
Using the Traditional Ex Vivo Whole Blood Model to Discriminate Bacteria by Their Inducible Host Responses
 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Whole Blood Infection
2.3. Membrane Cytokine Array
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Phagocytosis
2.6. Pairwise Discrimination of Bacteria
2.7. Linear Discriminant Analysis of Bacteria
2.8. Data Presentation and Statistical Analysis
3. Results
3.1. Establishment of Mediator Profiles to Use in Modelling Datasets
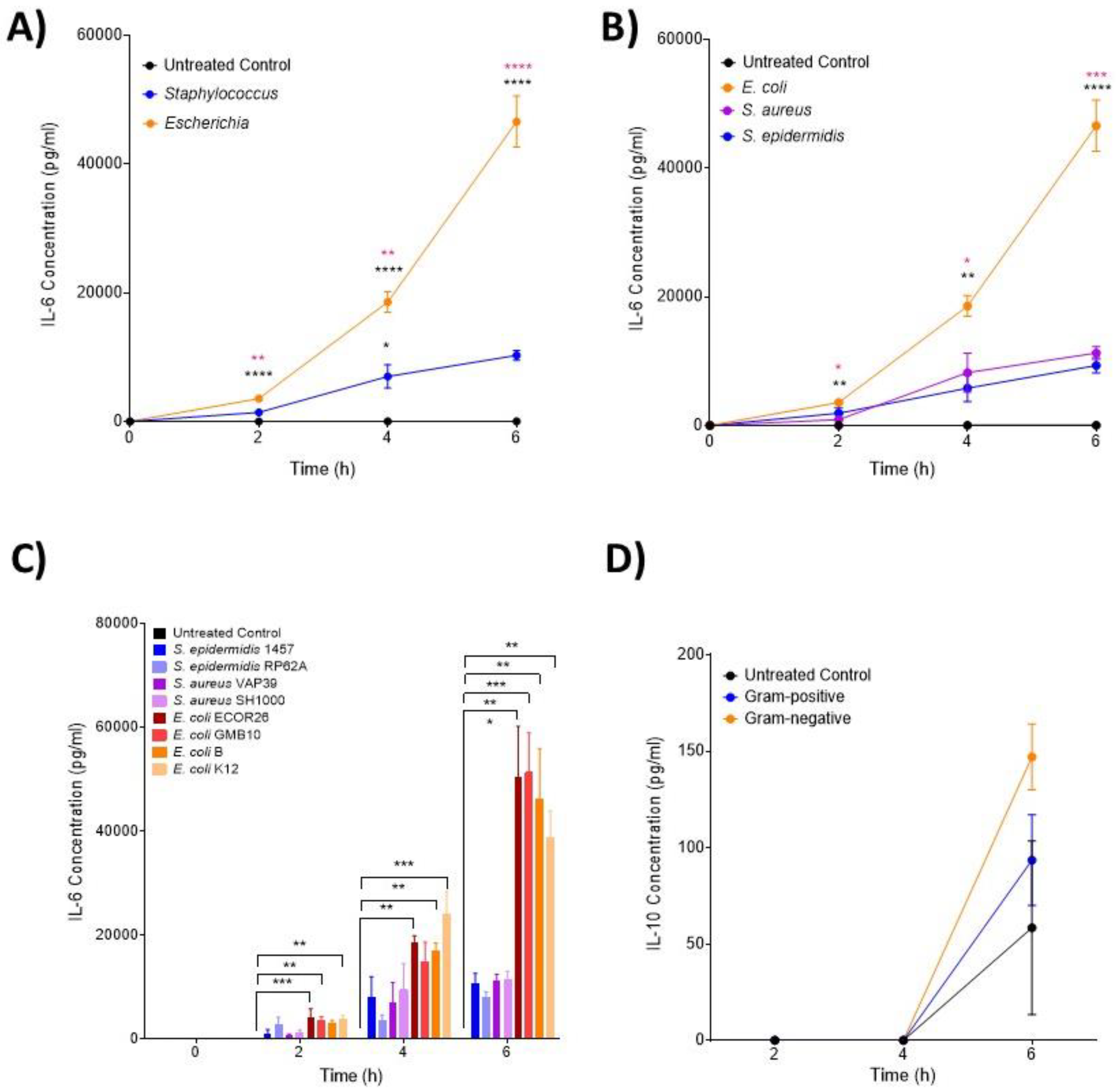
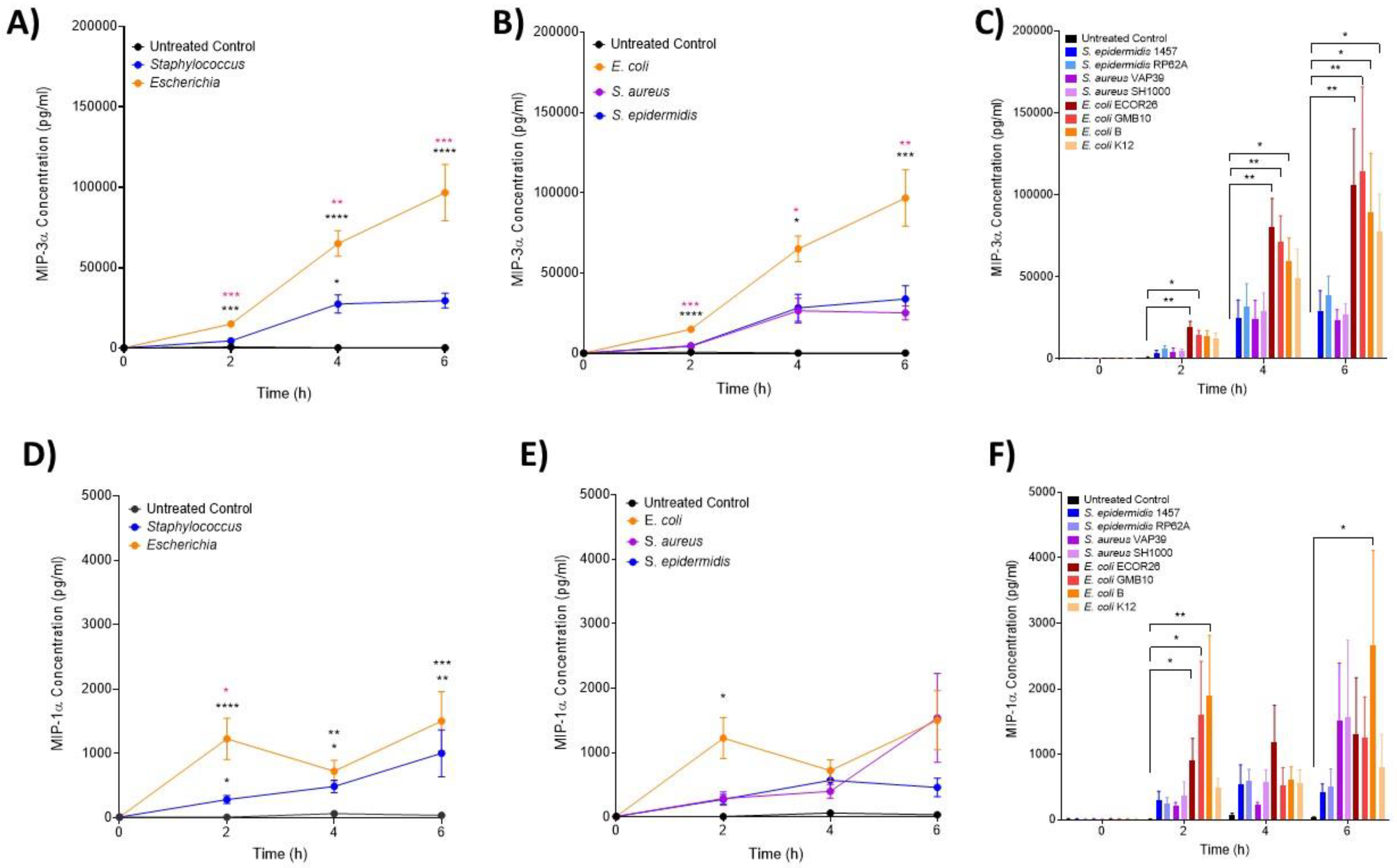
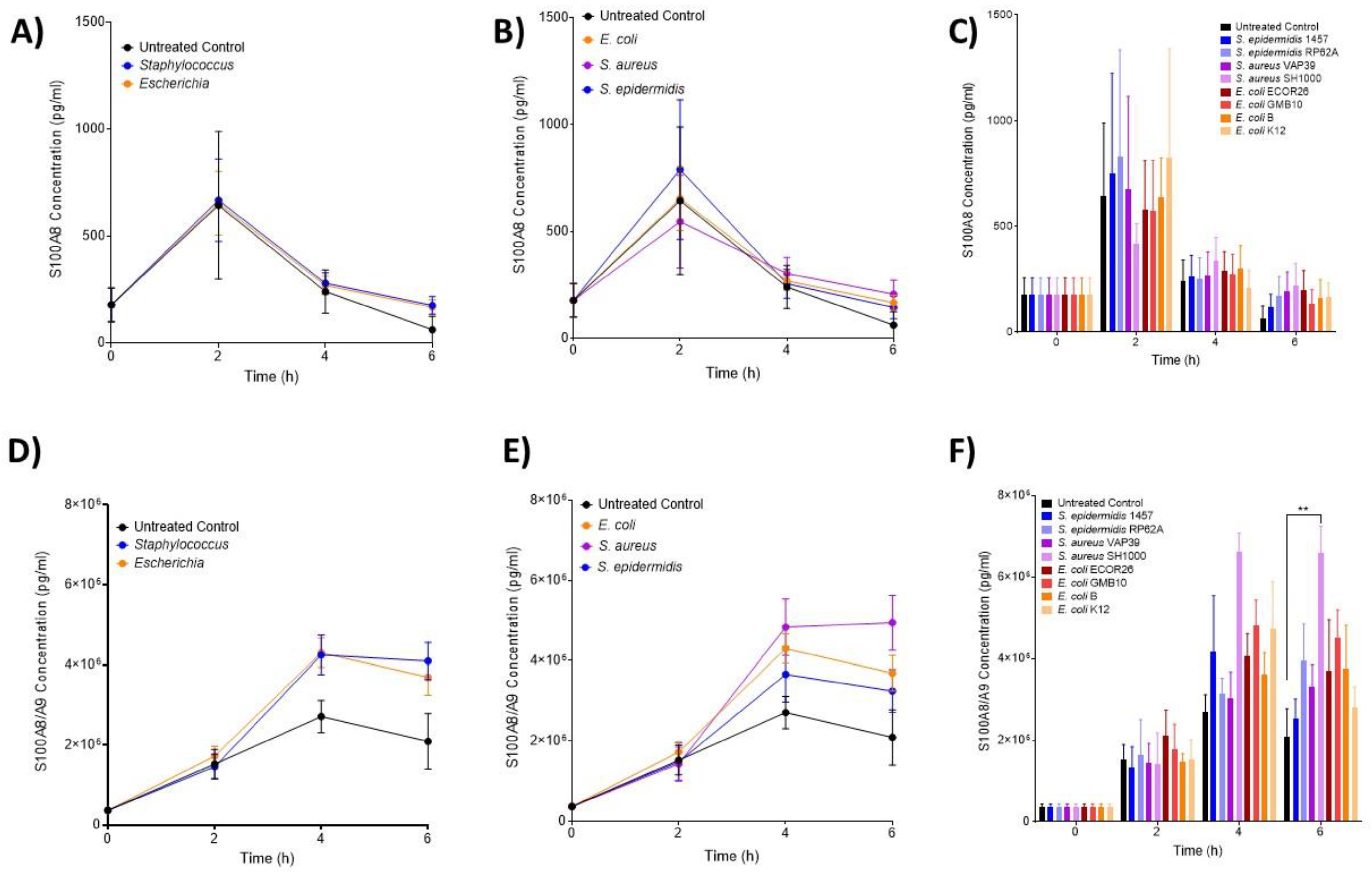
3.2. IL-6 and MIP Proteins Show Significant Early Induction in Escherichia Infection
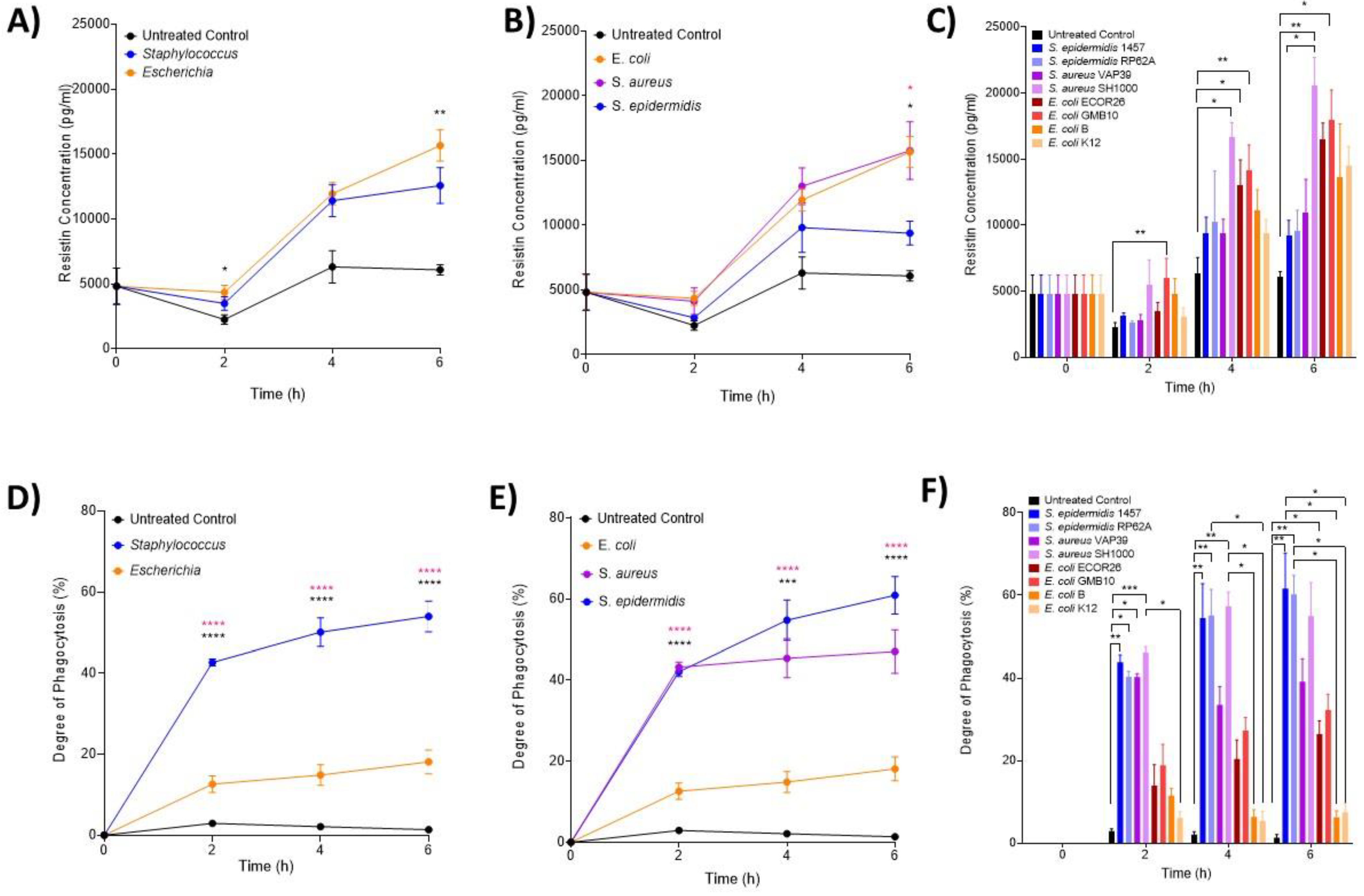
3.3. The Hormone Resistin Is Associated with All Infection and May Differentiate S. epidermidis and S. aureus Bacteria
3.4. Neutrophils Take up Significantly More Staphylococci Than Escherichia Bacteria
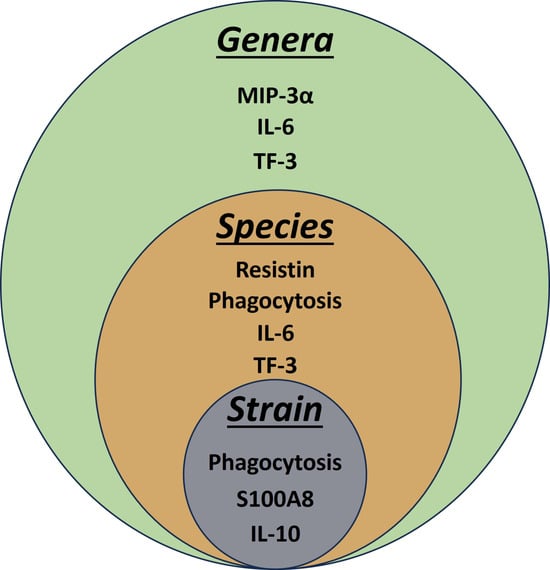
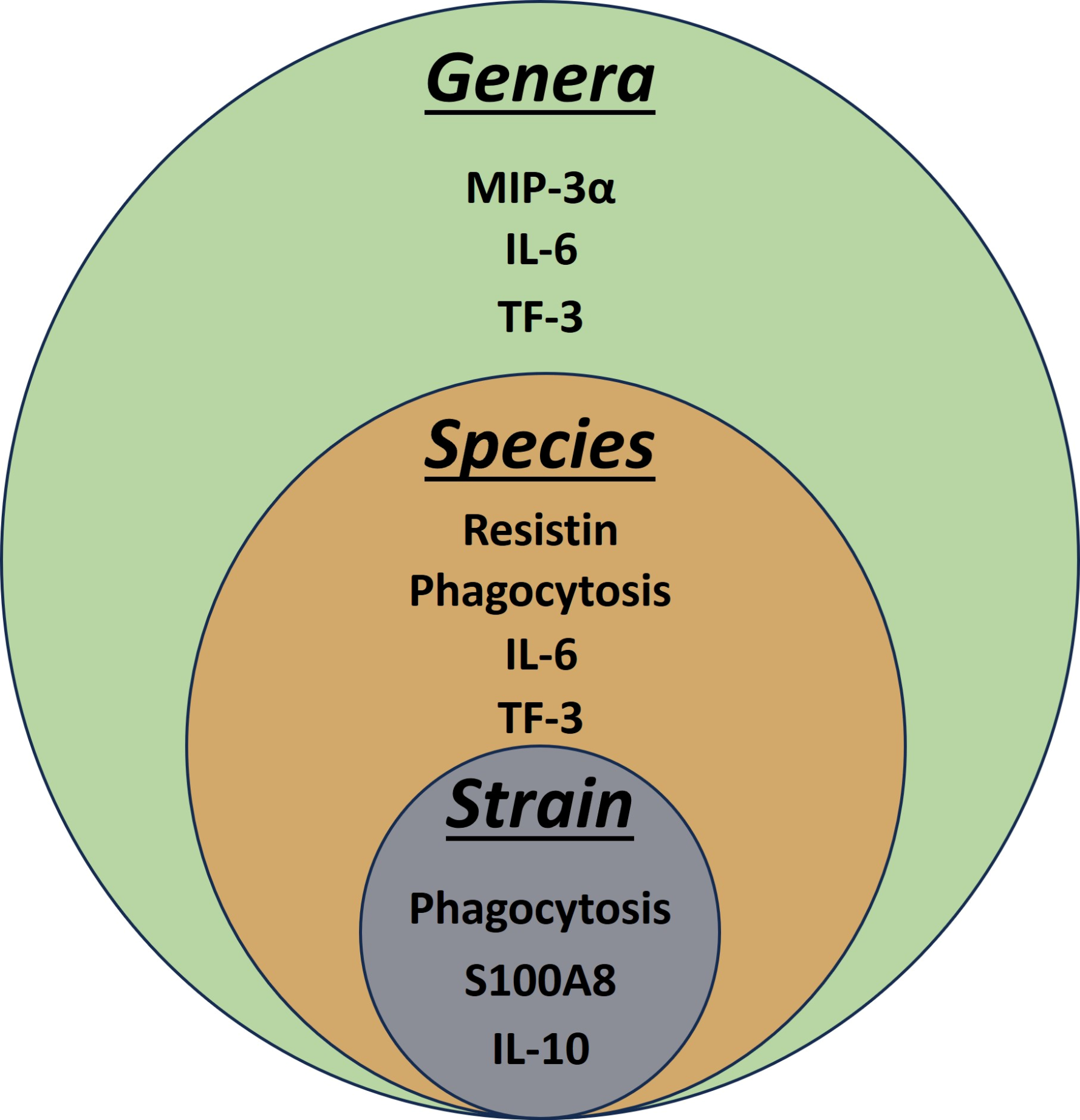
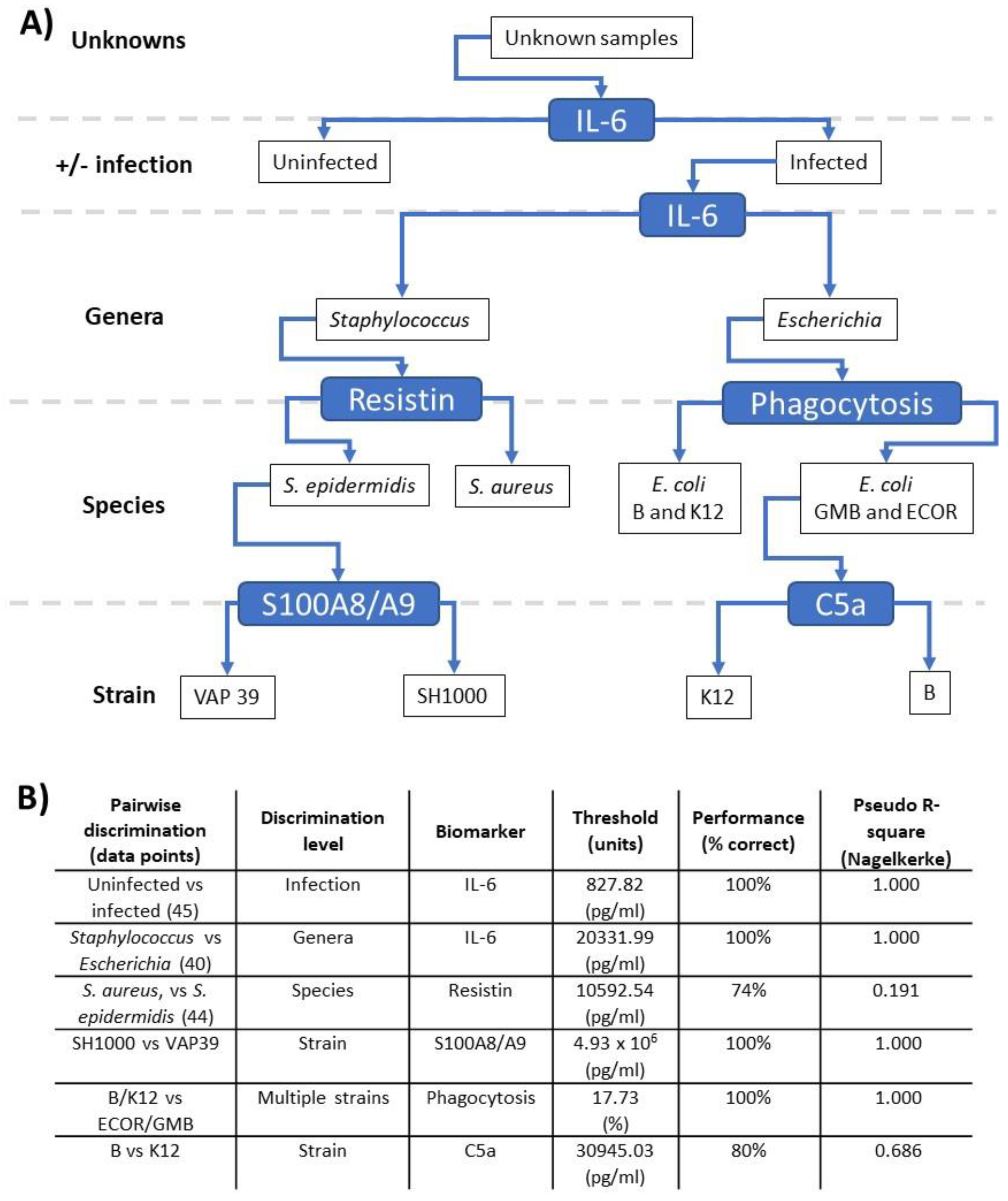
3.5. Pairwise Modelling Allows Bacterial Discrimination down to Strain Level
3.6. Linear Discriminant Analysis Provides Host Biomarker Panels for Bacterial Discrimination
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duffy, D.; Rouilly, V.; Libri, V.; Hasan, M.; Beitz, B.; David, M.; Urrutia, A.; Bisiaux, A.; Labrie, S.T.; Dubois, A.; et al. Functional analysis via standardized whole-blood stimulation systems defines the boundaries of a healthy immune response to complex stimuli. Immunity 2014, 40, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Liebers, V.; Kendzia, B.; Stubel, H.; Borowitzki, G.; Gering, V.; Monse, C.; Hagemeyer, O.; Merget, R.; Bruning, T.; Raulf, M. Cell Activation and Cytokine Release Ex Vivo: Estimation of Reproducibility of the Whole-Blood Assay with Fresh Human Blood. Adv. Exp. Med. Biol. 2018, 1108, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.; Ponte, C.G.; Hacker, M.A.; Antas, P.R. A whole blood assay as a simple, broad assessment of cytokines and chemokines to evaluate human immune responses to Mycobacterium tuberculosis antigens. Acta Trop. 2013, 127, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Spierenburg, E.A.J.; Portengen, L.; Smit, L.A.M.; Krop, E.J.M.; Hylkema, M.N.; Rijkers, G.T.; Heederik, D.; Wouters, I.M. Stability of individual LPS-induced ex vivo cytokine release in a whole blood assay over a five-year interval. J. Immunol. Methods 2018, 460, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, A.; Duffy, D.; Rouilly, V.; Posseme, C.; Djebali, R.; Illanes, G.; Libri, V.; Albaud, B.; Gentien, D.; Piasecka, B.; et al. Standardized Whole-Blood Transcriptional Profiling Enables the Deconvolution of Complex Induced Immune Responses. Cell Rep. 2016, 16, 2777–2791. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, S.B.; Emneus, J.; Wolff, A.; Jungersen, G. Revisiting the IFN-gamma release assay: Whole blood or PBMC cultures?—And other factors of influence. J. Immunol. Methods 2016, 434, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Liebers, V.; Stubel, H.; Duser, M.; Bruning, T.; Raulf-Heimsoth, M. Standardization of whole blood assay for determination of pyrogenic activity in organic dust samples. Int. J. Hyg. Environ. Health 2009, 212, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Damsgaard, C.T.; Lauritzen, L.; Calder, P.C.; Kjaer, T.M.; Frokiaer, H. Whole-blood culture is a valid low-cost method to measure monocytic cytokines—A comparison of cytokine production in cultures of human whole-blood, mononuclear cells and monocytes. J. Immunol. Methods 2009, 340, 95–101. [Google Scholar] [CrossRef]
- van Dooren, F.H.; Duijvis, N.W.; Velde, A.A.T. Analysis of cytokines and chemokines produced by whole blood, peripheral mononuclear and polymorphonuclear cells. J. Immunol. Methods 2013, 396, 128–133. [Google Scholar] [CrossRef]
