
Prospects for Prostate Cancer Chemotherapy: Cytotoxic Evaluation and Mechanistic Insights of Quinolinequinones with ADME/PK Profile

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Evaluation
2.1.1. In Vitro Single-Dose Anticancer Screening by NCI
2.1.2. In Vitro Five-Dose Anticancer Screening by NCI
2.1.3. MTT Assay
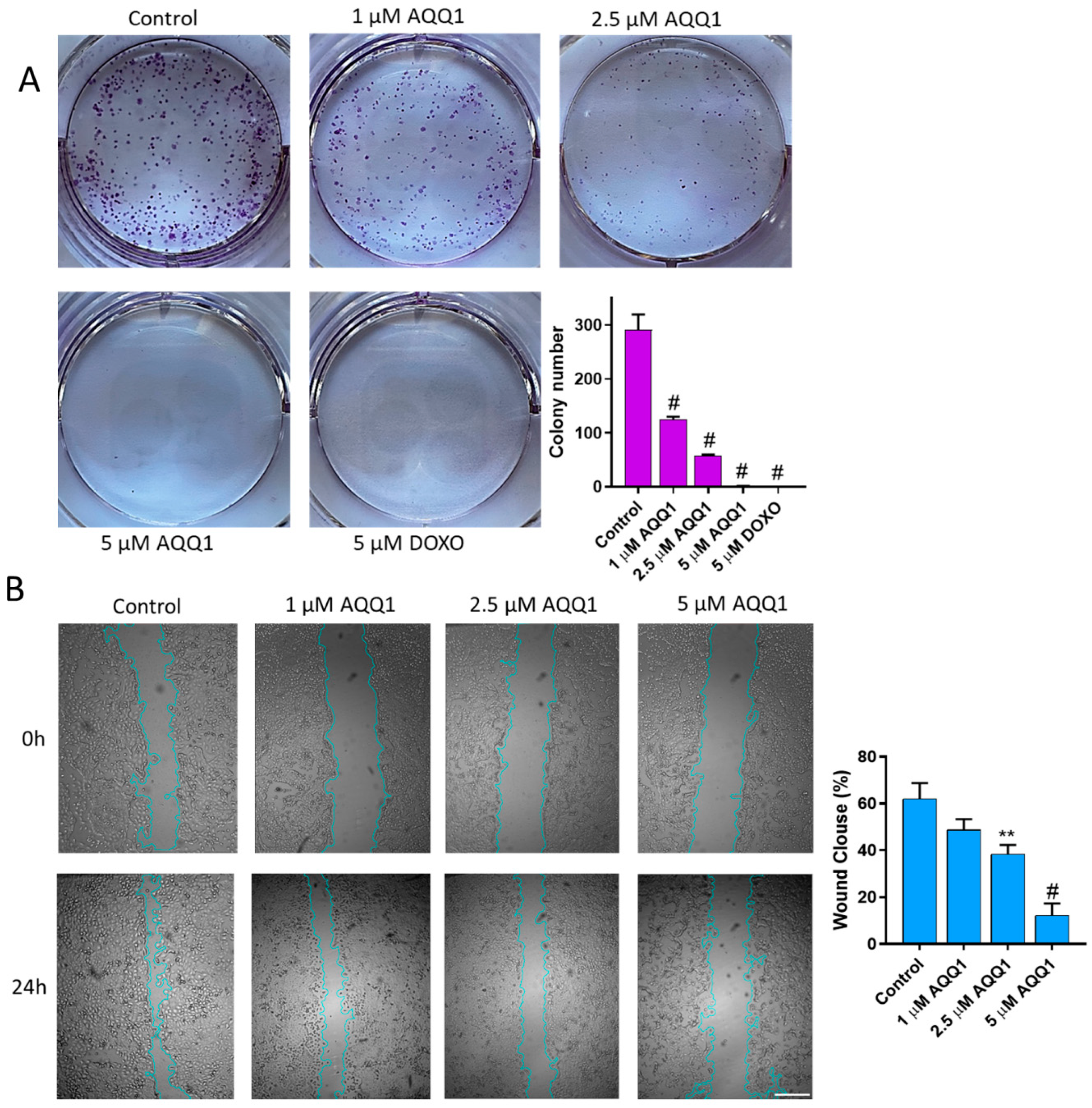
2.1.4. Colony Formation Assay
2.1.5. Scratch Assay
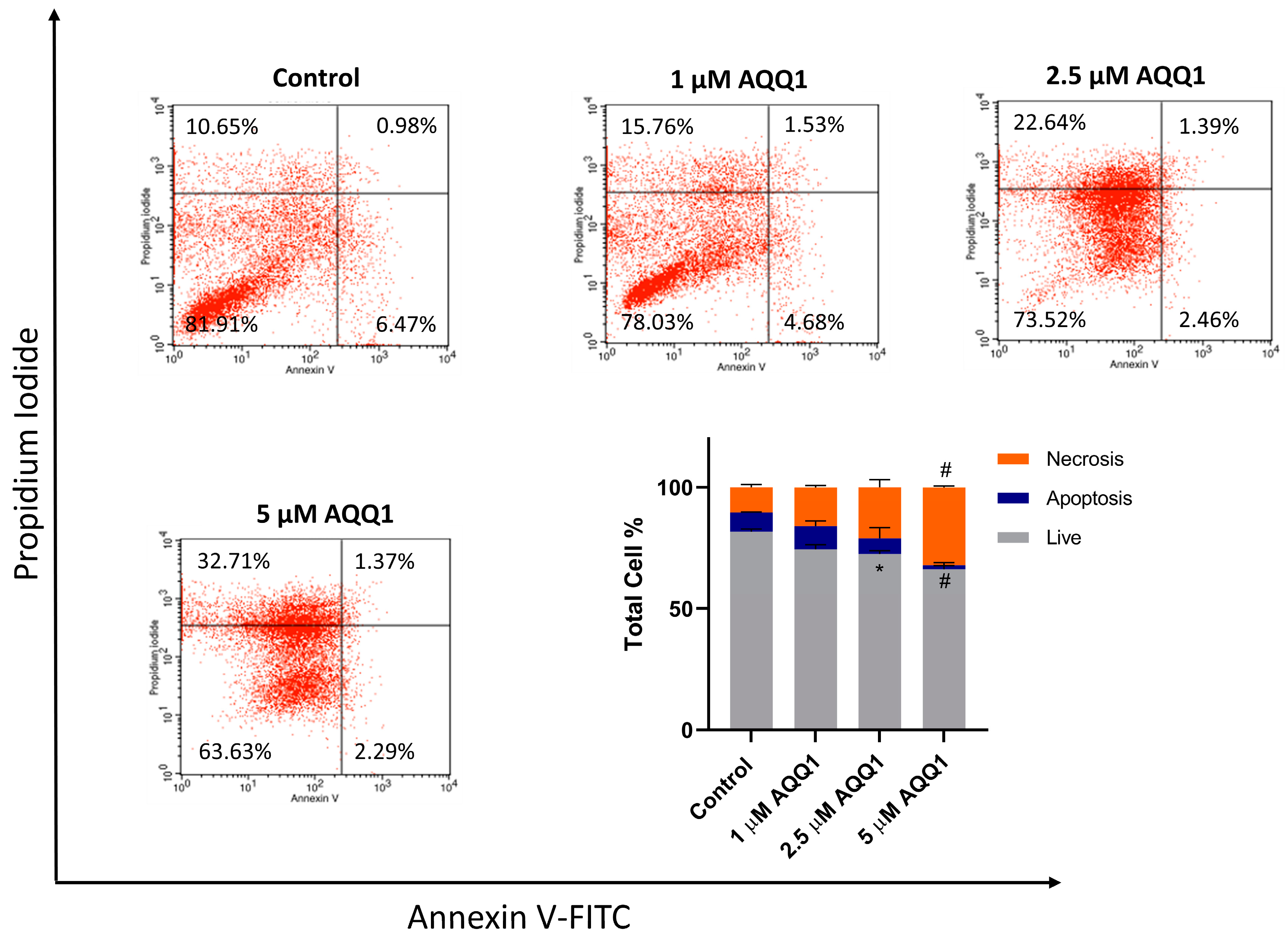
2.1.6. Apoptosis Necrosis Analysis
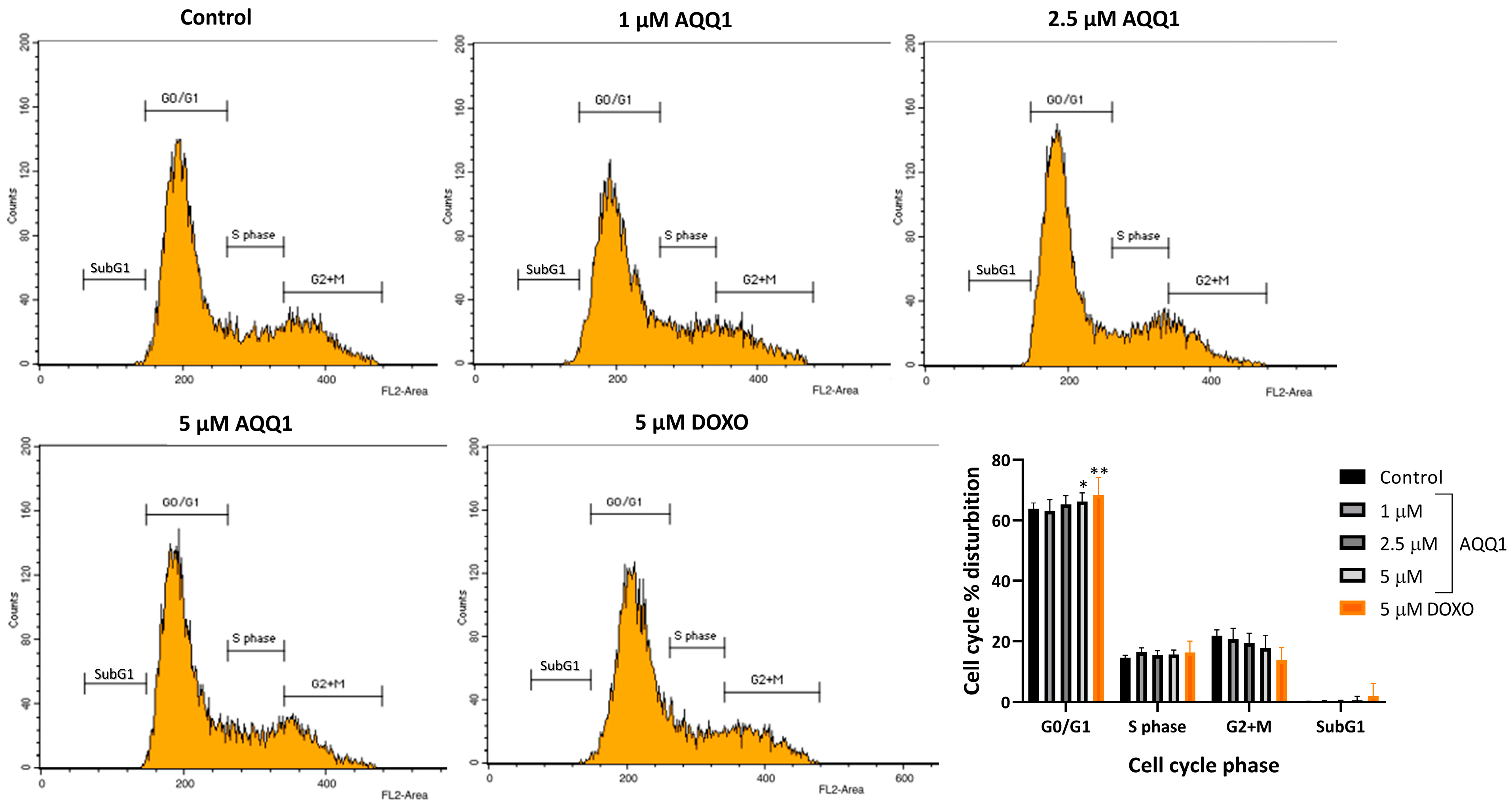
2.1.7. Cell Cycle Assay
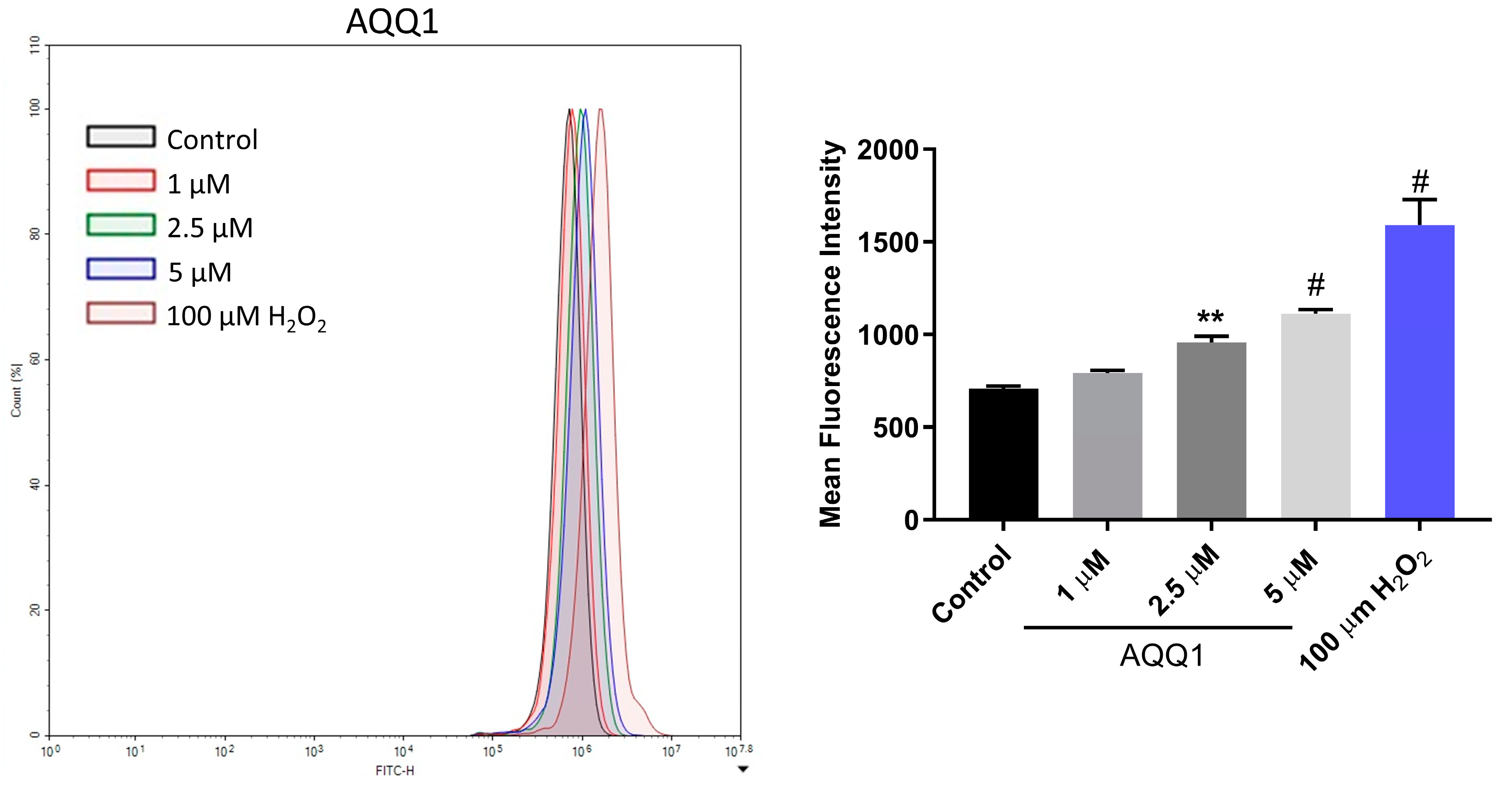
2.1.8. Oxidative Stress
2.1.9. Statistical Analysis
2.2. ADME and PK Profiling
2.2.1. In Vitro Metabolic Stability Study
