
CBP/P300 Inhibition Impairs CD4+ T Cell Activation: Implications for Autoimmune Disorders

 , and
, and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RNA-Sequencing
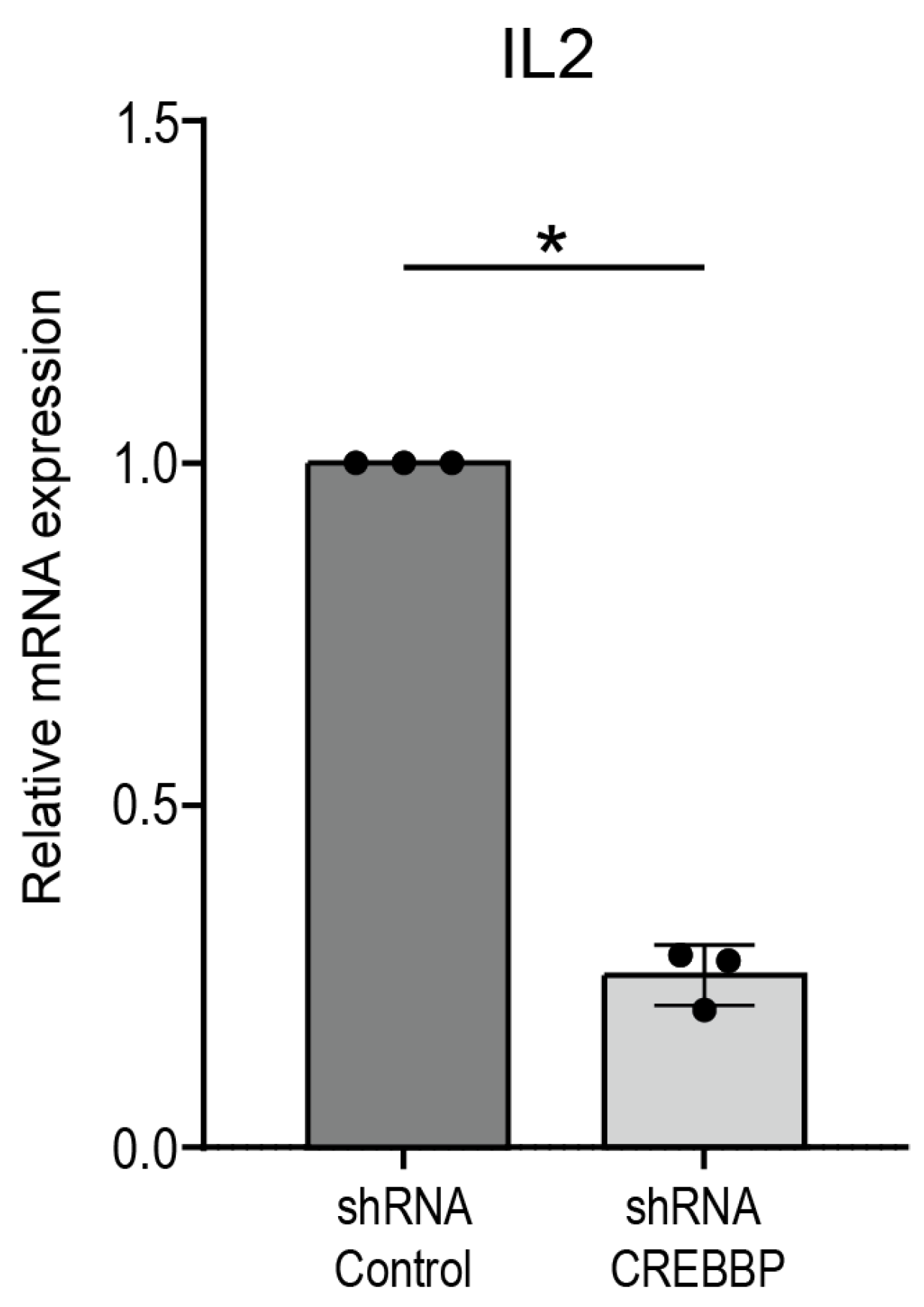
2.3. shRNA-Induced Knockdown of CREBBP in Jurkat Cells
2.4. qPCR Analysis
2.5. Luminex Assay
2.6. Analysis
3. Results
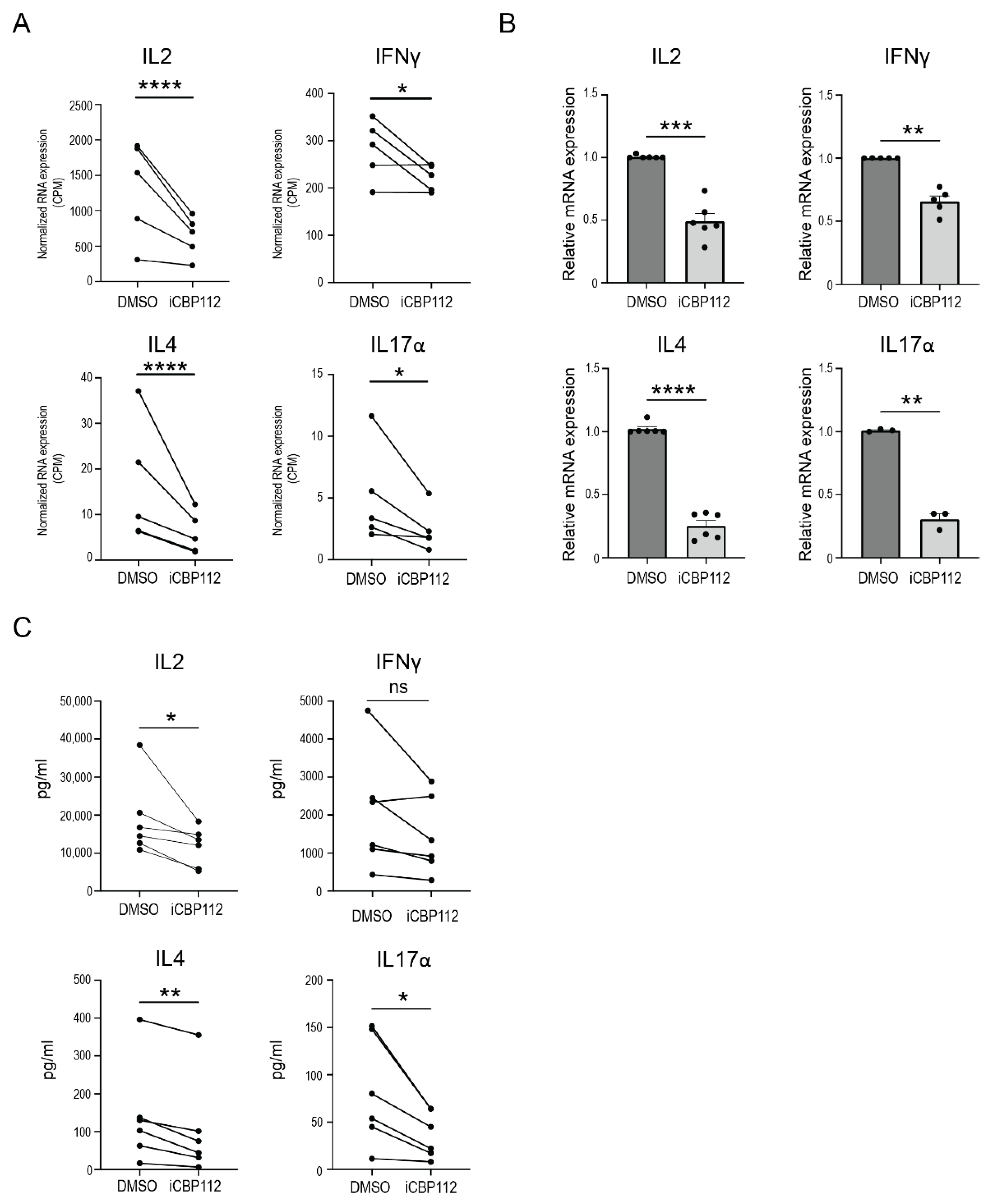
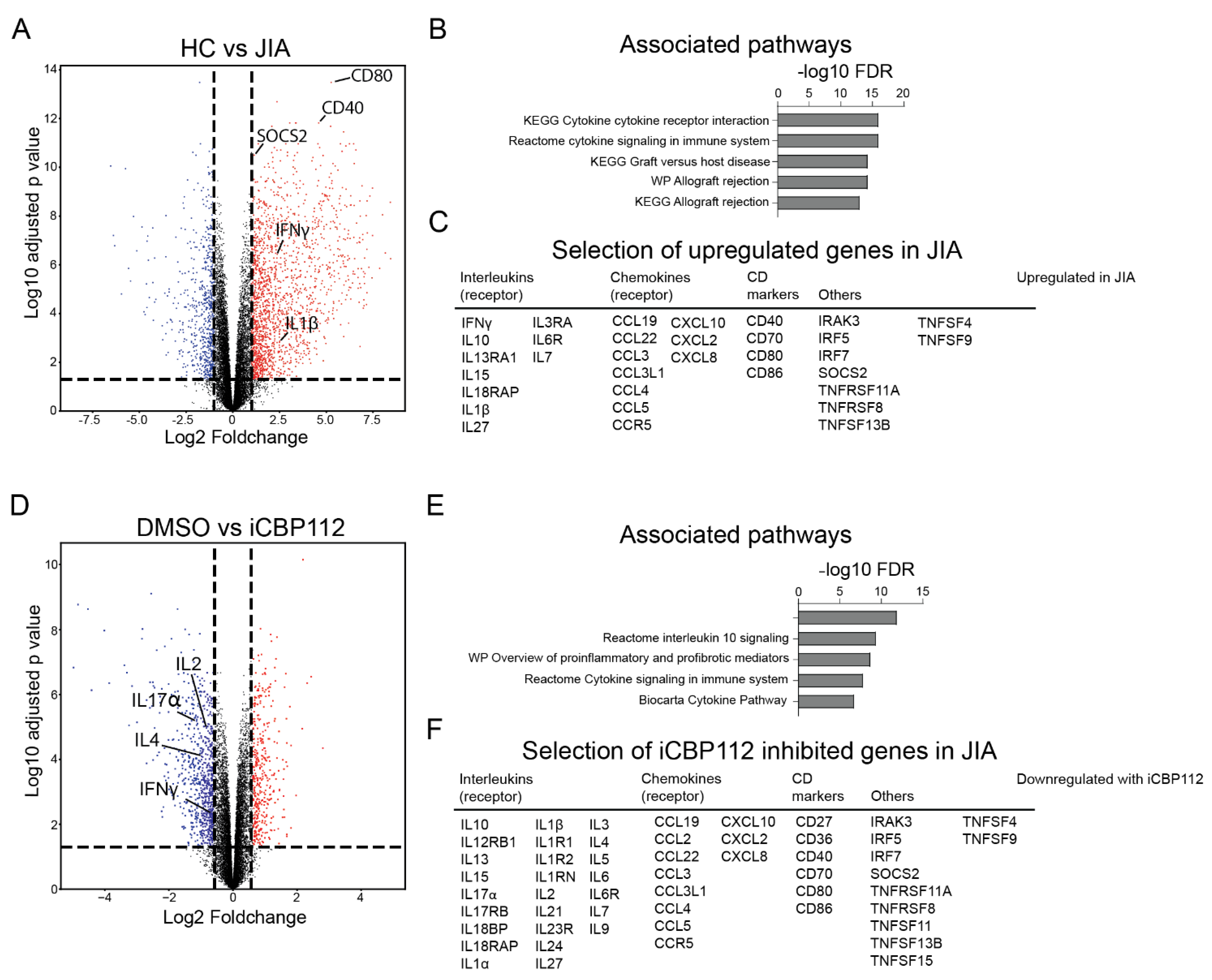
3.1. P300/CBP Regulate Pro-Inflammatory Pathways Associated with T Cell Activation
3.2. P300/CBP Inhibition Preferentially Inhibits the Expression of Proinflammatory Cytokines
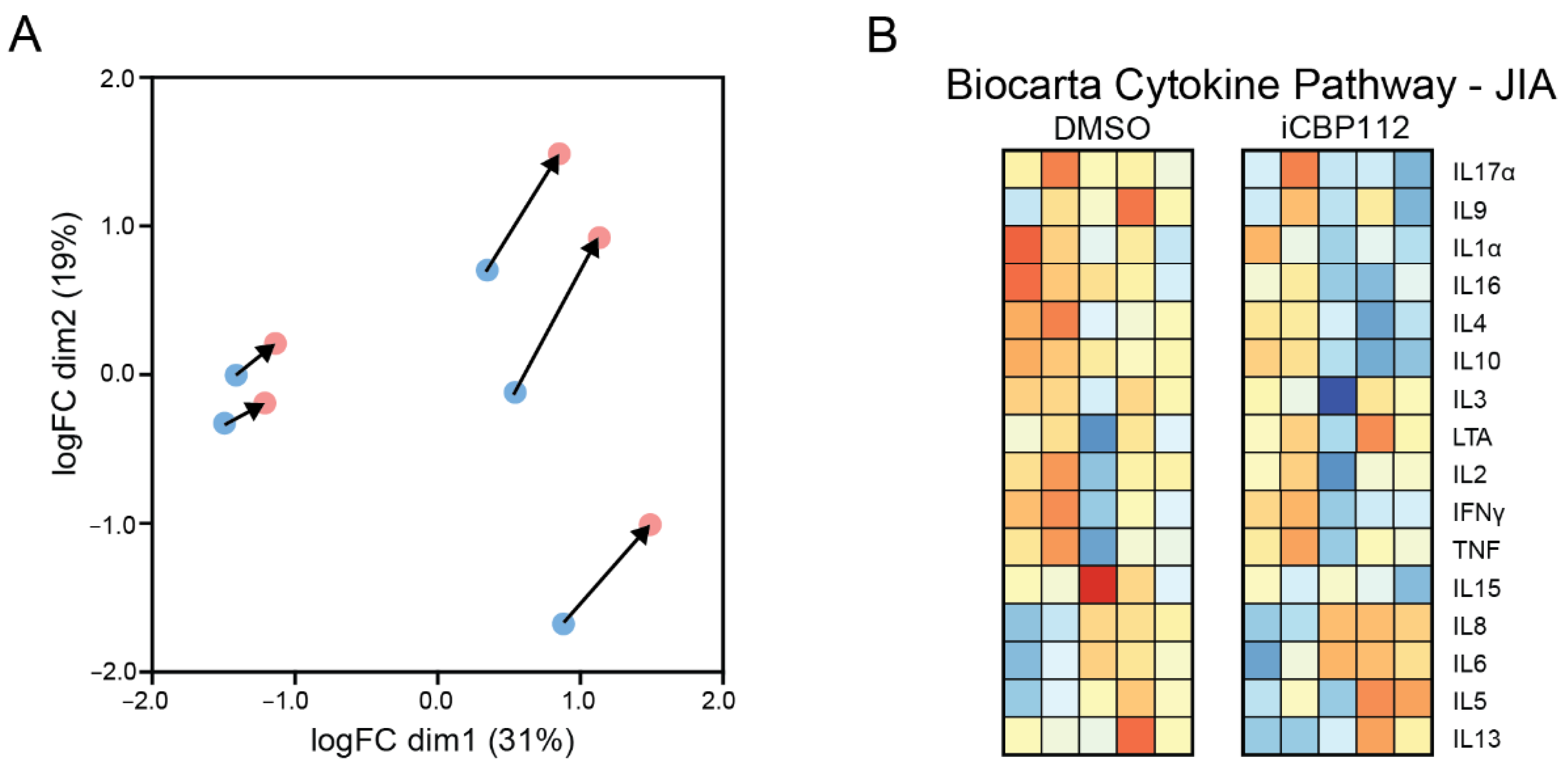
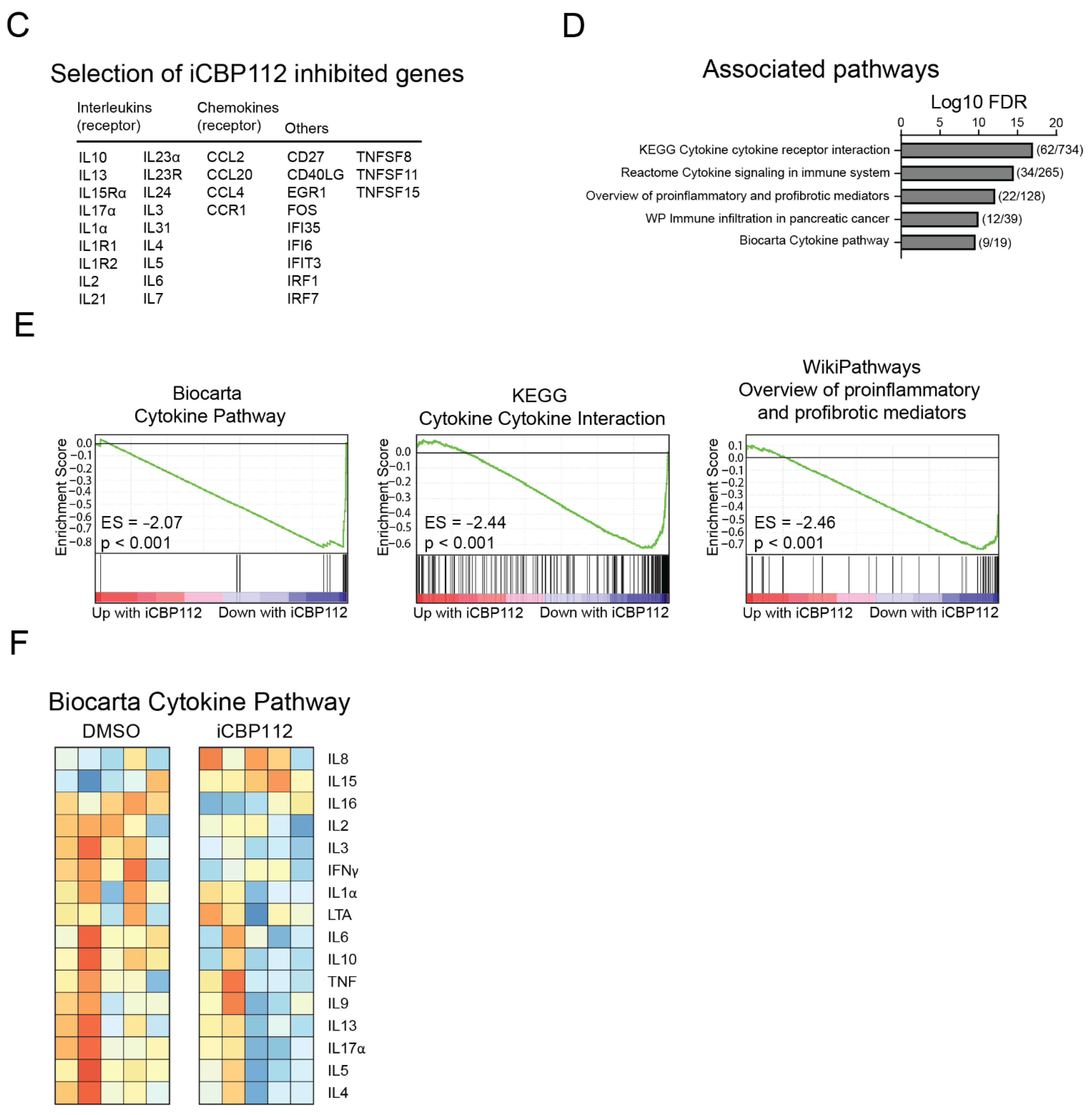
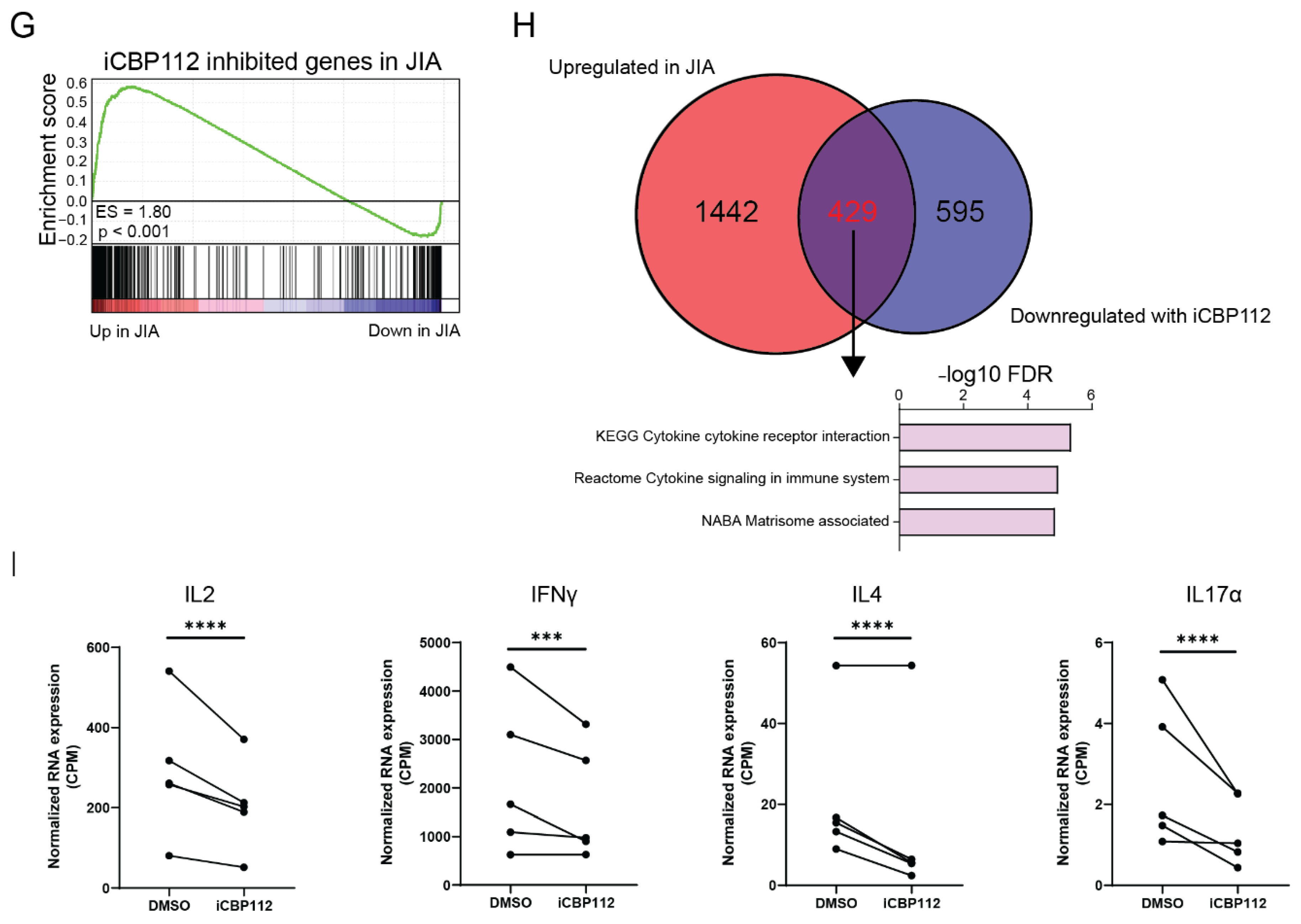
3.3. P300/CBP Inhibition Down-Regulates Genes Associated with Enhanced Expression in T Cells from Patients with Juvenile Idiopathic Arthritis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Glossary
| BRD | Bromodomain: A protein domain found in certain proteins that recognize acetylated lysine residues in histones, playing a role in gene regulation. |
| CBP | CREB-binding protein: A transcriptional coactivator that regulates gene expression by interacting with various transcription factors. |
| CD4 T cell | Helper T cell expressing CD4 receptor: A type of T lymphocyte that orchestrates immune responses by activating other immune cells, particularly B cells and macrophages. |
| CD8 T cell | Cytotoxic T cell expressing CD8 receptor: A type of T lymphocyte that directly kills infected or abnormal cells by releasing cytotoxic molecules. |
| H3K27ac | Histone H3 Lysine 27 acetylation: A histone modification associated with active gene transcription. |
| GATA3 | GATA-binding protein 3: A transcription factor important for the development and function of T cells, particularly Th2 cells. |
| IFNγ | Interferon-gamma: A cytokine produced by activated T cells and natural killer cells that plays a critical role in immune responses against viruses, intracellular bacteria, and tumors. |
