
A Network Pharmacology Study and Experimental Validation to Identify the Potential Mechanism of Heparan Sulfate on Alzheimer’s Disease-Related Neuroinflammation


Abstract
1. Introduction
2. Results
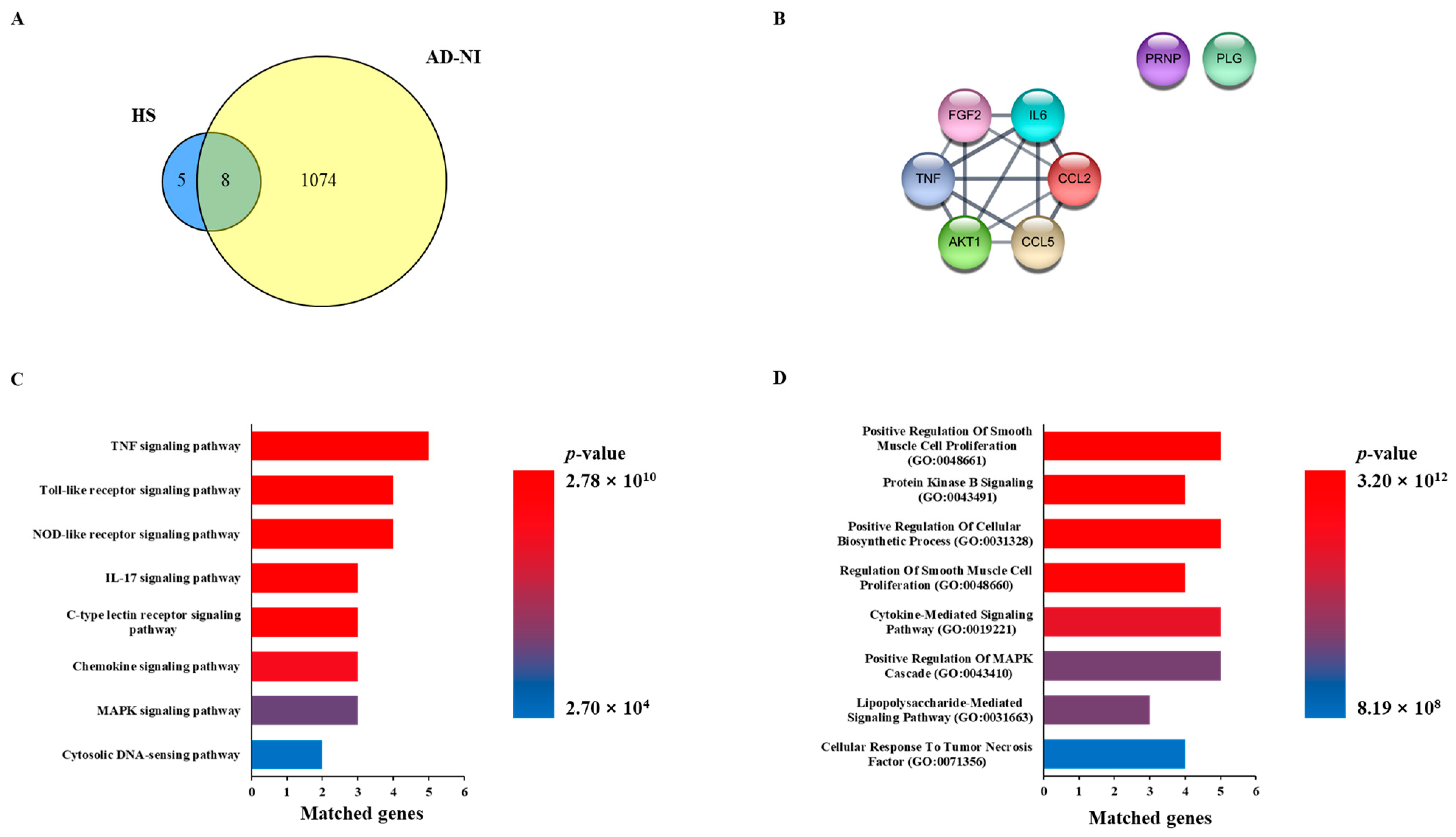
2.1. Network Pharmacology Analysis with HS and AD-NI
2.2. Functional Enrichment Analysis with HS and AD-NI
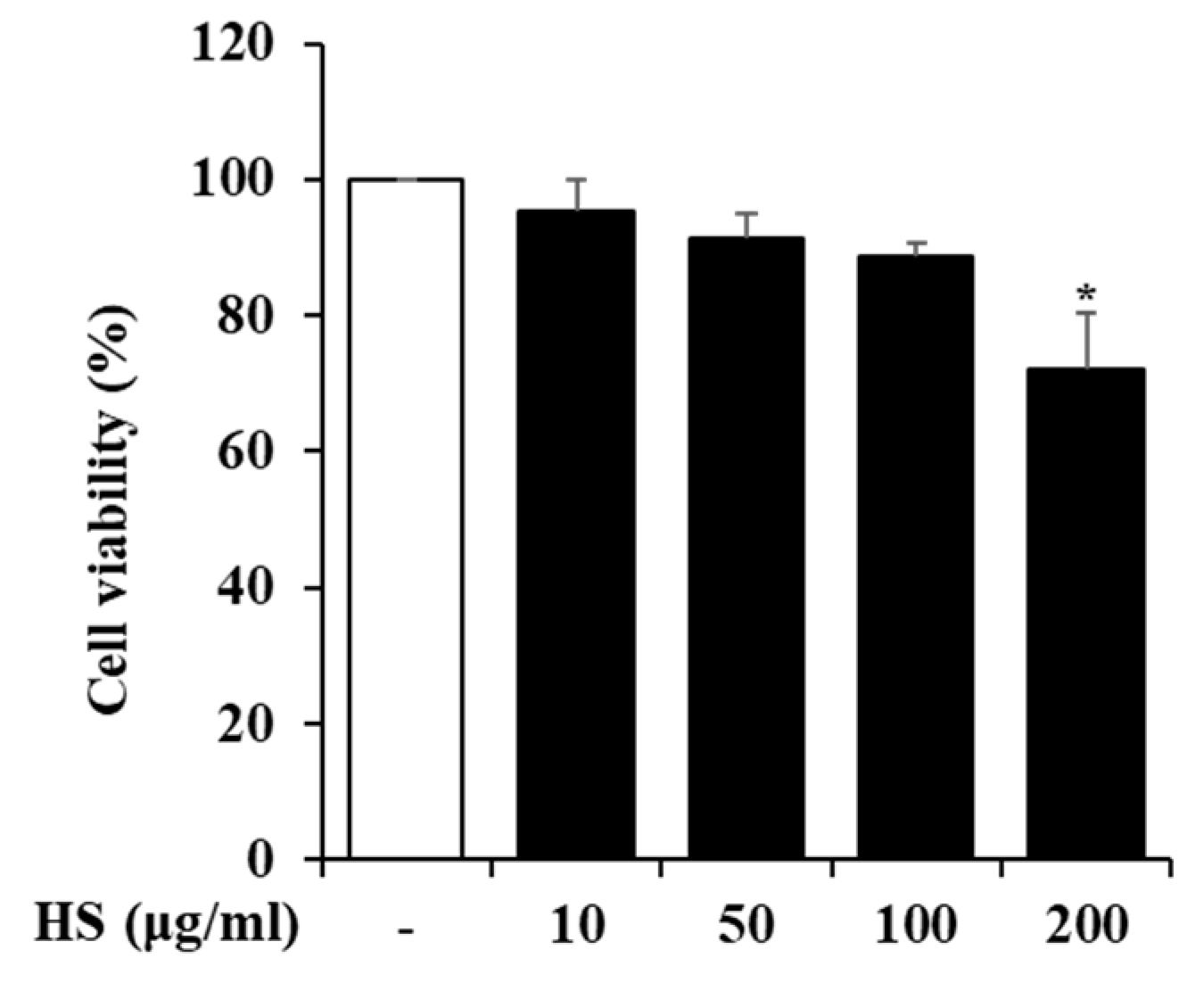
2.3. Influence of HS on BV2 Cell Viability
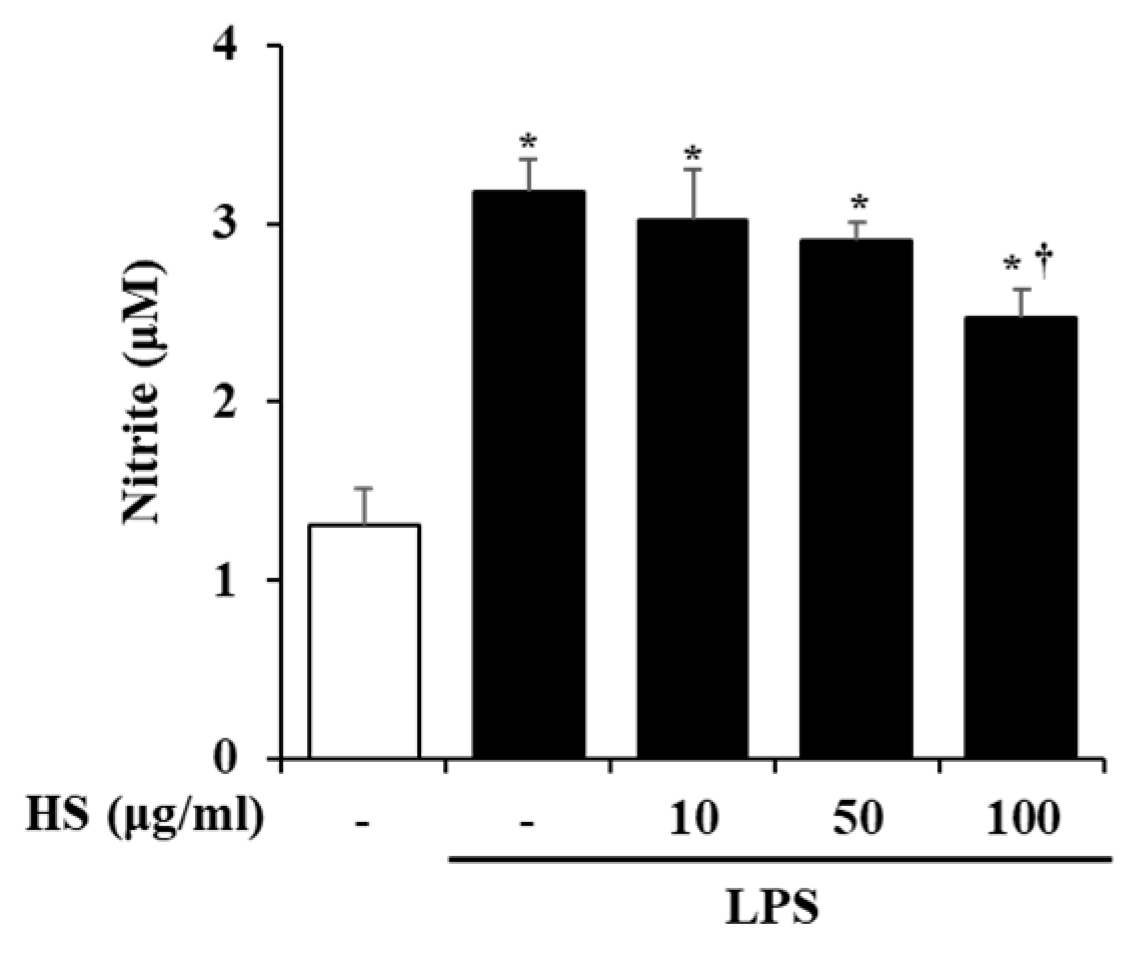
2.4. Effect of HS on Nitric Oxide (NO) Production in LPS-Stimulated BV2 Cells
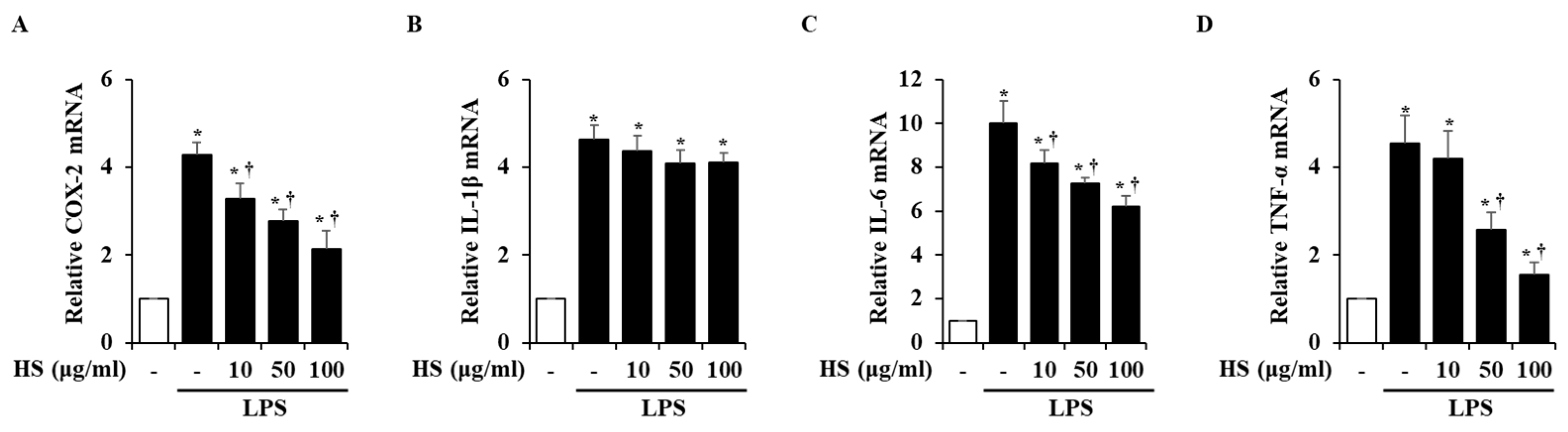
2.5. Effect of HS on Pro-Inflammatory Factor Expression in LPS-Stimulated BV2 Cells
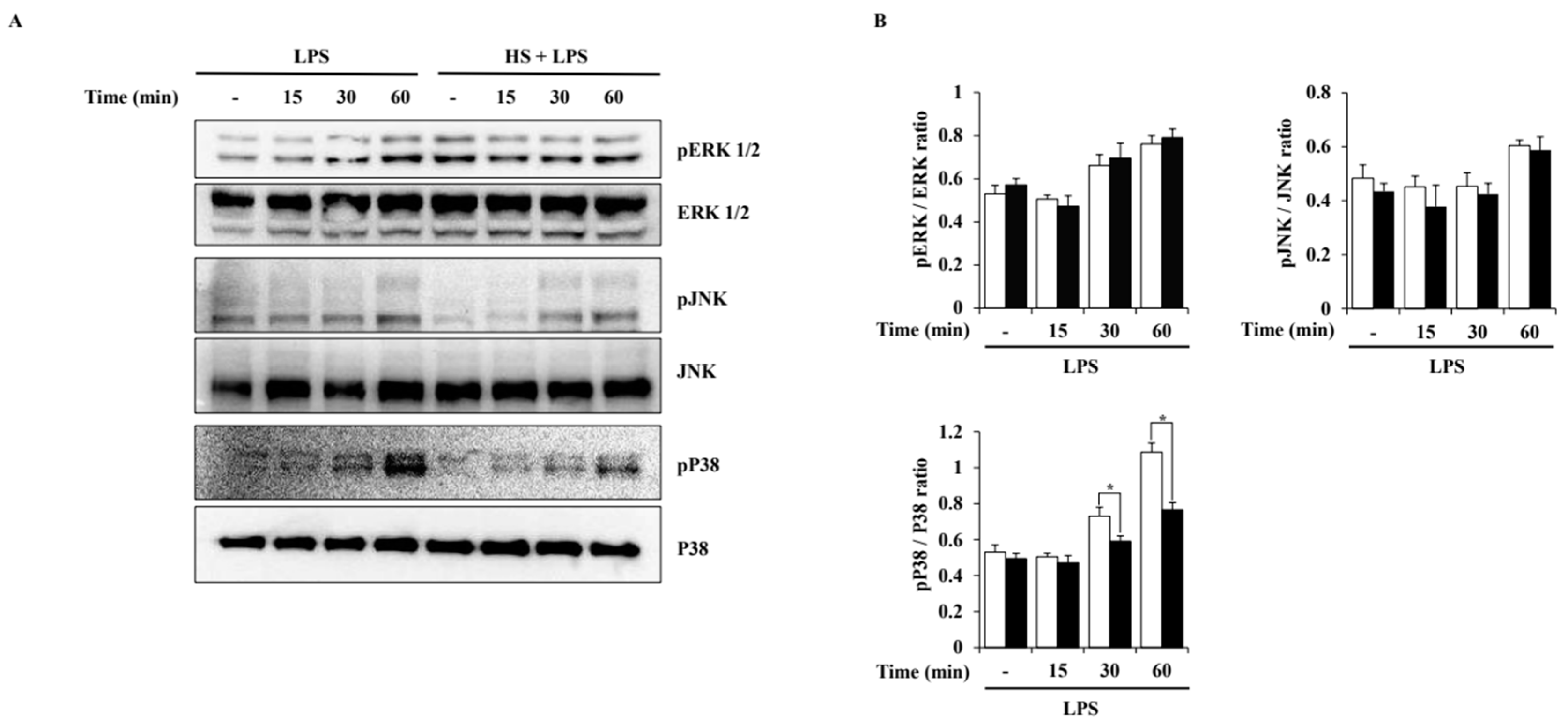
2.6. Effect of HS on Mitogen-Activated Protein Kinase (MAPK) Signaling Activation in LPS-Stimulated BV2 Cells
3. Discussion
4. Materials and Methods
4.1. Network Construction of Common Genes Between HS and AD-NI-Targeted Genes
4.2. Functional Enrichment Analysis
4.3. BV2 Cell Culture
4.4. MTT Assay
4.5. Measurement of NO
4.6. Real-Time RT-PCR
4.7. Western Blotting Assay
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Breijyeh, Z.; Karaman, R. Comprehensive review on Alzheimer’s disease: Causes and treatment. Molecules 2020, 25, 5789. [Google Scholar] [CrossRef] [PubMed]
- Porsteinsson, A.P.; Isaacson, R.; Knox, S.; Sabbagh, M.N.; Rubino, I. Diagnosis of early Alzheimer’s disease: Clinical practice in 2021. J. Prev. Alzheimer’s Dis. 2021, 8, 371–386. [Google Scholar] [CrossRef]
