
Hippocampal Transcriptome Analysis in a Mouse Model of Chronic Unpredictable Stress Insomnia

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Instruments and Reagents
2.3. CUMS and CUMS+Noise Model
2.4. Pentobarbital Sodium-Induced Sleep Test (PSST)
2.5. Behavioral Tests
2.5.1. Open-Field Test (OFT)
2.5.2. Elevated Plus Maze Test (EPM)
2.5.3. Tail Suspension Test (TST)
2.5.4. Forced Swimming Test (FST)
2.6. Sleep Recordings and Scoring
2.7. RNA Sequencing
2.8. WGCNA Analysis and Key Module Bioinformatics Analysis
2.9. Nissl Staining
2.10. Serum Corticosterone Measurement
2.11. RNA Extraction and Quantitative RT-PCR Detection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| Alb | TGC TTT TTC CAG GGG TGT GTT | TTA CTT CCT GCA CTA ATT TGG CA |
| Npsr1 | TGT GCC GAT GCT AGA TTC TTC C | CAG GAC CCA CAG GGT TAT CAG |
| P2rx1 | GGA TGG TGC TGG TAC GAA ACA | CAC TGA CAC ACT GCT GAT AAG G |
2.12. Statistical Analysis
3. Results
3.1. Mice Induced by CUMS and CUMS+Noise Showed Anxiety-like Behavior

3.2. Sleep Disturbances Successfully Induced by the CUMS+Noise 8-H Method
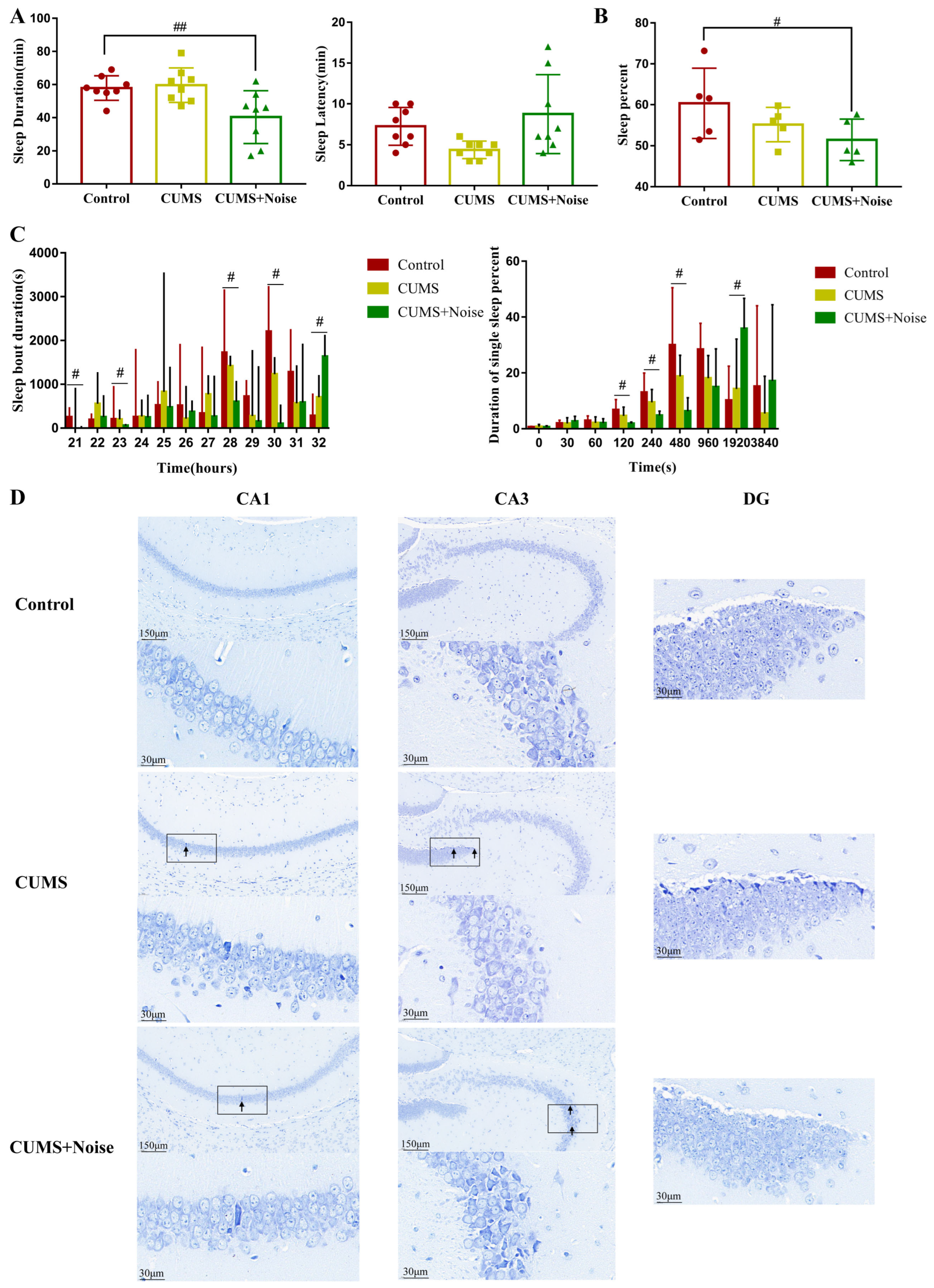
3.3. Investigation of Hub Genes and Key Pathways Regulating CUMS in Conjunction with Noise Exposure Through WGCNA Analysis and Bioinformatics Approaches
3.3.1. Key Gene Module Identification
3.3.2. Key Gene Module Expression and Gene Function
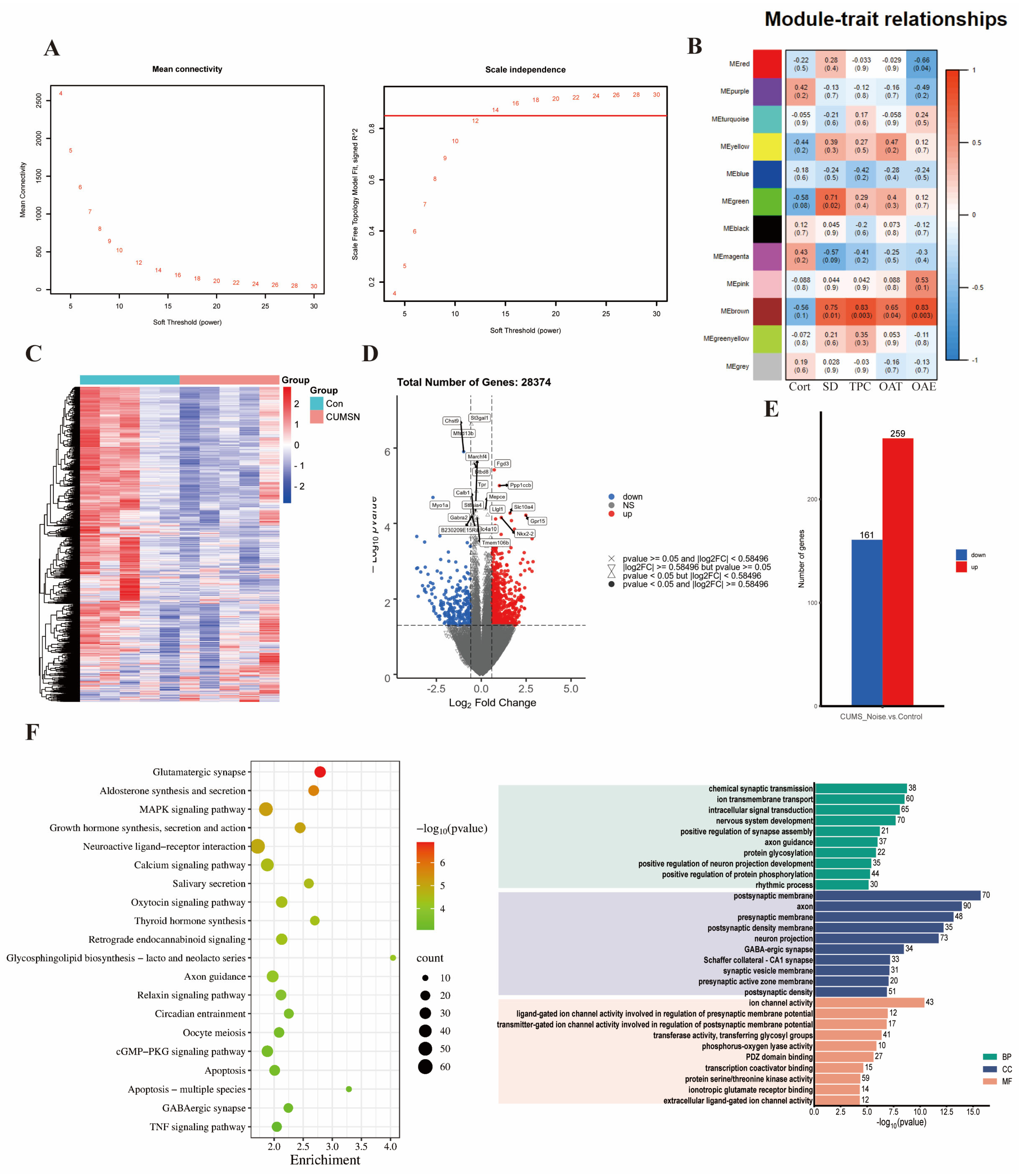
3.3.3. Selection and Verification of Key Module Hub Genes
3.4. Validation of the Altered Expression of Hippocampal Genes in Mice Subjected to CUMS in Conjunction with Noise Exposure

| Gene Symbol | log2Fold Change | p-Value | Degree | MCC | Pathway |
|---|---|---|---|---|---|
| Alb | 1.974 | 0.004 | 115 | 761,150,689 | Thyroid hormone synthesis |
| Npsr1 | −1.495 | 0.009 | 65 | 111,224,013,584 | Neuroactive ligand–receptor interaction |
| Rasl2-9 | 1.217 | 0.047 | 14 | 26,047 | Human T-cell leukemia virus 1 infection |
| Ccl22 | 1.420 | 0.016 | 12 | 11,569 | None |
| H2-Oa | 1.041 | 0.040 | 11 | 728 | Human T-cell leukemia virus 1 infection |
| Pou4f1 | 1.715 | 0.023 | 11 | 133 | None |
| Reg3g | −2.645 | 0.021 | 10 | 24 | None |
