
Molecular Ancestry Across Allelic Variants of SLC22A1, SLC22A2, SLC22A3, ABCB1, CYP2C8, CYP2C9, and CYP2C19 in Mexican-Mestizo DMT2 Patients

 , ,
, ,  ,
,  ,
,  and
and
Abstract

1. Introduction
2. Material and Methods
2.1. Study Design
2.2. Inclusion and Exclusion Criteria
2.3. Data Collection
2.4. Genotyping Procedure
2.5. Genomic Ancestry Analysis
2.6. Statistical Analysis
2.6.1. Descriptive Analysis
2.6.2. Inferential Analysis
2.6.3. Correlation Analysis
2.6.4. Clinical Biomarker and Ancestry Inference
3. Results
3.1. Cytochrome and Transporter Allelic and Genotypic Frequencies
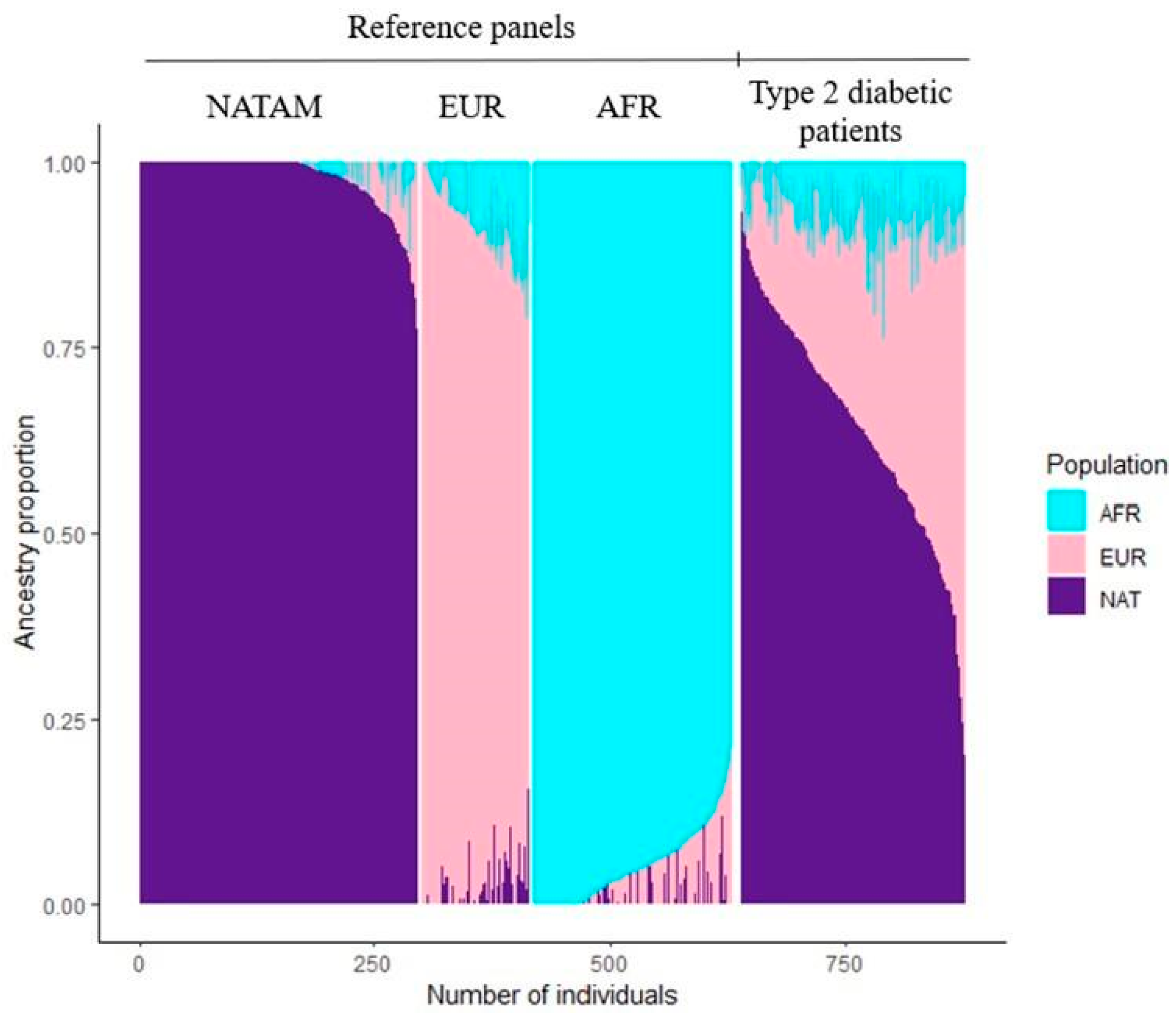
3.2. Ancestry Description
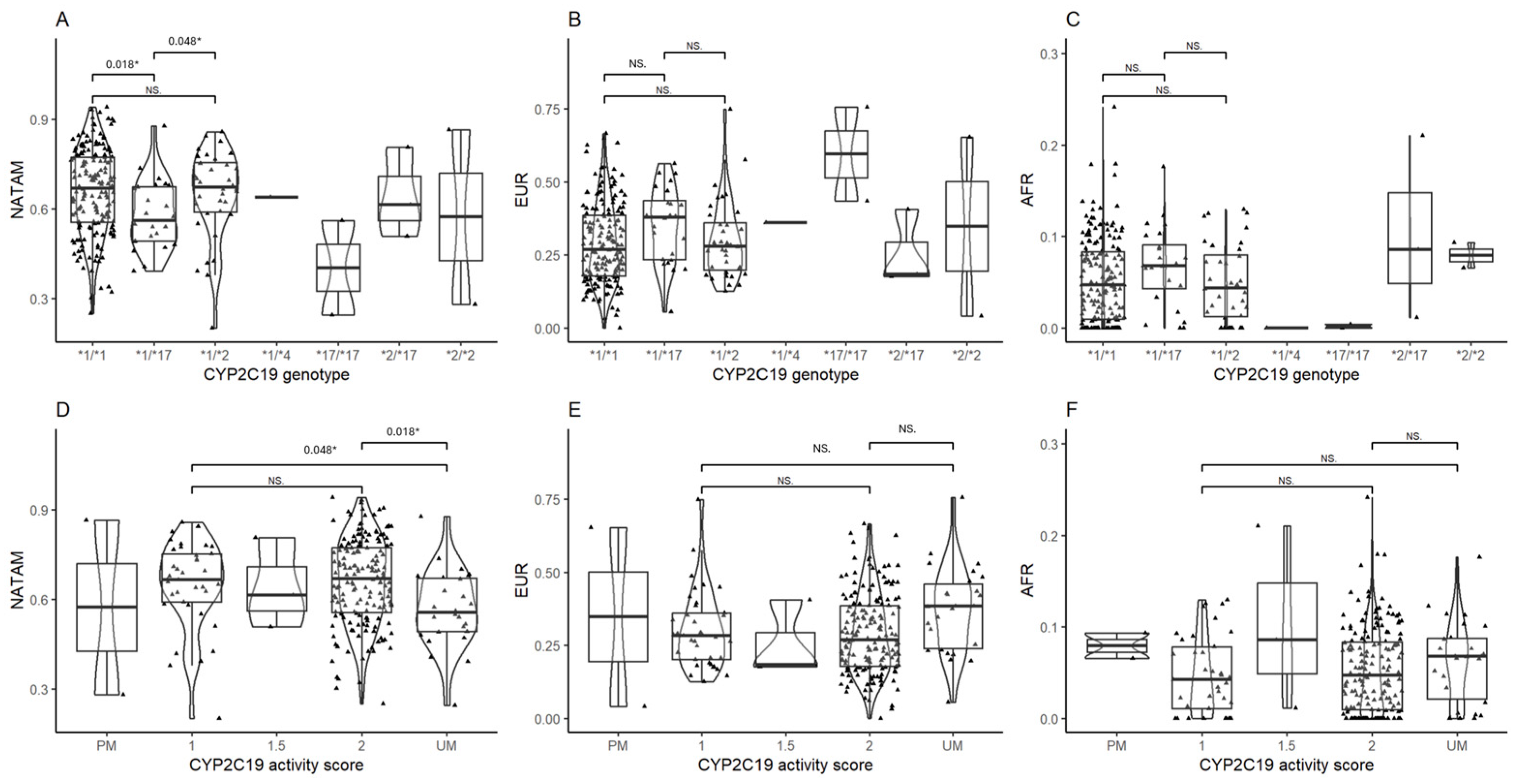
3.2.1. Ancestry Inference Across CYP2C8, CYP2C9, and CYP2C19
CYP2C8 Ancestry Inference
CYP2C9 Ancestry Inference
CYP2C19 Ancestry Inference
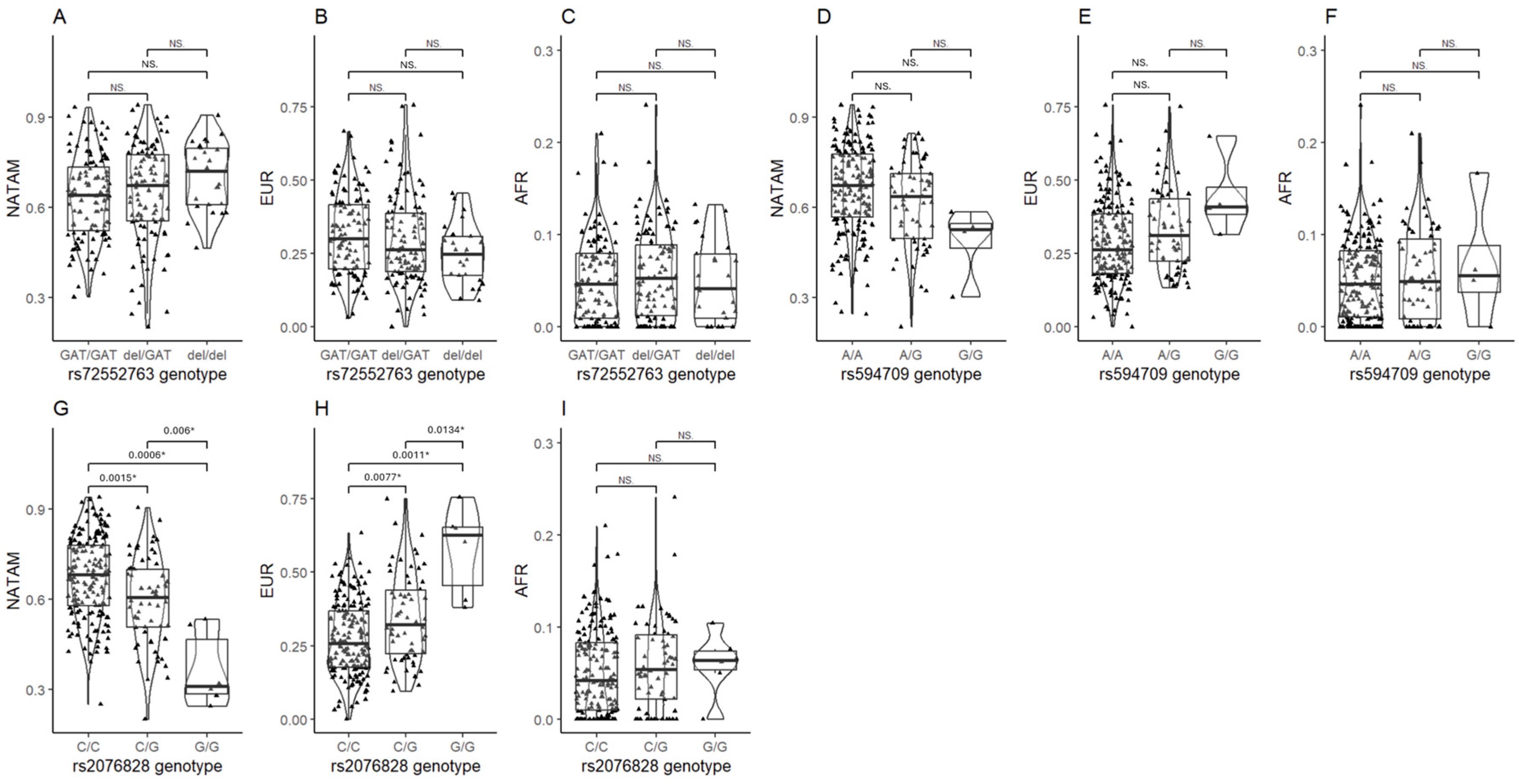
3.2.2. Ancestry Inference in Transporter SNVs
3.3. Correlation Analysis
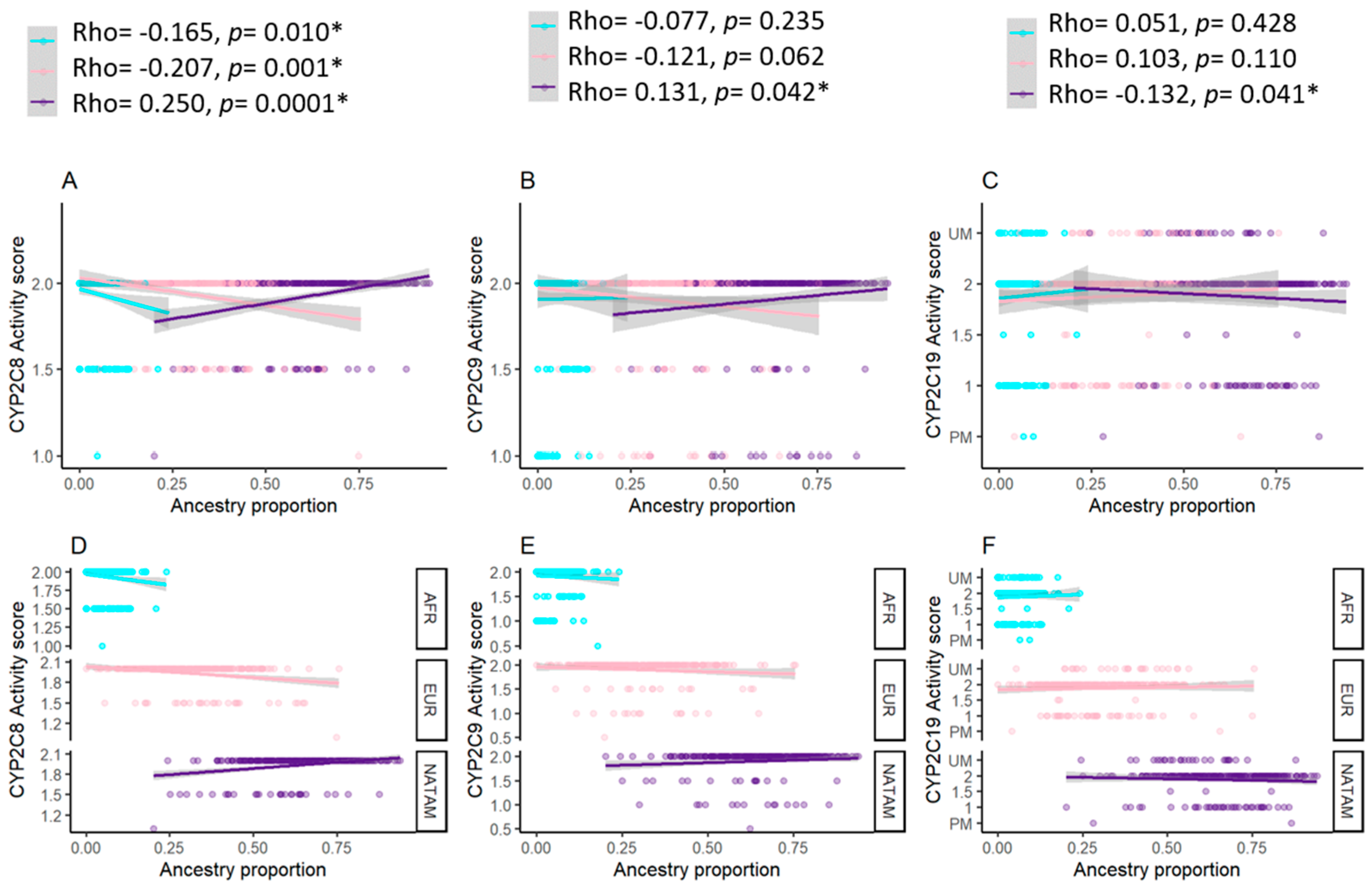
3.3.1. Correlation Analysis for CYP2C8
3.3.2. Correlation Analysis for CYP2C9
3.3.3. Correlation Analysis for CYP2C19
3.3.4. Correlation Analysis for Organic Cation Transporters (OCTs)
3.3.5. Correlation Analysis for Carried Allele Tally
3.3.6. Correlation Analysis for Activity Score and Ancestry Proportion
3.4. Clinical Biomarker and Ancestry Inference
4. Discussion
4.1. American Population Ancestry
4.2. Ancestry and Pharmacogenetics
4.2.1. CYP2C8
4.2.2. CYP2C9
4.2.3. CYP2C19
4.2.4. OCT Transporters
4.3. Ancestry, Clinical, and Pharmacogenetic Biomarkers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NATAM | Native-American |
| EUR | European |
| AFR | African |
| OCT | Organic Cation Transporter |
| SLC | Solute Carrier Family |
| CYP | P-450 Cytochrome |
| ABCB1 | ATP Binding Cassette Subfamily B Member 1 |
| SNV | Single Nucleotide Allelic Variant |
| DMT2 | Type 2 Diabetes Mellitus |
| GA | Genetic Ancestry |
References