- Weigandt, F.; Lexa, P.; Sonntag, H.G. A new test for the detection of pyrogens in pharmaceutical products. Examinations for the validation of the human whole blood assay. ALTEX 1998, 15, 13–17. [Google Scholar]
- Fennrich, S.; Fischer, M.; Hartung, T.; Lexa, P.; Montag-Lessing, T.; Sonntag, H.G.; Weigandt, M.; Wendel, A. Evaluation and further development of a pyrogenicity assay based on human whole blood. ALTEX 1998, 15, 123–128. [Google Scholar]
- Fischer, M.; Hartzsch, K.; Hartung, T.; Montag-Lessing, T. First Results in the prevaluation of the human whole blood assay for pyrogens in biological pharmaceuticals. ALTEX 1998, 15, 10–13. [Google Scholar]
- Pilat, C.; Kruger, K.; Frech, T.; Mooren, F.C. Exercise-induced cytokine changes in antigen stimulated whole-blood cultures compared to serum. J. Immunol. Methods 2017, 440, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Segre, E.; Fullerton, J.N. Stimulated Whole Blood Cytokine Release as a Biomarker of Immunosuppression in the Critically Ill: The Need for a Standardized Methodology. Shock 2016, 45, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Al-Ishaq, R.; Armstrong, J.; Gregory, M.; O’Hara, M.; Phiri, K.; Harris, L.G.; Rohde, H.; Siemssen, N.; Frommelt, L.; Mack, D.; et al. Effects of polysaccharide intercellular adhesin (PIA) in an ex vivo model of whole blood killing and in prosthetic joint infection (PJI): A role for C5a. Int. J. Med. Microbiol. 2015, 305, 948–956. [Google Scholar] [CrossRef]
- Opota, O.; Jaton, K.; Greub, G. Microbial diagnosis of bloodstream infection: Towards molecular diagnosis directly from blood. Clin. Microbiol. Infect. 2015, 21, 323–331. [Google Scholar] [CrossRef]
- Umemura, Y.; Ogura, H.; Takuma, K.; Fujishima, S.; Abe, T.; Kushimoto, S.; Hifumi, T.; Hagiwara, A.; Shiraishi, A.; Otomo, Y.; et al. Current spectrum of causative pathogens in sepsis: A prospective nationwide cohort study in Japan. Int. J. Infect. Dis. 2021, 103, 343–351. [Google Scholar] [CrossRef]
- Frendeus, B.; Wachtler, C.; Hedlund, M.; Fischer, H.; Samuelsson, P.; Svensson, M.; Svanborg, C. Escherichia coli P fimbriae utilize the Toll-like receptor 4 pathway for cell activation. Mol. Microbiol. 2001, 40, 37–51. [Google Scholar] [CrossRef]
- Lee, J.S.; Frevert, C.W.; Matute-Bello, G.; Wurfel, M.M.; Wong, V.A.; Lin, S.M.; Ruzinski, J.; Mongovin, S.; Goodman, R.B.; Martin, T. R TLR-4 pathway mediates the inflammatory response but not bacterial elimination in E. coli pneumonia. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 289, L731–L738. [Google Scholar] [CrossRef] [PubMed]
- Askarian, F.; Wagner, T.; Johannessen, M.; Nizet, V. Staphylococcus aureus modulation of innate immune responses through Toll-like (TLR), (NOD)-like (NLR) and C-type lectin (CLR) receptors. FEMS Microbiol. Rev. 2018, 42, 656–671. [Google Scholar] [CrossRef]
- Strunk, T.; Coombs, M.R.P.; Currie, A.J.; Richmond, P.; Golenbock, D.T.; Stoler-Barak, L.; Gallington, L.C.; Otto, M.; Burgner, D.; Levy, O. TLR2 mediates recognition of live Staphylococcus epidermidis and clearance of bacteremia. PLoS ONE 2010, 5, e10111. [Google Scholar] [CrossRef]
- Bi, D.; Qiao, L.; Bergelson, I.; Ek, C.J.; Duan, L.; Zhang, X.; Albertsson, A.M.; Pettengill, M.; Kronforst, K.; Ninkovic, J.; et al. Staphylococcus epidermidis Bacteremia Induces Brain Injury in Neonatal Mice via Toll-like Receptor 2-Dependent and -Independent Pathways. J. Infect. Dis. 2015, 212, 1480–1490. [Google Scholar] [CrossRef][Green Version]
- Conway-Morris, A.; Wilson, J.; Shankar-Hari, M. Immune Activation in Sepsis. Crit. Care Clin. 2018, 34, 29–42. [Google Scholar] [CrossRef]
- Samuel, L. Direct Detection of Pathogens in Bloodstream During Sepsis: Are We There Yet? J. Appl. Lab. Med. 2019, 3, 631–642. [Google Scholar] [CrossRef]
- Sweeney, T.E.; Liesenfeld, O.; May, L. Diagnosis of bacterial sepsis: Why are tests for bacteremia not sufficient? Expert Rev. Mol. Diagn. 2019, 19, 959–962. [Google Scholar] [CrossRef]