2.2.2. In Vivo Bioavailability Study of AQQ1 and AQQ2 in Male Sprague-Dawley Rats
2.2.3. Bioanalytical Method (LC/MS/MS) for AQQ1 and AQQ2
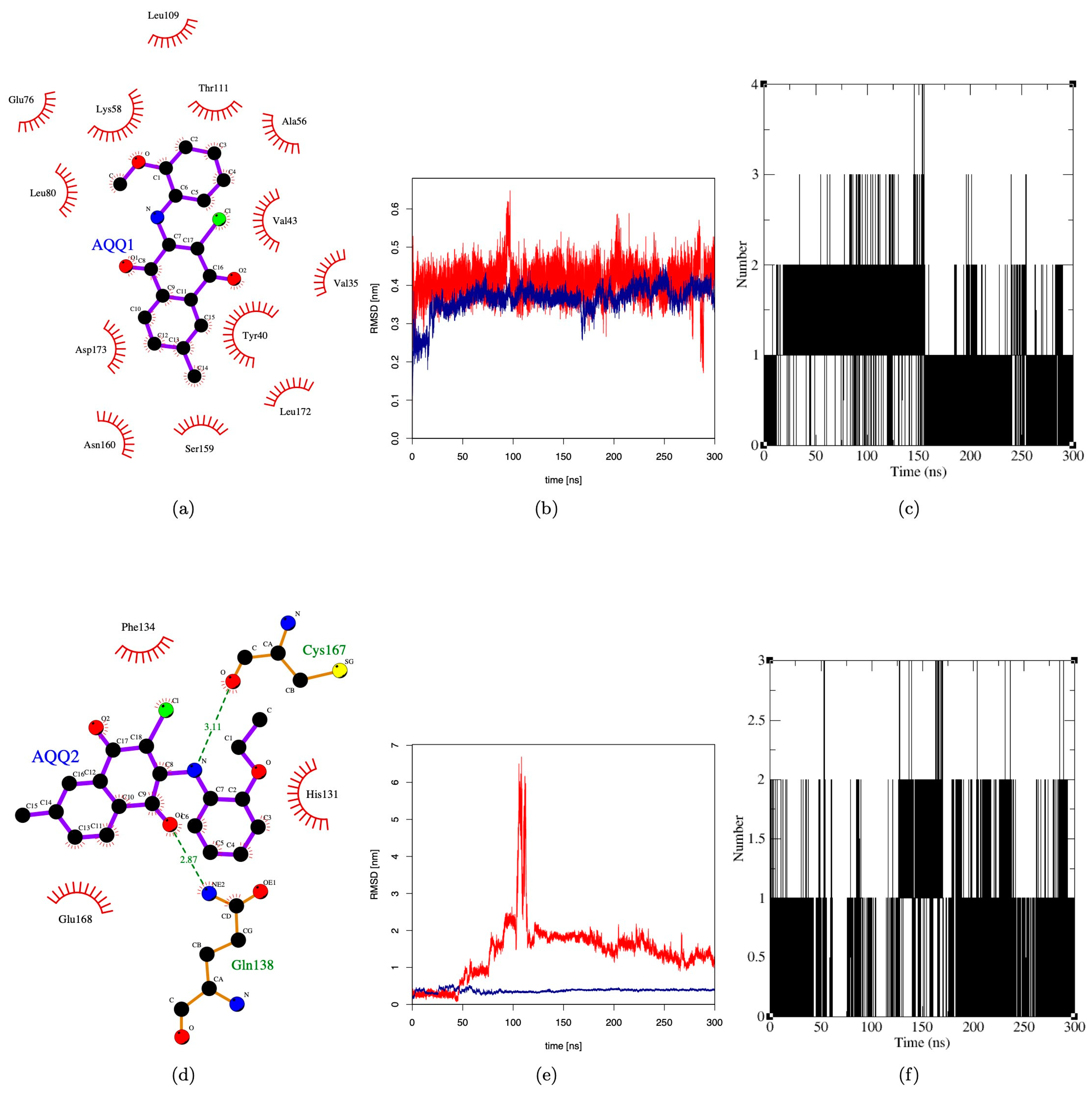
2.3. Molecular Docking and Molecular Dynamics
3. Results and Discussion
3.1. Biological Evaluation
3.1.1. Preliminary Anticancer Screening
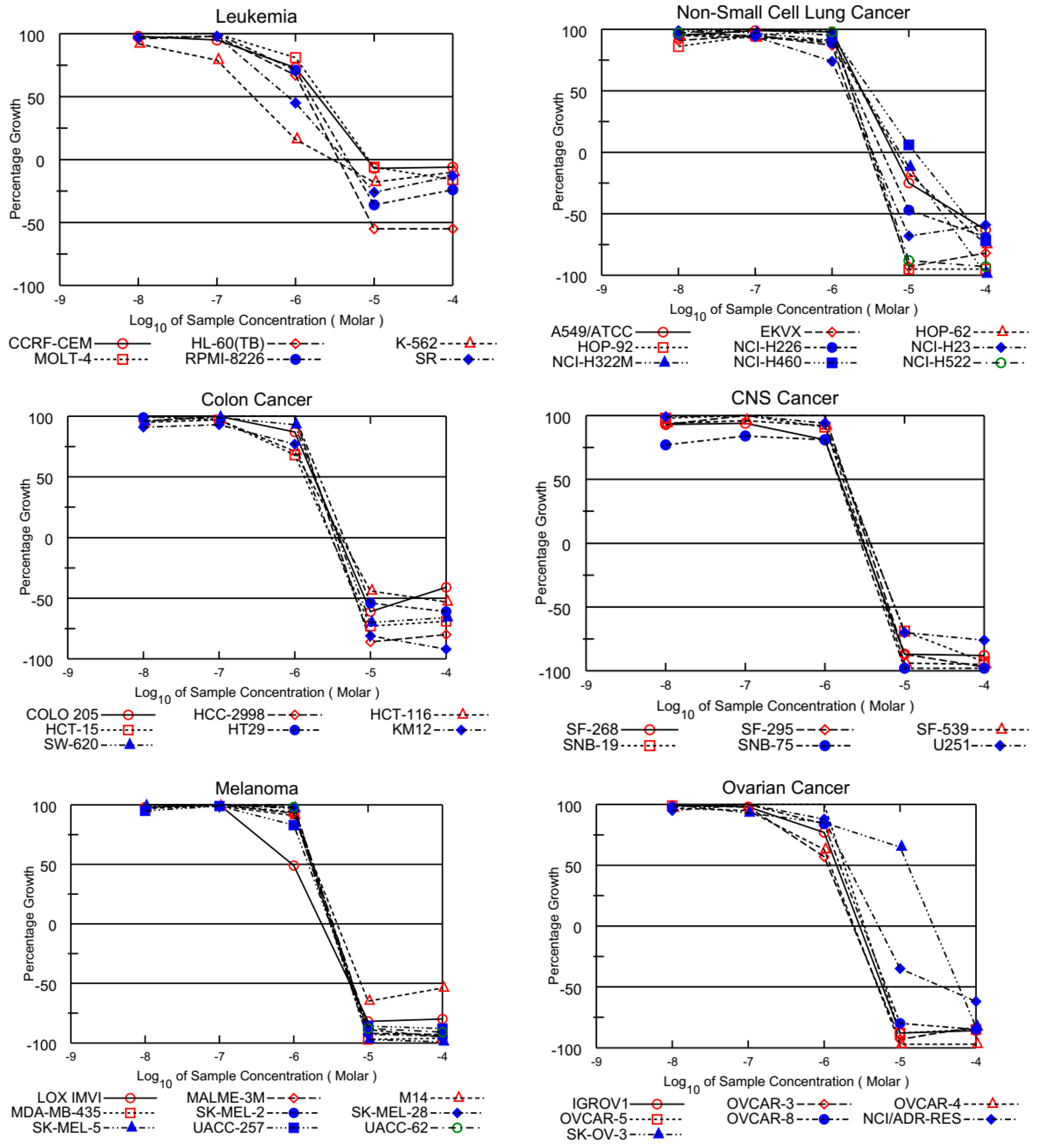
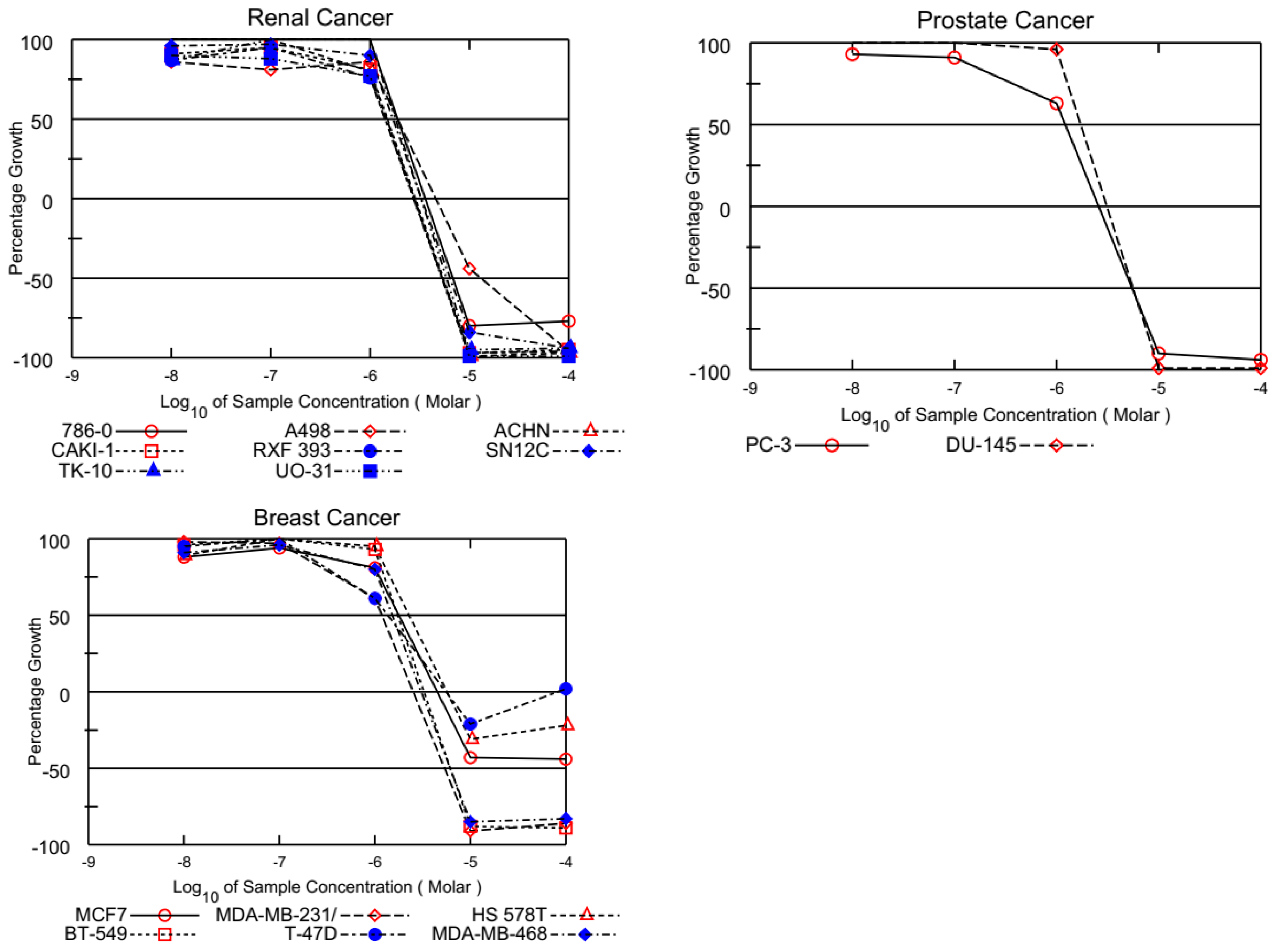
3.1.2. In Vitro Full Panel Five-Dose 60-Cell Lines Assay
3.1.3. Cytotoxicity Study of AQQ1 and AQQ2 with MTT Assay
3.2. Pharmacokinetic Profiling
3.2.1. In Vitro ADME Studies
3.2.2. In Vivo Pharmacokinetic Studies
3.3. In Silico Molecular Interaction Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- What Is Colorectal Cancer? Available