| IL2 | Interleukin-2: A cytokine produced by activated T cells that stimulates the growth and differentiation of T cells and other immune cells. |
| IL4 | Interleukin-4: A cytokine produced by activated T cells, mast cells, and basophils that promotes the differentiation of naive T cells into Th2 cells and the production of antibodies by B cells. |
| IL17α | Interleukin-17α: A cytokine produced by certain T cells that mediates inflammation and is associated with autoimmune diseases. |
| iCBP112 | A P300/CBP specific inhibitor. |
| JIA | Juvenile Idiopathic Arthritis: A chronic inflammatory disease affecting children that causes joint pain, swelling, and stiffness. |
| Jurkat cells | A human T lymphocyte cell line used in research to study T cell leukemia, HIV infection, and the immune system. |
| oJIA | Oligoarticular Juvenile Idiopathic Arthritis, a JIA subtype characterized by joint inflammation, in typically fewer than five joints, during the first six months of the disease. |
| NFAT | Nuclear factor of activated T cells: A family of transcription factors important for the immune response and other cellular processes. |
| PBMCs | Peripheral blood mononuclear cells: A type of blood cell containing a single round nucleus, including lymphocytes and monocytes. |
| P300 | E1A-binding protein P300: A histone acetyltransferase and transcriptional coactivator involved in the regulation of gene expression. |
| ROR-γt | Retinoic acid receptor-related orphan receptor gamma t: A transcription factor that regulates the development and function of certain immune cells, including Th17 cells. |
| SF | Synovial Fluid of knee joint: Fluid found in the knee joint that lubricates and nourishes the articular cartilage. |
| SE | Super enhancers: Clusters of enhancers that drive the expression of genes involved in cell identity and disease. |