- American Diabetes Association Professional Practice Committee. Pharmacologic Approaches to Glycemic Treatment: Standards of Care in Diabetes—2025. Diabetes Care 2025, 48 (Suppl. S1), S181–S206. [Google Scholar] [CrossRef] [PubMed]
- International Diabetes Federation. IDF Global Clinical Practice Recommendations for Managing Type 2 Diabetes—2025; International Diabetes Federation: Brussels, Belgium, 2025; Available online: https://idf.org/t2d-cpr-2025 (accessed on 6 May 2025).
- Eichelbaum, M.; Ingelman-Sundberg, M.; Evans, W.E. Pharmacogenomics and individualized drug therapy. Annu. Rev. Med. 2006, 57, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Damanhouri, Z.A.; Alkreathy, H.M.; Alharbi, F.A.; Abualhamail, H.; Ahmad, M.S. A Review of the Impact of Pharmacogenetics and Metabolomics on the Efficacy of Metformin in Type 2 Diabetes. Int. J. Med. Sci. 2023, 20, 142–150. [Google Scholar] [CrossRef]
- Wang, K.; Yang, A.; Shi, M.; Tam, C.C.H.; Lau, E.S.H.; Fan, B.; Lim, C.K.P.; Lee, H.M.; Kong, A.P.S.; Luk, A.O.Y.; et al. CYP2C19 Loss-of-function Polymorphisms are Associated with Reduced Risk of Sulfonylurea Treatment Failure in Chinese Patients with Type 2 Diabetes. Clin. Pharmacol. Ther. 2022, 111, 461–469. [Google Scholar] [CrossRef]
- Dawed, A.Y.; Donnelly, L.; Tavendale, R.; Carr, F.; Leese, G.; Palmer, C.N.; Pearson, E.R.; Zhou, K. CYP2C8 and SLCO1B1 Variants and Therapeutic Response to Thiazolidinediones in Patients With Type 2 Diabetes. Diabetes Care 2016, 39, 1902–1908. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.R.; Luzum, J.A.; Sangkuhl, K.; Gammal, R.S.; Sabatine, M.S.; Stein, C.M.; Kisor, D.F.; Limdi, N.A.; Lee, Y.M.; Scott, S.A.; et al. Clinical Pharmacogenetics Implementation Consortium Guideline for CYP2C19 Genotype and Clopidogrel Therapy: 2022 Update. Clin. Pharmacol. Ther. 2022, 112, 959–967. [Google Scholar] [CrossRef]
- Theken, K.N.; Lee, C.R.; Gong, L.; Caudle, K.E.; Formea, C.M.; Gaedigk, A.; Klein, T.E.; Agúndez, J.A.; Grosser, T. Clinical Pharmacogenetics Implementation Consortium Guideline (CPIC) for CYP2C9 and Nonsteroidal Anti-Inflammatory Drugs. Clin. Pharmacol. Ther. 2020, 108, 191–200. [Google Scholar] [CrossRef]