- Seymour, C.W.; Gesten, F.; Prescott, H.C.; Friedrich, M.E.; Iwashyna, T.J.; Phillips, G.S.; Lemeshow, S.; Osborn, T.; Terry, K.M.; Levy, M.M. Time to Treatment and Mortality during Mandated Emergency Care for Sepsis. N. Engl. J. Med. 2017, 376, 2235–2244. [Google Scholar] [CrossRef]
- Jin, M.; Khan, A.I. Procalcitonin: Uses in the clinical laboratory for the diagnosis of sepsis. Lab. Med. 2010, 41, 173–177. [Google Scholar] [CrossRef]
- Raveendran, A.V.; Kumar, A.; Gangadharan, S. Biomarkers and newer laboratory investigations in the diagnosis of sepsis. J. R. Coll. Physicians Edinb. 2019, 49, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Chalupa, P.; Beran, O.; Herwald, H.; Kasprikova, N.; Holub, M. Evaluation of potential biomarkers for the discrimination of bacterial and viral infections. Infection 2011, 39, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Huang, D.; Zeng, R.; Ye, Z.; Zhang, Y. Accuracy of serum interleukin (IL)-6 in sepsis diagnosis: A systematic review and meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 15238–15245. [Google Scholar] [PubMed]
- Vincent, J.L. The Clinical Challenge of Sepsis Identification and Monitoring. PLoS Med. 2016, 13, e1002022. [Google Scholar] [CrossRef]
- Gruys, E.; Toussaint, M.J.; Niewold, T.A.; Koopmans, S.J. Acute phase reaction and acute phase proteins. J. Zhejiang Univ. Sci. B 2005, 6, 1045–1056. [Google Scholar] [CrossRef]
- Schuetz, P.; Albrich, W.; Mueller, B. Procalcitonin for diagnosis of infection and guide to antibiotic decisions: Past, present and future. BMC Med. 2011, 9, 107. [Google Scholar] [CrossRef]
- Dolin, H.H.; Papadimos, T.J.; Chen, X.; Pan, Z.K. Characterization of Pathogenic Sepsis Etiologies and Patient Profiles: A Novel Approach to Triage and Treatment. Microbiol. Insights 2019, 12, 1178636118825081. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.; Nygard, S.; Fure, H.; Olstad, O.K.; Holden, M.; Lappegard, K.T.; Brekke, O.L.; Espevik, T.; Hovig, E.; Mollnes, T.E. CD14 and complement crosstalk and largely mediate the transcriptional response to Escherichia coli in human whole blood as revealed by DNA microarray. PLoS ONE 2015, 10, e0117261. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.; Olstad, O.K.; Holden, M.; Nygard, S.; Fure, H.; Lappegard, K.T.; Brekke, O.L.; Espevik, T.; Hovig, E.; Mollnes, T.E. Gene expression profiling of Gram-negative bacteria-induced inflammation in human whole blood: The role of complement and CD14-mediated innate immune response. Genom. Data 2015, 5, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Dix, A.; Hunniger, K.; Weber, M.; Guthke, R.; Kurzai, O.; Linde, J. Biomarker-based classification of bacterial and fungal whole-blood infections in a genome-wide expression study. Front. Microbiol. 2015, 6, 171. [Google Scholar] [CrossRef] [PubMed]
- Satria, R.D.; Jhan, M.K.; Chen, C.L.; Tseng, P.C.; Wang, Y.T.; Lin, C.F. Elevated TNF-alpha Induces Thrombophagocytosis by Mononuclear Cells in ex vivo Whole-Blood Co-Culture with Dengue Virus. J. Inflamm. Res. 2022, 15, 1717–1728. [Google Scholar] [CrossRef] [PubMed]
- Meric, G.; Kemsley, E.K.; Falush, D.; Saggers, E.J.; Lucchini, S. Phylogenetic distribution of traits associated with plant colonization in Escherichia coli. Environ. Microbiol. 2013, 15, 487–501. [Google Scholar] [CrossRef]
- Ochman, H.; Selander, R.K. Standard reference strains of Escherichia coli from natural populations. J. Bacteriol. 1984, 157, 690–693. [Google Scholar] [CrossRef]
- Bachmann, B.J. Pedigrees of some mutant strains of Escherichia coli K-12. Bacteriol. Rev. 1972, 36, 525–557. [Google Scholar] [CrossRef]
- Gray, C.H.; Tatum, E.L. X-Ray Induced Growth Factor Requirements in Bacteria. Proc. Natl. Acad. Sci. USA 1944, 30, 404–410. [Google Scholar] [CrossRef]
- Lederberg, J. E. coli K12. Microbiol. Today 2004, 31, 116. [Google Scholar]
