

Targeting the ZMYM2-ANXA9 Axis with FLT3 Inhibitor G749 Overcomes Oxaliplatin Resistance in Colorectal Cancer
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Method
2.1. Human Specimens
2.2. Mice
2.3. Cell Culture
2.4. Clonogenic Survival Assays
2.5. Drugs
2.6. ChIP–Quantitative PCR
2.7. Flow Cytometric Analysis
2.8. Western Blotting
2.9. Immunohistochemistry Staining
2.10. RNA Interference
2.11. Real-Time Quantitative PCR
2.12. Organoid Culture
2.13. Statistical Analysis
2.14. Replicates and Experimental Design
3. Result
3.1. Elevated ANXA9 Expression Is Correlated with Chemoresistance and Poor Prognosis
3.2. High Expression of ANXA9 Promotes Malignant Phenotypes in CRC Cells
3.3. FLT3 Inhibitor G749 Suppresses the Expression of ANXA9
3.4. G749 Inhibits Proliferation of CRC Tumor Both In Vitro and In Vivo
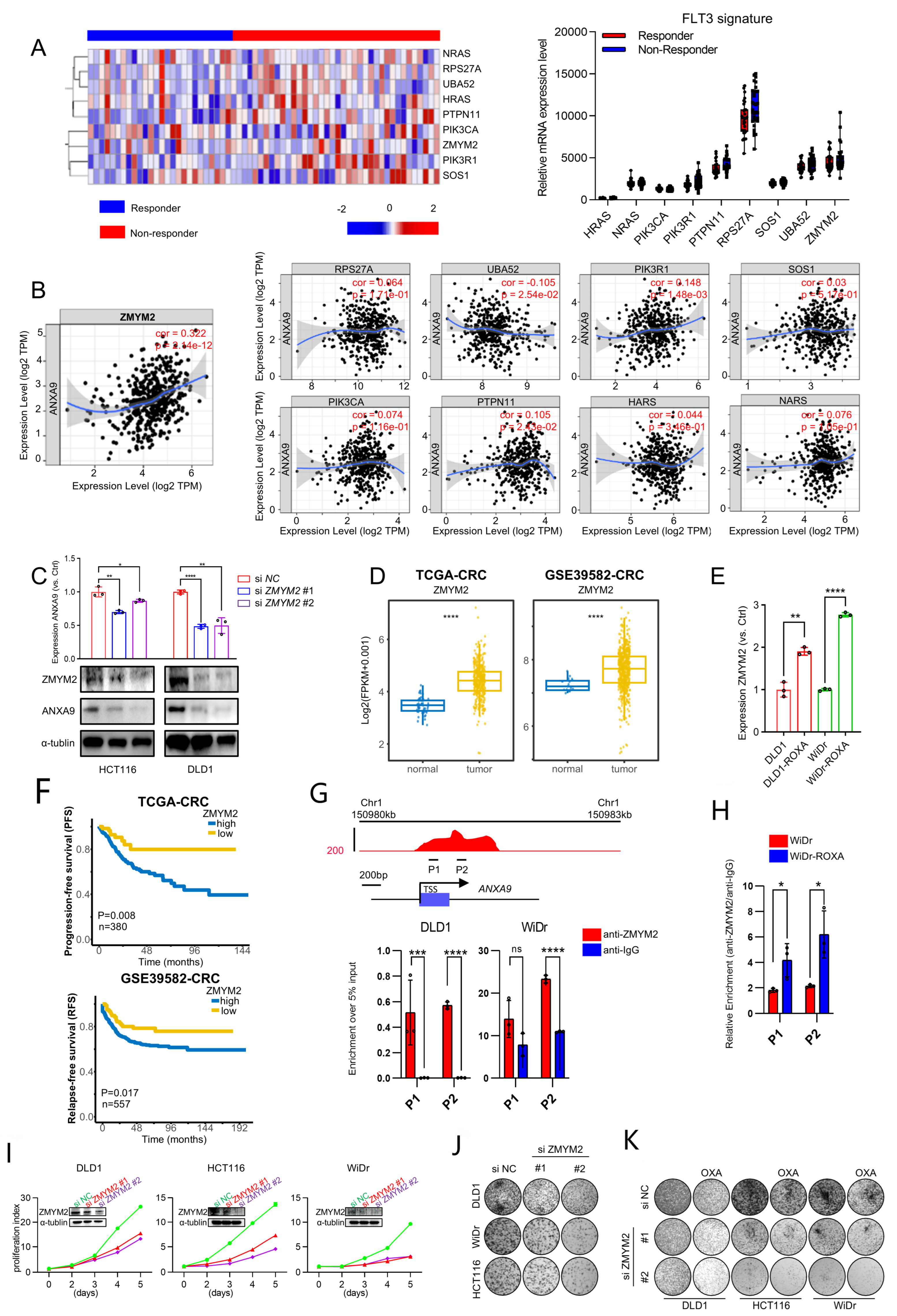
3.5. ZMYM2 Drives the Malignant Phenotypes in CRC Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Taieb, J.; Kourie, H.R.; Emile, J.F.; Le Malicot, K.; Balogoun, R.; Tabernero, J.; Mini, E.; Folprecht, G.; Van Laethem, J.L.; Mulot, C.; et al. Association of Prognostic Value of Primary Tumor Location in Stage III Colon Cancer with RAS and BRAF Mutational Status. JAMA Oncol. 2018, 4, e173695. [Google Scholar] [CrossRef] [PubMed]
- Marin, J.J.; Sanchez de Medina, F.; Castaño, B.; Bujanda, L.; Romero, M.R.; Martinez-Augustin, O.; Moral-Avila, R.D.; Briz, O. Chemoprevention, chemotherapy, and chemoresistance in colorectal cancer. Drug Metab. Rev. 2012, 44, 148–172. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.A.; Chen, Y.F.; Bao, Y.; Mahara, S.; Yatim, S.M.J.M.; Oguz, G.; Lee, P.L.; Feng, M.; Cai, Y.; Tan, E.Y.; et al. Hypoxic tumor microenvironment activates GLI2 via HIF-1α and TGF-β2 to promote chemoresistance in colorectal cancer. Proc. Natl. Acad. Sci. USA 2018, 115, E5990–E5999. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Li, Z.; Lee, P.L.; Guan, P.; Aau, M.Y.; Lee, S.T.; Feng, M.; Lim, C.Z.; Lee, E.Y.; Wee, Z.N.; et al. PDK1 signaling toward PLK1-MYC activation confers oncogenic transformation, tumor-initiating cell activation, and resistance to mTOR-targeted therapy. Cancer Discov. 2013, 3, 1156–1171. [Google Scholar] [CrossRef]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef]
- Fletcher, T.; Thompson, A.J.; Ashrafian, H.; Darzi, A. The measurement and modification of hypoxia in colorectal cancer: Overlooked but not forgotten. Gastroenterol. Rep. 2022, 10, goac42. [Google Scholar] [CrossRef]
- Wood, G.E.; Hockings, H.; Hilton, D.M.; Kermorgant, S. The role of MET in chemotherapy resistance. Oncogene 2021, 40, 1927–1941. [Google Scholar] [CrossRef]
- Shin, J.K.; Huh, J.W.; Lee, W.Y.; Yun, S.H.; Kim, H.C.; Cho, Y.B.; Park, Y.A. Clinical prediction model of pathological response following neoadjuvant chemoradiotherapy for rectal cancer. Sci. Rep. 2022, 12, 7145. [Google Scholar] [CrossRef]
