Intrinsic Type 1 Interferon (IFN1) Profile of Uncultured Human Bone Marrow CD45lowCD271+ Multipotential Stromal Cells (BM-MSCs): The Impact of Donor Age, Culture Expansion and IFNα and IFNβ Stimulation

 ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Cells
2.2. Sample Processing and Cell Sorting
2.3. Quantitative Polymerase Chain Reaction (qPCR)
2.4. BM-MSC Culture and IFN1 Stimulation
2.5. Intracellular ROS Measurements
2.6. Statistics
3. Results
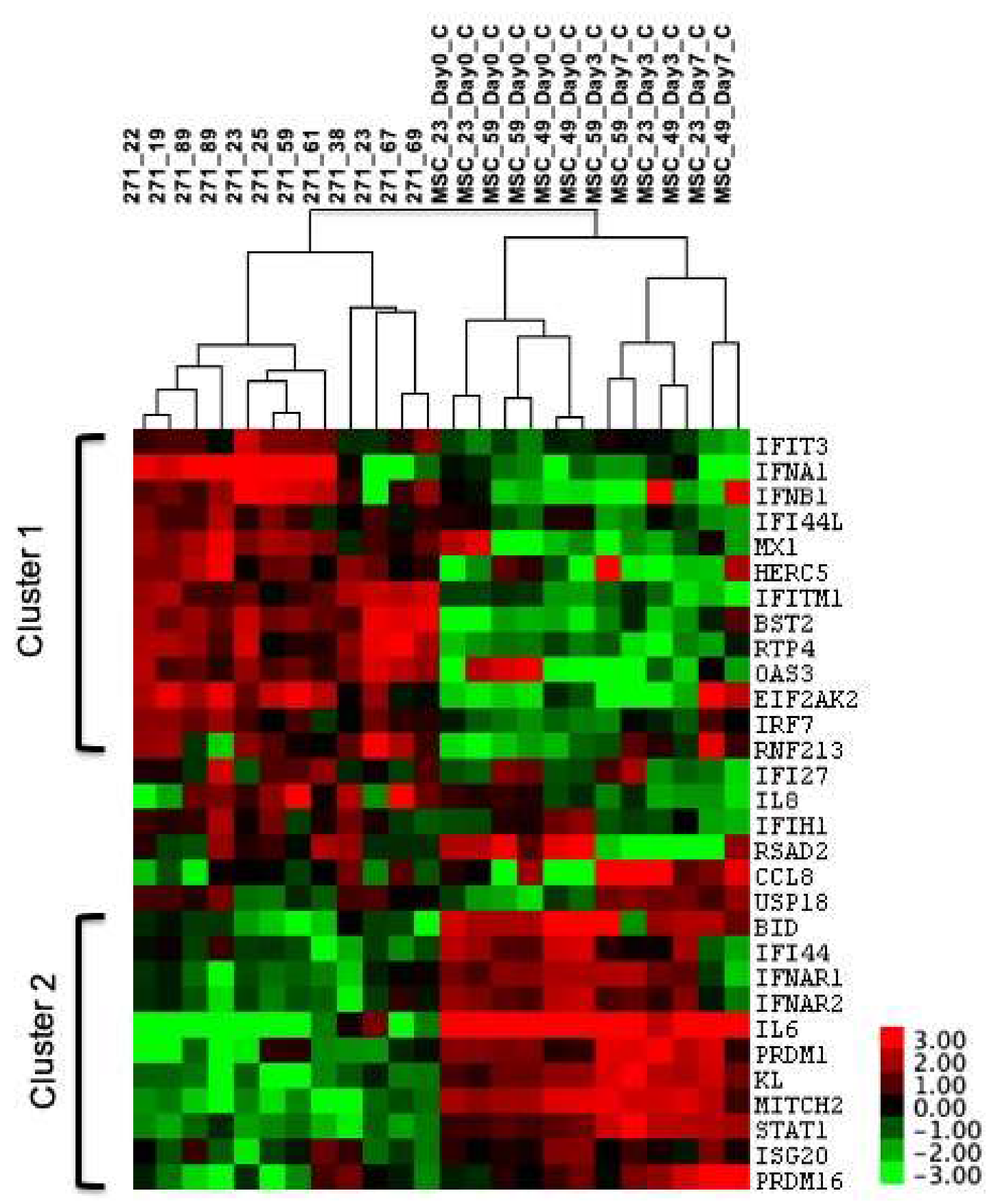
3.1. IFN1 Profile and Reduced Proliferation of Cultured BM-MSCs upon IFN1 Stimulation
3.2. IFN1 Profile of Uncultured CD45lowCD271+ BM-MSCs in Comparison to Cultured and IFN1 Stimulated BM-MSCs
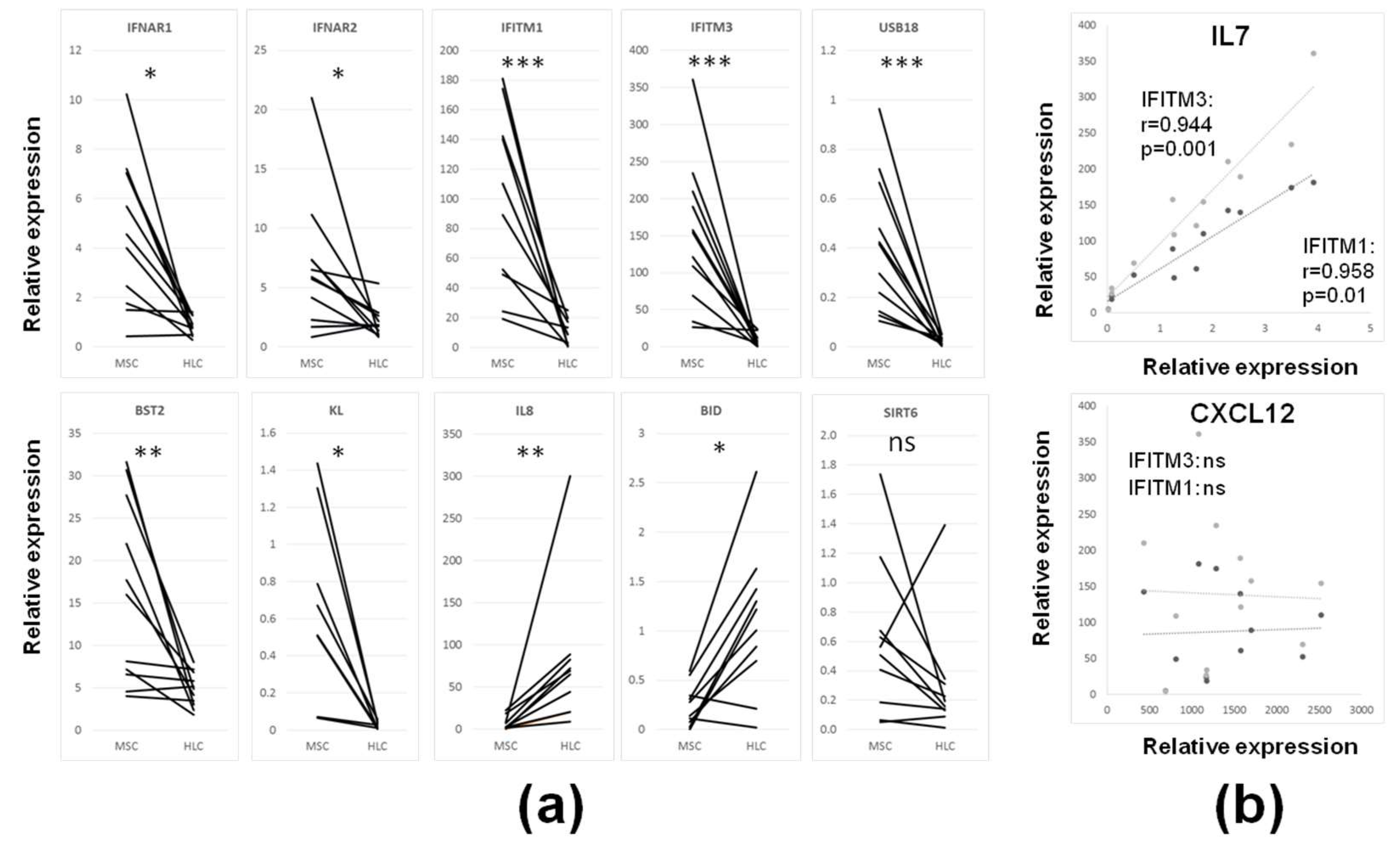
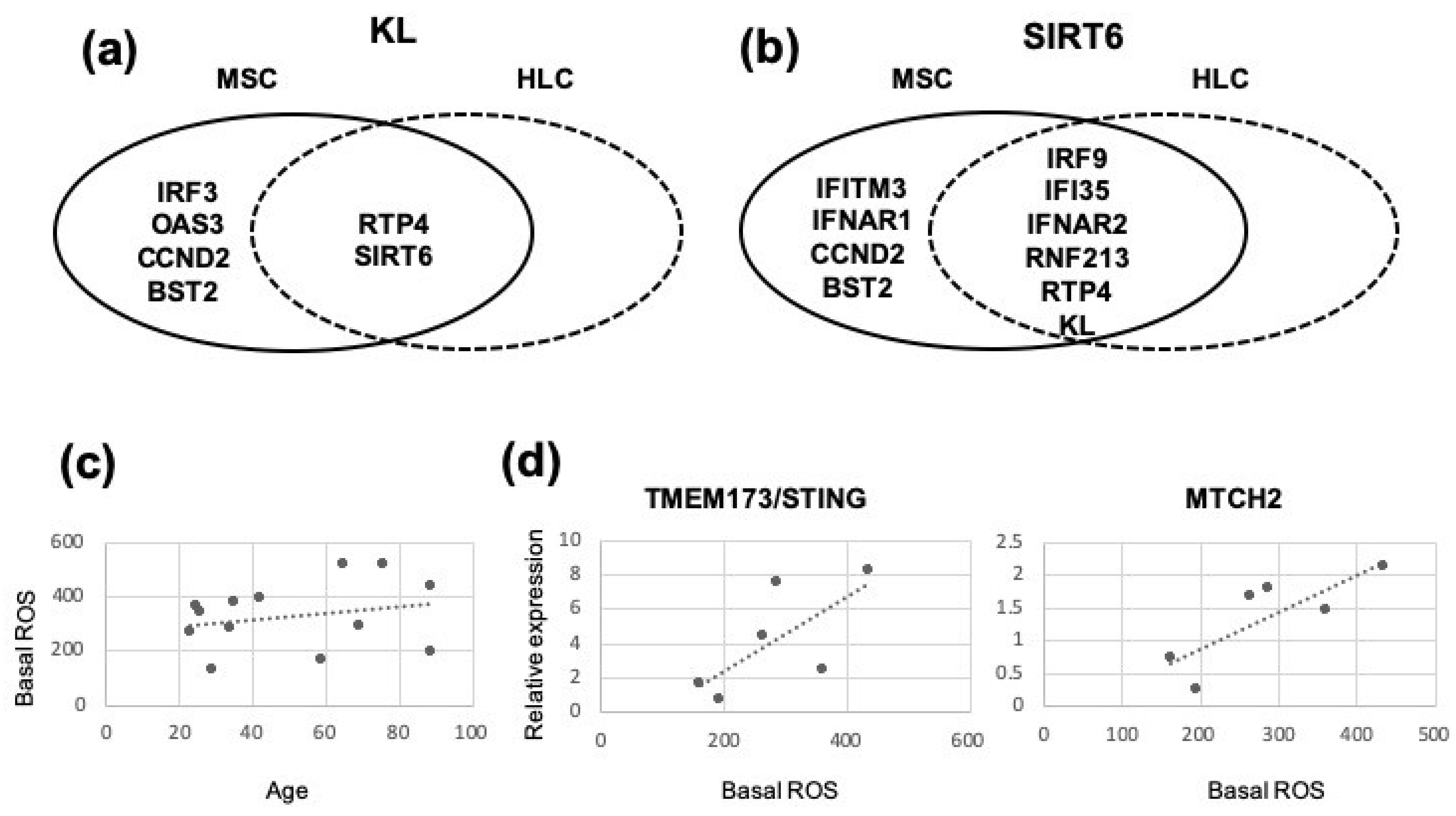
3.3. Intrinsic IFN1 Profile of Uncultured CD45lowCD271+ BM-MSCs in Comparison to Control CD45+CD271− BM-HLCs
3.4. Age-Related Changes in IFN1 Profile of CD45lowCD271+ BM-MSCs Compared to CD45+CD271− BM-HLCs
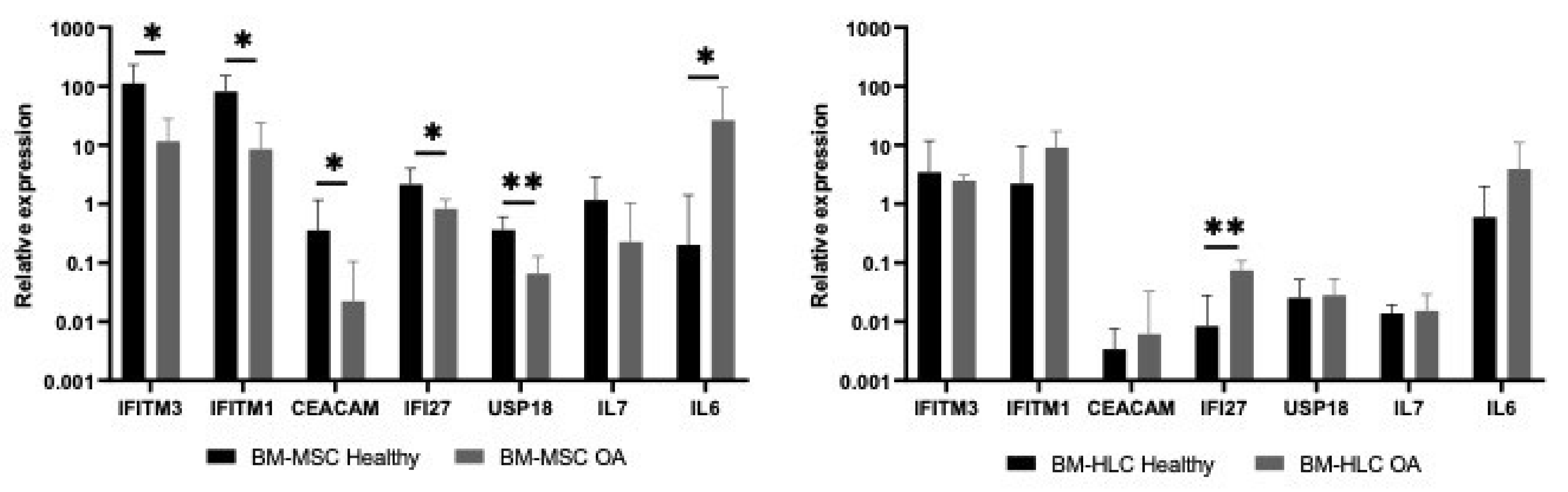
3.5. Differences in IFN1 Profile of CD45lowCD271+ BM-MSCs and CD45+CD271− BM-HLCs in Patients with Osteoarthritis (OA)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| IFN1 | Type 1 interferon |
| IFNα | Interferon alpha |
| IFNβ | Interferon beta |
| MSCs | Mesenchymal stromal cells |
| BM | Bone marrow |
| HSCs | Hematopoietic stem cells |