- Sohail, M.; Palma-Martínez, M.J.; Chong, A.Y.; Quinto-Cortés, C.D.; Barberena-Jonas, C.; Medina-Muñoz, S.G.; Ragsdale, A.; Delgado-Sánchez, G.; Cruz-Hervert, L.P.; Ferreyra-Reyes, L.; et al. Mexican Biobank advances population and medical genomics of diverse ancestries. Nature 2023, 622, 775–783. [Google Scholar] [CrossRef]
- 1000 Genomes Project Consortium; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef]
- Rodríguez-Rivera, N.S.; Cuautle-Rodríguez, P.; Castillo-Nájera, F.; Molina-Guarneros, J.A. Identification of genetic variants in pharmacogenetic genes associated with type 2 diabetes in a Mexican-Mestizo population. Biomed. Rep. 2017, 7, 21–28. [Google Scholar] [CrossRef]
- Cooper-DeHoff, R.M.; Niemi, M.; Ramsey, L.B.; Luzum, J.A.; Tarkiainen, E.K.; Straka, R.J.; Gong, L.; Tuteja, S.; Wilke, R.A.; Wadelius, M.; et al. The Clinical Pharmacogenetics Implementation Consortium Guideline for SLCO1B1, ABCG2, and CYP2C9 genotypes and Statin-Associated Musculoskeletal Symptoms. Clin. Pharmacol. Ther. 2022, 111, 1007–1021. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-Soares, F.; Peñas-Lledó, E.M.; Tarazona-Santos, E.; Sosa-Macías, M.; Terán, E.; López-López, M.; Rodeiro, I.; Moya, G.E.; Calzadilla, L.R.; Ramírez-Roa, R.; et al. Genomic Ancestry, CYP2D6, CYP2C9, and CYP2C19 Among Latin Americans. Clin. Pharmacol. Ther. 2020, 107, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Guevara, M.; Rodrigues-Soares, F.; de la Cruz, C.G.; de Andrés, F.; Rodríguez, E.; Peñas-Lledó, E.; Llerena, A.; CEIBA Consortium of the Ibero-American Network of Pharmacogenetics and Pharmacogenomics RIBEF. Afro-Latin American Pharmacogenetics of CYP2D6, CYP2C9, and CYP2C19 in Dominicans: A Study from the RIBEF-CEIBA Consortium. Pharmaceutics 2024, 16, 1399. [Google Scholar] [CrossRef]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. 2024. Available online: https://www.R-project.org/ (accessed on 6 May 2025).