- Tiwari, S.; Atluri, V.; Kaushik, A.; Yndart, A.; Nair, M. Alzheimer’s disease: Pathogenesis, diagnostics, and therapeutics. Int. J. Nanomed. 2019, 14, 5541–5554. [Google Scholar] [CrossRef]
- Kim, C.K.; Lee, Y.R.; Ong, L.; Gold, M.; Kalali, A.; Sarkar, J. Alzheimer’s disease: Key insights from two decades of clinical trial failures. J. Alzheimer’s Dis. 2022, 87, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Kinney, J.W.; Bemiller, S.M.; Murtishaw, A.S.; Leisgang, A.M.; Salazar, A.M.; Lamb, B.T. Inflammation as a central mechanism in Alzheimer’s disease. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2018, 4, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, B.L.; Lord, M.S.; Melrose, J.; Whitelock, J.M. The role of heparan sulfate in inflammation, and the development of biomimetics as anti-inflammatory strategies. J. Histochem. Cytochem. 2018, 66, 321–336. [Google Scholar] [CrossRef]
- Xu, D.; Olson, J.; Cole, J.N.; van Wijk, X.M.; Brinkmann, V.; Zychlinsky, A.; Nizet, V.; Esko, J.D.; Chang, Y.-C. Heparan sulfate modulates neutrophil and endothelial function in antibacterial innate immunity. Infect. Immun. 2015, 83, 3648–3656. [Google Scholar] [CrossRef]