| KL | Klotho |
| SIRT6 | Sirtuin 6 |
| ISGs | Interferon stimulated genes |
| DNA | Deoxyribonucleic acid |
| SSEA4 | Stage specific embryonic antigen 4 |
| cGAS/STING | Cyclic GMP-AMP synthase/Stimulator of interferon genes |
| IFNAR1 | Interferon alpha receptor 1 |
| IFNAR2 | Interferon alpha receptor 2 |
| JAK | Janus kinase |
| STAT1 | Signal transducer and activator of transcription1 |
| Mx1 | MX dynamine like GTPase 1 |
| HERC5 | HECT and RLD domain containing E3 ubiquitin protein ligase 5 |
| IFTM1 | Interferon induced trans membrane protein 1 |
| IFITM3 | Interferon induced trans membrane protein 3 |
| IRF7 | Interferon regulatory factor 7 |
| RPT4 | Receptor transporter protein 4 |
| ISG20 | Inteferon stimulated gene 20 |
| BID | BH3 interacting domain |
| MTCH2 | Mitochondrial homologue carrier 2 |
| ROS | Reactive oxygen species |
| OA | Osteoarthritis |
| OAS3 | 2′-5′-oligoadenylate synthetase 3 |
| SASP | Senescence associated secretory phenotype |
| IL6 | Interleukin 6 |
| IL8 | Interleukin 8 |
| USP18 | Ubiquitin specific peptidase 18 |
| FABP4 | Fatty acid binding protein 4 |
| PPARγ | Peroxisome proliferator activator receptor gamma |
| RUNX2 | Runt related transcription factor 2 |
| SPARC | Secreted protein acid and cysteine rich/Osteonectic |
| SOX9 | SRY-Box Transcription Factor 9 |
| ACAN | Aggrecan |
