
Amelioration of Morphological Pathology in Cardiac, Respiratory, and Skeletal Muscles Following Intraosseous Administration of Human Dystrophin Expressing Chimeric (DEC) Cells in Duchenne Muscular Dystrophy Model

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice and Animal Care
2.2. Creation of Human DEC Cells
2.3. Transplantation of DEC
2.4. Histological Analysis
2.5. Minimal Feret’s Diameter for Morphological Analysis
2.6. Statistical Analysis
3. Results
3.1. Local Safety and Tolerance Were Confirmed at 90 and 180 Days Following Transplantation of Human DEC Therapy to the mdx/scid Mice via Systemic-Intraosseous Route
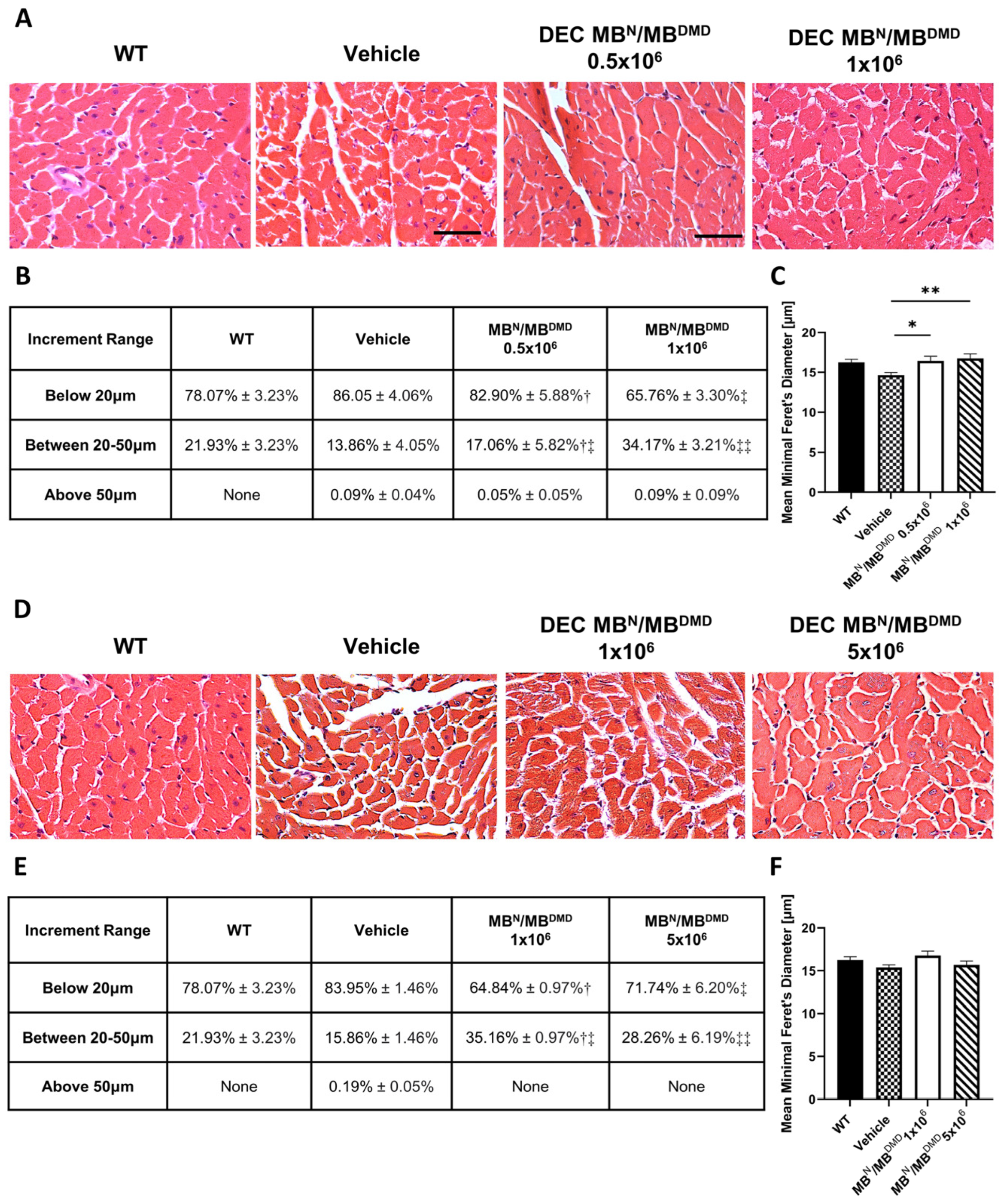
3.2. Transplantation of Human DEC Therapy Results in Improvement of Morphology of Cardiac Muscle at 90 and 180 Days after Intraosseous Transplantation
3.3. Human DEC Therapy Increases Overall Homogeneity of the Diaphragm at 90 and 180 Days Following Systemic Delivery via Intraosseous Route
3.4. Transplantation of Human DEC Therapy Normalizes Gastrocnemius Muscle Morphology at 90 and 180 Days Following Systemic-Intraosseous Transplantation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Ruiten, H.J.; Bettolo, C.M.; Cheetham, T.; Eagle, M.; Lochmuller, H.; Straub, V.; Bushby, K.; Guglieri, M. Why are some patients with Duchenne muscular dystrophy dying young: An analysis of causes of death in North East England. Eur. J. Paediatr. Neurol. 2016, 20, 904–909. [Google Scholar] [CrossRef]
- Maggio, I.; Liu, J.; Janssen, J.M.; Chen, X.; Gonçalves, M.A.F.V. Adenoviral vectors encoding CRISPR/Cas9 multiplexes rescue dystrophin synthesis in unselected populations of DMD muscle cells. Sci. Rep. 2016, 6, 37051. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, E.P.; Brown, R.H.; Kunkel, L.M. Dystrophin: The protein product of the duchenne muscular dystrophy locus. Cell 1987, 51, 919–928. [Google Scholar] [CrossRef]
- Kharraz, Y.; Guerra, J.; Pessina, P.; Serrano, A.L.; Muñoz-Cánoves, P. Understanding the process of fibrosis in duchenne muscular dystrophy. BioMed Res. Int. 2014, 2014, 965631. [Google Scholar] [CrossRef] [PubMed]