- Daegelen, P.; Studier, F.W.; Lenski, R.E.; Cure, S.; Kim, J.F. Tracing ancestors and relatives of Escherichia coli B, and the derivation of B strains REL606 and BL21(DE3). J. Mol. Biol. 2009, 394, 634–643. [Google Scholar] [CrossRef]
- Morris, A.C.; Kefala, K.; Wilkinson, T.S.; Dhaliwal, K.; Farrell, L.; Walsh, T.; Mackenzie, S.J.; Reid, H.; Davidson, D.J.; Haslett, C.; et al. C5a mediates peripheral blood neutrophil dysfunction in critically ill patients. Am. J. Respir. Crit. Care Med. 2009, 180, 19–28. [Google Scholar] [CrossRef]
- Morris, A.C.; Brittan, M.; Wilkinson, T.S.; McAuley, D.F.; Antonelli, J.; McCulloch, C.; Barr, L.C.; McDonald, N.A.; Dhaliwal, K.; Jones, R.O.; et al. C5a-mediated neutrophil dysfunction is RhoA-dependent and predicts infection in critically ill patients. Blood 2011, 117, 5178–5188. [Google Scholar] [CrossRef]
- Wilkinson, T.S.; Morris, A.C.; Kefala, K.; O’Kane, C.M.; Moore, N.R.; Booth, N.A.; McAuley, D.F.; Dhaliwal, K.; Walsh, T.S.; Haslett, C.; et al. Ventilator-associated pneumonia is characterized by excessive release of neutrophil proteases in the lung. Chest 2012, 142, 1425–1432. [Google Scholar] [CrossRef]
- O’Neill, A.J. Staphylococcus aureus SH1000 and 8325-4: Comparative genome sequences of key laboratory strains in staphylococcal research. Lett. Appl. Microbiol. 2010, 51, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Mack, D.; Siemssen, N.; Laufs, R. Parallel induction by glucose of adherence and a polysaccharide antigen specific for plastic-adherent Staphylococcus epidermidis: Evidence for functional relation to intercellular adhesion. Infect. Immun. 1992, 60, 2048–2057. [Google Scholar] [CrossRef] [PubMed]
- Christensen, G.D.; Simpson, W.A.; Bisno, A.L.; Beachey, E.H. Adherence of slime-producing strains of Staphylococcus epidermidis to smooth surfaces. Infect. Immun. 1982, 37, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.R.; Fouts, D.E.; Archer, G.L.; Mongodin, E.F.; Deboy, R.T.; Ravel, J.; Paulsen, I.T.; Kolonay, J.F.; Brinkac, L.; Beanan, M.; et al. Insights on evolution of virulence and resistance from the complete genome analysis of an early methicillin-resistant Staphylococcus aureus strain and a biofilm-producing methicillin-resistant Staphylococcus epidermidis strain. J. Bacteriol. 2005, 187, 2426–2438. [Google Scholar] [CrossRef]
- Evans, S.J.; Roberts, A.E.L.; Morris, A.C.; Simpson, A.J.; Harris, L.G.; Mack, D.; Jenkins, R.E.; Wilkinson, T.S. Contrasting effects of linezolid on healthy and dysfunctional human neutrophils: Reducing C5a-induced injury. Sci. Rep. 2020, 10, 16377. [Google Scholar] [CrossRef]
- Elemraid, M.A.; Rushton, S.P.; Thomas, M.F.; Spencer, D.A.; Gennery, A.R.; Clark, J.E. Utility of inflammatory markers in predicting the aetiology of pneumonia in children. Diagn. Microbiol. Infect. Dis. 2014, 79, 458–462. [Google Scholar] [CrossRef]
- R-Core-Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 21 March 2024).
- Opota, O.; Croxatto, A.; Prod’hom, G.; Greub, G. Blood culture-based diagnosis of bacteraemia: State of the art. Clin. Microbiol. Infect. 2015, 21, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Dietzman, D.E.; Fischer, G.W.; Schoenknecht, F.D. Neonatal Escherichia coli septicemia—bacterial counts in blood. J. Pediatr. 1974, 85, 128–130. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.P.; Carvalho, C.M. Burden of bacterial bloodstream infections and recent advances for diagnosis. Pathog. Dis. 2022, 80, ftac027. [Google Scholar] [CrossRef]
- Gahlot, R.; Nigam, C.; Kumar, V.; Yadav, G.; Anupurba, S. Catheter-related bloodstream infections. Int. J. Crit. Illn. Inj. Sci. 2014, 4, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Stranieri, I.; Kanunfre, K.A.; Rodrigues, J.C.; Yamamoto, L.; Nadaf, M.I.V.; Palmeira, P.; Okay, T.S. Assessment and comparison of bacterial load levels determined by quantitative amplifications in blood culture-positive and negative neonatal sepsis. Rev. Inst. Med. Trop. Sao Paulo 2018, 60, e61. [Google Scholar] [CrossRef] [PubMed]
