
Gene Expression Changes as Biomarkers of Immunosenescence in Bulgarian Individuals of Active Age
 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Groups
2.2. Immunophenotyping
2.3. Statistical Analysis
2.4. Isolation of RNA from PBMC of Each Sample, Quality Check and Preparation of cDNA
2.5. Pathway-Focused Gene Expression Analysis
2.6. Аnalysis of the Results
3. Results
3.1. Definition of Individuals with Senescent Immunophenotype (“Test” Group)
3.2. Gene Expression Analysis
4. Discussion
4.1. Potential Interventions and Therapeutic Strategies
4.2. Limitations of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kandilarov, E. Bulgaria political briefing: Bulgaria’s Population Continues to Decline and Age. China-CEE Inst. 2024, 75, 1–9. [Google Scholar]
- Asia-Pacific Centre for Environment and Health in WPR (ACE). UN Decade of Healthy Ageing: Plan of Action (2021–2030). pp. 1–31. Available online: https://www.who.int/westernpacific/publications/m/item/un-decade-of-healthy-ageing-plan-of-action-2021-2030 (accessed on 10 March 2025).
- Liu, Z.; Liang, Q.; Ren, Y.; Guo, C.; Ge, X.; Wang, L.; Cheng, Q.; Luo, P.; Zhang, Y.; Han, X. Immunosenescence: Molecular mechanisms and diseases. Signal Transduct. Target. Ther. 2023, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Naumova, E.; Ivanova, M. Associations of Cytokine Polymorphisms with Immunosenescence. In Handbook on Immunosenescence: Basic Understanding and Clinical Applications; Fulop, T., Franceschi, C., Hirokawa, K., Pawelec, G., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 615–629. Available online: https://www.researchgate.net/publication/286769660_Associations_of_cytokine_polymorphisms_with_immunosenescence/citation/download (accessed on 10 March 2025).
- Hayflick, L.; Moorhead, P.S. The serial cultivation of human diploid cell strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef] [PubMed]
- Escrig-Larena, J.I.; Delgado-Pulido, S.; Mittelbrunn, M. Mitochondria during T cell aging. Semin. Immunol. 2023, 69, 101808. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.; Micolucci, L.; Olivieri, F.; Procopio, A.D.; Rippo, M.R. MitomiRs in Human Inflamm-Aging. In Handbook of Immunosenescence: Basic Understanding and Clinical Implications; Fulop, T., Franceschi, C., Hirokawa, K., Pawelec, G., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–29. [Google Scholar]
- Olivieri, F.; Marchegiani, F.; Matacchione, G.; Giuliani, A.; Ramini, D.; Fazioli, F.; Sabbatinelli, J.; Bonafè, M. Sex/gender-related differences in inflammaging. Mech. Ageing Dev. 2023, 211, 111792. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F.; Bairey Merz, N.; Barnes, P.J.; Brinton, R.D.; Carrero, J.J.; DeMeo, D.L.; De Vries, G.J.; Epperson, C.N.; Govindan, R.; Klein, S.L.; et al. Sex and gender: Modifiers of health, disease, and medicine. Lancet 2020, 396, 565–582. [Google Scholar] [CrossRef]
- Yalcinkaya, A.; Yalcinkaya, R.; Sardh, F.; Landegren, N. Immune dynamics throughout life in relation to sex hormones and perspectives gained from gender-affirming hormone therapy. Front. Immunol. 2024, 15, 1501364. [Google Scholar] [CrossRef]
- Di Palo, A.; Siniscalchi, C.; Salerno, M. What microRNAs could tell us about the human X chromosome. Cell. Mol. Life Sci. 2020, 77, 4069–4080. [Google Scholar] [CrossRef]