| IL7 | Interleukin 7 |
| CXCL12 | C-X-C chemokine 12 |
| TNFSF11 | Tumour necrosis factor super family 11 |
| Ly6E | Lymphocyte antigen 6E |
| EIF2AK2 | Eukaryotic translation initiation factor 2-alpha kinase 2 |
| TMEM173 | Transmembrane protein 173/STING |
| IRF3 | Interferon regulatory factor 3 |
| CCND2 | CyclinD2 |
| IRF9 | Interferon regulatory factor 9 |
| IFI35 | Interferon induced 35 |
| RNF213 | Ring finger protein 213 |
| DDR | DNA-damage response |
| 3D | Three dimensional |
| SERPING1 | Serpin family G member1 |
| EDTA | Ethylene diamine tetra acetic acid |
| BMA | Bone marrow aspirate |
| DMSO | Dimethylsulfoxide |
| qPCR | quantitative Polymerised chain reaction |
| IFC | Integrated fluid circuits |
| DPBS | Dulbeccos phosphate buffer saline |
References
- Kehler, D.S. Age-related disease burden as a measure of population ageing. Lancet Public Health 2019, e123–e124. [Google Scholar] [CrossRef]
- World Health Organisation, Global Burden of Disease. 2004. Available online: http://www.who.int/healthinfo/global_burden_disease/GBD_report_2004update_full.pdf (accessed on 10 July 2020).
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- World Health Organisation, Fact Sheet Musculoskeletal Conditions. 2019. Available online: http://www.who.int/news-room/fact-sheets/detail/musculoskeletal-conditions (accessed on 10 July 2020).