- Kotecha, R.R.; Knezevic, A.; Arora, K.; Bandlamudi, C.; Kuo, F.; Carlo, M.I.; Fitzgerald, K.N.; Feldman, D.R.; Shah, N.J.; Reznik, E.; et al. Genomic ancestry in kidney cancer: Correlations with clinical and molecular features. Cancer 2024, 130, 692–701. [Google Scholar] [CrossRef]
- Nagar, S.D.; Conley, A.B.; Chande, A.T.; Rishishwar, L.; Sharma, S.; Mariño-Ramírez, L.; Aguinaga-Romero, G.; González-Andrade, F.; Jordan, I.K. Genetic ancestry and ethnic identity in Ecuador. HGG Adv. 2021, 2, 100050. [Google Scholar] [CrossRef]
- Sandoval, J.R.; Salazar-Granara, A.; Acosta, O.; Castillo-Herrera, W.; Fujita, R.; Pena, S.D.; Santos, F.R. Tracing the genomic ancestry of Peruvians reveals a major legacy of pre-Columbian ancestors. J. Hum. Genet. 2013, 58, 627–634. [Google Scholar] [CrossRef]
- Caro-Gomez, M.A.; Naranjo-González, C.A.; Gallego-Lopera, N.; Parra-Marín, M.V.; Valencia, D.M.; Arcos, E.G.; Villegas-Perrasse, A.; Bedoya-Berrío, G. Association of Native American ancestry and common variants in ACE, ADIPOR2, MTNR1B, GCK, TCF7L2 and FTO genes with glycemic traits in Colombian population. Gene 2018, 677, 198–210. [Google Scholar] [CrossRef]
- Rishishwar, L.; Conley, A.B.; Wigington, C.H.; Wang, L.; Valderrama-Aguirre, A.; Jordan, I.K. Ancestry, admixture and fitness in Colombian genomes. Sci. Rep. 2015, 5, 12376. [Google Scholar] [CrossRef]
- Azulay, R.S.; Rodrigues, V.; Lago, D.C.F.; de Almeida, A.G.F.P.; de Abreu, J.D.M.F.; Matos, L.; Andrade, C.; Nascimento, G.C.; Magalhães, M.; Facundo, A.; et al. Relationship Between C-Peptide Levels, Clinical Features, and Serum Data in a Brazilian Type 1 Diabetes Population with Large Variations in Genomic Ancestry. Int. J. Mol. Sci. 2024, 25, 11144. [Google Scholar] [CrossRef]
- Dyer, S.C.; Austine-Orimoloye, O.; Azov, A.G.; Barba, M.; Barnes, I.; Barrera-Enriquez, V.P.; Becker, A.; Bennett, R.; Beracochea, M.; Berry, A.; et al. Ensembl 2025. Nucleic Acids Res. 2025, 53, D948–D957. [Google Scholar] [CrossRef]
- Dorado, P.; Cavaco, I.; Cáceres, M.C.; Piedade, R.; Ribeiro, V.; Llerena, A. Relationship between CYP2C8 genotypes and diclofenac 5-hydroxylation in healthy Spanish volunteers. Eur. J. Clin. Pharmacol. 2008, 64, 967–970. [Google Scholar] [CrossRef] [PubMed]
- Ochoa, D.; Prieto-Pérez, R.; Román, M.; Talegón, M.; Rivas, A.; Galicia, I.; Abad-Santos, F.; Cabaleiro, T. Effect of gender and CYP2C9 and CYP2C8 polymorphisms on the pharmacokinetics of ibuprofen enantiomers. Pharmacogenomics 2015, 16, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rodriguez, R.; Novalbos, J.; Gallego-Sandin, S.; Roman-Martinez, M.; Torrado, J.; Gisbert, J.P.; Abad-Santos, F. Influence of CYP2C8 and CYP2C9 polymorphisms on pharmacokinetic and pharmacodynamic parameters of racemic and enantiomeric forms of ibuprofen in healthy volunteers. Pharmacol. Res. 2008, 58, 77–84. [Google Scholar] [CrossRef]
- Karaźniewicz-Łada, M.; Luczak, M.; Główka, F. Pharmacokinetic studies of enantiomers of ibuprofen and its chiral metabolites in humans with different variants of genes coding CYP2C8 and CYP2C9 isoenzymes. Xenobiotica 2009, 39, 476–485. [Google Scholar] [CrossRef]
- Campodónico, D.M.; Zubiaur, P.; Soria-Chacartegui, P.; Casajús, A.; Villapalos-García, G.; Navares-Gómez, M.; Gómez-Fernández, A.; Parra-Garcés, R.; Mejía-Abril, G.; Román, M.; et al. CYP2C8*3 and *4 define CYP2C8 phenotype: An approach with the substrate cinitapride. Clin. Transl. Sci. 2022, 15, 2613–2624. [Google Scholar] [CrossRef] [PubMed]
- Hirvensalo, P.; Tornio, A.; Neuvonen, M.; Tapaninen, T.; Paile-Hyvärinen, M.; Kärjä, V.; Männistö, V.T.; Pihlajamäki, J.; Backman, J.T.; Niemi, M. Comprehensive Pharmacogenomic Study Reveals an Important Role of UGT1A3 in Montelukast Pharmacokinetics. Clin. Pharmacol. Ther. 2018, 104, 158–168. [Google Scholar] [CrossRef]
- Llerena, A.; Dorado, P.; O’Kirwan, F.; Jepson, R.; Licinio, J.; Wong, M.L. Lower frequency of CYP2C9*2 in Mexican-Americans compared to Spaniards. Pharmacogenom. J. 2004, 4, 403–406. [Google Scholar] [CrossRef]
- Dorado, P.; Sosa-Macias, M.G.; Peñas-Lledó, E.M.; E Alanis-Bañuelos, R.; Wong, M.-L.; Licinio, J.; Lares-Asseff, I.; Llerena, A. CYP2C9 allele frequency differences between populations of Mexican-Mestizo, Mexican-Tepehuano, and Spaniards. Pharmacogenom. J. 2011, 11, 108–112. [Google Scholar] [CrossRef]
- Castelán-Martínez, O.D.; Hoyo-Vadillo, C.; Sandoval-García, E.; Sandoval-Ramírez, L.; González-Ibarra, M.; Solano-Solano, G.; Gómez-Díaz, R.A.; Parra, E.J.; Cruz, M.; Valladares-Salgado, A. Allele frequency distribution of CYP2C9 2 and CYP2C9 3 polymorphisms in six Mexican populations. Gene 2013, 523, 167–172. [Google Scholar] [CrossRef]