- Wu, L.; Zhao, N.; Jiang, W.; Wang, F. Effects of heparan sulfate from porcine mucosa on Aβ1–42-induced neurotoxicity in vitro and in vivo. Int. J. Biol. Macromol. 2022, 206, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Jiang, W.; Zhao, N.; Wang, F. Heparan sulfate from porcine mucosa promotes amyloid-beta clearance in APP/PS1 mice and alleviates Alzheimer’s pathology. Carbohydr. Polym. 2022, 285, 119205. [Google Scholar] [CrossRef] [PubMed]
- Noor, F.; Tahir ul Qamar, M.; Ashfaq, U.A.; Albutti, A.; Alwashmi, A.S.; Aljasir, M.A. Network pharmacology approach for medicinal plants: Review and assessment. Pharmaceuticals 2022, 15, 572. [Google Scholar] [CrossRef]
- Yang, L.; Yuan, S.; Wang, R.; Guo, X.; Xie, Y.; Wei, W.; Tang, L. Exploring the molecular mechanism of berberine for treating diabetic nephropathy based on network pharmacology. Int. Immunopharmacol. 2024, 126, 111237. [Google Scholar] [CrossRef]
- Nabirotchkin, S.; Peluffo, A.E.; Rinaudo, P.; Yu, J.; Hajj, R.; Cohen, D. Next-generation drug repurposing using human genetics and network biology. Curr. Opin. Pharmacol. 2020, 51, 78–92. [Google Scholar] [CrossRef]
- Kumar, B.; Thakur, A.; Dwivedi, A.R.; Kumar, R.; Kumar, V. Multi-target-directed ligands as an effective strategy for the treatment of Alzheimer’s disease. Curr. Med. Chem. 2022, 29, 1757–1803. [Google Scholar] [CrossRef] [PubMed]
- Kweon, B.; Oh, J.; Lim, Y.; Noh, G.; Yu, J.; Kim, D.; Jang, M.; Kim, D.; Bae, G. Anti-Inflammatory Effects of Honeysuckle Leaf Against Lipopolysaccharide-Induced Neuroinflammation on BV2 Microglia. Nutrients 2024, 16, 3954. [Google Scholar] [CrossRef]