- Serbezov, D.; Balabanski, L.; Hadjidekova, S.; Toncheva, D. Genomics of longevity: Recent insights from research on centenarians. Biotechnol. Biotechnol. Equip. 2018, 32, 1359–1366. [Google Scholar] [CrossRef]
- Naumova, E.; Mihaylova, A.; Ivanova, M.; Michailova, S.; Penkova, K.; Baltadjieva, D. Immunological markers contributing to successful aging in Bulgarians. Exp. Gerontol. 2004, 39, 637–644. [Google Scholar] [CrossRef]
- Babagana, M.; Oh, K.S.; Chakraborty, S.; Pacholewska, A.; Aqdas, M.; Sung, M.H. Hedgehog dysregulation contributes to tissue-specific inflammaging of resident macrophages. Aging 2021, 13, 19207–19229. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Comans-Bitter, W.M.; de Groot, R.; van den Beemd, R.; Neijens, H.J.; Hop, W.C.; Groeneveld, K.; Hooijkaas, H.; van Dongen, J.J. Immunophenotyping of blood lymphocytes in childhood. Reference values for lymphocyte subpopulations. J. Pediatr. 1997, 130, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Flores, R.R.; Jang, I.H.; Saathoff, A.; Robbins, P.D. Immune Senescence, Immunosenescence and Aging. Front. Aging 2022, 3, 900028. [Google Scholar] [CrossRef] [PubMed]
- Hima, L.; Patel, M.N.; Kannan, T.; Gour, S.; Pratap, U.P.; Priyanka, H.P.; Vasantharekha, R.; ThyagaRajan, S. Age-associated decline in neural, endocrine, and immune responses in men and women: Involvement of intracellular signaling pathways. J. Neuroimmunol. 2020, 345, 577290. [Google Scholar] [CrossRef]
- Wrona, M.V.; Ghosh, R.; Coll, K.; Chun, C.; Yousefzadeh, M.J. The 3 I’s of immunity and aging: Immunosenescence, inflammaging, and immune resilience. Front. Aging 2024, 5, 1490302. [Google Scholar] [CrossRef]
- Chaudhary, J.K.; Danga, A.K.; Kumari, A.; Bhardwaj, A.; Rath, P.C. Role of chemokines in aging and age-related diseases. Mech. Ageing Dev. 2025, 223, 112009. [Google Scholar] [CrossRef]
- Mukaida, N.; Sasaki, S.I.; Baba, T. CCL4 Signaling in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1231, 23–32. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Li, Y.; Lai, G. Study on relationship between chemokines and immune activation phenotype of breast cancer. J. Clin. Oncol. 2021, 39, e12564. [Google Scholar] [CrossRef]
- Chen, Y.; Liang, L.; Cao, S.; Hou, G.; Zhang, Q.; Ma, H.; Shi, B. Serum CCL21 as a Potential Biomarker for Cognitive Impairment in Spinal Cord Injury. BioMed Res. Int. 2020, 2020, 6692802. [Google Scholar] [CrossRef]
- Faura, J.; Bustamante, A.; Penalba, A.; Giralt, D.; Simats, A.; Martinez-Saez, E.; Alcolea, D.; Fortea, J.; Lleo, A.; Teunissen, C.E.; et al. CCL23: A Chemokine Associated with Progression from Mild Cognitive Impairment to Alzheimer’s Disease. J. Alzheimer’s Dis. JAD 2020, 73, 1585–1595. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Velasco-Velazquez, M.A.; Wang, M.; Li, Z.; Rui, H.; Peck, A.R.; Korkola, J.E.; Chen, X.; Xu, S.; DuHadaway, J.B.; et al. CCR5 Governs DNA Damage Repair and Breast Cancer Stem Cell Expansion. Cancer Res. 2018, 78, 1657–1671. [Google Scholar] [CrossRef] [PubMed]
- Sato, F.; Omura, S.; Martinez, N.E.; Tsunoda, I. Chapter 3—Animal Models of Multiple Sclerosis. In Neuroinflammation, 2nd ed.; Minagar, A., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 37–72. [Google Scholar] [CrossRef]