online: https://www.cdc.gov/cancer/colorectal/basic_info/what-is-colorectal-cancer.htm#:~:text=Colorectal%20cancer%20is%20a%20disease,the%20colon%20to%20the%20anus (accessed on 1 May 2023).
- Willett, W.C. Diet and cancer: One view at the start of the millennium. Cancer Epidemiol. Biomark. Prev. 2001, 10, 3–8. [Google Scholar]
- Skibber, J.; Minsky, B.; Hoff, P. Cancer of the Colon. In Principles & Practice of Oncology; DeVita, V.T., Hellman, S., Rosenberg, S.A., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2001; pp. 1216–1271. [Google Scholar]
- Markowitz, S.D.; Dawson, D.M.; Willis, J.; Willson, J.K. Focus on colon cancer. Cancer Cell 2002, 1, 233–236. [Google Scholar] [CrossRef]
- Breast Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/breast-cancer (accessed on 1 September 2021).
- Kelsey, J.L.; Bernstein, L. Epidemiology and prevention of breast cancer. Annu. Rev. Public Health 1996, 17, 47–67. [Google Scholar] [CrossRef]
- Prostate Enlargement (Benign Prostatic Hyperplasia). Available online: https://www.niddk.nih.gov/health-information/urologic-diseases/prostate-problems/prostate-enlargement-benign-prostatic-hyperplasia (accessed on 1 May 2023).
- Galani, P. Diagnosis & prognosis of prostate cancer. J. Adv. Med. Dent. Sci. Res. 2015, 3, S49–S53. [Google Scholar]
- What Is Prostate Cancer? Available online: https://www.cancer.org/cancer/prostate-cancer/about/what-is-prostate-cancer.html (accessed on 1 May 2023).