- She, Y.; Shao, C.-Y.; Liu, Y.-F.; Huang, Y.; Yang, J.; Wan, H.-T. Catalpol reduced LPS induced BV2 immunoreactivity through NF-κB/NLRP3 pathways: An in Vitro and in silico study. Front. Pharmacol. 2024, 15, 1415445. [Google Scholar] [CrossRef] [PubMed]
- Grabher, B.J. Effects of Alzheimer disease on patients and their family. J. Nucl. Med. Technol. 2018, 46, 335–340. [Google Scholar] [CrossRef]
- Zhan, X.; Stamova, B.; Jin, L.-W.; DeCarli, C.; Phinney, B.; Sharp, F.R. Gram-negative bacterial molecules associate with Alzheimer disease pathology. Neurology 2016, 87, 2324–2332. [Google Scholar] [CrossRef]
- Sheng, J.G.; Bora, S.H.; Xu, G.; Borchelt, D.R.; Price, D.L.; Koliatsos, V.E. Lipopolysaccharide-induced-neuroinflammation increases intracellular accumulation of amyloid precursor protein and amyloid β peptide in APPswe transgenic mice. Neurobiol. Dis. 2003, 14, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Miklossy, J. Chronic inflammation and amyloidogenesis in Alzheimer’s disease--role of Spirochetes. J. Prev. Alzheimer’s Dis. 2008, 13, 381–391. [Google Scholar] [CrossRef]
- Cho, H.-g.; Kim, D.-U.; Oh, J.-Y.; Park, S.-J.; Kweon, B.; Bae, G.-S. Anti-Neuroinflammatory Effects of Arecae pericarpium on LPS-Stimulated BV2 Cells. Curr. Issues Mol. Biol. 2024, 46, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Lyra e