- Rai, V.; Mathews, G.; Agrawal, D.K. Translational and Clinical Significance of DAMPs, PAMPs, and PRRs in Trauma-induced Inflammation. Arch. Clin. Biomed. Res. 2022, 6, 673–685. [Google Scholar] [CrossRef]
- Cicchinelli, S.; Pignataro, G.; Gemma, S.; Piccioni, A.; Picozzi, D.; Ojetti, V.; Franceschi, F.; Candelli, M. PAMPs and DAMPs in Sepsis: A Review of Their Molecular Features and Potential Clinical Implications. Int. J. Mol. Sci. 2024, 25, 962. [Google Scholar] [CrossRef]
- Bacconi, A.; Richmond, G.S.; Baroldi, M.A.; Laffler, T.G.; Blyn, L.B.; Carolan, H.E.; Frinder, M.R.; Toleno, D.M.; Metzgar, D.; Gutierrez, J.R.; et al. Improved sensitivity for molecular detection of bacterial and Candida infections in blood. J. Clin. Microbiol. 2014, 52, 3164–3174. [Google Scholar] [CrossRef]
- Tallosy, S.P.; Poles, M.Z.; Rutai, A.; Fejes, R.; Juhasz, L.; Burian, K.; Soki, J.; Szabo, A.; Boros, M.; Kaszaki, J. The microbial composition of the initial insult can predict the prognosis of experimental sepsis. Sci. Rep. 2021, 11, 22772. [Google Scholar] [CrossRef]
- Hessle, C.C.; Andersson, B.; Wold, A.E. Gram-positive and Gram-negative bacteria elicit different patterns of pro-inflammatory cytokines in human monocytes. Cytokine 2005, 30, 311–318. [Google Scholar] [CrossRef]
- Iwadou, H.; Morimoto, Y.; Iwagaki, H.; Sinoura, S.; Chouda, Y.; Kodama, M.; Yoshioka, T.; Saito, S.; Yagi, T.; Tanaka, N. Differential cytokine response in host defence mechanisms triggered by gram-negative and gram-positive bacteria, and the roles of gabexate mesilate, a synthetic protease inhibitor. J. Int. Med. Res. 2002, 30, 99–108. [Google Scholar] [CrossRef]
- Skovbjerg, S.; Martner, A.; Hynsjo, L.; Hessle, C.; Olsen, I.; Dewhirst, F.E.; Tham, W.; Wold, A.E. Gram-positive and gram-negative bacteria induce different patterns of cytokine production in human mononuclear cells irrespective of taxonomic relatedness. J. Interferon Cytokine Res. 2010, 30, 23–32. [Google Scholar] [CrossRef]
- Beran, O.; Potmesil, R.; Holub, M. Differences in Toll-like receptor expression and cytokine production after stimulation with heat-killed Gram-positive and Gram-negative bacteria. Folia Microbiol. 2011, 56, 283–287. [Google Scholar] [CrossRef]
- Abe, R.; Oda, S.; Sadahiro, T.; Nakamura, M.; Hirayama, Y.; Tateishi, Y.; Shinozaki, K.; Hirasawa, H. Gram-negative bacteremia induces greater magnitude of inflammatory response than Gram-positive bacteremia. Crit. Care 2010, 14, R27. [Google Scholar] [CrossRef]
- Surbatovic, M.; Popovic, N.; Vojvodic, D.; Milosevic, I.; Acimovic, G.; Stojicic, M.; Veljovic, M.; Jevdjic, J.; Djordjevic, D.; Radakovic, S. Cytokine profile in severe Gram-positive and Gram-negative abdominal sepsis. Sci. Rep. 2015, 5, 11355. [Google Scholar] [CrossRef]
- Xu, X.J.; Tang, Y.M.; Liao, C.; Song, H.; Yang, S.L.; Xu, W.Q.; Shi, S.W.; Zhao, N. Inflammatory cytokine measurement quickly discriminates gram-negative from gram-positive bacteremia in pediatric hematology/oncology patients with septic shock. Intensive Care Med. 2013, 39, 319–326. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, T.; Chen, H.J.; Chen, N.; Xing, Z.X.; Fu, X.Y. Risk prediction model for distinguishing Gram-positive from Gram-negative bacteremia based on age and cytokine levels: A retrospective study. World J. Clin. Cases 2023, 11, 4833–4842. [Google Scholar] [CrossRef]
- Guan, J.; Wang, Z.; Liu, X.; Jiang, Y.; Gao, Q.; Wu, Q.; Lu, H.; Wu, L.; Zhang, Z.; Lin, X.; et al. IL-6 and IL-10 Closely Correlate with Bacterial Bloodstream Infection. Iran. J. Immunol. 2020, 17, 185–203. [Google Scholar] [CrossRef]