- Sosa-Macías, M.; Lazalde-Ramos, B.P.; Galaviz-Hernández, C.; Rangel-Villalobos, H.; Salazar-Flores, J.; Martínez-Sevilla, V.M.; Martínez-Fierro, M.L.; Dorado, P.; Wong, M.L.; Licinio, J.; et al. Influence of admixture components on CYP2C9*2 allele frequency in eight indigenous populations from Northwest Mexico. Pharmacogenom. J. 2013, 13, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Macías, M.; Llerena, A. Cytochrome P450 genetic polymorphisms of Mexican indigenous populations. Drug Metabol. Drug Interact. 2013, 28, 193–208. [Google Scholar] [CrossRef]
- Fricke-Galindo, I.; Jung-Cook, H.; LLerena, A.; López-López, M. Interethnic variability of pharmacogenetic biomarkers in Mexican healthy volunteers: A report from the RIBEF (Ibero-American Network of Pharmacogenetics and Pharmacogenomics). Drug Metab. Pers. Ther. 2016, 31, 61–81. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Pozos, K.; Rivera-Santiago, C.; García-Rodríguez, M.H.; Ortiz-López, M.G.; Peña-Espinoza, B.I.; Granados-Silvestre, M.d.L.Á.; Llerena, A.; Menjívar, M. Genetic variability of CYP2C9*2 and CYP2C9*3 in seven indigenous groups from Mexico. Pharmacogenomics 2016, 17, 1881–1889. [Google Scholar] [CrossRef]
- Yee, J.; Heo, Y.; Kim, H.; Yoon, H.Y.; Song, G.; Gwak, H.S. Association Between the CYP2C9 Genotype and Hypoglycemia Among Patients With Type 2 Diabetes Receiving Sulfonylurea Treatment: A Meta-analysis. Clin. Ther. 2021, 43, 836–843.e4. [Google Scholar] [CrossRef]
- Marta, M.; Sánchez-Pozos, K.; Jaimes-Santoyo, J.; Monroy-Escutia, J.; Santiago, C.R.; Granados-Silvestre, M.d.L.Á.; Ortiz-López, M.G. Pharmacogenetic Evaluation of Metformin and Sulphonylurea Response in Mexican Mestizos with Type 2 Diabetes. Curr. Drug Metab. 2020, 21, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Ayala, A.; Rodríguez-Rivera, N.S.; Andrés, F.; Llerena, A.; Pérez-Silva, E.; Espinosa-Sánchez, A.G.; Molina-Guarneros, J.A. Pharmacogenetics of Metformin Transporters Suggests No Association with Therapeutic Inefficacy among Diabetes Type 2 Mexican Patients. Pharmaceuticals 2022, 15, 774. [Google Scholar] [CrossRef]
- Castelán-Martínez, O.D.; Hoyo-Vadillo, C.; Bazán-Soto, T.B.; Cruz, M.; Tesoro-Cruz, E.; Valladares-Salgado, A. CYP2C9*3 gene variant contributes independently to glycaemic control in patients with type 2 diabetes treated with glibenclamide. J. Clin. Pharm. Ther. 2018, 43, 768–774. [Google Scholar] [CrossRef]
- Zhou, Y.; Ingelman-Sundberg, M.; Lauschke, V.M. Worldwide Distribution of Cytochrome P450 Alleles: A Meta-analysis of Population-scale Sequencing Projects. Clin. Pharmacol. Ther. 2017, 102, 688–700. [Google Scholar] [CrossRef]
- Zhang, Y.; Si, D.; Chen, X.; Lin, N.; Guo, Y.; Zhou, H.; Zhong, D. Influence of CYP2C9 and CYP2C19 genetic polymorphisms on pharmacokinetics of gliclazide MR in Chinese subjects. Br. J. Clin. Pharmacol. 2007, 64, 67–74. [Google Scholar] [CrossRef]
- Tan, B.; Zhang, Y.F.; Chen, X.Y.; Zhao, X.H.; Li, G.X.; Zhong, D.F. The effects of CYP2C9 and CYP2C19 genetic polymorphisms on the pharmacokinetics and pharmacodynamics of glipizide in Chinese subjects. Eur. J. Clin. Pharmacol. 2010, 66, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.M.; Brasch-Andersen, C.; Green, H.; Nielsen, F.; Damkier, P.; Beck-Nielsen, H.; Brosen, K. The pharmacogenetics of metformin and its impact on plasma metformin steady-state levels and glycosylated hemoglobin A1c. Pharmacogenet. Genom. 2011, 21, 837–850. [Google Scholar] [CrossRef] [PubMed]
- Sundelin, E.; Gormsen, L.; Jensen, J.; Vendelbo, M.; Jakobsen, S.; Munk, O.; Christensen, M.; Brøsen, K.; Frøkiær, J.; Jessen, N. Genetic Polymorphisms in Organic Cation Transporter 1 Attenuates Hepatic Metformin Exposure in Humans. Clin. Pharmacol. Ther. 2017, 102, 841–848. [Google Scholar] [CrossRef]
- Ortega-Ayala, A.; De Andrés, F.; Llerena, A.; Bartolo-Montiel, C.M.; Acosta-Altamirano, G.; Molina-Guarneros, J.A. Longitudinal assessment of SNPs rs72552763 and rs622342 in SLC22A1 over HbA1c control among Mexican-Mestizo diabetic type 2 patients. Front. Pharmacol. 2024, 15, 1433519. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Ayala, A.; De Andrés, F.; Llerena, A.; Bartolo-Montiel, C.M.; Molina-Guarneros, J.A. Impact of SLC22A1 variants rs622342 and rs72552763 on HbA1c and metformin plasmatic concentration levels in patients with type 2 diabetes mellitus. Biomed. Rep. 2024, 21, 117. [Google Scholar] [CrossRef]
- Reséndiz-Abarca, C.A.; Flores-Alfaro, E.; Suárez-Sánchez, F.; Cruz, M.; Valladares-Salgado, A.; Alarcón-Romero, L.d.C.; Vázquez-Moreno, M.A.; Wacher-Rodarte, N.A.; Gómez-Zamudio, J.H. Altered Glycemic Control Associated with Polymorphisms in the SLC22A1 (OCT1) Gene in a Mexican Population with Type 2 Diabetes Mellitus Treated with Metformin: A Cohort Study. J. Clin. Pharmacol. 2019, 59, 1384–1390. [Google Scholar] [CrossRef]
- Xiao, D.; Guo, Y.; Li, X.; Yin, J.-Y.; Zheng, W.; Qiu, X.-W.; Xiao, L.; Liu, R.-R.; Wang, S.-Y.; Gong, W.-J.; et al. The Impacts of SLC22A1 rs594709 and SLC47A1 rs2289669 Polymorphisms on Metformin Therapeutic Efficacy in Chinese Type 2 Diabetes Patients. Int. J. Endocrinol. 2016, 2016, 4350712. [Google Scholar] [CrossRef]
