fanelli, F.; Ziparo, V.; James, J.H.; Perelle, B.A.; Fischer, J.E. Alterations in plasma and CSF amino acids, amines and metabolites in hepatic coma. Ann. Surg. 1978, 187, 343–350. [Google Scholar] [CrossRef]
- Watanabe, A.; Higashi, T.; Shiota, T.; Takei, N.; Sakata, T.; Fujiwara, M.; Nakatsukasa, H.; Kobayashi, M.; Nagashima, H. Alterations in neutral amino acid transport across the blood-brain barrier in hepatic failure. Res. Exp. Med. Zeitschrift Exp. Medizin Einschl. Exp. Chir. 1983, 183, 11–18. [Google Scholar] [CrossRef]
- Buxton, B.H.; Stewart, D.A.; Murray-Lyon, I.M.; Curzon, G.; Williams, R. Plasma amino acids in experimental acute hepatic failure and their relationship to brain tryptophan. Clin. Sci. Mol. Med. 1974, 46, 559–562. [Google Scholar] [CrossRef]
- Bergeron, M.; Layrargues, G.P.; Butterworth, R.F. Aromatic and branched-chain amino acids in autopsied brain tissue from cirrhotic patients with hepatic encephalopathy. Metab. Brain Dis. 1989, 4, 169–176. [Google Scholar] [CrossRef]
- Michalak, A.; Butterworth, R.F. Selective increases of extracellular brain concentrations of aromatic and branched-chain amino acids in relation to deterioration of neurological status in acute (ischemic) liver failure. Metab. Brain Dis. 1997, 12, 259–269. [Google Scholar] [CrossRef]
- Tossman, U.; Eriksson, S.; Delin, A.; Hagenfeldt, L.; Law, D.; Ungerstedt, U. Brain amino acids measured by intracerebral dialysis in portacaval shunted rats. J. Neurochem. 1983, 41, 1046–1051. [Google Scholar] [CrossRef]
- Saleem, D.M.; Haider, S.; Khan, M.M.; Shamsi, T.; Haleem, D.J. Role of tryptophan in the pathogenesis of hepatic encephalopathy. J. Pak. Med. Assoc. 2008, 58, 68–70. [Google Scholar]
- Borg, J.; Warter, J.M.; Schlienger, J.L.; Imler, M.; Marescaux, C.; Mack, G. Neurotransmitter modifications in human cerebrospinal fluid and serum during hepatic encephalopathy. J. Neurol. Sci. 1982, 57, 343–356. [Google Scholar] [CrossRef]
- Jellinger, K.; Riederer, P. Brain monoamines in metabolic (endotoxic) coma. A preliminary biochemical study in human postmortem material. J. Neural Transm. 1977, 41, 275–286. [Google Scholar] [CrossRef]
- Ding, S.; Liu, L.; Jing, H.; Xie, J.; Wang, X.; Mao, J.; Chen, B.; Zhuge, Q. Dopamine from cirrhotic liver contributes to the impaired learning and memory ability of hippocampus in minimal hepatic encephalopathy. Hepatol. Int. 2013, 7, 923–936. [Google Scholar] [CrossRef]
- Ding, S.; Wang, X.; Zhuge, W.; Yang, J.; Zhuge, Q. Dopamine induces glutamate accumulation in astrocytes to disrupt neuronal function leading to pathogenesis of minimal hepatic encephalopathy. Neuroscience 2017, 365, 94–113. [Google Scholar] [CrossRef]
- Avraham, Y.; Grigoriadis, N.C.; Magen, I.; Poutahidis, T.; Vorobiav, L.; Zolotarev, O.; Ilan, Y.; Mechoulam, R.; Berry, E.M. Capsaicin affects brain function in a model of hepatic encephalopathy associated with fulminant hepatic failure in mice. Br. J. Pharmacol. 2009, 158, 896–906. [Google Scholar] [CrossRef]
- Avraham, Y.; Grigoriadis, N.; Poutahidis, T.; Vorobiev, L.; Magen, I.; Ilan, Y.; Mechoulam, R.; Berry, E. Cannabidiol improves brain and liver function in a fulminant hepatic failure-induced model of hepatic encephalopathy in mice. Br. J. Pharmacol. 2011, 162, 1650–1658. [Google Scholar] [CrossRef]
- Bernacki, J.; Dobrowolska, A.; Nierwińska, K.; Małecki, A. Physiology and pharmacological role of the blood-brain barrier. Pharmacol. Rep. 2008, 60, 600–622. [Google Scholar]