| shRNA | Short hairpin RNA: A sequence of RNA that makes a tight hairpin turn and can be used to silence gene expression. |
| T-BET | T-box transcription factor TBX21: A transcription factor important for the development and function of Th1 cells and cytotoxic T cells. |
| TFs | Transcription factors: Proteins that bind to DNA and regulate the transcription of genes by controlling the initiation of RNA synthesis. |
| HAT | Histone acetyltransferase: An enzyme that adds acetyl groups to histone proteins, affecting chromatin structure and gene expression. |
Appendix A

References
- Avni, O.; Lee, D.; Macian, F.; Szabo, S.J.; Glimcher, L.H.; Rao, A. Th Cell Differentiation Is Accompanied by Dynamic Changes in Histone Acetylation of Cytokine Genes. Nat. Immunol. 2002, 3, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Beacon, T.H.; Delcuve, G.P.; López, C.; Nardocci, G.; Kovalchuk, I.; van Wijnen, A.J.; Davie, J.R. The Dynamic Broad Epigenetic (H3K4me3, H3K27ac) Domain as a Mark of Essential Genes. Clin. Epigenet. 2021, 13, 138. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for Gene List Enrichment Analysis and Candidate Gene Prioritization. Nucleic Acids Res. 2009, 37 (Suppl. S2), W305–W311. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yang, B.; Liu, X.; Zhang, X.D.; Zhang, L.; Liu, T. Histone Acetyltransferases CBP/P300 in Tumorigenesis and CBP/P300 Inhibitors as Promising Novel Anticancer Agents. Theranostics 2022, 12, 4935. [Google Scholar] [CrossRef] [PubMed]
- Chrivia, J.C.; Kwok, R.P.; Lamb, N.; Hagiwara, M.; Montminy, M.R.; Goodman, R.H. Phosphorylated CREB Binds Specifically to the Nuclear Protein CBP. Nature 1993, 365, 855–859. [Google Scholar] [CrossRef]
- Creyghton, M.P.; Cheng, A.W.; Welstead, G.G.; Kooistra, T.; Carey, B.W.; Steine, E.J.; Hanna, J.; Lodato, M.A.; Frampton, G.M.; Sharp, P.A.; et al. Histone H3K27ac Separates Active from Poised Enhancers and Predicts Developmental State. Proc. Natl. Acad. Sci. USA 2010, 107, 21931–21936. [Google Scholar] [CrossRef] [PubMed]