- Dangi, A.; Husain, I.; Jordan, C.Z.; Yu, S.; Natesh, N.; Shen, X.; Kwun, J.; Luo, X. Blocking CCL8-CCR8-Mediated Early Allograft Inflammation Improves Kidney Transplant Function. J. Am. Soc. Nephrol. JASN 2022, 33, 1876–1890. [Google Scholar] [CrossRef] [PubMed]
- Li, B.H.; Garstka, M.A.; Li, Z.F. Chemokines and their receptors promoting the recruitment of myeloid-derived suppressor cells into the tumor. Mol. Immunol. 2020, 117, 201–215. [Google Scholar] [CrossRef]
- Pelisch, N.; Rosas Almanza, J.; Stehlik, K.E.; Aperi, B.V.; Kroner, A. CCL3 contributes to secondary damage after spinal cord injury. J. Neuroinflammation 2020, 17, 362. [Google Scholar] [CrossRef]
- Yu, D.; Zhang, Z.; Ma, C.; Huang, S.; Xu, L.; Liang, J.; Li, H.; Fan, Q.; Liu, G.; Zhai, Z. CCL3 in the bone marrow microenvironment causes bone loss and bone marrow adiposity in aged mice. JCI Insight 2023, 18, e159107. [Google Scholar] [CrossRef]
- Fisher, J.S.; Adan-Barrientos, I.; Kumar, N.R.; Lancaster, J.N. The aged microenvironment impairs BCL6 and CD40L induction in CD4(+) T follicular helper cell differentiation. Aging Cell 2024, 23, e14140. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, W.; Hua, Y. Identification and validation of BCL6 and VEGFA as biomarkers and ageing patterns correlating with immune infiltrates in OA progression. Sci. Rep. 2023, 13, 2558. [Google Scholar] [CrossRef]
- Mitchell, C.A.; Verovskaya, E.V.; Calero-Nieto, F.J.; Olson, O.C.; Swann, J.W.; Wang, X.; Herault, A.; Dellorusso, P.V.; Zhang, S.Y.; Svendsen, A.F.; et al. Stromal niche inflammation mediated by IL-1 signalling is a targetable driver of haematopoietic ageing. Nat. Cell Biol. 2023, 25, 30–41. [Google Scholar] [CrossRef]
- Böni-Schnetzler, M.; Méreau, H.; Rachid, L.; Wiedemann, S.J.; Schulze, F.; Trimigliozzi, K.; Meier, D.T.; Donath, M.Y. IL-1beta promotes the age-associated decline of beta cell function. iScience 2021, 24, 103250. [Google Scholar] [CrossRef]
- Tylutka, A.; Walas, L.; Zembron-Lacny, A. Level of IL-6, TNF, and IL-1beta and age-related diseases: A systematic review and meta-analysis. Front. Immunol. 2024, 15, 1330386. [Google Scholar] [CrossRef] [PubMed]
- Prado, G.H.J.D.; Sardeli, A.V.; Lord, J.M.; Cavaglieri, C.R. The effects of ageing, BMI and physical activity on blood IL-15 levels: A systematic review and meta-analyses. Exp. Gerontol. 2022, 168, 111933. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.; Crane, E.D.; Kuo, Y.; Kim, A.; Crane, J.D. The exercise cytokine interleukin-15 rescues slow wound healing in aged mice. J. Biol. Chem. 2019, 294, 20024–20038. [Google Scholar] [CrossRef]
- Crooke, S.N.; Ovsyannikova, I.G.; Poland, G.A.; Kennedy, R.B. Immunosenescence: A systems-level overview of immune cell biology and strategies for improving vaccine responses. Exp. Gerontol. 2019, 124, 110632. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Ji, S.; Liu, L.; Liu, S.; Wang, B.; Ma, Y.; Cao, X. Promotion of TLR7-MyD88-dependent inflammation and autoimmunity in mice through stem-loop changes in Lnc-Atg16l1. Nat. Commun. 2024, 15, 10224. [Google Scholar] [CrossRef]