- Stockton, J.D.; Tee, L.; Whalley, C.; James, J.; Dilworth, M.; Wheat, R.; Nieto, T.; S-CORT Consortium; Geh, I.; Barros-Silva, J.D.; et al. Complete response to neoadjuvant chemoradiotherapy in rectal cancer is associated with RAS/AKT mutations and high tumour mutational burden. Radiat. Oncol. 2021, 16, 129. [Google Scholar] [CrossRef]
- Cercek, A.; Dos Santos Fernandes, G.; Roxburgh, C.S.; Ganesh, K.; Ng, S.; Sanchez-Vega, F.; Yaeger, R.; Segal, N.H.; Reidy-Lagunes, D.L.; Varghese, A.M.; et al. Mismatch Repair-Deficient Rectal Cancer and Resistance to Neoadjuvant Chemotherapy. Clin. Cancer Res. 2020, 26, 3271–3279. [Google Scholar] [CrossRef]
- Yuan, Y.; Sun, W.; Xie, J.; Zhang, Z.; Luo, J.; Han, X.; Xiong, Y.; Yang, Y.; Zhang, Y. RNA nanotherapeutics for hepatocellular carcinoma treatment. Theranostics 2025, 15, 965–992. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.A.; Ni Chonghaile, T.; Matchett, K.B.; Lynam-Lennon, N.; Kiely, P.A. Big Data-Led Cancer Research, Application, and Insights. Cancer Res. 2016, 76, 6167–6170. [Google Scholar] [CrossRef] [PubMed]
- Schloer, S.; Pajonczyk, D.; Rescher, U. Annexins in Translational Research: Hidden Treasures to Be Found. Int. J. Mol. Sci. 2018, 19, 1781. [Google Scholar] [CrossRef] [PubMed]
- Mirsaeidi, M.; Gidfar, S.; Vu, A.; Schraufnagel, D. Annexins family: Insights into their functions and potential role in pathogenesis of sarcoidosis. J. Transl. Med. 2016, 14, 89. [Google Scholar] [CrossRef]
- Yu, S.; Bian, H.; Gao, X.; Gui, L. Annexin A9 promotes invasion and metastasis of colorectal cancer and predicts poor prognosis. Int. J. Mol. Med. 2018, 41, 2185–2192. [Google Scholar] [CrossRef]
- Ma, S.; Lu, C.C.; Yang, L.Y.; Wang, J.-J.; Wang, B.-S.; Cai, H.-Q.; Hao, J.-J.; Xu, X.; Cai, Y.; Zhang, Y.; et al. ANXA2 promotes esophageal cancer progression by activating MYC-HIF1A-VEGF axis. J. Exp. Clin. Cancer Res. 2018, 37, 183. [Google Scholar] [CrossRef]
- Bizzarro, V.; Belvedere, R.; Migliaro, V.; Romano, E.; Parente, L.; Petrella, A. Hypoxia regulates ANXA1 expression to support prostate cancer cell invasion and aggressiveness. Cell Adhes. Migr. 2017, 11, 247–260. [Google Scholar] [CrossRef]
- Wang, K.; Li, J. Overexpression of ANXA3 is an independent prognostic indicator in gastric cancer and its depletion suppresses cell proliferation and tumor growth. Oncotarget 2016, 7, 86972–86984. [Google Scholar] [CrossRef]
- Bello, N.; Lopez-Kleine, L. Prog-Plot—A visual method to determine functional relationships for false discovery rate regression methods. J. Cell Sci. 2023, 136, jcs260312. [Google Scholar] [CrossRef]