- Phani, N.M.; Vohra, M.; Kakar, A.; Adhikari, P.; Nagri, S.K.; D’souza, S.C.; Umakanth, S.; Satyamoorthy, K.; Rai, P.S. Implication of critical pharmacokinetic gene variants on therapeutic response to metformin in Type 2 diabetes. Pharmacogenomics 2018, 19, 905–911. [Google Scholar] [CrossRef]
- Frenzel, D.; Köppen, C.; Bauer, O.B.; Karst, U.; Schröter, R.; Tzvetkov, M.V.; Ciarimboli, G. Effects of Single Nucleotide Polymorphism Ala270Ser (rs316019) on the Function and Regulation of hOCT2. Biomolecules 2019, 9, 578. [Google Scholar] [CrossRef]
- Chen, E.C.; Liang, X.; Yee, S.W.; Geier, E.G.; Stocker, S.L.; Chen, L.; Giacomini, K.M. Targeted disruption of organic cation transporter 3 attenuates the pharmacologic response to metformin. Mol. Pharmacol. 2015, 88, 75–83. [Google Scholar] [CrossRef]
- Florez, J.C.; Price, A.L.; Campbell, D.; Riba, L.; Parra, M.V.; Yu, F.; Duque, C.; Saxena, R.; Gallego, N.; Tello-Ruiz, M.; et al. Strong association of socioeconomic status with genetic ancestry in Latinos: Implications for admixture studies of type 2 diabetes. Diabetologia 2009, 52, 1528–1536. [Google Scholar] [CrossRef]
- Chande, A.T.; Rishishwar, L.; Conley, A.B.; Valderrama-Aguirre, A.; Medina-Rivas, M.A.; Jordan, I.K. Ancestry effects on type 2 diabetes genetic risk inference in Hispanic/Latino populations. BMC Med. Genet. 2020, 21 (Suppl. S2), 132. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Huff, C.D.; Yamamura, Y.; Wu, X.; Strom, S.S. The Relationship between Native American Ancestry, Body Mass Index and Diabetes Risk among Mexican-Americans. PLoS ONE 2015, 10, e0141260. [Google Scholar] [CrossRef] [PubMed]
- Norden-Krichmar, T.M.; Gizer, I.R.; Libiger, O.; Wilhelmsen, K.C.; Ehlers, C.L.; Schork, N.J. Correlation analysis of genetic admixture and social identification with body mass index in a Native American community. Am. J. Hum. Biol. 2014, 26, 347–360. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NATAM | EUR | AFR | Activity Score | NATAM | EUR | AFR | |
|---|---|---|---|---|---|---|---|
| CYP2C8 | |||||||
| *1/*1 | 67.56 (55.72–77.34) | 26.42 (18.20–38.50) | 4.49 (0.90–8.18) | 1 | - | - | - |
| *1/*3 | 58.46 (47.42–63.94) | 36.11 (29.17–44.69) | 6.98 (3.67–10.10) | 1.5 | 58.07 (42.54–63.91) | 37.05 (28.69–45.36) | 6.99 (4.47–10.47) |
| *1/*4 | 51.06 (41.81–61.34) | 39.48 (29.13–54.80) | 6.99 (5.09–12.46) | 2 | 67.56 (55.72–77.34) | 26.42 (18.20–38.50) | 4.49 (0.09–8.18) |
| *3/*4 | - | - | - | ||||
| p KW | 0.001 * | 0.009 * | 0.062 | p U | <0.001 ** | 0.003 * | 0.035 * |
| CYP2C9 | |||||||
| *1/*1 | 66.52 (55.08–77.17) | 27.34 (18.20–38.59) | 4.71 (1.00–8.39) | 1 | 64.05 (51.32–72.40) | 30.07 (25.95–42.20) | 2.57 (0.85–4.80) |
| *1/*2 | 56.45 (44.83–64.32) | 34.00 (26.99–44.88) | 8.31 (6.76–10.76) | 1.5 | 56.45 (43.33–64.32) | 34.00 (26.99–44.88) | 8.31 (6.76–10.76) |
| *1/*3 | 60.54 (49.38–73.31) | 30.34 (25.71–42.29) | 2.95 (0.91–5.02) | 2 | 66.52 (55.08–77.17) | 27.34 (18.20–38.59) | 4.71 (1.00–8.39) |
| *2/*2 | - | - | - | ||||
| *2/*3 | - | - | - | ||||
| p KW | 0.061 | 0.290 | 0.019 * | p KW | 0.137 | 0.175 | 0.010 * |
| CYP2C19 | |||||||
| *1/*1 | 66.81 (55.54–77.35) | 26.85 (17.76–38.62) | 4.68 (0.98–8.35) | 0 PM | - | - | - |
| *1/*2 | 64.01 (56.37–71.01) | 32.75 (22.62–40.43) | 2.58 (0.00–7.09) | 1 | 66.52 (59.13–75.07) | 28.28 (20.13–36.11) | 4.22 (1.06–7.80) |
| *1/*4 | - | - | - | 1.5 | - | - | - |
| *1/*17 | 56.09 (49.23–67.34) | 37.87 (23.38–43.72) | 6.81 (4.27–9.11) | 2 | 66.81 (55.54–77.35) | 26.85 (17.76–38.62) | 4.68 (0.98–8.35) |
| *2/*2 | - | - | - | >2 UM | 55.53 (49.13–67.07) | 38.35 (23.88–46.00) | 6.76 (2.10–8.75) |
| *17/*17 | - | - | p KW | 0.041 * | 0.083 | 0.465 | |
| *2/*17 | - | - | - | ||||
| p KW | 0.018 * | 0.057 | 0.243 |
| Gen | ID | Genotype | NATAM | EUR | AFR |
|---|---|---|---|---|---|
| SLC22A1 | rs72552763 | GAT/GAT | 64.04 (52.23–73.34) | 30.07 (19.74–41.54) | 4.68 (0.93–7.95) |
| GAT/del | 67.16 (55.31–77.33) | 26.42 (18.83–38.77) | 5.30 (1.22–8.90) | ||
| del/del | 72.10 (60.82–79.64) | 24.78 (17.54–30.76) | 4.19 (0.97–7.94) | ||
| p KW | 0.089 | 0.090 | 0.597 | ||
| rs622342 | A/A | 64.26 (53.02–73.33) | 30.07 (19.85–40.81) | 4.71 (0.93–7.95) | |
| A/C | 68.34 (55.68–77.58) | 23.88 (17.63–38.71) | 4.50 (1.14–8.87) | ||
| C/C | 62.94 (56.71–77.94) | 29.01 (18.90–38.06) | 6.04 (1.39–8.42) | ||
| p KW | 0.430 | 0.225 | 0.474 | ||
| rs12208357 | CC | 65.50 (54.96–76.44) | 28.40 (18.49–39.76) | 4.71 (1.02–8.35) | |
| CT | 64.40 (54.20–68.03) | 27.40 (22.84–33.44) | 9.38 (4.69–10.80) | ||
| TT | - | - | - | ||
| p U | 0.485 | 0.972 | 0.097 | ||
| rs2282143 | CC | 64.90 (53.94–75.92) | 28.70 (19.56–40.13) | 4.77 (0.99–8.66) | |
| CT | 70.60 (62.67–78.55) | 21.30 (16.33–32.62) | 5.57 (1.81–7.20) | ||
| TT | - | - | - | ||
| p KW | 0.075 | 0.063 | 0.733 | ||
| rs594709 | AA | 67.20 (56.65–77.54) | 26.30 (18.09–38.47) | 4.68 (1.10–8.28) | |
| AG | 63.70 (49.48–71.06) | 31.30 (22.28–43.67) | 4.93 (0.88–9.54) | ||
| GG | 52.60 (46.43–54.63) | 41.00 (38.16–47.44) | 5.62 (3.77–8.88) | ||