- Crompton, J.G.; Narayanan, M.; Cuddapah, S.; Roychoudhuri, R.; Ji, Y.; Yang, W.; Patel, S.J.; Sukumar, M.; Palmer, D.C.; Peng, W.; et al. Lineage Relationship of CD8+ T Cell Subsets Is Revealed by Progressive Changes in the Epigenetic Landscape. Cell. Mol. Immunol. 2016, 13, 502–513. [Google Scholar] [CrossRef]
- Drazic, A.; Myklebust, L.M.; Ree, R.; Arnesen, T. The World of Protein Acetylation. Biochim. Et Biophys. Acta—Proteins Proteom. 2016, 1864, 1372–1401. [Google Scholar] [CrossRef]
- Eckner, R.; Ewen, M.E.; Newsome, D.; Gerdes, M.; DeCaprio, J.A.; Lawrence, J.B.; Livingston, D.M. Molecular Cloning and Functional Analysis of the Adenovirus E1A-Assoclated 300-KD Protein (P300) Reveals a Protein with Properties OI a Transcriptional Adaptor. Genes Dev. 1994, 8, 869–884. [Google Scholar] [CrossRef]
- Fields, P.E.; Lee, G.R.; Kim, S.T.; Bartsevich, V.V.; Flavell, R.A. Th2-Specific Chromatin Remodeling and Enhancer Activity in the Th2 Cytokine Locus Control Region. Immunity 2004, 21, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, T.; Kasper, L.H.; Boussouar, F.; Jeevan, T.; van Deursen, J.; Brindle, P.K. Histone Acetyltransferase CBP Is Vital To Demarcate Conventional and Innate CD8+ T-Cell Development. Mol. Cell. Biol. 2009, 29, 3894–3904. [Google Scholar] [CrossRef]
- García-Rodríguez, C.; Rao, A. Nuclear Factor of Activated T Cells (NFAT)-Dependent Transactivation Regulated by the Coactivators P300/CREB-Binding Protein (CBP). J. Exp. Med. 1998, 187, 2031–2036. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Taylor, A.; Chin, M.; Huang, H.-R.; Conery, A.R.; Mertz, J.A.; Salmeron, A.; Dakle, P.J.; Mele, D.; Cote, A.; et al. Regulatory T Cell Modulation by CBP/EP300 Bromodomain Inhibition. J. Biol. Chem. 2016, 291, 13014–13027. [Google Scholar] [CrossRef] [PubMed]