- Chen, D.Y.; Lin, C.C.; Chen, Y.M.; Lan, J.L.; Hung, W.T.; Chen, H.H.; Lai, K.L.; Hsieh, C.W. Involvement of TLR7 MyD88-dependent signaling pathway in the pathogenesis of adult-onset Still’s disease. Arthritis Res. Ther. 2013, 15, R39. [Google Scholar] [CrossRef]
- Lopez-Lee, C.; Kodama, L.; Fan, L.; Zhu, D.; Zhu, J.; Wong, M.Y.; Ye, P.; Norman, K.; Foxe, N.R.; Ijaz, L.; et al. Tlr7 drives sex differences in age- and Alzheimer’s disease-related demyelination. Science 2024, 386, eadk7844. [Google Scholar] [CrossRef]
- Magerus, A.; Bercher-Brayer, C.; Rieux-Laucat, F. The genetic landscape of the FAS pathway deficiencies. Biomed. J. 2021, 44, 388–399. [Google Scholar] [CrossRef]
- Kim, H.-J.; Kim, H.; Lee, J.-H.; Hwangbo, C. Toll-like receptor 4 (TLR4): New insight immune and aging. Immun. Ageing 2023, 20, 67. [Google Scholar] [CrossRef]
- Li, X.; Goobie, G.C.; Zhang, Y. Toll-interacting protein impacts on inflammation, autophagy, and vacuole trafficking in human disease. J. Mol. Med. 2021, 99, 21–31. [Google Scholar] [CrossRef]
- Tran Van Hoi, E.; De Glas, N.A.; Portielje, J.E.A.; Van Heemst, D.; Van Den Bos, F.; Jochems, S.P.; Mooijaart, S.P. Biomarkers of the ageing immune system and their association with frailty—A systematic review. Exp. Gerontol. 2023, 176, 112163. [Google Scholar] [CrossRef] [PubMed]
- Martín-Vázquez, E.; Cobo-Vuilleumier, N.; López-Noriega, L.; Lorenzo, P.I.; Gauthier, B.R. The PTGS2/COX2-PGE(2) signaling cascade in inflammation: Pro or anti? A case study with type 1 diabetes mellitus. Int. J. Biol. Sci. 2023, 19, 4157–4165. [Google Scholar] [CrossRef] [PubMed]
- Ricciotti, E.; Haines, P.G.; Beerens, M.; Kartoun, U.; Castro, C.; Tang, S.Y.; Ghosh, S.; Das, U.S.; Lahens, N.F.; Wang, T.; et al. Suppression of cyclooxygenase-2 predisposes to heart failure with preserved ejection fraction. bioRxiv 2024. [Google Scholar] [CrossRef]
- Xu, Z.; Chen, C. The Downregulation of PTGS2 Mediated by ncRNAs is Tightly Correlated with Systemic Sclerosis-Interstitial Lung Disease. Front. Genet. 2021, 12, 795034. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Chen, S.; Zhou, F.; Xie, T.; Cao, H. Construction and evaluation of Alzheimer’s disease diagnostic prediction model based on genes involved in mitophagy. Front. Aging Neurosci. 2023, 15, 1146660. [Google Scholar] [CrossRef]
- Kim, H.-J.; Kim, K.-W.; Yu, B.-P.; Chung, H.-Y. The effect of age on cyclooxygenase-2 gene expression: NF-κB activation and IκBα degradation. Free Radic. Biol. Med. 2000, 28, 683–692. [Google Scholar] [CrossRef]
- Audia, S.; Brescia, C.; Dattilo, V.; Torchia, N.; Trapasso, F.; Amato, R. The IL-23R and Its Genetic Variants: A Hitherto Unforeseen Bridge Between the Immune System and Cancer Development. Cancers 2025, 17, 55. [Google Scholar] [CrossRef]
- Nagase, H.; Ueki, S.; Fujieda, S. The roles of IL-5 and anti-IL-5 treatment in eosinophilic diseases: Asthma, eosinophilic granulomatosis with polyangiitis, and eosinophilic chronic rhinosinusitis. Allergol. Int. 2020, 69, 178–186. [Google Scholar] [CrossRef]
- Liongue, C.; Almohaisen, F.L.J.; Ward, A.C. B Cell Lymphoma 6 (BCL6): A Conserved Regulator of Immunity and Beyond. Int. J. Mol. Sci. 2024, 25, 10968. [Google Scholar] [CrossRef]