- Santoso, K.T.; Menorca, A.; Cheung, C.Y.; Cook, G.M.; Stocker, B.L.; Timmer, M.S.M. The synthesis and evaluation of quinolinequinones as anti-mycobacterial agents. Bioorgan Med. Chem. 2019, 27, 3532–3545. [Google Scholar] [CrossRef]
- Ciftci, H.; Sever, B.; Kaya, N.; Bayrak, N.; Yildiz, M.; Yildirim, H.; Tateishi, H.; Otsuka, M.; Fujita, M.; TuYuN, A.F. Studies on 1,4-Quinone Derivatives Exhibiting Anti-Leukemic Activity along with Anti-Colorectal and Anti-Breast Cancer Effects. Molecules 2023, 28, 77. [Google Scholar] [CrossRef]
- Yilmaz Goler, A.M.; Tarbin Jannuzzi, A.; Biswas, A.; Mondal, S.; Basavanakatti, V.N.; Jayaprakash Venkatesan, R.; Yildirim, H.; Yildiz, M.; Celik Onar, H.; Bayrak, N.; et al. Analysis of Quinolinequinone Analogs with Promising Cytotoxic Activity against Breast Cancer. Chem. Biodivers. 2023, 20, e202300848. [Google Scholar] [CrossRef]
- Mataraci-Kara, E.; Bayrak, N.; Yildiz, M.; Yildirim, H.; TuYuN, A.F. Exploring the Relationships between Structure and Antimicrobial Potency of Quinolinequinones. Antibiotics 2022, 11, 1397. [Google Scholar] [CrossRef]
- Ricci, M.S.; Zong, W.-X. Chemotherapeutic Approaches for Targeting Cell Death Pathways. Oncologist 2006, 11, 342–357. [Google Scholar] [CrossRef]
- Razaghi, A.; Heimann, K.; Schaeffer, P.M.; Gibson, S.B. Negative regulators of cell death pathways in cancer: Perspective on biomarkers and targeted therapies. Apoptosis 2018, 23, 93–112. [Google Scholar] [CrossRef]
- Boyd, M.R.; Pauli, K.D. Some Practical Considerations and Applications of the National-Cancer-Institute in-Vitro Anticancer Drug Discovery Screen. Drug Dev. Res. 1995, 34, 91–109. [Google Scholar] [CrossRef]
- Monks, A.; Scudiero, D.; Skehan, P.; Shoemaker, R.; Paull, K.; Vistica, D.; Hose, C.; Langley, J.; Cronise, P.; Vaigrowolff, A.; et al. Feasibility of a High-Flux Anticancer Drug Screen Using a Diverse Panel of Cultured Human Tumor-Cell Lines. J. Natl. Cancer I 1991, 83, 757–766. [Google Scholar] [CrossRef]
- Grever, M.R.; Schepartz, S.A.; Chabner, B.A. The National-Cancer-Institute—Cancer Drug Discovery and Development Program. Semin. Oncol. 1992, 19, 622–638. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Suarez-Arnedo, A.; Torres Figueroa, F.; Clavijo, C.; Arbelaez, P.; Cruz, J.C.; Munoz-Camargo, C. An image J plugin for the high throughput image analysis of in vitro scratch wound healing assays. PLoS ONE 2020, 15, e0232565. [Google Scholar] [CrossRef]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS Using Oxidized DCFDA and Flow-Cytometry. Adv. Protoc. Oxidative Stress II 2010, 594, 57–72. [Google Scholar]
- Rakshit, G.; Jayaprakash, V. Tuberculosis and HIV responses threatened by nCOVID-19: A situation prompting an in silico investigation of reported MbtA inhibitors for combined inhibition of SARS-CoV-2 and HIV-TB co-infection. Struct. Chem. 2023, 34, 655–679. [Google Scholar] [CrossRef]
- Shilkar, D.; Mohd Siddique, M.U.; Bua, S.; Yasmin, S.; Patil, M.; Timiri, A.K.; Supuran, C.T.; Jayaprakash, V. Carbonic anhydrase inhibitory activity of phthalimide-capped benzene sulphonamide derivatives. J. Enzym. Inhib. Med. Chem. 2023, 38, 2235089. [Google Scholar] [CrossRef]
- Rakshit, G.; Biswas, A.; Jayaprakash, V. In Silico Drug Repurposing Studies for the Discovery of Novel Salicyl-AMP Ligase (MbtA)Inhibitors. Pathogens 2023, 12, 1433. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Santos-Martins, D.; Solis-Vasquez, L.; Tillack, A.F.; Sanner, M.F.; Koch, A.; Forli, S. Accelerating AutoDock4 with GPUs and Gradient-Based Local Search. J. Chem. Theory Comput. 2021, 17, 1060–1073. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple Ligand–Protein Interaction Diagrams for Drug Discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]
- Bayrak, N. A new family of azanaphthoquinones for antimicrobial evaluation. Chem. Cent. J. 2018, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, L.; Donato, M.T.; Gómez-Lechón, M.J. General Cytotoxicity Assessment by Means of the MTT Assay. Methods Mol. Biol. 2015, 1250, 333–348. [Google Scholar] [PubMed]
- Kadela, M.; Jastrzebska, M.; Bebenek, E.; Chrobak, E.; Latocha, M.; Kusz, J.; Ksiazek, M.; Boryczka, S. Synthesis, Structure and Cytotoxic Activity of Mono- and Dialkoxy Derivatives of 5,8-Quinolinedione. Molecules 2016, 21, 156. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.P.; Thatikonda, S.; Tokala, R.; Kumari, S.S.; Lakshmi, U.J.; Godugu, C.; Shankaraiah, N.; Kamal, A. Sulfamic acid promoted one-pot synthesis of phenanthrene fused-dihydrodibenzo-quinolinones: Anticancer activity, tubulin polymerization inhibition and apoptosis inducing studies. Bioorgan. Med. Chem. 2018, 26, 1996–2008. [Google Scholar] [CrossRef] [PubMed]