| p KW | 0.008 * | 0.020 * | 0.854 | ||
| rs683369 | CC | 65.60 (55.55–76.97) | 28.40 (18.32–39.08) | 4.69 (1.02–8.35) | |
| CG | 61.50 (50.35–70.72) | 27.70 (22.60–42.31) | 6.67 (2.90–10.32) | ||
| GG | - | - | |||
| p U | 0.122 | 0.316 | 0.214 | ||
| rs628031 | GG | 67.20 (57.16–77.71) | 26.30 (18.08–38.50) | 4.59 (1.14–8.13) | |
| GA | 63.70 (49.26–71.06) | 31.30 (22.28–44.69) | 5.09 (0.88–9.54) | ||
| AA | 54.30 (51.88–58.37) | 35.30 (31.43–41.63) | 10.40 (5.02–16.68) | ||
| p KW | 0.022 * | 0.118 | 0.354 | ||
| SLC22A2 | rs316019 | C/C | 66.52 (55.70–77.02) | 27.22 (18.23–38.48) | 4.80 (1.08–8.59) |
| C/A | 56.08 (47.69–67.88) | 39.40 (25.90–48.44) | 4.83 (0.97–7.57) | ||
| A/A | - | - | - | ||
| p U | 0.009 * | 0.005 * | 0.831 | ||
| SLC22A3 | rs2076828 | C/C | 68.25 (57.82–78.04) | 25.83 (17.73–36.82) | 4.21 (0.98–8.29) |
| C/G | 60.77 (50.69–70.05) | 32.17 (22.17–43.88) | 5.44 (2.19–9.17) | ||
| G/G | 31.09 (28.56–46.70) | 62.58 (45.37–65.25) | 6.38 (5.32–7.35) | ||
| p KW | <0.001 * | <0.001 * | 0.609 | ||
| ABCB1 | rs2032582 | G/G | 64.08 (54.77–76.81) | 30.63 (17.53–38.96) | 5.00 (1.89–7.76) |
| G/A | 67.21 (60.81–74.26) | 28.68 (21.54–35.23) | 1.32 (0.13–5.74) | ||
| A/A | - | - | - | ||
| G/T | 63.68 (51.06–72.10) | 28.84 (20.91–40.59) | 5.52 (2.19–9.25) | ||
| T/T | 70.66 (55.35–79.12) | 23.22 (17.12–36.62) | 4.62 (0.00–8.04) | ||
| T/A | 73.99 (56.67–82.36) | 21.08 (14.38–38.90) | 2.06 (0.75–8.16) | ||
| p KW | 0.222 | 0.291 | 0.067 | ||
| rs1128503 | C/C | 63.38 (53.93–74.93) | 30.15 (19.10–41.09) | 3.59 (1.16–8.04) | |
| C/T | 65.40 (53.85–77.22) | 29.13 (19.70–38.87) | 5.52 (1.69–8.88) | ||
| T/T | 69.40 (55.14–77.04) | 24.78 (18.20–36.79) | 4.00 (0.39–8.12) | ||
| p KW | 0.291 | 0.380 | 0.297 | ||
| rs1045642 | C/C | 66.17 (56.33–77.83) | 26.85 (17.62–38.44) | 4.88 (1.24–8.41) | |
| C/T | 63.88 (52.05–76.02) | 29.10 (19.75–40.65) | 4.68 (1.01–8.68) | ||
| T/T | 68.60 (54.14–74.83) | 27.36 (20.15–40.85) | 5.43 (1.64–8.22) | ||
| p KW | 0.569 | 0.458 | 0.984 |
| Allelic Variant of CYP2C8 | |||
|---|---|---|---|
| Ancestry Native-American | |||
| wt | *3 | *4 | |
| Rho s | 0.250 | −0.178 | −0.194 |
| p s | <0.001 * | 0.006 * | 0.002 * |
| Ancestry EUR | |||
| Rho s | −0.207 | 0.162 | 0.149 |
| p s | 0.001 * | 0.012 * | 0.023 * |
| Ancestry AFR | |||
| Rho s | −0.165 | 0.102 | 0.146 |
| p s | 0.010 * | 0.117 | 0.023 * |
| Allele Variant of CYP2C9 | ||||
|---|---|---|---|---|
| Ancestry Native-American | ||||
| wt | *2 | *3 | Activity score | |
| Rho s | 0.135 | −0.133 | −0.050 | 0.131 |
| p s | 0.036 * | 0.039 * | 0.436 | 0.042 * |
| Ancestry EUR | ||||
| Rho s | −0.122 | 0.092 | 0.060 | −0.121 |
| p s | 0.059 | 0.154 | 0.352 | 0.062 |
| Ancestry AFR | ||||
| Rho s | −0.086 | 0.171 | −0.037 | −0.077 |
| p s | 0.182 | 0.007 * | 0.561 | 0.235 |
| Allele Variant of CYP2C19 | |||||
|---|---|---|---|---|---|
| Ancestry Native-American | |||||
| wt | *2 | *4 | *17 | Activity score | |
| Rho s | 0.123 | 0.025 | - | −0.198 | −0.132 |
| p s | 0.057 | 0.697 | - | 0.002 * | 0.041 * |
| Ancestry EUR | |||||
| Rho s | −0.096 | 0.004 | - | 0.162 | 0.103 |
| p s | 0.026 * | 0.909 | - | 0.011 * | 0.110 |
| Ancestry AFR | |||||
| Rho s | −0.047 | −0.044 | - | 0.137 | 0.051 |
| p s | 0.270 | 0.303 | - | 0.001 * | 0.428 |
| SLC22A1 | SLC22A3 | ||
|---|---|---|---|
| rs72552763 | rs594709 | rs2076828 | |
| GAT | A | C | |
| Ancestry Native-American | |||
| Rho s | −0.125 | 0.177 | 0.289 |
| p s | 0.054 | 0.005 * | <0.001 * |
| Ancestry EUR | |||
| Rho s | 0.132 | −0.162 | −0.258 |
| p s | 0.041 * | 0.012 * | <0.001 * |
| Ancestry AFR | |||
| Rho s | −0.034 | −0.035 | −0.064 |
| p s | 0.597 | 0.584 | 0.322 |
| Variable | Native-American (n = 37) | Admixture (n = 201) | p |
|---|---|---|---|
| HbA1c (%) ‖ | 7.33 (6.72–9.93) | 6.86 (6.10–8.58) | 0.037 * |
| HbA1c control (<7%) ‖ | |||
| Yes | 12 (35.29%) | 86 (52.12%) | 0.109 |
| No | 22 (64.70%) | 79 (47.87%) | |
| Glucose (mg/dL) | 137.93 (108.78–160.73) | 129 (111–173) | 0.959 |
| Height (m) | 1.46 (1.42–1.56) | 1.55 (1.50–1.62) | <0.001 * |
| Weight (kg) | 58.80 (53.40–66.30) | 70.00 (62.00–80.75) | <0.001 * |
| BMI (kg/m2) | 26.49 (25.26–30.72) | 28.88 (25.85–33.55) | 0.022 * |
| Diagnosis period (y) | 7.00 (5.00–15.00) | 6.00 (4.00–12.00) | 0.214 |
| Treatment | |||
| Metformin | 9 (28.12%) | 70 (40.69%) | 0.471 |
| Metformin + glibenclamide | 18 (56.25%) | 84 (48.83%) | |
| Glibenclamide | 3 (9.37%) | 8 (4.65%) | |
| Others | 2 (6.25%) | 10 (5.81%) | |
| Metformin dose (mg/kg/day) | 1.20 (0.77–1.67) | 1.05 (0.64–1.50) | 0.294 |
| Triglycerides | 171.00 (129.25–238.00) | 178.15 (129.25–238.00) | 0.779 |
| Total cholesterol | 195.00 (169.60–215.00) | 199.50 (169.60–224.75) | 0.558 |
| CYP2C8 genotype | 0.275 | ||
| *1/*1 | 36 (97.29%) | 171 (85.92%) | |
| *1/*3 | 1 (2.70%) | 18 (9.04%) | |
| *1/*4 | 0 (0.00%) | 9 (4.52%) | |
| *3/*4 | 0 (0.00%) | 1 (0.50%) | |
| CYP2C8 activity score | 0.153 | ||