| Qprt | 1.047 | 0.029 | 9 | 12 | None |
| Actg2 | −1.655 | 0.020 | 8 | 37 | None |
| Slc45a2 | 1.266 | 0.004 | 8 | 14 | None |
| Wnt8b | 1.387 | 0.035 | 8 | 8 | None |
| Abca12 | −1.493 | 0.011 | 7 | 19 | None |
| Cnga3 | 1.317 | 0.005 | 7 | 15 | None |
| P2rx1 | −1.168 | 0.005 | 7 | 14 | Neuroactive ligand–receptor interaction; Calcium signaling pathway |
| Serpina1b | 1.434 | 0.023 | 7 | 7 | None |
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CUMS | Chronic unpredictable mild stress |
| WGCNA | Weighted Gene Co-Expression Network approach |
| SD | Sleep deprivation |
| TPC | Time spent in the center |
| TDMC% | The percentage of total distance moved in the center |
| TDMP% | The percentage of total distance moved in the periphery |
| TPC | Time spent in the center |
| TPP | Time spent in the periphery |
| OAT% | The percentage of time spent in the open arms |
| OAE% | The percentage of entries to the open arms |
| OATDM% | The percentage of total distance moved to the open arms |
References
- Yoo, J.; Slavish, D.; Dietch, J.R.; Kelly, K.; Ruggero, C.; Taylor, D.J. Daily reactivity to stress and sleep disturbances: Unique risk factors for insomnia. Sleep 2023, 46, zsac256. [Google Scholar] [CrossRef] [PubMed]
- Meaklim, H.; Junge, M.F.; Varma, P.; Finck, W.A.; Jackson, M.L. Pre-existing and post-pandemic insomnia symptoms are associated with high levels of stress, anxiety, and depression globally during the COVID-19 pandemic. J. Clin. Sleep Med. 2021, 17, 2085–2097. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.N.; Walker, M.P. The role of sleep in emotional brain function. Annu. Rev. Clin. Psychol. 2014, 10, 679–708. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Li, Y.; Chen, M.; Li, H.; Liang, C.; Chen, Y.; Long, C.; Yang, Y.; Ye, J.; Mao, Y.; et al. Insomnia and stress: The mediating roles of frontoparietal network. Brain Imaging Behav. 2024, 18, 1355–1365. [Google Scholar] [CrossRef]
- Kraus, S.S.; Rabin, L.A. Sleep America: Managing the crisis of adult chronic insomnia and associated conditions. J. Affect. Disord. 2012, 138, 192–212. [Google Scholar] [CrossRef]