- Hammitzsch, A.; Tallant, C.; Fedorov, O.; O’mahony, A.; Brennan, P.E.; Hay, D.A.; Martinez, F.O.; Al-Mossawi, M.H.; de Wit, J.; Vecellio, M.; et al. CBP30, a Selective CBP/P300 Bromodomain Inhibitor, Suppresses Human Th17 Responses. Proc. Natl. Acad. Sci. USA 2015, 112, 10768–10773. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, R.D.; Larjo, A.; Tripathi, S.K.; Wagner, U.; Luu, Y.; Lonnberg, T.; Raghav, S.K.; Lee, L.K.; Lund, R.; Ren, B.; et al. Global Chromatin State Analysis Reveals Lineage-Specific Enhancers during the Initiation of Human T Helper 1 and T Helper 2 Cell Polarization. Immunity 2013, 38, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- He, Z.-X.; Wei, B.-F.; Zhang, X.; Gong, Y.-P.; Ma, L.-Y.; Zhao, W. 2021. Current Development of CBP/P300 Inhibitors in the Last Decade. Eur. J. Med. Chem. 2020, 209, 112861. [Google Scholar] [CrossRef] [PubMed]
- Hulsen, T.; de Vlieg, J.; Alkema, W. BioVenn—A Web Application for the Comparison and Visualization of Biological Lists Using Area-Proportional Venn Diagrams. BMC Genom. 2008, 9, 488. [Google Scholar] [CrossRef] [PubMed]
- De Jager, W.; Prakken, B.J.; Bijlsma, J.W.; Kuis, W.; Rijkers, G.T. Improved Multiplex Immunoassay Performance in Human Plasma and Synovial Fluid Following Removal of Interfering Heterophilic Antibodies. J. Immunol. Methods 2005, 300, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Kasper, L.H.; Fukuyama, T.; Biesen, M.A.; Boussouar, F.; Tong, C.; De Pauw, A.; Murray, P.J.; Van Deursen, J.M.; Brindle, P.K. Conditional Knockout Mice Reveal Distinct Functions for the Global Transcriptional Coactivators CBP and P300 in T-Cell Development. Mol. Cell. Biol. 2006, 26, 789–809. [Google Scholar] [CrossRef]
- Krošel, M.; Gabathuler, M.; Maciukiewicz, M.; Moser, L.; Lee, G.I.; Marks, M.; Tomšič, M.; Distler, O.; Ospelt, C.; Klein, K. Individual Functions of the Histone Acetyl Transferases CBP and P300 in Regulating the Inflammatory Response of Synovial Fibroblasts. J. Autoimmun. 2021, 123, 102709. [Google Scholar] [CrossRef] [PubMed]