- Singh, N.; Ecker, G.F. Insights into the Structure, Function, and Ligand Discovery of the Large Neutral Amino Acid Transporter 1, LAT1. Int. J. Mol. Sci. 2018, 19, 1278. [Google Scholar] [CrossRef]
- Dickens, D.; Chiduza, G.N.; Wright, G.S.A.; Pirmohamed, M.; Antonyuk, S.V.; Hasnain, S.S. Modulation of LAT1 (SLC7A5) transporter activity and stability by membrane cholesterol. Sci. Rep. 2017, 7, 43580. [Google Scholar] [CrossRef] [PubMed]
- Kanai, Y.; Segawa, H.; Miyamoto, K.I.; Uchino, H.; Takeda, E.; Endou, H. Expression cloning and characterization of a transporter for large neutral amino acids activated by the heavy chain of 4F2 antigen (CD98). J. Biol. Chem. 1998, 273, 23629–23632. [Google Scholar] [CrossRef]
- Lin, J.; Raoof, D.A.; Thomas, D.G.; Greenson, J.K.; Giordano, T.J.; Robinson, G.S.; Bourner, M.J.; Bauer, C.T.; Orringer, M.B.; Beer, D.G. L-type amino acid transporter-1 overexpression and melphalan sensitivity in Barrett’s adenocarcinoma. Neoplasia 2004, 6, 74–84. [Google Scholar] [CrossRef]
- Kageyama, T.; Nakamura, M.; Matsuo, A.; Yamasaki, Y.; Takakura, Y.; Hashida, M.; Kanai, Y.; Naito, M.; Tsuruo, T.; Minato, N.; et al. The 4F2hc/LAT1 complex transports L-DOPA across the blood-brain barrier. Brain Res. 2000, 879, 115–121. [Google Scholar] [CrossRef]
- Takahashi, Y.; Nishimura, T.; Higuchi, K.; Noguchi, S.; Tega, Y.; Kurosawa, T.; Deguchi, Y.; Tomi, M. Transport of Pregabalin Via L-Type Amino Acid Transporter 1 (SLC7A5) in Human Brain Capillary Endothelial Cell Line. Pharm. Res. 2018, 35, 246. [Google Scholar] [CrossRef] [PubMed]
- Dickens, D.; Webb, S.D.; Antonyuk, S.; Giannoudis, A.; Owen, A.; Rädisch, S.; Hasnain, S.S.; Pirmohamed, M. Transport of gabapentin by LAT1 (SLC7A5). Biochem. Pharmacol. 2013, 85, 1672–1683. [Google Scholar] [CrossRef]
- Zevenbergen, C.; Meima, M.E.; Lima de Souza, E.C.; Peeters, R.P.; Kinne, A.; Krause, G.; Visser, W.E.; Visser, T.J. Transport of Iodothyronines by Human L-Type Amino Acid Transporters. Endocrinology 2015, 156, 4345–4355. [Google Scholar] [CrossRef][Green Version]
- Kanai, Y.; Endou, H. Functional properties of multispecific amino acid transporters and their implications to transporter-mediated toxicity. J. Toxicol. Sci. 2003, 28, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Papin-Michault, C.; Bonnetaud, C.; Dufour, M.; Almairac, F.; Coutts, M.; Patouraux, S.; Virolle, T.; Darcourt, J.; Burel-Vandenbos, F. Study of LAT1 Expression in Brain Metastases: Towards a Better Understanding of the Results of Positron Emission Tomography Using Amino Acid Tracers. PLoS ONE 2016, 11, e0157139. [Google Scholar] [CrossRef]
- Dadone-Montaudié, B.; Ambrosetti, D.; Dufour, M.; Darcourt, J.; Almairac, F.; Coyne, J.; Virolle, T.; Humbert, O.; Burel-Vandenbos, F. [18F] FDOPA standardized uptake values of brain tumors are not exclusively dependent on LAT1 expression. PLoS ONE 2017, 12, e0184625. [Google Scholar] [CrossRef]
- Hutterer, M.; Ebner, Y.; Riemenschneider, M.J.; Willuweit, A.; McCoy, M.; Egger, B.; Schröder, M.; Wendl, C.; Hellwig, D.; Grosse, J.; et al. Epileptic Activity Increases Cerebral Amino Acid Transport Assessed by 18F-Fluoroethyl-l-Tyrosine Amino Acid PET: A Potential Brain Tumor Mimic. J. Nucl. Med. 2017, 58, 129–137. [Google Scholar] [CrossRef]