- Riemann, D.; Nissen, C.; Palagini, L.; Otte, A.; Perlis, M.L.; Spiegelhalder, K. The neurobiology, investigation, and treatment of chronic insomnia. Lancet Neurol. 2015, 14, 547–558. [Google Scholar] [CrossRef]
- Kalmbach, D.A.; Anderson, J.R.; Drake, C.L. The impact of stress on sleep: Pathogenic sleep reactivity as a vulnerability to insomnia and circadian disorders. J. Sleep Res. 2018, 27, e12710. [Google Scholar] [CrossRef]
- Monti, J.M.; Monti, D. Sleep disturbance in generalized anxiety disorder and its treatment. Sleep Med. Rev. 2000, 4, 263–276. [Google Scholar] [CrossRef]
- Johnson, E.O.; Roth, T.; Breslau, N. The association of insomnia with anxiety disorders and depression: Exploration of the direction of risk. J. Psychiatr. Res. 2006, 40, 700–708. [Google Scholar] [CrossRef]
- Mason, E.C.; Harvey, A.G. Insomnia before and after treatment for anxiety and depression. J. Affect. Disord. 2014, 168, 415–421. [Google Scholar] [CrossRef]
- Zhang, L.G.; Cheng, L.F.; Wang, T.T.; Wang, L.L.; Zhou, S.J.; Luo, Y.H.; Chen, J.X. Chain mediating effect of insomnia, depression, and anxiety on the relationship between nightmares and cognitive deficits in adolescents. J. Affect. Disord. 2023, 322, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Roberge, E.M.; Bryan, C.J. An integrated model of chronic trauma-induced insomnia. Clin. Psychol. Psychother. 2021, 28, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, X.; Yan, D.; Wang, G.; Wang, Q.; Ren, X.; Liu, T. Establishment of insomnia model of chronic unpredictable stress in rats. Heliyon 2023, 9, e18338. [Google Scholar] [CrossRef]
- Xu, Y.X.; Liu, G.Y.; Ji, Z.Z.; Li, Y.Y.; Wang, Y.L.; Wu, X.Y.; Liu, J.L.; Ma, D.X.; Zhong, M.K.; Gao, C.B.; et al. Restraint stress induced anxiety and sleep in mice. Front. Psychiatry 2023, 14, 1090420. [Google Scholar] [CrossRef]
- Radwan, B.; Jansen, G.; Chaudhury, D. Abnormal sleep signals vulnerability to chronic social defeat stress. Front. Neurosci. 2021, 14, 610655. [Google Scholar] [CrossRef]
- Davis, C.J.; Schmidt, M.A.; Hemmer, K.; Krömmelbein, N.; Seilheimer, B. Multicomponent drug Neurexan mitigates acute stress-induced insomnia in rats. J. Sleep Res. 2022, 31, e13550. [Google Scholar] [CrossRef]