- Tietze, K.; Dalpke, A.; Morath, S.; Mutters, R.; Heeg, K.; Nonnenmacher, C. Differences in innate immune responses upon stimulation with gram-positive and gram-negative bacteria. J. Periodontal. Res. 2006, 41, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Hoerr, V.; Zbytnuik, L.; Leger, C.; Tam, P.P.; Kubes, P.; Vogel, H.J. Gram-negative and Gram-positive bacterial infections give rise to a different metabolic response in a mouse model. J. Proteome Res. 2012, 11, 3231–3245. [Google Scholar] [CrossRef] [PubMed]
- Barber, A.E.; Fleming, B.A.; Mulvey, M.A. Similarly Lethal Strains of Extraintestinal Pathogenic Escherichia coli Trigger Markedly Diverse Host Responses in a Zebrafish Model of Sepsis. mSphere 2016, 1, e00062-16. [Google Scholar] [CrossRef] [PubMed]
- Schroder, N.W.; Morath, S.; Alexander, C.; Hamann, L.; Hartung, T.; Zahringer, U.; Gobel, U.B.; Weber, J.R.; Schumann, R.R. Lipoteichoic acid (LTA) of Streptococcus pneumoniae and Staphylococcus aureus activates immune cells via Toll-like receptor (TLR)-2, lipopolysaccharide-binding protein (LBP), and CD14, whereas TLR-4 and MD-2 are not involved. J. Biol. Chem. 2003, 278, 15587–15594. [Google Scholar] [CrossRef] [PubMed]
- Volz, T.; Nega, M.; Buschmann, J.; Kaesler, S.; Guenova, E.; Peschel, A.; Rocken, M.; Gotz, F.; Biedermann, T. Natural Staphylococcus aureus-derived peptidoglycan fragments activate NOD2 and act as potent costimulators of the innate immune system exclusively in the presence of TLR signals. FASEB J. 2010, 24, 4089–4102. [Google Scholar] [CrossRef] [PubMed]
- Hajjar, A.M.; O’Mahony, D.S.; Ozinsky, A.; Underhill, D.M.; Aderem, A.; Klebanoff, S.J.; Wilson, C.B. Cutting edge: Functional interactions between toll-like receptor (TLR) 2 and TLR1 or TLR6 in response to phenol-soluble modulin. J. Immunol. 2001, 166, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Takada, H.; Kaneko, T.; Kato, I.; Golenbock, D.; Hara, Y. Structural requirements of muramylpeptides for induction of Toll-like receptor 2-mediated NF-kappaB activation in CHO cells. J. Endotoxin. Res. 2000, 6, 407–410. [Google Scholar] [CrossRef]
- Stevens, N.T.; Sadovskaya, I.; Jabbouri, S.; Sattar, T.; O’Gara, J.P.; Humphreys, H.; Greene, C.M. Staphylococcus epidermidis polysaccharide intercellular adhesin induces IL-8 expression in human astrocytes via a mechanism involving TLR2. Cell Microbiol. 2009, 11, 421–432. [Google Scholar] [CrossRef]
- O’Grady, N.P.; Tropea, M.; Preas, H.L., II; Reda, D.; Vandivier, R.W.; Banks, S.M.; Suffredini, A.F. Detection of macrophage inflammatory protein (MIP)-1alpha and MIP-1beta during experimental endotoxemia and human sepsis. J. Infect. Dis. 1999, 179, 136–141. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, B.; Ning, B. Evaluating IL-6 and IL-10 as rapid diagnostic tools for Gram-negative bacteria and as disease severity predictors in pediatric sepsis patients in the intensive care unit. Front. Immunol. 2022, 13, 1043968. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, Y.; Yu, B.; Zhang, Z.; Fan, Y.; Wang, L.; Cheng, M.; Yan, P.; Zhao, W. Evaluation of the diagnostic and prognostic values of serum HSP90alpha in sepsis patients: A retrospective study. PeerJ 2022, 10, e12997. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, T.; Matsumoto, H.; Matsubara, T.; Matsuura, H.; Hirose, T.; Shimizu, K.; Ogura, H.; Kang, S.; Tanaka, T.; Shimazu, T. Adipocytokine Profile Reveals Resistin Forming a Prognostic-Related Cytokine Network in the Acute Phase of Sepsis. Shock 2021, 56, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Vassiliadi, D.A.; Tzanela, M.; Kotanidou, A.; Orfanos, S.E.; Nikitas, N.; Armaganidis, A.; Koutsilieris, M.; Roussos, C.; Tsagarakis, S.; Dimopoulou, I. Serial changes in adiponectin and resistin in critically ill patients with sepsis: Associations with sepsis phase, severity, and circulating cytokine levels. J. Crit. Care 2012, 27, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Sunden-Cullberg, J.; Nystrom, T.; Lee, M.L.; Mullins, G.E.; Tokics, L.; Andersson, J.; Norrby-Teglund, A.; Treutiger, C.J. Pronounced elevation of resistin correlates with severity of disease in severe sepsis and septic shock. Crit. Care Med. 2007, 35, 1536–1542. [Google Scholar] [CrossRef] [PubMed]