- Florencio-Silva, R.; Sasso, S.G.R.; Sasso-Cerri, I.; Simões, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. BioMed. Res. Int. 2015, 1–17. [Google Scholar] [CrossRef]
- Kovtonyuk, L.V.; Fritsch, K.; Feng, X.; Manz, M.G.; Takizawa, H. Inflamm-Aging of Hematopoiesis, Hematopoietic Stem Cells, and the Bone Marrow Microenvironment. Front. Immunol. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Garagnani, P.; Vitale, G.; Capri, M.; Salvioli, S. Inflammaging and ‘Garb-aging’. Trends Endocrinol. Metab. 2017, 28, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Pang, W.W.; Schrier, S.L.; Weisman, I.L. Age-associated changes in human hematopoietic stem cells and Causes and Mechanisms of Hematopoietic Stem Cell Aging. Semin. Hematol. 2017, 54, 39–42. [Google Scholar] [CrossRef]
- Becerikli, M.; Jaurich, H.; Schira, J.; Schulte, M.; Döbele, C.; Wallner, C.; Abraham, S.; Wagner, J.M.; Dadras, M.; Kneser, U.; et al. Age-dependent alterations in osteoblast and osteoclast activity in human cancellous bone. J. Cell Mol. Med. 2017, 21, 2773–2781. [Google Scholar] [CrossRef]
- Ganani, D.; Crippa, S.; Volpe, L.D.; Rossella, V.; Conti, A.; Lettera, E.; Rivis, S.; Ometti, M.; Fraschini, G.; Bernardo, M.E.; et al. An early-senescence state in aged mesenchymal stromal cells contributes to hematopoietic stem and progenitor cell clonogenic impairment through the activation of a pro-inflammatory program. Aging Cell 2019, 18, 1–21. [Google Scholar] [CrossRef]
- Farr, J.N.; Khosla, S. Cellular senescence in bone. Bone 2019, 121, 121–133. [Google Scholar] [CrossRef]
- Pontikoglou, C.; Deschaseaux, F.; Sensebé, L.; Papadaki, H.A. Bone Marrow Mesenchymal Stem Cells: Biological Properties and Their Role in Hematopoiesis and Hematopoietic Stem Cell Transplantation. Stem Cell Rev. Rep. 2011, 7, 569–589. [Google Scholar] [CrossRef]
- Kawamura, H.; Nakatsuka, R.; Matsuoka, Y.; Sumide, K.; Fujioka, T.; Asano, H.; Iida, H.; Sonoda, Y. TGF-β Signaling Accelerates Senescence of Human Bone-Derived CD271 and SSEA-4 Double-Positive Mesenchymal Stromal Cells. Stem Cell Rep. 2018, 10, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Josephson, A.M.; Bradaschia-Correa, V.; Lee, S.; Leclerc, K.; Patel, K.S.; Lopez, E.M.; Litwa, H.P.; Neibart, S.S.; Kadiyala, M.; Wong, M.Z.; et al. Age-related inflammation triggers skeletal stem/progenitor cell dysfunction. Proc. Natl. Acad. Sci. USA 2019, 14, 6995–7004. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, P.; El-Jawhari, J.J.; Burska, A.N.; Ponchel, F.; Giannoudis, P.V.; Jones, E.A. The Analysis of In Vivo Aging in Human Bone Marrow Mesenchymal Stromal Cells Using Colony-Forming Unit-Fibroblast Assay and the CD45lowCD271+ Phenotype. Stem Cells Int. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Katlinskaya, Y.V.; Carbone, C.J.; Zhao, B.; Katlinski, K.V.; Zheng, H.; Guha, M.; Li, N.; Chen, Q.; Yang, T.; et al. DNA damage-induced type I interferon promotes senescence and inhibits stem cell function. Cell Rep. 2015, 11, 785–797. [Google Scholar] [CrossRef]
- Li, T.; Chen, Z.J. The cGAS-cGAMP-STING pathway connects DNA damage to inflammation, senescence, and cancer. J. Exp. Med. 2018, 215, 1287–1299. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Thi, V.L.D.; Huang, Y.; Billerbeck, E.; Saha, D.; Hoffmann, H.-H.; Wang, Y.; Silva, L.A.V.; Sarbanes, S.; Sun, T.; et al. Intrinsic immunity shapes viral resistance of stem cells. Cell 2018, 172, 423–438. [Google Scholar] [CrossRef]
- Kreienkamp, R.; Graziano, S.; Coll-Bonfill, N.; Bedia-Diaz, G.; Cybulla, E.; Vindigni, A.; Dorsett, D.; Kubben, N.; Batista, L.F.Z.; Gonzalo, S. A Cell-Intrinsic Interferon-like Response Links Replication Stress to Cellular Aging Caused by Progerin. Cell Rep. 2018, 22. [Google Scholar] [CrossRef]
- James, S.; Fox, J.; Afsari, F.; Lee, J.; Clough, S.; Knight, C.; Ashmore, J.; Ashton, P.; Preham, O.; Hoogduijn, M.; et al. Multiparameter Analysis of Human Bone Marrow Stromal Cells Identifies Distinct Immunomodulatory and Differentiation-Competent Subtypes. Stem Cell Rep. 2015, 4, 1004–1015. [Google Scholar] [CrossRef]
- Banchereau, J.; Pacual, V. Type I interferon in systemic lupus erythematosus and other autoimmune diseases. Immunity 2006, 25, 383–392. [Google Scholar] [CrossRef]
- Gu, Z.; Jiang, J.; Tan, W.; Xia, Y.; Cao, H.; Meng, Y.; Da, Z.; Liu, H.; Chen, C. p53/p21 Pathway involved in mediating cellular senescence of bone marrow-derived mesenchymal stem cells from systemic lupus erythematosus patients. Clin. Dev. Immunol. 2013, 2013, 1–13. [Google Scholar] [CrossRef]
- Gao, L.; Bird, A.K.; Meednu, N.; Dauenhauer, K.; Liesveld, J.; Anolik, J.; Looney, R.J. Bone Marrow-Derived Mesenchymal Stem Cells From Patients With Systemic Lupus Erythematosus Have a Senescence-Associated Secretory Phenotype Mediated by a Mitochondrial Antiviral Signaling Protein-Interferon-β Feedback Loop. Arthritis Rheumatol. 2017, 69, 1623–1635. [Google Scholar] [CrossRef]