- Miyazaki, K.; Itoh, N.; Ohyama, S.; Kadota, K.; Oishi, K. Continuous exposure to a novel stressor based on water aversion induces abnormal circadian locomotor rhythms and sleep-wake cycles in mice. PLoS ONE 2013, 8, e55452. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, T.; Zhao, Y.; Cai, E.; Zhu, H.; Liu, S. Panaxynol attenuates CUMS-induced anxiety and depressive-like behaviors via regulating neurotransmitters, synapses and the HPA axis in mice. Food Funct. 2020, 11, 1235–1244. [Google Scholar] [CrossRef]
- Li, Z.R.; Liu, D.G.; Xie, S.; Wang, Y.H.; Han, Y.S.; Li, C.Y.; Zou, M.S.; Jiang, H.X. Sleep deprivation leads to further impairment of hippocampal synaptic plasticity by suppressing melatonin secretion in the pineal gland of chronically unpredictable stress rats. Eur. J. Pharmacol. 2022, 930, 175149. [Google Scholar] [CrossRef]
- Kang, Z.; Zheng, Z.; Guo, W. Efficacy of agomelatine on sleep disorders and lateral habenula neuronal activity in chronic restraint stress depression model mice. Psychopharmacology 2025, 242, 353–360. [Google Scholar] [CrossRef]
- Yasugaki, S.; Liu, C.Y.; Kashiwagi, M.; Kanuka, M.; Honda, T.; Miyata, S.; Yanagisawa, M.; Hayashi, Y. Effects of 3 weeks of water immersion and restraint stress on sleep in mice. Front. Neurosci. 2019, 13, 1072. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.J.; Wang, Z.; Jin, S.W.; Liu, X.M.; Liu, Y.G.; Zhang, S.S.; Pan, R.L.; Jiang, N.; Liao, Y.H.; Yan, M.Z.; et al. Melatonin-related dysfunction in chronic restraint stress triggers sleep disorders in mice. Front. Pharmacol. 2023, 14, 1210393. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.; Roberts, H.; Kwan, M.-P.; Helbich, M. Transportation noise exposure and anxiety: A systematic review and meta-analysis. Environ. Res. 2020, 191, 110118. [Google Scholar] [CrossRef]
- Dong, Y.J.; Jiang, N.H.; Zhan, L.H.; Teng, X.; Fang, X.; Lin, M.Q.; Xie, Z.Y.; Luo, R.; Li, L.Z.; Li, B.; et al. Soporific effect of modified Suanzaoren Decoction on mice models of insomnia by regulating Orexin-A and HPA axis homeostasis. Biomed. Pharmacother. 2021, 143, 112141. [Google Scholar] [CrossRef]