- Bai, F.; Zhang, P.; Fu, Y.; Chen, H.; Zhang, M.; Huang, Q.; Li, D.; Li, B.; Wu, K. Targeting ANXA1 abrogates Treg-mediated immune suppression in triple-negative breast cancer. J. Immunother. Cancer 2020, 8, e000169. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhou, X.; Deng, X.; Ye, D.; Liu, D.; Zhou, B.; Zheng, W.; Wang, X.; Wang, Y.; Borkhuu, O.; et al. miR-186-ANXA9 signaling inhibits tumorigenesis in breast cancer. Front. Oncol. 2023, 13, 1166666. [Google Scholar] [CrossRef]
- Zhang, T.; Yu, S.; Zhao, S. ANXA9 as a novel prognostic biomarker associated with immune infiltrates in gastric cancer. PeerJ 2021, 9, e12605. [Google Scholar] [CrossRef] [PubMed]
- Perl, A.E.; Martinelli, G.; Cortes, J.E.; Neubauer, A.; Berman, E.; Paolini, S.; Montesinos, P.; Baer, M.R.; Larson, R.A.; Ustun, C.; et al. Gilteritinib or Chemotherapy for Relapsed or Refractory FLT3-Mutated AML. N. Engl. J. Med. 2019, 381, 1728–1740. [Google Scholar] [CrossRef]
- Stone, R.M.; Mandrekar, S.J.; Sanford, B.L.; Laumann, K.; Geyer, S.; Bloomfield, C.D.; Thiede, C.; Prior, T.W.; Döhner, K.; Marcucci, G.; et al. Midostaurin plus Chemotherapy for Acute Myeloid Leukemia with a FLT3 Mutation. N. Engl. J. Med. 2017, 377, 454–464. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, R.; Liu, J.; Yang, X.; Xiao, Y.; Xu, X.; Liu, X.; Zeng, W.; Zhang, S.; Wang, L. Antitumor activity of gilteritinib, an inhibitor of AXL, in human solid tumors. Cell Death Discov. 2025, 11, 124. [Google Scholar] [CrossRef] [PubMed]
- Renneville, A.; Gasser, J.A.; Grinshpun, D.E.; Jean Beltran, P.M.; Udeshi, N.D.; Matyskiela, M.E.; Clayton, T.; McConkey, M.; Viswanathan, K.; Tepper, A.; et al. Avadomide induces degradation of ZMYM2 fusion oncoproteins in hematologic malignancies. Blood Cancer Discov. 2021, 2, 250–265. [Google Scholar] [CrossRef]
- Aydin, E.; Tokat, U.M.; Adibi, A.; Ozgu, E.; Bilgic, S.N.; Demiray, M. Case report: Precision guided reactive cancer management: Molecular complete response in heavily pretreated metastatic CRC by dual immunotherapy and sorafenib. Front. Oncol. 2024, 14, 1405170. [Google Scholar]
- Munthe-Kaas, M.C.; Forthun, R.B.; Brendehaug, A.; Eek, A.K.M.; Høysæter, T.B.; Osnes, L.T.; Prescott, T.; Spetalen, S.; Hovland, R. Partial Response to Sorafenib in a Child with a Myeloid/Lymphoid Neoplasm, Eosinophilia, and a ZMYM2-FLT3 Fusion. J. Pediatr. Hematol. Oncol. 2021, 43, e508–e511. [Google Scholar] [CrossRef]
- Graham-Paquin, A.L.; Saini, D.; Sirois, J.; Hossain, I.; Katz, M.S.; Zhuang, Q.K.-W.; Kwon, S.Y.; Yamanaka, Y.; Bourque, G.; Bouchard, M.; et al. ZMYM2 is essential for methylation of germline genes and active transposons in embryonic development. Nucleic Acids Res. 2023, 51, 7314–7329. [Google Scholar] [CrossRef]