- Kung, A.L.; Rebel, V.I.; Bronson, R.T.; Ch’Ng, L.-E.; Sieff, C.A.; Livingston, D.M.; Yao, T.-P. Gene Dose-Dependent Control of Hematopoiesis and Hematologic Tumor Suppression by CBP. Genes Dev. 2000, 14, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. Voom: Precision Weights Unlock Linear Model Analysis Tools for RNA-Seq Read Counts. Genome Biol. 2014, 15, R29. [Google Scholar] [CrossRef] [PubMed]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.-F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1α-Responsive Genes Involved in Oxidative Phosphorylation Are Coordinately Downregulated in Human Diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.L.T.; Jones, S.A.; Prier, J.E.; Russ, B.E. Transcriptional Enhancers in the Regulation of T Cell Differentiation. Front. Immunol. 2015, 6, 462. [Google Scholar] [CrossRef] [PubMed]
- Ogryzko, V.V.; Schiltz, R.; Russanova, V.; Howard, B.H.; Nakatani, Y. The Transcriptional Coactivators P300 and CBP Are Histone Acetyltransferases. Cell 1996, 87, 953–959. [Google Scholar] [CrossRef]
- Sullivan, K.E. Inflammation in Juvenile Idiopathic Arthritis. Rheum. Dis. Clin. N. Am. 2007, 33, 365–388. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, B.A.; Laxer, R.M.; Andersson, U.; Silverman, E.D. Local Synthesis of Both Macrophage and T Cell Cytokines by Synovial Fluid Cells from Children with Juvenile Rheumatoid Arthritis. Clin. Exp. Immunol. 1994, 96, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Julé, A.M.; Hoyt, K.J.; Wei, K.; Gutierrez-Arcelus, M.; Taylor, M.L.; Ng, J.; Lederer, J.A.; Case, S.M.; Chang, M.H.; Cohen, E.M.; et al. Th1 Polarization Defines the Synovial Fluid T Cell Compartment in Oligoarticular Juvenile Idiopathic Arthritis. JCI Insight 2021, 6, e149185. [Google Scholar] [CrossRef]
- O’Shea, J.J.; Lahesmaa, R.; Vahedi, G.; Laurence, A.; Kanno, Y. Genomic Views of STAT Function in CD4+ T Helper Cell Differentiation. Nat. Rev. Immunol. 2011, 11, 239–250. [Google Scholar] [CrossRef]