- Sun, N.; Zhang, L.; Zhang, C.; Yuan, Y. miR-144-3p inhibits cell proliferation of colorectal cancer cells by targeting BCL6 via inhibition of Wnt/β-catenin signaling. Cell. Mol. Biol. Lett. 2020, 25, 19. [Google Scholar] [CrossRef]
- Cheng, N.-L.; Weng, N.-P. MicroRNA-125b Modulates Inflammatory Chemokine CCL4 Expression and Its Reduction May Cause CCL4 Increase in Circulation with Age. In Handbook of Immunosenescence; Springer International Publishing: Cham, Switzerland, 2019; pp. 1225–1239. [Google Scholar] [CrossRef]
- Chang, T.T.; Lin, L.Y.; Chen, C.; Chen, J.W. CCL4 contributes to aging related angiogenic insufficiency through activating oxidative stress and endothelial inflammation. Angiogenesis 2024, 27, 475–499. [Google Scholar] [CrossRef] [PubMed]
- Mannick, J.B.; Teo, G.; Bernardo, P.; Quinn, D.; Russell, K.; Klickstein, L.; Marshall, W.; Shergill, S. Targeting the biology of ageing with mTOR inhibitors to improve immune function in older adults: Phase 2b and phase 3 randomised trials. Lancet. Healthy Longev. 2021, 2, e250–e262. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Lu, C.; Meng, S.; Dun, L.; Yin, N.; An, H.; Xu, H.; Liu, G.; Cai, Y. Silencing of PTGS2 exerts promoting effects on angiogenesis endothelial progenitor cells in mice with ischemic stroke via repression of the NF-κB signaling pathway. J. Cell. Physiol. 2019, 234, 23448–23460. [Google Scholar] [CrossRef] [PubMed]
- Wight, A.E.; Sido, J.M.; Degryse, S.; Ao, L.; Nakagawa, H.; Qiu Vivian, Y.; Shen, X.; Oseghali, O.; Kim, H.J.; Cantor, H. Antibody-mediated blockade of the IL23 receptor destabilizes intratumoral regulatory T cells and enhances immunotherapy. Proc. Natl. Acad. Sci. USA 2022, 119, e2200757119. [Google Scholar] [CrossRef]
- Wertheimer, T.; Zwicky, P.; Rindlisbacher, L.; Sparano, C.; Vermeer, M.; de Melo, B.M.S.; Haftmann, C.; Rückert, T.; Sethi, A.; Schärli, S.; et al. IL-23 stabilizes an effector Treg cell program in the tumor microenvironment. Nat. Immunol. 2024, 25, 512–524. [Google Scholar] [CrossRef]
- Principe, S.; Benfante, A.; Calzetta, L.; Rogliani, P.; Scichilone, N. Age does not affect the efficacy of anti-IL-5/IL-5R in severe asthmatics. World Allergy Organ. J. 2019, 12, 100081. [Google Scholar] [CrossRef]
- Wong, F.; Zheng, E.J.; Valeri, J.A.; Donghia, N.M.; Anahtar, M.N.; Omori, S.; Li, A.; Cubillos-Ruiz, A.; Krishnan, A.; Jin, W.; et al. Discovery of a structural class of antibiotics with explainable deep learning. Nature 2024, 626, 177–185. [Google Scholar] [CrossRef]
- Johnson, S.; Gerding, D.N. Bezlotoxumab. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2019, 68, 699–704. [Google Scholar] [CrossRef]
- Cisneros, B.; Garcia-Aguirre, I.; Unzueta, J.; Arrieta-Cruz, I.; Gonzalez-Morales, O.; Dominguez-Larrieta, J.M.; Tamez-Gonzalez, A.; Leyva-Gomez, G.; Magana, J.J. Immune system modulation in aging: Molecular mechanisms and therapeutic targets. Front. Immunol. 2022, 13, 1059173. [Google Scholar] [CrossRef]
- Calder, P.C.; Ortega, E.F.; Meydani, S.N.; Adkins, Y.; Stephensen, C.B.; Thompson, B.; Zwickey, H. Nutrition, Immunosenescence, and Infectious Disease: An Overview of the Scientific Evidence on Micronutrients and on Modulation of the Gut Microbiota. Adv. Nutr. 2022, 13, S1–S26. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Ref Seq Number | Symbol | Description |