Silva, N.M.; Gonçalves, R.A.; Pascoal, T.A.; Lima-Filho, R.A.; Resende, E.d.P.F.; Vieira, E.L.; Teixeira, A.L.; de Souza, L.C.; Peny, J.A.; Fortuna, J.T. Pro-inflammatory interleukin-6 signaling links cognitive impairments and peripheral metabolic alterations in Alzheimer’s disease. Transl. Psychiatry 2021, 11, 251. [Google Scholar] [CrossRef] [PubMed]
- Plantone, D.; Pardini, M.; Righi, D.; Manco, C.; Colombo, B.M.; De Stefano, N. The Role of TNF-α in Alzheimer’s Disease: A Narrative Review. Cells 2023, 13, 54. [Google Scholar] [CrossRef]
- Salter, M.W.; Stevens, B. Microglia emerge as central players in brain disease. Nat. Med. 2017, 23, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Leng, F.; Edison, P.J.N.R.N. Neuroinflammation and microglial activation in Alzheimer disease: Where do we go from here? Nat. Rev. Neurol. 2021, 17, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Muzio, L.; Viotti, A.; Martino, G. Microglia in neuroinflammation and neurodegeneration: From understanding to therapy. Front. Neurosci. 2021, 15, 742065. [Google Scholar] [CrossRef]
- Solé-Domènech, S.; Cruz, D.L.; Capetillo-Zarate, E.; Maxfield, F.R. The endocytic pathway in microglia during health, aging and Alzheimer’s disease. Ageing Res. Rev. 2016, 32, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.S.; Koh, S.-H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef]
- Spittau, B. Aging microglia—Phenotypes, functions and implications for age-related neurodegenerative diseases. Front. Aging Neurosci. 2017, 9, 194. [Google Scholar] [CrossRef]