- Churchman, S.M.; Ponchel, F.; Boxall, S.A.; Cuthbert, R.; Kouroupis, D.; Roshdy, T.; Giannoudis, P.V.; Emery, P.; McGonagle, D.; Jones, E.A. Transcriptional Profile of Native CD271+ Multipotential Stromal Cells Evidence for Multiple Fates, With Prominent Osteogenic and Wnt Pathway Signaling Activity. Arthritis Rheum. 2012, 64, 2632–2643. [Google Scholar] [CrossRef]
- Ghazanfari, R.; Zacharaki, D.; Li, H.; Lim, H.C.; Soneji, S.; Scheding, S. Human Primary Bone Marrow Mesenchymal Stromal Cells and Their in Vitro Progenies Display Distinct Transcriptional Profile Signatures. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Weber, A.; Chan, P.M.B.; Wen, C. Do immune cells lead the way in subchondral bone disturbance on osteoarthritis? Prog. Biophys. Mol. Biol. 2019, 148, 21–31. [Google Scholar] [CrossRef]
- Ni, Z.; Zhao, X.; Dai, X.; Zhao, L.; Xia, J. The role of interferon regulatory factor 5 in macrophage inflammation during osteoarthritis. Inflammation 2019, 42, 1821–1829. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.; English, A.; Churchman, S.M.; Kouroupis, D.; Boxall, S.A.; Kinsey, S.; Giannoudis, P.V.; Emery, P.; McGonagle, D. Large-scale extraction and characterization of CD271+ multipotential stromal cells from trabecular bone in health and osteoarthritis: Implications for bone regeneration strategies based on uncultured or minimally cultured multipotential stromal cells published correction appears in Arthritis Rheum. Arthritis Rheum. 2010, 62, 1944–1954. [Google Scholar] [CrossRef]
- Ilas, D.C.; Churchman, S.M.; Baboolal, T.G.; Giannoudis, P.V.; Aderintho, J.; McGonagle, D.; Jones, E. The simultaneous analysis of mesenchymal stem cells and early osteocytes accumulation in osteoarthritic femoral head sclerotic bone. Rheumatology 2019, 58, 1777–1783. [Google Scholar] [CrossRef] [PubMed]
- El-Jawhari, J.J.; Ganguly, P.; Churchman, S.M.; Jones, E.; Giannnoudis, P.V. The biological fitness of bone progenitor cells in reamer/irrigator/aspirator waste. J. Bone Joint Surg. Am. 2019, 101, 2111–2119. [Google Scholar] [CrossRef] [PubMed]
- Purcell, M.; Kruger, A.; Tainsky, M.A. Gene expression profiling of replicative and induced senescence. Cell Cycle 2014, 15, 3927–3937. [Google Scholar] [CrossRef]
- Oreffo, R.O.; Romberg, S.; Virdi, A.S.; Joyner, C.J.; Berven, S.; Triffitt, J.T. Effects of interferon alpha on human osteoprogenitor cell growth and differentiation in vitro. J. Cell. Biochem. 1999, 74, 372–385. [Google Scholar] [CrossRef]
- Hatzfeld, A.; Eid, P.; Peiffer, I.; Li, M.L.; Barbet, R.; Oostendorp, R.A.J.; Haydont, V.; Monier, M.-N.; Milon, L.; Fortunel, N.; et al. A Sub-Population of High Proliferative Potential-Quiescent Human Mesenchymal Stem Cells Is Under the Reversible Control of Interferon Alpha/Beta. Leukemia 2007, 21, 714–724. [Google Scholar] [CrossRef]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-stimulated gene: A complex web of host defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef]
- El-Sherbiny, Y.; El-Jawhari, J.J.; Moseley, T.A.; McGonagle, D.; Jones, E. T Cell Immunomodulation by Clinically Used Allogeneic Human Cancellous Bone Fragments: A Potential Novel Immunotherapy Tool. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, F.; Borger, D.K.; Wei, Q.Z.; Pinho, S.; Maryanovich, M.; Zahalka, A.H.; Suzuki, M.; Cruz, C.D.; Wang, Z.C.; Xu, C.L.; et al. Engineering a haematopoietic stem cell niche by revitalizing mesenchymal stromal cells. Nat. Cell Biol. 2019, 21, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Tadogan, A.; Kumar, S.; Allies, G.; Bausinger, J.; Beckel, F.; Hofemeister, H.; Mulaw, M.; Madan, V.; Scharffetter-Kochanek, K.; Feuring-Buske, M.; et al. DNA Damage-Induced HSPC Malfunction Depends on ROS Accumulation Downstream of IFN-1 Signaling and Bid Mobilization. Cell Stem Cell 2016, 19, 752–767. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, K. Molecular Profiles of Cell-to-Cell Variation in the Regenerative Potential of Mesenchymal Stromal Cells. Stem Cells Int. 2019, 2019, 1–14. [Google Scholar] [CrossRef]
- Flint, S.; Jovanovic, V.; Teo, B.W.; Mak, A.; Thumboo, J.; McKinney, E.F.; Lee, J.C.; MacAry, P.; Kemeny, D.M.; Jayne, D.; et al. Leucocyte subset-specific type 1 interferon signatures in SLE and other immune-mediated diseases. RMD 2016, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wu, J.; Du, F.; Chen, X.; Chen, Z.J. Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 2013, 339, 1–15. [Google Scholar] [CrossRef]
- Liu, F.; Wu, S.; Ren, H.; Gu, J. Klotho supresses RIG-1 mediated senescence associated inflammation. Nat. Cell Biol. 2011, 13, 254–262. [Google Scholar] [CrossRef]