|---|---|---|---|
| A01 | NM_001706 | BCL6 | B-cell CLL/lymphoma 6 |
| A02 | NM_000064 | C3 | Complement component 3 |
| A03 | NM_004054 | C3AR1 | Complement component 3a receptor 1 |
| A04 | NM_002986 | CCL11 | Chemokine (C-C motif) ligand 11 |
| A05 | NM_005408 | CCL13 | Chemokine (C-C motif) ligand 13 |
| A06 | NM_004590 | CCL16 | Chemokine (C-C motif) ligand 16 |
| A07 | NM_002987 | CCL17 | Chemokine (C-C motif) ligand 17 |
| A08 | NM_006274 | CCL19 | Chemokine (C-C motif) ligand 19 |
| A09 | NM_002982 | CCL2 | Chemokine (C-C motif) ligand 2 |
| A10 | NM_002989 | CCL21 | Chemokine (C-C motif) ligand 21 |
| A11 | NM_002990 | CCL22 | Chemokine (C-C motif) ligand 22 |
| A12 | NM_005064 | CCL23 | Chemokine (C-C motif) ligand 23 |
| B01 | NM_002991 | CCL24 | Chemokine (C-C motif) ligand 24 |
| B02 | NM_002983 | CCL3 | Chemokine (C-C motif) ligand 3 |
| B03 | NM_002984 | CCL4 | Chemokine (C-C motif) ligand 4 |
| B04 | NM_002985 | CCL5 | Chemokine (C-C motif) ligand 5 |
| B05 | NM_006273 | CCL7 | Chemokine (C-C motif) ligand 7 |
| B06 | NM_005623 | CCL8 | Chemokine (C-C motif) ligand 8 |
| B07 | NM_001295 | CCR1 | Chemokine (C-C motif) receptor 1 |
| B08 | NM_001123396 | CCR2 | Chemokine (C-C motif) receptor 2 |
| B09 | NM_001837 | CCR3 | Chemokine (C-C motif) receptor 3 |
| B10 | NM_005508 | CCR4 | Chemokine (C-C motif) receptor 4 |
| B11 | NM_001838 | CCR7 | Chemokine (C-C motif) receptor 7 |
| B12 | NM_000591 | CD14 | CD14 molecule |
| C01 | NM_001250 | CD40 | CD40 molecule, TNF receptor superfamily member 5 |
| C02 | NM_000074 | CD40LG | CD40 ligand |
| C03 | NM_005194 | CEBPB | CCAAT/enhancer binding protein (C/EBP), beta |
| C04 | NM_000567 | CRP | C-reactive protein, pentraxin-related |
| C05 | NM_000757 | CSF1 | Colony-stimulating factor 1 (macrophage) |
| C06 | NM_001511 | CXCL1 | Chemokine (C-X-C motif) ligand 1 (melanoma growth stimulating activity, alpha) |
| C07 | NM_001565 | CXCL10 | Chemokine (C-X-C motif) ligand 10 |
| C08 | NM_002089 | CXCL2 | Chemokine (C-X-C motif) ligand 2 |
| C09 | NM_002090 | CXCL3 | Chemokine (C-X-C motif) ligand 3 |
| C10 | NM_002994 | CXCL5 | Chemokine (C-X-C motif) ligand 5 |
| C11 | NM_002993 | CXCL6 | Chemokine (C-X-C motif) ligand 6 (granulocyte chemotactic protein 2) |
| C12 | NM_002416 | CXCL9 | Chemokine (C-X-C motif) ligand 9 |
| D01 | NM_000634 | CXCR1 | Chemokine (C-X-C motif) receptor 1 |
| D02 | NM_001557 | CXCR2 | Chemokine (C-X-C motif) receptor 2 |
| D03 | NM_003467 | CXCR4 | Chemokine (C-X-C motif) receptor 4 |
| D04 | NM_000639 | FASLG | Fas ligand (TNF superfamily, member 6) |
| D05 | NM_005252 | FOS | FBJ murine osteosarcoma viral oncogene homolog |
| D06 | NM_000619 | IFNG | Interferon, gamma |
| D07 | NM_000572 | IL10 | Interleukin 10 |
| D08 | NM_000628 | IL10RB | Interleukin 10 receptor, beta |
| D09 | NM_000585 | IL15 | Interleukin 15 |
| D10 | NM_002190 | IL17A | Interleukin 17A |
| D11 | NM_001562 | IL18 | Interleukin 18 (interferon-gamma-inducing factor) |
| D12 | NM_000575 | IL1A | Interleukin 1, alpha |
| E01 | NM_000576 | IL1B | Interleukin 1, beta |
| E02 | NM_000877 | IL1R1 | Interleukin 1 receptor, type I |
| E03 | NM_002182 | IL1RAP | Interleukin 1 receptor accessory protein |