- Rosselli, M.; Keller, R.; Dubey, R.K. Role of nitric oxide in the biology, physiology and pathophysiology of reproduction. Hum. Reprod. Update 1998, 4, 3–24. [Google Scholar] [CrossRef]
- Sharma, J.; Al-Omran, A.; Parvathy, S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Olivera, G.C.; Ren, X.; Vodnala, S.K.; Lu, J.; Coppo, L.; Leepiyasakulchai, C.; Holmgren, A.; Kristensson, K.; Rottenberg, M.E. Nitric oxide protects against infection-induced neuroinflammation by preserving the stability of the blood-brain barrier. PLoS Pathog. 2016, 12, e1005442. [Google Scholar] [CrossRef]
- Liy, P.M.; Puzi, N.N.A.; Jose, S.; Vidyadaran, S. Nitric oxide modulation in neuroinflammation and the role of mesenchymal stem cells. Exp. Biol. Med. 2021, 246, 2399–2406. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.E. Inflammation in the Pathogenesis of Chronic Diseases: The COX-2 Controversy; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007; Volume 42. [Google Scholar]
- Becher, B.; Spath, S.; Goverman, J. Cytokine networks in neuroinflammation. Nat. Rev. Immunol. 2017, 17, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Soltani Khaboushan, A.; Yazdanpanah, N.; Rezaei, N. Neuroinflammation and proinflammatory cytokines in epileptogenesis. Mol. Neurobiol. 2022, 59, 1724–1743. [Google Scholar] [CrossRef] [PubMed]