- Kuang, W.-B.; Huang, R.-Z.; Qin, J.-L.; Lu, X.; Qin, Q.-P.; Zou, B.-Q.; Chen, Z.-F.; Liang, H.; Zhang, Y. Design, synthesis and pharmacological evaluation of new 3-(1H-benzimidazol-2-yl)quinolin-2(1H)-one derivatives as potential antitumor agents. Eur. J. Med. Chem. 2018, 157, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Gulappa, T.; Reddy, R.S.; Suman, S.; Nyakeriga, A.M.; Damodaran, C. Molecular interplay between cdk4 and p21 dictates G0/G1 cell cycle arrest in prostate cancer cells. Cancer Lett. 2013, 337, 177–183. [Google Scholar] [CrossRef]
- Kim, G.-J.; Jo, H.-J.; Lee, K.-J.; Choi, J.W.; An, J.H. Oleanolic acid induces p53-dependent apoptosis via the ERK/JNK/AKT pathway in cancer cell lines in prostatic cancer xenografts in mice. Oncotarget 2018, 9, 26370–26386. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Cell Lines | AQQ1 | AQQ2 | AQQ3 | AQQ4 | AQQ5 | |
|---|---|---|---|---|---|---|
| Leukemia | ||||||
| GP | CCRF-CEM | −4.94 | −6.33 | 0.63 | 15.29 | 2.72 |
| HL-60(TB) | −48.05 | −36.75 | −46.39 | 9.95 | −13.44 | |
| K-562 | −21.12 | 1.26 | −4.17 | 7.18 | 4.73 | |
| MOLT-4 | −39.00 | −24.89 | −46.73 | 69.83 | 8.29 | |
| RPMI-8226 | −30.44 | −35.12 | −19.35 | 47.33 | −39.66 | |
| SR | −27.92 | −12.75 | −27.42 | 0.92 | −3.19 | |
| Non-Small Cell Lung Cancer | ||||||
| GP | A549/ATCC | 101.36 | 59.11 | 46.18 | 107.06 | 42.41 |
| EKVX | 28.52 | −2.24 | −97.48 | 50.81 | 74.66 | |
| HOP-62 | 74.12 | 84.25 | 72.15 | 110.03 | 0.85 | |
| HOP-92 | −83.81 | −89.25 | −86.99 | 112.07 | −12.23 | |
| NCI-H226 | 29.65 | 17.62 | −52.55 | 97.11 | 68.59 | |
| NCI-H23 | −8.37 | −67.81 | −61.00 | 59.03 | −55.61 | |
| NCI-H322M | 71.14 | 28.84 | 32.37 | 99.05 | 89.75 | |
| NCI-H460 | 70.83 | 12.07 | 13.41 | 100.46 | 2.02 | |
| NCI-H522 | −48.50 | −43.48 | −61.57 | 78.84 | −80.61 | |
| Colon Cancer | ||||||
| GP | COLO 205 | 111.34 | 99.24 | 98.22 | 107.70 | −80.52 |
| HCC-2998 | 58.90 | 20.35 | −55.70 | 103.69 | 14.44 | |
| HCT-116 | −80.64 | −79.44 | −83.64 | 55.84 | −97.84 | |
| HCT-15 | −47.03 | −76.56 | −83.19 | 82.29 | −66.21 | |
| HT29 | 75.04 | 19.96 | 15.90 | 109.56 | 24.91 | |
| KM12 | 69.40 | 60.77 | 57.40 | 98.23 | 39.30 | |
| SW-620 | −63.13 | −66.65 | −55.33 | 72.23 | −90.16 | |
| CNS Cancer | ||||||
| GP | SF-268 | 47.42 | 32.75 | 32.33 | 84.69 | 37.00 |
| SF-295 | 96.61 | 90.72 | 51.05 | 103.62 | 51.57 | |
| SF-539 | 1.78 | −37.33 | −58.82 | 103.46 | −30.92 | |
| SNB-19 | 38.92 | 46.47 | 44.68 | 94.62 | 30.42 | |
| SNB-75 | 105.38 | 101.38 | 96.54 | 110.92 | −4.09 | |
| U251 | 62.79 | 37.89 | 15.60 | 105.80 | 36.76 | |
| Melanoma | ||||||
| GP | LOX IMVI | −72.99 | −82.20 | −76.37 | −3.09 | −82.62 |
| MALME-3M | −55.92 | −71.39 | −90.58 | 94.54 | −89.37 | |
| M14 | ND a | ND a | ND a | ND a | −95.99 | |
| MDA-MB-435 | −85.96 | −85.72 | −83.74 | −67.16 | −87.43 | |
| SK-MEL-2 | 78.82 | −15.72 | −49.77 | 100.13 | 5.03 | |
| SK-MEL-28 | 14.13 | 14.55 | 8.93 | 101.65 | −73.06 | |
| SK-MEL-5 | −93.34 | −97.76 | −98.64 | 86.57 | −74.93 | |
| UACC-257 | −82.67 | −88.33 | −89.19 | 91.03 | −92.05 | |
| UACC-62 | −45.84 | −29.14 | −43.31 | 71.83 | −43.56 | |
| Ovarian Cancer | ||||||
| GP | IGROV1 | −83.51 | −57.52 | −82.43 | 19.63 | −23.13 |
| OVCAR-3 | CN b | CN b | CN b | 13.16 | CN b | |
| OVCAR-4 | −98.22 | −91.97 | −97.52 | 71.38 | −93.28 | |
| OVCAR-5 | 78.91 | 40.78 | 54.83 | 104.66 | 22.00 | |
| OVCAR-8 | 9.87 | 0.93 | 1.03 | 92.63 | −0.82 | |
| NCI/ADR-RES | 7.08 | −9.58 | −15.63 | 94.66 | −1.26 | |
| SK-OV-3 | 88.34 | 85.34 | 79.78 | 91.82 | 93.82 | |
| Renal Cancer | ||||||
| GP | 786-0 | 9.17 | −88.18 | 8.39 | 98.99 | −59.47 |
| A498 | 71.81 | 78.84 | 100.96 | 82.55 | 71.10 | |
| ACHN | −99.93 | −99.15 | −98.42 | 95.00 | −99.62 | |
| CAKI-1 | ND a | ND a | ND a | ND a | −96.53 | |
| RXF 393 | ND a | ND a | ND a | ND a | −97.24 | |
| SN12C | −49.46 | −50.97 | −57.33 | 90.46 | −63.27 | |
| TK-10 | 147.78 | 124.08 | 121.08 | 148.82 | −50.07 | |
| UO-31 | −65.68 | −92.96 | −65.05 | 100.30 | −94.65 | |
| Prostate Cancer | ||||||
| GP | PC-3 | 27.39 | 4.34 | 10.40 | 70.56 | 27.99 |
| DU-145 | −57.26 | −85.02 | −92.73 | 93.00 | −92.99 | |
| Breast Cancer | ||||||
| GP | MCF7 | −60.38 | −59.68 | −66.97 | 59.00 | −51.73 |
| MDA-MB-231/ATCC | −57.30 | −63.63 | −71.74 | 72.96 | −28.56 | |
| HS 578T | 72.15 | 1.05 | 48.97 | 118.57 | −32.78 | |
| BT-549 | 83.54 | 44.40 | 46.48 | 126.22 | 24.11 | |
| T-47D | −44.73 | −40.00 | −43.85 | 14.93 | −56.01 | |
| MDA-MB-468 | −79.74 | −90.72 | −95.15 | 58.11 | −78.49 | |
| Molecule | AQQ1 (NCI: D-829432/1) | AQQ2 (NCI: D-829433/1) | AQQ3 (NCI: D-829431/1) | AQQ5 (NCI: D-830803/1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Panel/Cell Line | GI50 | TGI | LC50 | GI50 | TGI | LC50 | GI50 | TGI | LC50 | GI50 | TGI | LC50 |