- Edge, S.B.; Compton, C.C. The American Joint Committee on Cancer: The 7th edition of the AJCC cancer staging manual and the future of TNM. Ann. Surg. Oncol. 2010, 17, 1471–1474. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Deng, P.; Chen, Y.; Lin, D.; Liu, S.; Hong, J.; Guan, P.; Chen, J.; Zhong, M.-E.; Chen, J.; et al. Pharmacological modulation of RB1 activity mitigates resistance to neoadjuvant chemotherapy in locally advanced rectal cancer. Proc. Natl. Acad. Sci. USA 2024, 121, e1990348175. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Cui, R.X.; He, Q.M.; Huang, B.J.; Sun, Y.; Xie, D.; Zeng, J.; Wang, H.Y.; Ma, J. Reduced expression of Dicer11 is associated with poor prognosis in patients with nasopharyngeal carcinoma. Med. Oncol. 2013, 30, 360. [Google Scholar] [CrossRef]
- Jie, X.; Fong, W.P.; Zhou, R.; Zhao, Y.; Zhao, Y.; Meng, R.; Zhang, S.; Dong, X.; Zhang, T.; Yang, K.; et al. USP9X-mediated KDM4C deubiquitination promotes lung cancer radioresistance by epigenetically inducing TGF-β2 transcription. Cell Death Differ. 2021, 28, 2095–2111. [Google Scholar] [CrossRef] [PubMed]
- Daver, N.; Schlenk, R.F.; Russell, N.H.; Levis, M.J. Targeting FLT3 mutations in AML: Review of current knowledge and evidence. Leukemia 2019, 33, 299–312. [Google Scholar] [CrossRef]
- Hasegawa, H.; Taniguchi, H.; Nakamura, Y.; Kato, T.; Fujii, S.; Ebi, H.; Shiozawa, M.; Yuki, S.; Masuishi, T.; Kato, K.; et al. FMS-like tyrosine kinase 3 (FLT3) amplification in patients with metastatic colorectal cancer. Cancer Sci. 2021, 112, 314–322. [Google Scholar] [CrossRef]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic Signaling Pathways in The Cancer Genome Atlas. Cell 2018, 173, 321–337.e10. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, D.; Xu, Y.; Zhan, H.; Liang, Y.; Liu, R.; Liu, J.; Luo, D.; Chen, X.; Cai, J.; Zou, Y. Targeting the ZMYM2-ANXA9 Axis with FLT3 Inhibitor G749 Overcomes Oxaliplatin Resistance in Colorectal Cancer. Biomedicines 2025, 13, 1247. https://doi.org/10.3390/biomedicines13051247
Lin D, Xu Y, Zhan H, Liang Y, Liu R, Liu J, Luo D, Chen X, Cai J, Zou Y. Targeting the ZMYM2-ANXA9 Axis with FLT3 Inhibitor G749 Overcomes Oxaliplatin Resistance in Colorectal Cancer. Biomedicines. 2025; 13(5):1247. https://doi.org/10.3390/biomedicines13051247
Chicago/Turabian StyleLin, Dezheng, Yucheng Xu, Huanmiao Zhan, Yufan Liang, Riyun Liu, Jun Liu, Dandong Luo, Xiaochuan Chen, Jiawei Cai, and Yifeng Zou. 2025. "Targeting the ZMYM2-ANXA9 Axis with FLT3 Inhibitor G749 Overcomes Oxaliplatin Resistance in Colorectal Cancer" Biomedicines 13, no. 5: 1247. https://doi.org/10.3390/biomedicines13051247
APA StyleLin, D., Xu, Y., Zhan, H., Liang, Y., Liu, R., Liu, J., Luo, D., Chen, X., Cai, J., & Zou, Y. (2025). Targeting the ZMYM2-ANXA9 Axis with FLT3 Inhibitor G749 Overcomes Oxaliplatin Resistance in Colorectal Cancer. Biomedicines, 13(5), 1247. https://doi.org/10.3390/biomedicines13051247