- Cordeirogomes, A.; Hara, T.; Lim, V.Y.; Herndler-Brandstetter, D.; Nevius, E.; Sugiyama, T.; Tani-Ichi, S.; Schlenner, S.; Richie, E.; Rodewald, H.-R.; et al. Hematopoietic Stem Cell Niches Produce Lineage Instructive Signals to Control Multipotent Progenitor Differentiation. Immunity 2016, 45, 1219–1231. [Google Scholar] [CrossRef]
- Naismith, E.N.; Pangrazzi, L. The impact of oxidative stress, inflammation, and senescence on the maintenance of immunological memory in the bone marrow in old age. Biosci. Rep. 2019, 39, 1–12. [Google Scholar] [CrossRef]
- Sugiyama, T.; Kohara, H.; Noda, M.; Nagasawa, T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 2006, 25, 977–988. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.; Tasdogan, A.; Fehling, H.J. The IFN-1 > BID > ROS pathway: Linking DNA damage with HSPC malfunction. Cell Cycle 2017, 16, 819–820. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, B.C.; Sarhan, J.; Poltorak, A. Host-intrinsic interferon status in infection and immunity. Trend Mol. Med. 2019, 24, 658–668. [Google Scholar] [CrossRef]
- Jazwinski, S.M.; Kim, S. Examination of the Dimensions of Biological Age. Front. Genet. 2019, 10, 1–7. [Google Scholar] [CrossRef]
- Doss, F.; Menard, J.; Hauschild, M.; Kreutzer, H.-J.; Mittlmeier, T.; Müller-Steinhardt, M.; Müller, B. Elevated IL-6 levels in the synovial fluid of osteoarthritis patients stem from plasma cells. Scand. J. Rheumatol. 2007, 36, 136–139. [Google Scholar] [CrossRef]
- Honke, N.; Shaabani, N.; Zhang, D.-E.; Hardt, C.; Lang, K.S. Multiple functions of USP18. Cell Death Dis. 2016, 7, e2444. [Google Scholar] [CrossRef]
- Li, Y.; Ma, M.X.; Qin, B.; Lin, L.-T.; Richardson, C.D.; Feld, J.; McGilvray, I.D.; Chen, L. The Ubiquitin-Specific Protease 18 Promotes Hepatitis C Virus Production by Increasing Viral Infectivity. Mediat. Inflamm. 2019, 3124745. [Google Scholar] [CrossRef]
- Schreiber, G. The molecular basis for differential type I interferon signaling. J. Biol. Chem. 2017, 292, 7285–7294. [Google Scholar] [CrossRef] [PubMed]
- Woeckel, V.J.; Eijken, M.; van de Peppel, J.; Chiba, H.; van der Eerden, B.C.J.; van Leeuwen, J.P.T.M. IFNβ impairs extracellular matrix formation leading to inhibition of mineralization by effects in the early stage of human osteoblast differentiation. J. Cell Physiol. 2012, 227, 2668–2676. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; MacArthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef]
- Hiroshi, T.; Kojiro, S.; Akinori, T.; Tadatsugu, T. Interplay between interferon and other cytokine systems in bone metabolism. Immunol. Rev. 2005, 208, 181–193. [Google Scholar] [CrossRef]
- Takeuchi, T.; Shimakawa, G.; Tamura, M.; Yokosawa, H.; Arata, Y. ISG15 Regulates RANKL-induced Osteoclastogenic Differentiation of RAW264 Cells. Biol. Pharm Bull. 2015, 38, 452–486. [Google Scholar] [CrossRef]
- Van Roon, J.A.G.; Lafeber, F.P.J.G. Role of interleukin-7 in degenerative and inflammatory joint diseases. Arthritis Res. Ther. 2008, 10. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhi, X.; Wang, J.; Su, J. RANKL Signaling in Bone Marrow Mesenchymal Stem Cells Negatively Regulates Osteoblastic Bone Formation. Bone Res. 2018, 6, 34. [Google Scholar] [CrossRef]
- Tormin, A.; Li, Q.; Brune, J.C.; Walsh, S.; Schütz, B.; Ehinger, M.; Ditzel, N.; Kassem, M.; Scheding, S. CD146 expression on primary nonhematopoietic bone marrow stem cells is correlated with in situ localization. Blood 2011, 117, 5067–5077. [Google Scholar] [CrossRef]
- Souliotis, V.L.; Vlachogiannis, N.I.; Pappa, M.; Argyriou, A.; Ntouros, P.A.; Sfikakis, P.P. DNA Damage Response and Oxidative Stress in Systemic Autoimmunity. Int. J. Mol. Sci. 2020, 21, 55. [Google Scholar] [CrossRef]
- Harichandan, A.; Buhring, H.-J. Prospective isolation of human MSCs. Best Pract. Res. Clin. Haematol. 2011, 24, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Mo, M.; Wang, S.; Zhou, Y.; Li, H.; Wu, Y. Mesenchymal stem cell subpopulations: Phenotype, property and therapeutic potential. Cell. Mol. Life Sci. 2016, 73, 3311–3321. [Google Scholar] [CrossRef]
- Wolock, S.L.; Krishnan, I.; Tenen, D.E.; Matkins, V.; Camacho, V.; Patel, S.; Agarwal, P.; Bhatia, R.; Tenen, D.G.; Klein, A.M.; et al. Mapping Distinct Bone Marrow Niche Populations and Their Differentiation Paths. Cell Rep. 2019, 28, 302–311. [Google Scholar] [CrossRef]
- Gough, D.J.; Messina, N.L.; Clarke, C.J.P.; Johnstone, R.W.; Levy, D.E. Constitutive type I interferon modulates homeostatic balance through tonic signaling. Immunity 2013, 36, 166–174. [Google Scholar] [CrossRef]