- Peeters, J.G.; Vervoort, S.J.; Tan, S.C.; Mijnheer, G.; de Roock, S.; Vastert, S.J.; Nieuwenhuis, E.E.; van Wijk, F.; Prakken, B.J.; Creyghton, M.P.; et al. Inhibition of Super-Enhancer Activity in Autoinflammatory Site-Derived T Cells Reduces Disease-Associated Gene Expression. Cell Rep. 2015, 12, 1986–1996. [Google Scholar] [CrossRef] [PubMed]
- Piccirillo, A.R.; Cattley, R.T.; D’Cruz, L.M.; Hawse, W.F. Histone Acetyltransferase CBP Is Critical for Conventional Effector and Memory T-Cell Differentiation in Mice. J. Biol. Chem. 2019, 294, 2397–2406. [Google Scholar] [CrossRef]
- Rada-Iglesias, A.; Wysocka, J. One Genome, Many Epigenomes Epigenomics of Human Embryonic Stem Cells and Induced Pluripotent Stem Cells: Insights into Pluripotency and Implications for Disease. Genome Med. 2011, 3, 36. [Google Scholar] [CrossRef]
- Raisner, R.; Kharbanda, S.; Jin, L.; Jeng, E.; Chan, E.; Merchant, M.; Haverty, P.M.; Bainer, R.; Cheung, T.; Arnott, D.; et al. Enhancer Activity Requires CBP/P300 Bromodomain-Dependent Histone H3K27 Acetylation. Cell Rep. 2018, 24, 1722–1729. [Google Scholar] [CrossRef] [PubMed]
- Roh, T.-Y.; Cuddapah, S.; Zhao, K. Active Chromatin Domains Are Defined by Acetylation Islands Revealed by Genome-Wide Mapping. Genes Dev. 2005, 19, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Roh, T.-Y.; Wei, G.; Farrell, C.M.; Zhao, K. Genome-Wide Prediction of Conserved and Nonconserved Enhancers by Histone Acetylation Patterns. Genome Res. 2007, 17, 74–81. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Vahedi, G.; Kanno, Y.; Furumoto, Y.; Jiang, K.; Parker, S.C.J.; Erdos, M.R.; Davis, S.R.; Roychoudhuri, R.; Restifo, N.P.; Gadina, M.; et al. Super-Enhancers Delineate Disease-Associated Regulatory Nodes in T Cells. Nature 2015, 520, 558–562. [Google Scholar] [CrossRef]