- Gierlikowska, B.; Stachura, A.; Gierlikowski, W.; Demkow, U. The Impact of Cytokines on Neutrophils’ Phagocytosis and NET Formation during Sepsis-A Review. Int. J. Mol. Sci. 2022, 23, 5076. [Google Scholar] [CrossRef] [PubMed]
- Miajlovic, H.; Smith, S.G. Bacterial self-defence: How Escherichia coli evades serum killing. FEMS Microbiol. Lett. 2014, 354, 1–9. [Google Scholar] [CrossRef]
- Guerra, F.E.; Borgogna, T.R.; Patel, D.M.; Sward, E.W.; Voyich, J.M. Epic Immune Battles of History: Neutrophils vs. Staphylococcus aureus. Front. Cell Infect. Microbiol. 2017, 7, 286. [Google Scholar] [CrossRef]
- Osterud, B. Tissue factor expression by monocytes: Regulation and pathophysiological roles. Blood Coagul. Fibrinolysis 1998, 9 (Suppl. 1), S9–S14. [Google Scholar]
- Kopp, R.; Bernsberg, R.; Kashefi, A.; Mottaghy, K.; Rossaint, R.; Kuhlen, R. Effect of hirudin versus heparin on hemocompatibility of blood contacting biomaterials: An in vitro study. Int. J. Artif. Organs 2005, 28, 1272–1277. [Google Scholar] [CrossRef]
- Zhang, T.; Garstka, M.A.; Li, K. The Controversial C5a Receptor C5aR2: Its Role in Health and Disease. J. Immunol. Res. 2017, 2017, 8193932. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.O.; Larsen, G.L.; Henson, P.M. In vivo clearance and tissue distribution of C5a and C5a des arginine complement fragments in rabbits. J. Clin. Investig. 1982, 70, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.M.; Popper, S.J.; Gupta, S.; Davong, V.; Vaidya, K.; Chanthongthip, A.; Dittrich, S.; Robinson, M.T.; Vongsouvath, M.; Mayxay, M.; et al. A robust host-response-based signature distinguishes bacterial and viral infections across diverse global populations. Cell Rep. Med. 2022, 3, 100842. [Google Scholar] [CrossRef]
- Mahle, R.E.; Suchindran, S.; Henao, R.; Steinbrink, J.M.; Burke, T.W.; McClain, M.T.; Ginsburg, G.S.; Woods, C.W.; Tsalik, E.L. Validation of a Host Gene Expression Test for Bacterial/Viral Discrimination in Immunocompromised Hosts. Clin. Infect. Dis. 2021, 73, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Chawla, D.G.; Cappuccio, A.; Tamminga, A.; Sealfon, S.C.; Zaslavsky, E.; Kleinstein, S.H. Benchmarking transcriptional host response signatures for infection diagnosis. Cell Syst. 2022, 13, 974.e7–988.e7. [Google Scholar] [CrossRef] [PubMed]
- Sohn, E. Diagnosis: Frontiers in blood testing. Nature 2017, 549, S16–S18. [Google Scholar] [CrossRef]
- Ng, S.; Strunk, T.; Jiang, P.; Muk, T.; Sangild, P.T.; Currie, A. Precision Medicine for Neonatal Sepsis. Front. Mol. Biosci. 2018, 5, 70. [Google Scholar] [CrossRef]
- Mejias, A.; Suarez, N.M.; Ramilo, O. Detecting specific infections in children through host responses: A paradigm shift. Curr. Opin. Infect. Dis. 2014, 27, 228–235. [Google Scholar] [CrossRef]
- Morris, A.C.; Gadsby, N.; McKenna, J.P.; Hellyer, T.P.; Dark, P.; Singh, S.; Walsh, T.S.; McAuley, D.F.; Templeton, K.; Simpson, A.J.; et al. 16S pan-bacterial PCR can accurately identify patients with ventilator-associated pneumonia. Thorax 2017, 72, 1046–1048. [Google Scholar] [CrossRef]
- Thavasu, P.W.; Longhurst, S.; Joel, S.P.; Slevin, M.L.; Balkwill, F.R. Measuring cytokine levels in blood. Importance of anticoagulants, processing, and storage conditions. J. Immunol. Methods 1992, 153, 115–124. [Google Scholar] [CrossRef]
- Grievink, H.W.; Moerland, M. Sample Aging Profoundly Reduces Monocyte Responses in Human Whole Blood Cultures. J. Immunol. Res. 2018, 2018, 8901485. [Google Scholar] [CrossRef] [PubMed]
- Reid, W.D.; Close, A.J.; Humphrey, S.; Chaloner, G.; Lacharme-Lora, L.; Rothwell, L.; Kaiser, P.; Williams, N.J.; Humphrey, T.J.; Wigley, P.; et al. Cytokine responses in birds challenged with the human food-borne pathogen Campylobacter jejuni implies a Th17 response. R. Soc. Open Sci. 2016, 3, 150541. [Google Scholar] [CrossRef] [PubMed]