- Childs, B.; van Deursen, J. An evolving picture of cell senescence. Nature 2019, 566, 46–47. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, E.T.; Gonzalez-Nieto, D.; Ghiaur, G.; Dunn, S.K.; Ficker, A.M.; Murali, B.; Madhu, M.; Gutstein, D.E.; Fishman, G.I.; Barrio, L.C.; et al. Connexin-43 prevents hematopoietic stem cell senescence through transfer of reactive oxygen species to bone marrow stromal cells. Proc. Natl. Acad. Sci. USA 2012, 109, 9071–9076. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Id | MSC (Median) | HLC (Median) | Fold Difference | p Value |
|---|---|---|---|---|---|
| SERPING | Serine protease inhibitor, clade G (C1 inhibitor), member 1 | 22 | 0.094 | 233.3 | 0.0003 |
| IFI27 | Interferon gamma-inducible protein 27 | 1.551 | 0.01 | 155.1 | <0.0001 |
| PRDM16 | PR domain containing 16 | 0.179 | 0.002 | 105.3 | 0.042 |
| IL7 | Interleukin 7 | 1.481 | 0.016 | 93.7 | 0.0005 |
| CEACAM | Carcinoembryonic antigen-related cell adhesion molecule 1 | 0.31 | 0.007 | 44.3 | 0.0021 |
| IFITM3 | Interferon induced transmembrane protein 3 | 137.9 | 3.79 | 36.4 | <0.0001 |
| KL | Klotho | 0.51 | 0.02 | 25.5 | 0.0083 |
| USP18 | Ubiquitin specific peptidase 18 | 0.35 | 0.02 | 17.5 | <0.0001 |
| LRP1 | LDL Receptor Related Protein 1 | 48.92 | 4.03 | 12.1 | 0.0007 |
| RTP4 | Receptor (chemosensory) transporter protein 4 | 0.96 | 0.08 | 12 | 0.0036 |
| IFITM1 | Interferon induced transmembrane protein 1 | 75.05 | 6.33 | 11.9 | 0.0005 |
| LY6E | Lymphocyte antigen 6 complex, locus E | 24.58 | 2.4 | 10.2 | <0.0001 |
| SPATS2L | Spermatogenesis associated, serine-rich 2-like | 3.509 | 0.405 | 8.7 | 0.0001 |
| IFI6 | Interferon inducible alpha protein 6 | 3.285 | 0.438 | 7.5 | 0.0145 |
| IFIT5 | Interferon induced protein with tetratricopeptide repeats 5 | 0.633 | 0.11 | 5.8 | 0.0284 |
| CCND2 | CyclinD2 | 2.359 | 0.415 | 5.7 | 0.0013 |
| SOCS1 | Suppressor of cytokine signalling 3 | 2.558 | 0.461 | 5.6 | 0.0003 |
| ABCA1 | ATP Binding Cassette Subfamily A Member 1 | 1.173 | 0.24 | 4.9 | 0.0249 |
| IFNAR1 | Interferon alpha receptor 1 | 4.28 | 0.88 | 4.9 | 0.0018 |
| IFI44L | Interferon induced protein 44 like | 1.507 | 0.31 | 4.9 | 0.008 |
| OASL | 2′-5′-oligoadenylate synthetase | 1.014 | 0.22 | 4.6 | 0.0284 |
| PLSCR1 | Phospholipid scramblase 1 | 4.117 | 0.998 | 4.1 | 0.0068 |
| EIF2AK2 | Eukaryotic translation initiation factor 2-alpha kinase 2 | 3.807 | 0.956 | 4 | 0.0023 |
| OAS3 | 2′-5′-oligoadenylate synthetase 3 | 2.55 | 0.7 | 3.6 | 0.0121 |
| IFI35 | Interferon inducible alpha protein 35 | 2.68 | 0.76 | 3.5 | 0.0387 |
| NT5C3B | 5′-nucleotidase cytosolic IIIB | 0.56 | 0.159 | 3.5 | 0.0018 |
| RSAD2 | Radical S-adenosyl methionine domain containing 2 | 0.434 | 0.132 | 3.3 | 0.0169 |
| IFNAR2 | Interferon alpha receptor 2 | 5.82 | 1.84 | 3.2 | 0.0332 |
| IFI44 | Interferon inducible alpha protein 44 | 3.1 | 1.01 | 3.1 | 0.0205 |
| EPSTI1 | Epithelial stromal interaction 1 | 1.59 | 0.57 | 2.8 | 0.0045 |
| IFIH1 | Interferon induced with helicase C domain 1 | 0.45 | 0.17 | 2.7 | 0.0106 |
| UBE2L6 | Ubiquitin/ISG15-conjugating enzyme E2 L6 | 11.86 | 4.5 | 2.6 | 0.0083 |
| SAMD9L | Sterile alpha motif domain containing 9-like | 1.67 | 0.642 | 2.6 | 0.0387 |
| BST2 | Bone marrow stromal antigen 2, Tetherin (CD317) | 12.06 | 4.97 | 2.4 | 0.0083 |
| IFI16 | Interferon inducible alpha protein 16 | 1.664 | 0.809 | 2.1 | 0.0068 |
| RGS1 | Regulator of G protein signalling 1 | 0.621 | 32.29 | 52 | 0.0184 |
| IL8 | Interleukin 8 | 2.919 | 67.21 | 23 | 0.0022 |
| TNF | Tumour necrosis factor | 0.17 | 3.67 | 21.6 | 0.013 |
| IRF5 | Interferon regulatory factor 5 | 0.129 | 1.285 | 10 | 0.0157 |
| HPSE | Heparanase | 0.097 | 0.732 | 7.6 | 0.0171 |
| FCGR1B | Fc fragment of IgG receptor 1B | 0.19 | 1.04 | 5.5 | 0.0031 |
| BID | BH3 interacting domain | 0.21 | 1 | 4.8 | 0.0188 |
| PRDM1 | PR domain containing 1 | 0.51 | 1.72 | 3.4 | 0.0148 |
| LAIR1 | Leukocyte associated immunoglobulin like receptor1 | 0.43 | 1.4 | 3.3 | 0.0278 |
| CASP1 | Caspase1 | 3.06 | 6.16 | 2 | 0.0205 |
| Gene ID | Gene Name | Young (Median) | Old (Median) | Fold Difference | p Value |
|---|---|---|---|---|---|
| IL7R | Interleukin7 receptor | 0.491 | 0.098 | 5.01 | 0.026 |
| PRKRA | Protein Kinase, Interferon-Inducible Double Stranded RNA Dependent Activator | 0.970 | 0.200 | 4.85 | 0.0152 |
| SCARB1 | Scavenger Receptor Class B Member 1 | 0.431 | 0.131 | 3.29 | 0.0087 |
| CXCL10 | C-X-C motif chemokine 10 | 0.580 | 0.180 | 3.22 | 0.0667 |
| RTP4 | Receptor transporter protein 4 | 0.150 | 0.050 | 3.00 | 0.0152 |
| CEACAM | Carcinoembryonic antigen-related cell adhesion molecule 1 | 0.008 | 0.003 | 2.67 | 0.0667 |
| Tp53 | Tumour protein 53 | 2.930 | 1.210 | 2.42 | 0.0043 |
| IRF9 | Interferon regulatory factor 9 | 1.520 | 0.628 | 2.42 | 0.026 |
| IRF3 | Interferon regulatory factor 3 | 1.561 | 0.660 | 2.37 | 0.0411 |
| ABCG1 | ATP Binding Cassette Subfamily G Member 1 | 0.070 | 0.030 | 2.33 | 0.0649 |