- Hu, H.; Li, F.; Cheng, S.; Qu, T.; Shen, F.; Cheng, J.; Chen, L.; Zhao, Z.; Hu, H. Alternate-day fasting ameliorated anxiety-like behavior in high-fat diet-induced obese mice. J. Nutr. Biochem. 2024, 124, 109526. [Google Scholar] [CrossRef]
- Xu, F.R.; Wei, Z.H.; Xu, X.X.; Zhang, X.G.; Wei, C.J.; Qi, X.-M.; Li, Y.H.; Gao, X.L.; Wu, Y. The hypothalamic steroidogenic pathway mediates susceptibility to inflammation-evoked depression in female mice. J. Neuroinflamm. 2023, 20, 293. [Google Scholar] [CrossRef]
- Badran, M.; Puech, C.; Barrow, M.B.; Runion, A.R.; Gozal, D. Solriamfetol enhances wakefulness and improves cognition and anxiety in a murine model of OSA. Sleep Med. 2023, 107, 89–99. [Google Scholar] [CrossRef]
- Mang, G.M.; Nicod, J.; Emmenegger, Y.; Donohue, K.D.; O’Hara, B.F.; Franken, P. Evaluation of a piezoelectric system as an alternative to electroencephalogram/electromyogram recordings in mouse sleep studies. Sleep 2014, 37, 1383–1392. [Google Scholar] [CrossRef]
- Paulose, J.K.; Wang, C.; O’Hara, B.F.; Cassone, V.M. The effects of aging on sleep parameters in a healthy, melatonin-competent mouse model. Nat. Sci. Sleep 2019, 11, 113–121. [Google Scholar] [CrossRef]
- Yang, S.; Yi, L.; Xia, X.; Chen, X.; Hou, X.; Zhang, L.; Yang, F.; Liao, J.; Han, Z.; Fu, Y. Transcriptome comparative analysis of amygdala-hippocampus in depression: A rat model induced by chronic unpredictable mild stress (CUMS). J. Affect. Disord. 2023, 334, 258–270. [Google Scholar] [CrossRef]
- Yan, B.; Liao, P.; Han, Z.; Zhao, J.; Gao, H.; Liu, Y.; Chen, F.; Lei, P. Association of aging related genes and immune microenvironment with major depressive disorder. J. Affect. Disord. 2025, 369, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Sun, P.; Li, Z.; Li, J.; Lv, X.; Chen, S.; Zhu, X.; Chai, X.; Zhao, S. Eucommiae cortex polysaccharides attenuate gut microbiota dysbiosis and neuroinflammation in mice exposed to chronic unpredictable mild stress: Beneficial in ameliorating depressive-like behaviors. J. Affect. Disord. 2023, 334, 278–292. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, X.; Liu, Z.Z.; Tein, J.Y.; Jia, C.X. Life stress, insomnia, and anxiety/depressive symptoms in adolescents: A three-wave longitudinal study. J. Affect. Disord. 2023, 322, 91–98. [Google Scholar] [CrossRef]