- Cacquevel, M.; Lebeurrier, N.; Cheenne, S.; Vivien, D. Cytokines in neuroinflammation and Alzheimer’s disease. Curr. Drug Targets 2004, 5, 529–534. [Google Scholar] [CrossRef]
- Corrêa, S.A.; Eales, K.L. The role of p38 MAPK and its substrates in neuronal plasticity and neurodegenerative disease. J. Signal Transduct. 2012, 2012, 649079. [Google Scholar] [CrossRef]
- Bachstetter, A.D.; Xing, B.; de Almeida, L.; Dimayuga, E.R.; Watterson, D.M.; Van Eldik, L.J. Microglial p38α MAPK is a key regulator of proinflammatory cytokine up-regulation induced by toll-like receptor (TLR) ligands or beta-amyloid (Aβ). J. Neuroinflammation 2011, 8, 79. [Google Scholar] [CrossRef] [PubMed]
- Perea, J.R.; Bolós, M.; Cuadros, R.; García, E.; García-Escudero, V.; Hernández, F.; McManus, R.M.; Heneka, M.T.; Avila, J. p38 inhibition decreases Tau toxicity in microglia and improves their phagocytic function. Mol. Neurobiol. 2022, 59, 1632–1648. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | |
|---|---|---|
| COX-2 | F (5′−3′) | GGTGGCTGTTTTGGTAGGCTG |
| R (5’−3′) | GGGTTGCTGGGGGAAGAAATG | |
| IL-1β | F (5′−3′) | CCTCGTGCTGTCGGACCCAT |
| R (5′−3′) | CAGGCTTGTGCTCTGCTTGTGA | |
| IL-6 | F (5′−3′) | CCGGAGAGGAGACTTCACAG |
| R (5′−3′) | CAGAATTGCCATTGCACAAC | |
| TNF-α | F (5′−3′) | AACTAGTGGTGCCAGCCGAT |
| R (5′−3′) | CTTCACAGAGCAATGACTCC | |
| GAPDH | F (5′−3′) | TGTGTCCGTCGTGGATCTGA |