- Vahedi, G.; Takahashi, H.; Nakayamada, S.; Sun, H.-W.; Sartorelli, V.; Kanno, Y.; O’shea, J.J. STATs Shape the Active Enhancer Landscape of T Cell Populations. Cell 2012, 151, 981–993. [Google Scholar] [CrossRef]
- Wu, D.; Smyth, G.K. Smyth. Camera: A Competitive Gene Set Test Accounting for Inter-Gene Correlation. Nucleic Acids Res. 2012, 40, e133. [Google Scholar] [CrossRef]
- Zeng, L.; Zhou, M.-M. Bromodomain: An Acetyl-Lysine Binding Domain. FEBS Lett. 2002, 513, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Wen, H.; Shi, X. CBP/p300: Intramolecular and intermolecular regulations. Front. Biol. 2018, 13, 168–179. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Picavet, L.W.; Samat, A.A.K.; Calis, J.; Nijhuis, L.; Scholman, R.; Mokry, M.; Tough, D.F.; Prinjha, R.K.; Vastert, S.J.; van Loosdregt, J. CBP/P300 Inhibition Impairs CD4+ T Cell Activation: Implications for Autoimmune Disorders. Biomedicines 2024, 12, 1344. https://doi.org/10.3390/biomedicines12061344
Picavet LW, Samat AAK, Calis J, Nijhuis L, Scholman R, Mokry M, Tough DF, Prinjha RK, Vastert SJ, van Loosdregt J. CBP/P300 Inhibition Impairs CD4+ T Cell Activation: Implications for Autoimmune Disorders. Biomedicines. 2024; 12(6):1344. https://doi.org/10.3390/biomedicines12061344
Chicago/Turabian StylePicavet, Lucas Wilhelmus, Anoushka A. K. Samat, Jorg Calis, Lotte Nijhuis, Rianne Scholman, Michal Mokry, David F. Tough, Rabinder K. Prinjha, Sebastiaan J. Vastert, and Jorg van Loosdregt. 2024. "CBP/P300 Inhibition Impairs CD4+ T Cell Activation: Implications for Autoimmune Disorders" Biomedicines 12, no. 6: 1344. https://doi.org/10.3390/biomedicines12061344
APA StylePicavet, L. W., Samat, A. A. K., Calis, J., Nijhuis, L., Scholman, R., Mokry, M., Tough, D. F., Prinjha, R. K., Vastert, S. J., & van Loosdregt, J. (2024). CBP/P300 Inhibition Impairs CD4+ T Cell Activation: Implications for Autoimmune Disorders. Biomedicines, 12(6), 1344. https://doi.org/10.3390/biomedicines12061344