| TMEM173/STING | Transmembrane protein 173/Stimulator of interferon genes | 2.293 | 1.033 | 2.22 | 0.026 |
| IFIT5 | Interferon induced protein with tetratricopeptide repeats 5 | 0.124 | 0.058 | 2.14 | 0.0411 |
| MTCH2 | Mitochondrial carrier homolog 2 | 1.610 | 0.790 | 2.04 | 0.0152 |
| Gene ID | Gene Name | MSC (Median) | HLC (Median) | Fold Difference | p Value |
|---|---|---|---|---|---|
| PRDM16 | PR domain containing 16 | 0.11 | 0.001 | 110 | 0.055 |
| SERPING | Serine protease inhibitor, clade G (C1 inhibitor), member 1 | 8.8 | 0.086 | 102.3 | 0.0006 |
| Kl | Klotho | 0.1 | 0.002 | 50 | 0.0175 |
| LRP1 | LDL Receptor Related Protein 1 | 8 | 0.31 | 25.8 | 0.0111 |
| IL7 | Interleukin 7 | 0.22 | 0.015 | 14.7 | 0.0012 |
| IFI27 | Interferon gamma-inducible protein 27 | 0.82 | 0.07 | 11.7 | 0.0006 |
| OAS1 | 2′-5′-oligoadenylate synthetase 1 | 0.23 | 0.024 | 9.6 | 0.022 |
| CHMP5 | Charged multivesicular body protein 5 | 1.78 | 0.21 | 8.5 | 0.0728 |
| IL6 | Interleukin 6 | 26.57 | 3.93 | 6.8 | 0.0728 |
| SPATS2L | Spermatogenesis associated, serine-rich 2-like | 2.12 | 0.317 | 6.7 | 0.0012 |
| TLR4 | Toll Like Receptor 4 | 0.88 | 0.16 | 5.5 | 0.011 |
| NT5C3B | 5′-nucleotidase cytosolic IIIB | 0.37 | 0.07 | 5.3 | 0.0041 |
| CCL8 | Chemokine C-C motif ligand 8 | 0.12 | 0.023 | 5.2 | 0.0006 |
| CXCL10 | C-X-C motif chemokine 10 | 0.56 | 0.11 | 5.1 | 0.038 |
| ABCA1 | ATP Binding Cassette Subfamily A Member 1 | 2.88 | 0.59 | 4.9 | 0.0728 |
| IFITM3 | Interferon induced transmembrane protein 3 | 11.6 | 2.47 | 4.7 | 0.026 |
| IFI35 | Interferon alpha-inducible protein 35 | 1.29 | 0.28 | 4.6 | 0.0728 |
| RTP4 | Receptor (chemosensory) transporter protein 4 | 0.12 | 0.03 | 4 | 0.0728 |
| PRKRA | Protein Kinase, Interferon-Inducible Double Stranded RNA Dependent Activator | 0.83 | 0.21 | 4.0 | 0.073 |
| GBP1 | Guanylate binding protein 1 | 3.3 | 0.84 | 3.9 | 0.038 |
| STING | Stimulator of interferon genes | 1.25 | 0.46 | 2.7 | 0.073 |
| PLSCR1 | Phospholipid scramblase 1 | 2.04 | 0.81 | 2.5 | 0.0262 |
| IFI44L | Interferon induced protein 44 like | 0.35 | 0.16 | 2.2 | 0.0262 |
| IFIT5 | Interferon induced protein with tetratricopeptide repeats 5 | 0.26 | 0.12 | 2.2 | 0.0006 |
| IFI16 | Interferon inducible alpha protein 16 | 1.164 | 0.56 | 2.1 | 0.0041 |
| IFNAR2 | Interferon alpha receptor 2 | 1.36 | 0.66 | 2.1 | 0.035 |
| IFNG | Interferon gamma | 0.003 | 1.29 | 430 | 0.0167 |
| RGS1 | Regulator of G protein signalling 1 | 0.18 | 21.98 | 122.1 | 0.0006 |
| LAMP3 | Lysosome associated membrane glycoprotein 3 | 0.02 | 0.48 | 24 | 0.0023 |
| FCGR1B | Fc fragment of IgG receptor IB | 0.003 | 0.065 | 21.7 | 0.0513 |
| IL7R | Interleukin 7 receptor | 0.19 | 2.89 | 15.2 | 0.0012 |
| SIGLEC1 | Sialic acid binding Ig like Lectin 1 | 0.008 | 0.08 | 10 | 0.0714 |
| PRDM1 | PR domain containing 1, with ZNF domain | 0.71 | 3.6 | 5.1 | 0.011 |
| LAIR1 | Leukocyte associated immunoglobulin like receptor1 | 0.13 | 0.61 | 4.7 | 0.0023 |
| TGFB | PR domain containing 1, with ZNF domain | 2.9 | 13.23 | 4.6 | 0.0012 |
| ISG20 | Interferon stimulated exonuclease gene 20kDa | 1.17 | 5.081 | 4.3 | 0.0006 |
| HPSE | Heparanase | 0.06 | 0.16 | 2.7 | 0.053 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganguly, P.; Burska, A.N.; Davis, C.L.M.; El-Jawhari, J.J.; Giannoudis, P.V.; Jones, E.A. Intrinsic Type 1 Interferon (IFN1) Profile of Uncultured Human Bone Marrow CD45lowCD271+ Multipotential Stromal Cells (BM-MSCs): The Impact of Donor Age, Culture Expansion and IFNα and IFNβ Stimulation. Biomedicines 2020, 8, 214. https://doi.org/10.3390/biomedicines8070214
Ganguly P, Burska AN, Davis CLM, El-Jawhari JJ, Giannoudis PV, Jones EA. Intrinsic Type 1 Interferon (IFN1) Profile of Uncultured Human Bone Marrow CD45lowCD271+ Multipotential Stromal Cells (BM-MSCs): The Impact of Donor Age, Culture Expansion and IFNα and IFNβ Stimulation. Biomedicines. 2020; 8(7):214. https://doi.org/10.3390/biomedicines8070214
Chicago/Turabian StyleGanguly, Payal, Agata N. Burska, Charlotte L.M. Davis, Jehan J. El-Jawhari, Peter V. Giannoudis, and Elena A. Jones. 2020. "Intrinsic Type 1 Interferon (IFN1) Profile of Uncultured Human Bone Marrow CD45lowCD271+ Multipotential Stromal Cells (BM-MSCs): The Impact of Donor Age, Culture Expansion and IFNα and IFNβ Stimulation" Biomedicines 8, no. 7: 214. https://doi.org/10.3390/biomedicines8070214
APA StyleGanguly, P., Burska, A. N., Davis, C. L. M., El-Jawhari, J. J., Giannoudis, P. V., & Jones, E. A. (2020). Intrinsic Type 1 Interferon (IFN1) Profile of Uncultured Human Bone Marrow CD45lowCD271+ Multipotential Stromal Cells (BM-MSCs): The Impact of Donor Age, Culture Expansion and IFNα and IFNβ Stimulation. Biomedicines, 8(7), 214. https://doi.org/10.3390/biomedicines8070214