| 1 | 0 (0.00%) | 1 (0.50%) | |
| 1.5 | 1 (2.70%) | 27 (13.56%) | |
| 2 | 36 (97.29%) | 171 (85.92%) | |
| CYP2C9 genotype | 0.710 | ||
| *1/*1 | 35 (94.59%) | 173 (86.06%) | |
| *1/*2 | 1 (2.70%) | 14 (6.96%) | |
| *1/*3 | 1 (2.70%) | 12 (5.97%) | |
| *2/*2 | 0 (0.00%) | 1 (0.49%) | |
| *2/*3 | 0 (0.00%) | 1 (0.49%) | |
| CYP2C9 activity score | 0.552 | ||
| 0.5 | 0 (0.00%) | 1 (0.49%) | |
| 1 | 1 (2.70%) | 13 (6.46%) | |
| 1.5 | 1 (2.70%) | 14 (6.96%) | |
| 2 | 35 (94.59%) | 173 (86.06%) | |
| CYP2C19 genotype | 0.252 | ||
| *1/*1 | 31 (83.78%) | 139 (69.15%) | |
| *1/*17 | 1 (2.70%) | 23 (11.44%) | |
| *1/*2 | 3 (8.10%) | 33 (16.41%) | |
| *1/*4 | 0 (0.00%) | 1 (0.49%) | |
| *17/*17 | 0 (0.00%) | 2 (0.99%) | |
| *2/*17 | 1 (2.70%) | 2 (0.99%) | |
| *2/*2 | 1 (2.70%) | 1 (0.49%) | |
| CYP2C19 activity score | 0.101 | ||
| PM | 1 (2.70%) | 1 (0.49%) | |
| 1 | 3 (8.10%) | 34 (16.91%) | |
| 1.5 | 1 (2.70%) | 2 (0.99%) | |
| 2 | 31 (83.78%) | 139 (69.15%) | |
| UM | 1 (2.70%) | 25 (12.43%) | |
| SLC22A1 (rs72552763) | 0.244 | ||
| GAT/GAT | 13 (35.13%) | 97 (48.25%) | |
| GAT/del | 18 (48.64%) | 85 (42.28%) | |
| del/del | 6 (16.21%) | 19 (9.45) | |
| SLC22A1 (rs594709) | 0.059 | ||
| AA | 33 (89.18%) | 142 (70.64%) | |
| AG | 4 (10.81%) | 55 (27.36%) | |
| GG | 0 (0.00%) | 4 (1.99%) | |
| SLC22A3 (rs2076828) | 0.010 * | ||
| CC | 34 (91.89%) | 136 (67.66%) | |
| CG | 3 (8.10%) | 59 (29.35%) | |
| GG | 0 (0.00%) | 6 (2.98%) |
| HbA1c Control | |||
|---|---|---|---|
| Yes (HbA1c < 7%) n = 98 | No (HbA1c ≥ 7%) n = 101 | p | |
| Native-American ancestry | 64.18 (51.44–73.45) | 67.16 (58.93–78.06) | 0.018 * |
| European ancestry | 29.25 (19.92–42.40) | 25.16 (17.52–35.09) | 0.022 * |
| African ancestry | 5.05 (1.54–9.15) | 4.19 (0.65–8.64) | 0.341 |
| Diagnosis period (y) | 6 (4–10) | 8 (4–15) | 0.140 |
| CYP2C8 genotype | 0.264 | ||
| *1/*1 | 85 (87.62%) | 92 (92.00%) | |
| *1/*3 | 6 (6.18%) | 7 (7.00%) | |
| *1/*4 | 5 (5.15%) | 1 (1.00%) | |
| *3/*4 | 1 (1.03%) | 0 (0.00%) | |
| CYP2C8 activity score | 0.426 | ||
| 1 | 1 (1.03%) | 0 (0.00%) | |
| 1.5 | 11 (11.34%) | 8 (8.00%) | |
| 2 | 85 (87.62%) | 92 (92.00%) | |
| CYP2C9 genotype | 0.610 | ||
| *1/*1 | 87 (88.77%) | 88 (87.12%) | |
| *1/*2 | 4 (4.08%) | 6 (5.94%) | |
| *1/*3 | 7 (7.14%) | 5 (4.95%) | |
| *2/*2 | 0 (0.00%) | 1 (0.99%) | |
| *2/*3 | 0 (0.00%) | 1 (0.99%) | |
| CYP2C9 activity score | 0.696 | ||
| 0.5 | 0 (0.00%) | 1 (0.99%) | |
| 1 | 7 (7.14%) | 6 (5.94%) | |
| 1.5 | 4 (4.08%) | 6 (5.94%) | |
| 2 | 87 (88.77%) | 88 (87.12%) | |
| CYP2C19 genotype | 0.502 | ||
| *1/*1 | 70 (71.42%) | 76 (75.24%) | |
| *1/*17 | 10 (10.20%) | 8 (7.92%) | |
| *1/*2 | 16 (16.32%) | 12 (11.88%) | |
| *1/*4 | 0 (0.00%) | 1 (0.99%) | |
| *17/*17 | 0 (0.00%) | 2 (1.98%) | |
| *2/*17 | 2 (2.04%) | 1 (0.99%) | |
| *2/*2 | 0 (0.00%) | 1 (0.99%) | |
| CYP2C19 activity score | 0.764 | ||
| PM | 0 (0.00%) | 1 (0.99%) | |
| 1 | 16 (16.32%) | 13 (12.87%) | |
| 1.5 | 2 (2.04%) | 1 (0.99%) | |
| 2 | 70 (71.42%) | 76 (75.24%) | |
| UM | 10 (10.20%) | 10 (9.90%) | |
| SLC22A1 (rs72552763) | 0.730 | ||
| GAT/GAT | 43 (43.87%) | 46 (45.54%) | |
| GAT/del | 44 (44.89%) | 47 (46.53%) | |
| del/del | 11 (11.22%) | 8 (7.92%) | |
| SLC22A1 (rs594709) | 0.819 | ||
| AA | 70 (71.42%) | 74 (73.26%) | |
| AG | 26 (26.53%) | 26 (25.74%) | |
| GG | 2 (2.04%) | 1 (0.99%) | |
| SLC22A3 (rs2076828) | 0.347 | ||
| CC | 68 (69.38%) | 79 (78.21%) | |
| CG | 28 (28.57%) | 21 (20.79%) | |
| GG | 2 (2.04%) | 1 (0.99%) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortega-Ayala, A.; de la Cruz, C.G.; Dorado, P.; Rodrigues-Soares, F.; Castillo-Nájera, F.; LLerena, A.; Molina-Guarneros, J. Molecular Ancestry Across Allelic Variants of SLC22A1, SLC22A2, SLC22A3, ABCB1, CYP2C8, CYP2C9, and CYP2C19 in Mexican-Mestizo DMT2 Patients. Biomedicines 2025, 13, 1156. https://doi.org/10.3390/biomedicines13051156
Ortega-Ayala A, de la Cruz CG, Dorado P, Rodrigues-Soares F, Castillo-Nájera F, LLerena A, Molina-Guarneros J. Molecular Ancestry Across Allelic Variants of SLC22A1, SLC22A2, SLC22A3, ABCB1, CYP2C8, CYP2C9, and CYP2C19 in Mexican-Mestizo DMT2 Patients. Biomedicines. 2025; 13(5):1156. https://doi.org/10.3390/biomedicines13051156
Chicago/Turabian StyleOrtega-Ayala, Adiel, Carla González de la Cruz, Pedro Dorado, Fernanda Rodrigues-Soares, Fernando Castillo-Nájera, Adrián LLerena, and Juan Molina-Guarneros. 2025. "Molecular Ancestry Across Allelic Variants of SLC22A1, SLC22A2, SLC22A3, ABCB1, CYP2C8, CYP2C9, and CYP2C19 in Mexican-Mestizo DMT2 Patients" Biomedicines 13, no. 5: 1156. https://doi.org/10.3390/biomedicines13051156
APA StyleOrtega-Ayala, A., de la Cruz, C. G., Dorado, P., Rodrigues-Soares, F., Castillo-Nájera, F., LLerena, A., & Molina-Guarneros, J. (2025). Molecular Ancestry Across Allelic Variants of SLC22A1, SLC22A2, SLC22A3, ABCB1, CYP2C8, CYP2C9, and CYP2C19 in Mexican-Mestizo DMT2 Patients. Biomedicines, 13(5), 1156. https://doi.org/10.3390/biomedicines13051156