| E04 | NM_000577 | IL1RN | Interleukin 1 receptor antagonist |
| E05 | NM_020525 | IL22 | Interleukin 22 |
| E06 | NM_016584 | IL23A | Interleukin 23, alpha subunit p19 |
| E07 | NM_144701 | IL23R | Interleukin 23 receptor |
| E08 | NM_000879 | IL5 | Interleukin 5 (colony-stimulating factor, eosinophil) |
| E09 | NM_000600 | IL6 | Interleukin 6 (interferon, beta 2) |
| E10 | NM_000565 | IL6R | Interleukin 6 receptor |
| E11 | NM_000584 | CXCL8 | Interleukin 8 |
| E12 | NM_000590 | IL9 | Interleukin 9 |
| F01 | NM_000211 | ITGB2 | Integrin, beta 2 (complement component 3 receptor 3 and 4 subunit) |
| F02 | NM_000893 | KNG1 | Kininogen 1 |
| F03 | NM_000595 | LTA | Lymphotoxin alpha (TNF superfamily, member 1) |
| F04 | NM_002341 | LTB | Lymphotoxin beta (TNF superfamily, member 3) |
| F05 | NM_015364 | LY96 | Lymphocyte antigen 96 |
| F06 | NM_002468 | MYD88 | Myeloid differentiation primary response gene (88) |
| F07 | NM_003998 | NFKB1 | Nuclear factor of kappa light polypeptide gene enhancer in B-cells 1 |
| F08 | NM_000625 | NOS2 | Nitric oxide synthase 2, inducible |
| F09 | NM_000176 | NR3C1 | Nuclear receptor subfamily 3, group C, member 1 (glucocorticoid receptor) |
| F10 | NM_000963 | PTGS2 | Prostaglandin-endoperoxide synthase 2 (prostaglandin G/H synthase and cyclooxygenase) |
| F11 | NM_003821 | RIPK2 | Receptor-interacting serine-threonine kinase 2 |
| F12 | NM_000450 | SELE | Selectin E |
| G01 | NM_001039661 | TIRAP | Toll-interleukin 1 receptor (TIR) domain containing adaptor protein |
| G02 | NM_003263 | TLR1 | Toll-like receptor 1 |
| G03 | NM_003264 | TLR2 | Toll-like receptor 2 |
| G04 | NM_003265 | TLR3 | Toll-like receptor 3 |
| G05 | NM_138554 | TLR4 | Toll-like receptor 4 |
| G06 | NM_003268 | TLR5 | Toll-like receptor 5 |
| G07 | NM_006068 | TLR6 | Toll-like receptor 6 |
| G08 | NM_016562 | TLR7 | Toll-like receptor 7 |
| G09 | NM_017442 | TLR9 | Toll-like receptor 9 |
| G10 | NM_000594 | TNF | Tumor necrosis factor |
| G11 | NM_003807 | TNFSF14 | Tumor necrosis factor (ligand) superfamily, member 14 |
| G12 | NM_019009 | TOLLIP | Toll interacting protein |
| H01 | NM_001101 | ACTB | Actin, beta |
| H02 | NM_004048 | B2M | Beta-2-microglobulin |
| H03 | NM_002046 | GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| H04 | NM_000194 | HPRT1 | Hypoxanthine phosphoribosyltransferase 1 |
| H05 | NM_001002 | RPLP0 | Ribosomal protein, large, P0 |
| H06 | SA_00105 | HGDC | Human Genomic DNA Contamination |
| H07 | SA_00104 | RTC | Reverse Transcription Control |
| H08 | SA_00104 | RTC | Reverse Transcription Control |
| H09 | SA_00104 | RTC | Reverse Transcription Control |
| H10 | SA_00103 | PPC | Positive PCR Control |
| H11 | SA_00103 | PPC | Positive PCR Control |
| H12 | SA_00103 | PPC | Positive PCR Control |
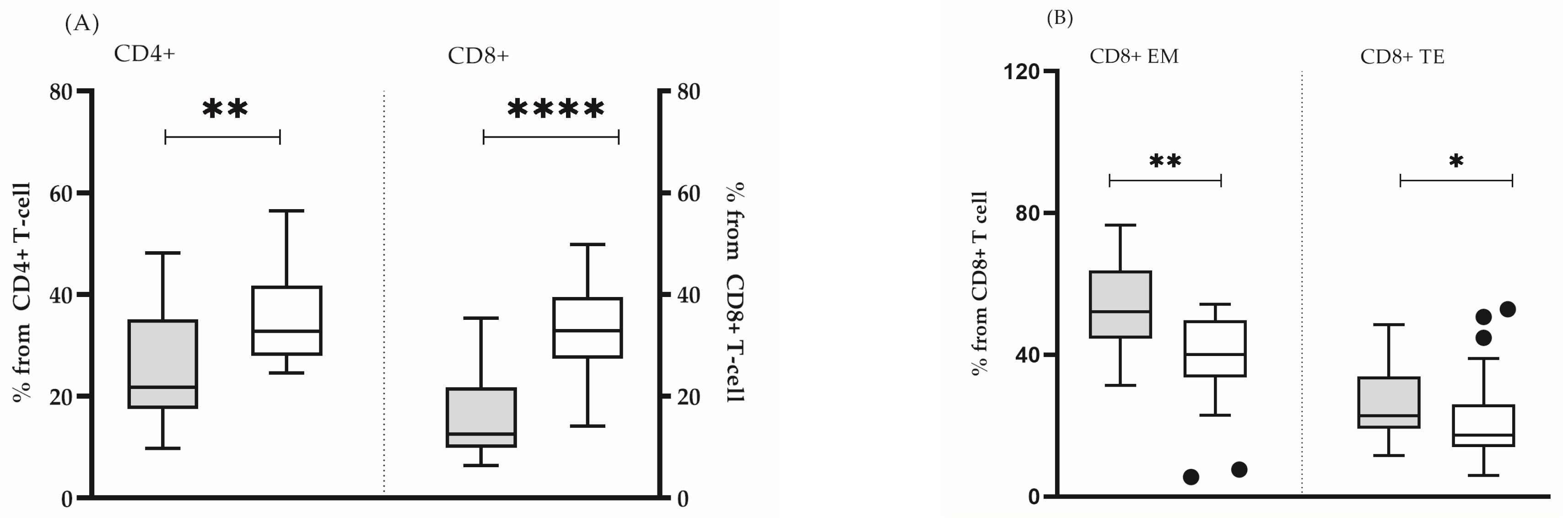
| Healthy Individuals (“Control Group”) (n = 39) | Healthy Immunosenescent Individuals (“Test Group”) (n = 16) | p Value | |
|---|---|---|---|
| years | 37.69 ± 10.0 | 44.00 ± 12.65 | 0.09 |
| male/female | 18/21 | 5/11 | 0.47 |
| Lymphocyte population | |||
| Lymphocyte (AC) | 2227 ± 518 | 1912 ± 562 | 0.05 |
| CD3+CD4+ T (AC) | 1005 ± 258 | 851 ± 213 | 0.05 |
| CD3+CD8 T (AC) | 492 ± 158 | 461 ± 188 | 0.44 |