- Lim, B.-C.; Cho, K.-Y.; Lim, J.-S.; Lee, R.-S.; Kim, H.-S.; Kim, M.-K.; Kim, J.-H.; Woo, Y.-J.; Kim, J.-K.; Kim, D.K.; et al. Increased expression of L-amino acid transporters in balloon cells of tuberous sclerosis. Child’s Nerv. Syst. ChNS Off. J. Int. Soc. Pediatr. Neurosurg. 2011, 27, 63–70. [Google Scholar] [CrossRef]
- Bao, X.; Wu, J.; Xie, Y.; Kim, S.; Michelhaugh, S.; Jiang, J.; Mittal, S.; Sanai, N.; Li, J. Protein Expression and Functional Relevance of Efflux and Uptake Drug Transporters at the Blood-Brain Barrier of Human Brain and Glioblastoma. Clin. Pharmacol. Ther. 2020, 107, 1116–1127. [Google Scholar] [CrossRef]
- Cai, L.; Kirchleitner, S.V.; Zhao, D.; Li, M.; Tonn, J.-C.; Glass, R.; Kälin, R.E. Glioblastoma Exhibits Inter-Individual Heterogeneity of TSPO and LAT1 Expression in Neoplastic and Parenchymal Cells. Int. J. Mol. Sci. 2020, 21, 612. [Google Scholar] [CrossRef] [PubMed]
- DeMorrow, S.; Cudalbu, C.; Davies, N.; Jayakumar, A.R.; Rose, C.F. 2021 ISHEN guidelines on animal models of hepatic encephalopathy. Liver Int. Off. J. Int. Assoc. Study Liver 2021, 41, 1474–1488. [Google Scholar] [CrossRef]
- Qin, Y.; Xu, P.; Wu, T.; Qian, C.; Fan, Y.; Gen, D.; Zhu, L.; Kong, W.; Yang, H.; Xu, F.; et al. Bile duct ligation enhances AZT CNS toxicity partly by impairing the expression and function of BCRP in rat brain. Acta Pharmacol. Sin. 2020, 41, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Forrest, G.; Sills, G.J.; Leach, J.P.; Brodie, M.J. Determination of gabapentin in plasma by high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1996, 681, 421–425. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Sun, B.; Li, Y.; Xu, P.; Liu, C.; Liu, L.; Liu, X. Hyperammonemia enhances the function and expression of P-glycoprotein and Mrp2 at the blood-brain barrier through NF-κB. J. Neurochem. 2014, 131, 791–802. [Google Scholar] [CrossRef]
- Xu, P.; Ling, Z.-L.; Zhang, J.; Li, Y.; Shu, N.; Zhong, Z.-Y.; Chen, Y.; Di, X.-Y.; Wang, Z.-J.; Liu, L.; et al. Unconjugated bilirubin elevation impairs the function and expression of breast cancer resistance protein (BCRP) at the blood-brain barrier in bile duct-ligated rats. Acta Pharmacol. Sin. 2016, 37, 1129–1140. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Xu, P.; Sun, B.; Zhong, Z.; Liu, C.; Ling, Z.; Chen, Y.; Shu, N.; Zhao, K.; et al. Acute liver failure impairs function and expression of breast cancer-resistant protein (BCRP) at rat blood-brain barrier partly via ammonia-ROS-ERK1/2 activation. J. Neurochem. 2016, 138, 282–294. [Google Scholar] [CrossRef]
- Kimura, M.; Matsumura, Y.; Konno, T.; Miyauchi, Y.; Maeda, H. Enzymatic removal of bilirubin toxicity by bilirubin oxidase in vitro and excretion of degradation products in vivo. Proc. Soc. Exp. Biol. Med. 1990, 195, 64–69. [Google Scholar] [CrossRef]
- Togawa, H.; Shinkai, S.; Mizutani, T. Induction of human UGT1A1 by bilirubin through AhR dependent pathway. Drug Metab. Lett. 2008, 2, 231–237. [Google Scholar] [CrossRef]
- Sinal, C.J.; Bend, J.R. Aryl hydrocarbon receptor-dependent induction of cyp1a1 by bilirubin in mouse hepatoma hepa 1c1c7 cells. Mol. Pharmacol. 1997, 52, 590–599. [Google Scholar] [CrossRef]
- Shinde, R.; McGaha, T.L. The Aryl Hydrocarbon Receptor: Connecting Immunity to the Microenvironment. Trends Immunol. 2018, 39, 1005–1020. [Google Scholar] [CrossRef] [PubMed]