| Leukemia | ||||||||||||
| CCRF-CEM | 1.94 | 8.21 | >100 | 1.41 | >100 | >100 | 2.11 | >100 | >100 | 2.23 | ND a | >100 |
| HL-60(TB) | 1.37 | 3.52 | 9.03 | 1.03 | 2.92 | 8.29 | 1.47 | 4.18 | >100 | 1.38 | 4.65 | >100 |
| K-562 | 0.290 | 2.94 | >100 | 0.341 | ND a | >100 | 1.17 | >100 | >100 | 0.381 | >100 | >100 |
| MOLT-4 | 2.27 | 8.60 | >100 | 2.03 | 8.09 | >100 | 2.49 | >100 | >100 | 1.50 | 7.00 | >100 |
| RPMI-8226 | 1.57 | 4.61 | >100 | 1.20 | 4.71 | >100 | 1.67 | 5.33 | >100 | 1.79 | 5.19 | >100 |
| SR | 0.794 | 4.25 | >100 | 0.543 | 4.14 | >100 | 1.09 | >100 | >100 | 0.350 | 2.43 | >100 |
| Non-Small Cell Lung Cancer | ||||||||||||
| A549/ATCC | 2.44 | 6.24 | 45.2 | 2.06 | 4.64 | 12.9 | 2.09 | 4.64 | 17.5 | 2.83 | 8.97 | 38.8 |
| EKVX | 1.61 | 3.05 | 5.78 | 1.56 | 2.98 | 5.70 | 1.61 | 3.12 | 6.05 | 1.60 | 3.12 | 6.08 |
| HOP-62 | 2.41 | 6.98 | 37.5 | 2.27 | 6.74 | 28.8 | 1.74 | 3.23 | 6.01 | 4.26 | 14.1 | 37.7 |
| HOP-92 | 1.85 | 3.30 | 5.91 | 1.80 | 3.33 | 6.13 | 2.01 | 3.51 | 6.12 | 1.73 | 3.17 | 5.83 |
| NCI-H226 | 1.93 | 4.52 | 14.1 | 1.78 | 4.24 | 16.2 | 1.84 | 4.52 | >100 | 2.82 | 8.35 | 46.1 |
| NCI-H23 | 1.47 | 3.30 | 7.45 | 1.44 | 3.55 | 8.71 | 1.35 | 2.85 | 6.03 | 1.53 | 3.15 | 6.49 |
| NCI-H322M | 2.50 | 7.69 | 27.6 | 2.22 | 7.91 | 29.7 | 1.68 | 3.74 | 8.29 | 3.42 | 12.8 | 35.8 |
| NCI-H460 | 3.22 | 12 | 52.7 | 3.02 | 11.4 | 45.4 | 3.08 | 10.4 | 49.7 | 2.68 | 8.07 | 82.6 |
| NCI-H522 | 1.81 | 3.36 | 6.24 | 1.74 | 3.28 | 6.18 | 1.75 | 3.22 | 5.94 | 1.56 | 3.06 | 6.02 |
| Colon Cancer | ||||||||||||
| COLO 205 | 1.78 | 3.87 | ND a | 1.73 | 3.66 | 7.74 | 1.86 | 3.64 | 7.12 | 1.68 | 3.08 | 5.63 |
| HCC-2998 | 1.87 | 3.46 | 6.42 | 1.94 | 3.52 | 6.39 | 1.76 | 3.25 | 6.00 | 1.81 | 3.26 | 5.88 |
| HCT-116 | 1.53 | 4.16 | 44.3 | 1.05 | 2.31 | 5.05 | 1.32 | 2.79 | 5.92 | 1.53 | 2.88 | 5.45 |
| HCT-15 | 1.33 | 3.02 | 6.83 | 1.13 | 2.55 | 5.76 | 1.30 | 2.94 | 6.66 | 1.54 | 3.92 | ND a |
| HT29 | 2.13 | 4.49 | 9.46 | 3.40 | 15.2 | 99.5 | 2.87 | 7.08 | 62.5 | 1.87 | 3.48 | 6.48 |
| KM12 | 1.49 | 3.08 | 6.40 | 1.41 | 2.89 | 5.92 | 1.44 | 3.07 | 6.51 | 13.0 | 26.6 | 54.7 |
| SW-620 | 1.84 | 3.73 | 7.56 | 1.60 | 3.21 | 6.41 | 1.83 | 3.51 | 6.73 | 1.76 | 3.47 | 6.85 |
| CNS Cancer | ||||||||||||
| SF-268 | 1.53 | 3.05 | 6.06 | 1.47 | 2.94 | 5.87 | 1.55 | 3.09 | 6.14 | 3.25 | 14.4 | 72.7 |
| SF-295 | 1.85 | 3.42 | 6.32 | 2.03 | 4.20 | 8.70 | 1.86 | 3.43 | 6.30 | 1.99 | 5.41 | 20.8 |
| SF-539 | 1.68 | 3.13 | 5.81 | 1.67 | 3.09 | 5.71 | 1.68 | 3.13 | 5.84 | 1.79 | 3.22 | 5.78 |
| SNB-19 | 1.81 | 3.72 | 7.64 | 1.80 | 3.97 | 8.73 | 1.68 | 3.16 | 5.94 | 5.82 | 18.8 | 44.2 |
| SNB-75 | 1.50 | 2.84 | 5.39 | 1.63 | 3.43 | 7.21 | 1.46 | 2.79 | 5.33 | 1.94 | 5.37 | 23.2 |
| U251 | 1.85 | 3.73 | 7.53 | 1.70 | 3.27 | 6.28 | 1.80 | 3.54 | 6.94 | 1.92 | 4.44 | 10.8 |
| Melanoma | ||||||||||||
| LOX IMVI | 0.952 | 2.36 | 5.70 | 0.839 | 2.22 | 5.33 | 1.06 | 2.33 | 5.14 | 1.10 | 2.32 | 4.88 |
| MALME-3M | 1.71 | 3.19 | 5.95 | 1.36 | 2.77 | 5.64 | 1.65 | 3.16 | 6.02 | 1.76 | 3.28 | 6.11 |
| M14 | 1.82 | 3.81 | 7.96 | 1.46 | 2.83 | 5.50 | 1.68 | 3.23 | 6.21 | 1.69 | 3.09 | 5.66 |
| MDA-MB-435 | 1.70 | 3.11 | 5.67 | 0.266 | 0.671 | 2.45 | 0.442 | 1.53 | 3.99 | 1.72 | 3.57 | 7.39 |
| SK-MEL-2 | 1.81 | 3.34 | 6.19 | 1.44 | 2.86 | 5.69 | 1.63 | 3.03 | 5.64 | 1.59 | 3.10 | 6.03 |
| SK-MEL-28 | 1.85 | 3.34 | 6.02 | 1.71 | 3.14 | 5.79 | 1.73 | 3.14 | 5.70 | 1.79 | 3.23 | 5.85 |
| SK-MEL-5 | 1.75 | 3.17 | 5.74 | 1.74 | 3.13 | 5.65 | 1.72 | 3.11 | 5.62 | 1.62 | 2.97 | 5.47 |
| UACC-257 | 1.57 | 3.09 | 6.11 | 1.28 | 2.61 | 5.34 | 1.39 | 2.78 | 5.58 | 1.51 | 2.85 | 5.37 |
| UACC-62 | 1.82 | 3.37 | 6.25 | 1.80 | 3.36 | 6.28 | 1.81 | 3.29 | 5.97 | 1.77 | 3.42 | 6.60 |
| Ovarian Cancer | ||||||||||||
| IGROV1 | 1.46 | 2.92 | 5.86 | 1.37 | 2.82 | 5.84 | 1.36 | 2.76 | 5.61 | 1.73 | 3.55 | 7.26 |
| OVCAR-3 | 1.12 | 2.40 | 5.15 | 0.759 | 2.03 | 4.54 | 0.962 | 2.19 | 4.87 | ND a | ND a | ND a |
| OVCAR-4 | 1.21 | 2.49 | 5.11 | 1.00 | 2.22 | 4.91 | 1.13 | 2.37 | 4.97 | 1.40 | 2.82 | 5.66 |
| OVCAR-5 | 1.86 | 3.42 | 6.29 | 1.84 | 3.33 | 6.03 | 1.92 | 3.46 | 6.25 | 2.38 | 5.79 | 23.7 |
| OVCAR-8 | 1.61 | 3.25 | 6.59 | 1.49 | 2.92 | 5.70 | 1.57 | 3.01 | 5.77 | 1.64 | 3.28 | 6.56 |
| NCI/ADR-RES | 2.03 | 5.16 | 34.6 | 1.89 | 5.09 | >100 | 1.70 | 4.11 | 9.93 | 1.81 | 4.47 | 21.7 |
| SK-OV-3 | 12.6 | 27.5 | 59.8 | 13.5 | 27.1 | 54.2 | 11.0 | 30.1 | 82.3 | 12.6 | 25.1 | 50.1 |
| Renal Cancer | ||||||||||||
| 786-0 | 1.91 | 3.61 | 6.82 | 1.80 | 3.25 | 5.86 | 1.72 | 3.19 | 5.89 | 1.52 | 2.88 | 5.44 |
| A498 | 1.90 | 4.60 | 13.1 | 2.65 | 10.8 | 34.8 | 1.56 | 5.52 | 24.4 | 2.35 | 11.4 | 33.8 |