- Pripp, A.H.; Stanisic, M. The correlation between pro- and anti-inflammatory cytokines in chronic subdural hematoma patients assessed with factor analysis. PLoS ONE 2014, 9, e90149. [Google Scholar] [CrossRef] [PubMed]
- Picat, M.Q.; Pellegrin, I.; Bitard, J.; Wittkop, L.; Proust-Lima, C.; Liquet, B.; Moreau, J.F.; Bonnet, F.; Blanco, P.; Thiebaut, R. Integrative Analysis of Immunological Data to Explore Chronic Immune T-Cell Activation in Successfully Treated HIV Patients. PLoS ONE 2017, 12, e0169164. [Google Scholar] [CrossRef]
- Bradbury, J.; Brooks, L.; Myers, S.P. Are the Adaptogenic Effects of Omega 3 Fatty Acids Mediated via Inhibition of Proinflammatory Cytokines? Evid. Based Complement. Alternat. Med. 2012, 2012, 209197. [Google Scholar] [CrossRef][Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | Correct Predictions (Observations) | Correct Prediction (%) |
|---|---|---|
| Strain | 16/40 | 40 |
| Species | 36/40 | 90 |
| Genera | 38/40 | 95 |
| a: Variables Predicting Strain | |||
| Biomarker | Wilks Lambda | p Value Overall | p Value Diff |
| Phagocytosis | 0.564 | 0.0064 | 0.0064 |
| S100A8 | 0.396 | 0.0050 | 0.1053 |
| IL-10 | 0.259 | 0.0017 | 0.0546 |
| b: Variables Predicting Species | |||
| Biomarker | Wilks Lambda | p Value Overall | p Value Diff |
| IL-6 | 0.313 | 0.0000 | 0.0000 |
| Phagocytosis | 0.193 | 0.0000 | 0.0187 |
| Resistin | 0.133 | 0.0000 | 0.0888 |
| TF3 | 0.095 | 0.0000 | 0.1527 |
| c: Variables Predicting Genera | |||
| Biomarker | Wilks Lambda | p Value Overall | p Value Diff |
| IL-6 | 0.391 | 0.0000 | 0.0000 |
| TF3 | 0.336 | 0.0000 | 0.0179 |
| MIP3 | 0.286 | 0.0000 | 0.0166 |
| Advantages | Limitations |
|---|---|
| Simple and rapid to perform | Lack of endothelial cells and other immune mediator-producing cells |
| Can measure both soluble and cellular mediators of inflammation | Presence of anticoagulants interfering with complement and coagulation pathways |
| Versatile application to a variety of assays and disease situations | Absence of liver and acute phase proteins |
| Only basic healthy volunteer ethics required | Potential for donor variation |
| Experiments completed in one day | Requires checks on plastic/vehicle activation |
| Use of a primary biological tissue/fluid/suspension | Longer experiments (>8 h) not feasible |
| Complements the human endotoxemia model without compromising patient safety | Impact of disease severity (sepsis vs. septic shock) not possible to measure |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chick, H.M.; Rees, M.E.; Lewis, M.L.; Williams, L.K.; Bodger, O.; Harris, L.G.; Rushton, S.; Wilkinson, T.S. Using the Traditional Ex Vivo Whole Blood Model to Discriminate Bacteria by Their Inducible Host Responses. Biomedicines 2024, 12, 724. https://doi.org/10.3390/biomedicines12040724
Chick HM, Rees ME, Lewis ML, Williams LK, Bodger O, Harris LG, Rushton S, Wilkinson TS. Using the Traditional Ex Vivo Whole Blood Model to Discriminate Bacteria by Their Inducible Host Responses. Biomedicines. 2024; 12(4):724. https://doi.org/10.3390/biomedicines12040724
Chicago/Turabian StyleChick, Heather M., Megan E. Rees, Matthew L. Lewis, Lisa K. Williams, Owen Bodger, Llinos G. Harris, Steven Rushton, and Thomas S. Wilkinson. 2024. "Using the Traditional Ex Vivo Whole Blood Model to Discriminate Bacteria by Their Inducible Host Responses" Biomedicines 12, no. 4: 724. https://doi.org/10.3390/biomedicines12040724
APA StyleChick, H. M., Rees, M. E., Lewis, M. L., Williams, L. K., Bodger, O., Harris, L. G., Rushton, S., & Wilkinson, T. S. (2024). Using the Traditional Ex Vivo Whole Blood Model to Discriminate Bacteria by Their Inducible Host Responses. Biomedicines, 12(4), 724. https://doi.org/10.3390/biomedicines12040724