| R (5′−3′) | TTGCTGTTGAAGTCGCAGGAG | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-U.; Kweon, B.; Oh, J.; Lim, Y.; Noh, G.; Yu, J.; Kang, H.-R.; Kwon, T.; Lee, K.y.; Bae, G.-S. A Network Pharmacology Study and Experimental Validation to Identify the Potential Mechanism of Heparan Sulfate on Alzheimer’s Disease-Related Neuroinflammation. Biomedicines 2025, 13, 103. https://doi.org/10.3390/biomedicines13010103
Kim D-U, Kweon B, Oh J, Lim Y, Noh G, Yu J, Kang H-R, Kwon T, Lee Ky, Bae G-S. A Network Pharmacology Study and Experimental Validation to Identify the Potential Mechanism of Heparan Sulfate on Alzheimer’s Disease-Related Neuroinflammation. Biomedicines. 2025; 13(1):103. https://doi.org/10.3390/biomedicines13010103
Chicago/Turabian StyleKim, Dong-Uk, Bitna Kweon, Jinyoung Oh, Yebin Lim, Gyeongran Noh, Jihyun Yu, Hyang-Rin Kang, Tackmin Kwon, Kwang youll Lee, and Gi-Sang Bae. 2025. "A Network Pharmacology Study and Experimental Validation to Identify the Potential Mechanism of Heparan Sulfate on Alzheimer’s Disease-Related Neuroinflammation" Biomedicines 13, no. 1: 103. https://doi.org/10.3390/biomedicines13010103
APA StyleKim, D.-U., Kweon, B., Oh, J., Lim, Y., Noh, G., Yu, J., Kang, H.-R., Kwon, T., Lee, K. y., & Bae, G.-S. (2025). A Network Pharmacology Study and Experimental Validation to Identify the Potential Mechanism of Heparan Sulfate on Alzheimer’s Disease-Related Neuroinflammation. Biomedicines, 13(1), 103. https://doi.org/10.3390/biomedicines13010103