- Chellappa, S.L.; Aeschbach, D. Sleep and anxiety: From mechanisms to interventions. Sleep Med. Rev. 2022, 61, 101583. [Google Scholar] [CrossRef]
- Blake, M.J.; Trinder, J.A.; Allen, N.B. Mechanisms underlying the association between insomnia, anxiety, and depression in adolescence: Implications for behavioral sleep interventions. Clin. Psychol. Rev. 2018, 63, 25–40. [Google Scholar] [CrossRef]
- Fernandez, F.X.; Perlis, M.L. Animal models of human insomnia. J. Sleep Res. 2023, 32, e13845. [Google Scholar] [CrossRef]
- Kon, K.; Ode, K.L.; Mano, T.; Fujishima, H.; Takahashi, R.R.; Tone, D.; Shimizu, C.; Shiono, S.; Yada, S.; Matsuzawa, K.; et al. Cortical parvalbumin neurons are responsible for homeostatic sleep rebound through CaMKII activation. Nat. Commun. 2024, 15, 6054. [Google Scholar] [CrossRef]
- Schussler, G.C. Thyroxine-binding proteins. Thyroid 1990, 1, 25–34. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Brent, G.A. The Role of Thyroid Hormone in Neuronal Protection. Compr. Physiol. 2021, 11, 2075–2095. [Google Scholar] [CrossRef]
- Tesfaye, E.; Getnet, M.; Anmut Bitew, D.; Adugna, D.G.; Maru, L. Brain functional connectivity in hyperthyroid patients: Systematic review. Front. Neurosci. 2024, 18, 1383355. [Google Scholar] [CrossRef]
- Wang, M.; Lu, X.; Zheng, X.; Xu, C.; Liu, J. The relationship between sleep duration and thyroid function in the adult US population: NHANES 2007–2012. PLoS ONE 2023, 18, e0291799. [Google Scholar] [CrossRef]
- Rodrigues, N.C.; da Cruz, N.S.; de Paula Nascimento, C.; da Conceição, R.R.; da Silva, A.C.M.; Olivares, E.L.; Marassi, M.P. Sleep deprivation alters thyroid hormone economy in rats. Exp. Physiol. 2015, 100, 193–202. [Google Scholar] [CrossRef]
- Mello da Luz, M.H.; Volejnik Pino, J.M.; Monico-Neto, M.; de Amorim, P.N.; Moreira Antunes, H.K.; Porcionatto, M.A.; Lee, K.S. Sleep deprivation modulates APOE and LDL receptor-related protein 1 through thyroid hormone T4 and impairs Abeta clearance in hippocampus of rats. Biochim. Biophys. Acta-Mol. Basis Dis. 2003, 1869, 166729. [Google Scholar] [CrossRef]