- Bock, K.W.; Köhle, C. The mammalian aryl hydrocarbon (Ah) receptor: From mediator of dioxin toxicity toward physiological functions in skin and liver. Biol. Chem. 2009, 390, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Longhi, M.S.; Vuerich, M.; Kalbasi, A.; Kenison, J.E.; Yeste, A.; Csizmadia, E.; Vaughn, B.; Feldbrugge, L.; Mitsuhashi, S.; Wegiel, B.; et al. Bilirubin suppresses Th17 immunity in colitis by upregulating CD39. JCI Insight 2017, 2, e92791. [Google Scholar] [CrossRef]
- Tomblin, J.K.; Arthur, S.; Primerano, D.A.; Chaudhry, A.R.; Fan, J.; Denvir, J.; Salisbury, T.B. Aryl hydrocarbon receptor (AHR) regulation of L-Type Amino Acid Transporter 1 (LAT-1) expression in MCF-7 and MDA-MB-231 breast cancer cells. Biochem. Pharmacol. 2016, 106, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Le Vee, M.; Jouan, E.; Lecureur, V.; Fardel, O. Aryl hydrocarbon receptor-dependent up-regulation of the heterodimeric amino acid transporter LAT1 (SLC7A5)/CD98hc (SLC3A2) by diesel exhaust particle extract in human bronchial epithelial cells. Toxicol. Appl. Pharmacol. 2016, 290, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, J.; Zielińska, M. Exchange-mode glutamine transport across CNS cell membranes. Neuropharmacology 2019, 161, 107560. [Google Scholar] [CrossRef]
- Errasti-Murugarren, E.; Palacín, M. Heteromeric Amino Acid Transporters in Brain: From Physiology to Pathology. Neurochem. Res. 2021, 1–14. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Sham | BDL |
|---|---|---|
| Body weight (BW) (g) | 238.38 ± 9.34 | 234.88 ± 18.96 |
| Liver weight (% BW) | 3.28 ± 0.18 | 6.04 ± 0.91 ** |
| Spleen weight (% BW) | 0.24 ± 0.03 | 0.53 ± 0.11 ** |
| ALT (IU/L) | 28.23 ± 5.62 | 40.75 ± 9.04 ** |
| AST (IU/L) | 51.00 ± 9.70 | 122.15 ± 46.55 ** |
| ALP (IU/L) | 358.38 ± 84.89 | 681.94 ± 207.21 ** |
| Total bilirubin (μmol/L) | <3.4 | 99.76 ± 80.11 ** |
| Total Bile Acids (μmol/L) | 34.25 ± 13.24 | 130.55 ± 73.18 ** |
| Serum ammonia (μmol/L) | 222.32 ± 16.23 | 314.77 ± 49.56 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.; Yang, H.; Qin, L.; Wang, S.; Xie, L.; Yang, L.; Kong, W.; Zhu, L.; Liu, L.; Liu, X. Bile Duct Ligation Upregulates Expression and Function of L-Amino Acid Transporter 1 at Blood–Brain Barrier of Rats via Activation of Aryl Hydrocarbon Receptor by Bilirubin. Biomedicines 2021, 9, 1320. https://doi.org/10.3390/biomedicines9101320
Zheng X, Yang H, Qin L, Wang S, Xie L, Yang L, Kong W, Zhu L, Liu L, Liu X. Bile Duct Ligation Upregulates Expression and Function of L-Amino Acid Transporter 1 at Blood–Brain Barrier of Rats via Activation of Aryl Hydrocarbon Receptor by Bilirubin. Biomedicines. 2021; 9(10):1320. https://doi.org/10.3390/biomedicines9101320
Chicago/Turabian StyleZheng, Xiaoke, Hanyu Yang, Lan Qin, Siqian Wang, Lei Xie, Lu Yang, Weimin Kong, Liang Zhu, Li Liu, and Xiaodong Liu. 2021. "Bile Duct Ligation Upregulates Expression and Function of L-Amino Acid Transporter 1 at Blood–Brain Barrier of Rats via Activation of Aryl Hydrocarbon Receptor by Bilirubin" Biomedicines 9, no. 10: 1320. https://doi.org/10.3390/biomedicines9101320
APA StyleZheng, X., Yang, H., Qin, L., Wang, S., Xie, L., Yang, L., Kong, W., Zhu, L., Liu, L., & Liu, X. (2021). Bile Duct Ligation Upregulates Expression and Function of L-Amino Acid Transporter 1 at Blood–Brain Barrier of Rats via Activation of Aryl Hydrocarbon Receptor by Bilirubin. Biomedicines, 9(10), 1320. https://doi.org/10.3390/biomedicines9101320