| B cells (AC) | 238 (89–716) | 186 (71–458) | 0.17 |
| NK cells (AC) | 317 ± 163 | 262 ± 129 | 0.27 |
| CD4/CD8 | 2.0 (1.1–3.7) | 2.76 (1.1–3.1) | 0.55 |
| CD4+ naive | 32.7 (24.6–56.0) | 21.8 (9.8–48.0) | 0.005 |
| CD4+ EM | 31.8 (14.1–56.3) | 38.2 (5.0–55.5) | 0.076 |
| CD4+ CM | 25.0 (6.1–46) | 27.3 (16.3–52.0) | 0.314 |
| CD4+ TE | 5.7 (1.1–19.2) | 5.9 (1.4–21.8) | 0.545 |
| CD8+ naive | 32.9 (12–49.8) | 12.6 (6.4–35.3) | 0.0001 |
| CD8+ EM | 40.1 (5.5–54.2) | 52.1 (31.4–76.7) | 0.002 |
| CD8+ CM | 4.0 (0.3–25.2) | 4.7 (1.3–8.3) | 0.360 |
| CD8+ TE | 17.4 (6.0–52.9) | 22.8 (11.6–48.5) | 0.047 |
| CD8+CD57+CD27- | 8.1 (0.2–21) | 7.6 (1.9–48.2) | 0.206 |
| CD4+CD38ABS | 3417 (1817–4481) | 3662 (2568–7611) | 0.241 |
| CD8+CD38ABS | 1903 (1048–2788) | 2154 (1270–8351) | 0.251 |
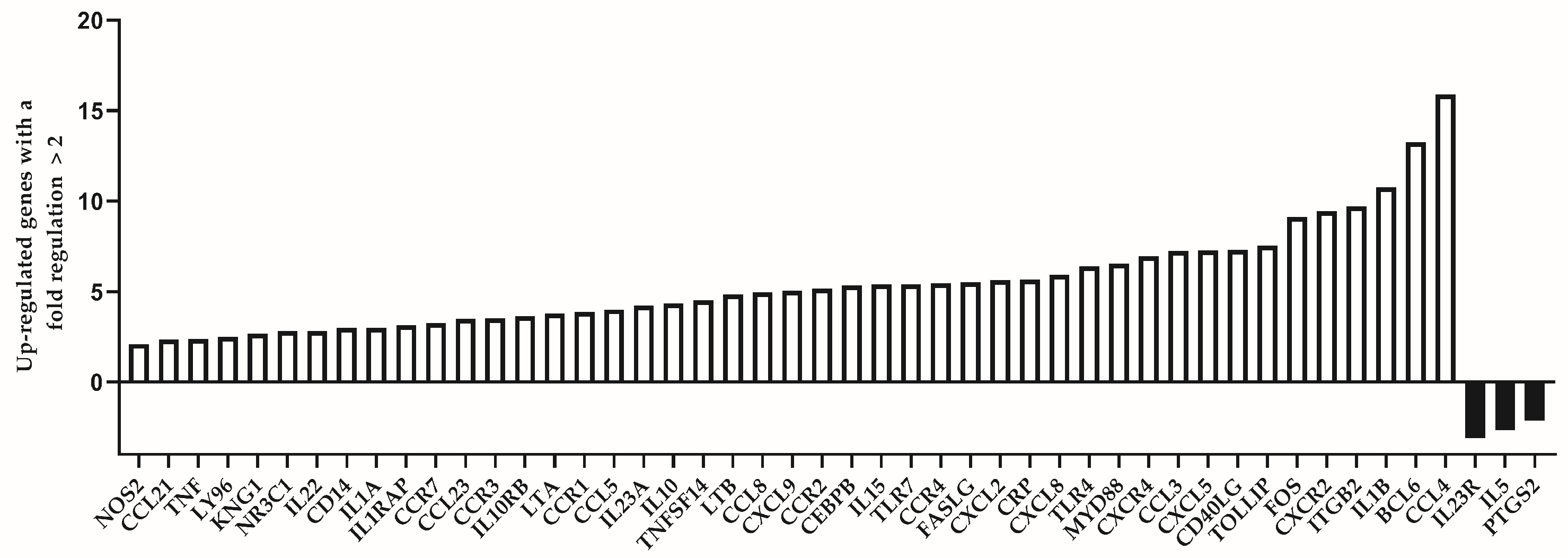
| Number | Up-Regulated Genes with a Fold Regulation > 2 | |
| Symbol | Fold Regulation | |
| 1 | NOS2 | 2.10 |
| 2 | CCL21 | 2.37 |
| 3 | TNF | 2.39 |
| 4 | LY96 | 2.49 |
| 5 | KNG1 | 2.68 |
| 6 | NR3C1 | 2.81 |
| 7 | IL22 | 2.83 |
| 8 | CD14 | 2.99 |
| 9 | IL1A | 2.99 |
| 10 | IL1RAP | 3.14 |
| 11 | CCR7 | 3.26 |
| 12 | CCL23 | 3.50 |
| 13 | CCR3 | 3.53 |
| 14 | IL10RB | 3.66 |
| 15 | LTA | 3.79 |
| 16 | CCR1 | 3.89 |
| 17 | CCL5 | 4.00 |
| 18 | IL23A | 4.24 |
| 19 | IL10 | 4.34 |
| 20 | TNFSF14 | 4.51 |
| 21 | LTB | 4.84 |
| 22 | CCL8 | 4.97 |
| 23 | CXCL9 | 5.05 |
| 24 | CCR2 | 5.16 |
| 25 | CEBPB | 5.34 |
| 26 | IL15 | 5.39 |
| 27 | TLR7 | 5.40 |
| 28 | CCR4 | 5.44 |
| 29 | FASLG | 5.53 |
| 30 | CXCL2 | 5.64 |
| 31 | CRP | 5.65 |
| 32 | CXCL8 | 5.92 |
| 33 | TLR4 | 6.41 |
| 34 | MYD88 | 6.56 |
| 35 | CXCR4 | 6.94 |
| 36 | CCL3 | 7.25 |
| 37 | CXCL5 | 7.26 |
| 38 | CD40LG | 7.31 |
| 39 | TOLLIP | 7.53 |
| 40 | FOS | 9.11 |
| 41 | CXCR2 | 9.43 |
| 42 | ITGB2 | 9.70 |
| 43 | IL1B | 10.76 |
| 44 | BCL6 | 13.25 |
| 45 | CCL4 | 15.91 |
| Down-Regulated Genes with a Fold Regulation < −2 | ||
| 1 | IL23R | −3.10 |
| 2 | IL5 | −2.66 |
| 3 | PTGS2 | −2.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolova, D.; Todorova, Y.; Hammoudeh, Z.; Rukova, B.; Emilova, R.; Aleksova, M.; Koleva, V.; Nikolova, M. Gene Expression Changes as Biomarkers of Immunosenescence in Bulgarian Individuals of Active Age. Biomedicines 2025, 13, 721. https://doi.org/10.3390/biomedicines13030721
Nikolova D, Todorova Y, Hammoudeh Z, Rukova B, Emilova R, Aleksova M, Koleva V, Nikolova M. Gene Expression Changes as Biomarkers of Immunosenescence in Bulgarian Individuals of Active Age. Biomedicines. 2025; 13(3):721. https://doi.org/10.3390/biomedicines13030721
Chicago/Turabian StyleNikolova, Dragomira, Yana Todorova, Zora Hammoudeh, Blaga Rukova, Radoslava Emilova, Milena Aleksova, Vesselina Koleva, and Maria Nikolova. 2025. "Gene Expression Changes as Biomarkers of Immunosenescence in Bulgarian Individuals of Active Age" Biomedicines 13, no. 3: 721. https://doi.org/10.3390/biomedicines13030721
APA StyleNikolova, D., Todorova, Y., Hammoudeh, Z., Rukova, B., Emilova, R., Aleksova, M., Koleva, V., & Nikolova, M. (2025). Gene Expression Changes as Biomarkers of Immunosenescence in Bulgarian Individuals of Active Age. Biomedicines, 13(3), 721. https://doi.org/10.3390/biomedicines13030721