| ACHN | 1.47 | 2.80 | 5.34 | 1.35 | 2.69 | 5.35 | 1.57 | 2.92 | 5.45 | 1.67 | 3.04 | 5.51 |
| CAKI-1 | 1.50 | 2.87 | 5.47 | 1.45 | 2.84 | 5.57 | 1.60 | 2.97 | 5.51 | 1.76 | 3.15 | 5.65 |
| RXF 393 | 1.42 | 2.76 | 5.36 | 1.44 | 2.78 | 5.39 | 1.57 | 2.96 | 5.61 | 1.63 | 2.99 | 5.48 |
| SN12C | 1.70 | 3.29 | 6.36 | 1.59 | 3.04 | 5.80 | 1.58 | 2.99 | 5.66 | 1.71 | 3.25 | 6.19 |
| TK-10 | 2.34 | 3.86 | 6.36 | 2.25 | 3.72 | 6.18 | 2.30 | 3.77 | 6.18 | 2.32 | 3.78 | 6.15 |
| UO-31 | 1.42 | 2.73 | 5.26 | 1.40 | 2.71 | 5.23 | 1.43 | 2.75 | 5.28 | 1.69 | 3.06 | 5.53 |
| Prostate Cancer | ||||||||||||
| PC-3 | 1.21 | 2.58 | 5.49 | 1.26 | 2.77 | 6.11 | 1.29 | 2.77 | 5.93 | 2.56 | 10.4 | 43.6 |
| DU-145 | 1.72 | 3.11 | 5.62 | 1.63 | 2.99 | 5.49 | 1.72 | 3.13 | 5.68 | 1.84 | 3.55 | 6.83 |
| Breast Cancer | ||||||||||||
| MCF7 | 1.78 | 4.49 | >100 | 1.33 | 3.65 | ND a | 1.54 | 3.82 | 9.44 | 1.82 | 5.33 | >100 |
| MDA-MB-231/ATCC | 1.19 | 2.53 | 5.37 | 1.07 | 2.38 | 5.31 | 1.17 | 2.50 | 5.34 | 1.46 | 3.08 | 6.48 |
| HS 578T | 2.27 | 5.64 | >100 | 2.11 | 5.25 | >100 | 2.22 | 5.82 | >100 | 2.43 | 5.78 | >100 |
| BT-549 | 1.72 | 3.27 | 6.19 | 1.52 | 2.88 | 5.46 | 1.43 | 2.80 | 5.49 | 1.97 | 3.40 | 5.87 |
| T-47D | 1.37 | ND a | >100 | 0.467 | 3.17 | >100 | 1.01 | 3.91 | >100 | 1.50 | 3.62 | 8.74 |
| MDA-MB-468 | 1.52 | 3.06 | 6.16 | 1.45 | 2.88 | 5.71 | 1.49 | 2.92 | 5.75 | 1.23 | 2.57 | 5.37 |
| (µM) | DU-145 | MDA-MB-231 | HCT-116 | HUVEC | |
|---|---|---|---|---|---|
| AQQ1 | IC50 | 4.18 ± 0.73 | 8.27 ± 0.91 | 5.83 ± 0.76 | 5.39 ± 0.73 |
| AQQ2 | IC50 | 4.17 ± 0.80 | 13.33 ± 1.12 | 9.18 ± 0.96 | 6.39 ± 0.80 |
| DOXO | IC50 | <100 | 59.06 ± 9.06 | 24.35 ± 6.90 | 57.48 ± 9.82 |
| Source | Parameters | AQQ1 | AQQ2 | Verapamil | Atenolol |
|---|---|---|---|---|---|
| Log P | 1.85 | 1.77 | −0.21 | −0.33 | |
| Log D | 2.37 | 2.35 | 1.87 | −1.77 | |
| Mouse liver microsomes | % Metabolism in 30 min | 63 | 96.5 | 80.0 | |
| Half-life (min) | 20.5 | 5.5 | 11.0 | ||
| CLint (L/min/mg protein) | 68.5 | 249 | 130.0 | ||
| Rat liver microsomes | % Metabolism in 30 min | 100 | 100 | 79.0 | |
| Half-life (min) | 2 | 3 | 11.0 | ||
| CLint (L/min/mg protein) | 655.5 | 501 | 121.0 | ||
| Dog liver microsomes | % Metabolism in 30 min | 41.5 | 79.5 | 81.0 | |
| Half-life (min) | 31 | 17 | 11.0 | ||
| CLint (L/min/mg protein) | 45 | 84 | 131.0 | ||
| Human liver microsomes | % Metabolism in 30 min | 39.5 | 47 | 79.0 | |
| Half-life (min) | >30 | 26.5 | 11.0 | ||
| CLint (L/min/mg protein) | 39 | 52 | 129.0 |
| Parameters | AQQ1 | AQQ2 | ||
|---|---|---|---|---|
| IV | PO | IV | PO | |
| C0 (IV only)/Cmax (PO only) (ng/mL) | 90.8 ± 71.3 | 435 ± 18.1 | 23.2 ± 12.8 | 282 ± 79.4 |
| Tmax (h) | NA | 0.5 | NA | 1 |
| AUClast (h × ng/mL) | 77.5 ± 9.81 | 3810 ± 111 | 77 ± 21 | 2300 ± 416 |
| AUCINF (h × ng/mL) | 115 ± 45.7 | 4000 ± 179 | 110 ± 18.1 | 2370 ± 375 |
| T1/2 (h) | 9.35 ± 8.75 | 5.22 ± 0.843 | 12.8 ± 2.95 | 4.72 ± 1.16 |
| Vd (L/kg) | 90.3 ± 40 | NC b | 147 ± 9.71 | NC b |
| Cl (mL/min/kg) | 161 ± 62.9 | NC b | 154 ± 26.9 | NC b |
| MRTlast (h) | 3.77 ± 0.279 | 7.47 ± 0.367 | 6.84 ± 1.84 | 6.61 ± 0.787 |
| F% | NA a | 9.83 | NAa | 5.96 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jannuzzi, A.T.; Yilmaz Goler, A.M.; Biswas, A.; Mondal, S.; Basavanakatti, V.N.; Yıldırım, H.; Yıldız, M.; Bayrak, N.; Jayaprakash, V.; TuYuN, A.F. Prospects for Prostate Cancer Chemotherapy: Cytotoxic Evaluation and Mechanistic Insights of Quinolinequinones with ADME/PK Profile. Biomedicines 2024, 12, 1241. https://doi.org/10.3390/biomedicines12061241
Jannuzzi AT, Yilmaz Goler AM, Biswas A, Mondal S, Basavanakatti VN, Yıldırım H, Yıldız M, Bayrak N, Jayaprakash V, TuYuN AF. Prospects for Prostate Cancer Chemotherapy: Cytotoxic Evaluation and Mechanistic Insights of Quinolinequinones with ADME/PK Profile. Biomedicines. 2024; 12(6):1241. https://doi.org/10.3390/biomedicines12061241
Chicago/Turabian StyleJannuzzi, Ayse Tarbin, Ayse Mine Yilmaz Goler, Abanish Biswas, Subodh Mondal, Vinay N. Basavanakatti, Hatice Yıldırım, Mahmut Yıldız, Nilüfer Bayrak, Venkatesan Jayaprakash, and Amaç Fatih TuYuN. 2024. "Prospects for Prostate Cancer Chemotherapy: Cytotoxic Evaluation and Mechanistic Insights of Quinolinequinones with ADME/PK Profile" Biomedicines 12, no. 6: 1241. https://doi.org/10.3390/biomedicines12061241
APA StyleJannuzzi, A. T., Yilmaz Goler, A. M., Biswas, A., Mondal, S., Basavanakatti, V. N., Yıldırım, H., Yıldız, M., Bayrak, N., Jayaprakash, V., & TuYuN, A. F. (2024). Prospects for Prostate Cancer Chemotherapy: Cytotoxic Evaluation and Mechanistic Insights of Quinolinequinones with ADME/PK Profile. Biomedicines, 12(6), 1241. https://doi.org/10.3390/biomedicines12061241