- Vasudevan, N.; Morgan, M.; Pfaff, D.; Ogawa, S. Distinct behavioral phenotypes in male mice lacking the thyroid hormone receptor α1 or β isoforms. Horm. Behav. 2013, 63, 742–751. [Google Scholar] [CrossRef]
- Chae, W.R.; Baumert, J.; Nübel, J.; Brasanac, J.; Gold, S.M.; Hapke, U.; Otte, C. Associations between individual depressive symptoms and immunometabolic characteristics in major depression. Eur. Neuropsychopharmacol. 2023, 71, 25–40. [Google Scholar] [CrossRef]
- Daujat-Chavanieu, M.; Kot, M. Albumin is a secret factor involved in multidirectional interactions among the serotoninergic, immune and endocrine systems that supervises the mechanism of CYP1A and CYP3A regulation in the liver. Pharmacol. Ther. 2020, 215, 107616. [Google Scholar] [CrossRef]
- Mestre, T.A.; Zurowski, M.; Fox, S.H. 5-Hydroxytryptamine 2A receptor antagonists as potential treatment for psychiatric disorders. Expert Opin. Investig. Drugs 2013, 22, 411–421. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Tong, C.; Cao, N.; Tian, D.; Du, L.; Xu, Y.; Wang, W.; Chen, Z.; Zhai, S. Hippocampal Transcriptome Analysis in a Mouse Model of Chronic Unpredictable Stress Insomnia. Biomedicines 2025, 13, 1205. https://doi.org/10.3390/biomedicines13051205
Zhang S, Tong C, Cao N, Tian D, Du L, Xu Y, Wang W, Chen Z, Zhai S. Hippocampal Transcriptome Analysis in a Mouse Model of Chronic Unpredictable Stress Insomnia. Biomedicines. 2025; 13(5):1205. https://doi.org/10.3390/biomedicines13051205
Chicago/Turabian StyleZhang, Shuo, Changqing Tong, Na Cao, Dong Tian, Linshan Du, Ya Xu, Weiguang Wang, Zijie Chen, and Shuangqing Zhai. 2025. "Hippocampal Transcriptome Analysis in a Mouse Model of Chronic Unpredictable Stress Insomnia" Biomedicines 13, no. 5: 1205. https://doi.org/10.3390/biomedicines13051205
APA StyleZhang, S., Tong, C., Cao, N., Tian, D., Du, L., Xu, Y., Wang, W., Chen, Z., & Zhai, S. (2025). Hippocampal Transcriptome Analysis in a Mouse Model of Chronic Unpredictable Stress Insomnia. Biomedicines, 13(5), 1205. https://doi.org/10.3390/biomedicines13051205