- Porcari, P.; Hall, M.G.; Clark, C.A.; Greally, E.; Straub, V.; Blamire, A.M. Time-dependent diffusion MRI as a probe of microstructural changes in a mouse model of Duchenne muscular dystrophy. NMR Biomed. 2020, 33, e4276. [Google Scholar] [CrossRef]
- Briguet, A.; Courdier-Fruh, I.; Foster, M.; Meier, T.; Magyar, J.P. Histological parameters for the quantitative assessment of muscular dystrophy in the mdx-mouse. Neuromuscul. Disord. 2004, 14, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, P.V.; Hooper, A.C. Sarcomere length and fibre diameter distributions in four different mouse skeletal muscles. J. Anat. 1971, 110 Pt 2, 249–257. [Google Scholar]
- Cernisova, V.; Lu-Nguyen, N.; Trundle, J.; Herath, S.; Malerba, A.; Popplewell, L. Microdystrophin Gene Addition Significantly Improves Muscle Functionality and Diaphragm Muscle Histopathology in a Fibrotic Mouse Model of Duchenne Muscular Dystrophy. Int. J. Mol. Sci. 2023, 24, 8174. [Google Scholar] [CrossRef]
- Florczyk-Soluch, U.; Polak, K.; Dulak, J. The multifaceted view of heart problem in Duchenne muscular dystrophy. Cell. Mol. Life Sci. 2021, 78, 5447–5468. [Google Scholar] [CrossRef]
- Abdel-Salam, E.; Abdel-Meguid, I.; Korraa, S.S. Markers of degeneration and regeneration in Duchenne muscular dystrophy. Acta Myol. 2009, 28, 94–100. [Google Scholar]
- Rodrigues, M.; Echigoya, Y.; Maruyama, R.; Lim, K.R.; Fukada, S.I.; Yokota, T. Impaired regenerative capacity and lower revertant fibre expansion in dystrophin-deficient mdx muscles on DBA/2 background. Sci. Rep. 2016, 6, 38371. [Google Scholar] [CrossRef]
- Barthélémy, F.; Wein, N. Personalized gene and cell therapy for Duchenne Muscular Dystrophy. Neuromuscul. Disord. 2018, 28, 803–824. [Google Scholar] [CrossRef]
- Biressi, S.; Filareto, A.; Rando, T.A. Stem cell therapy for muscular dystrophies. J. Clin. Investig. 2020, 130, 5652–5664. [Google Scholar] [CrossRef]
- Sienkiewicz, D.; Kulak, W.; Okurowska-Zawada, B.; Paszko-Patej, G.; Kawnik, K. Duchenne muscular dystrophy: Current cell therapies. Ther. Adv. Neurol. Disord. 2015, 8, 166–177. [Google Scholar] [CrossRef]
- Maggio, I.; Chen, X.; Gonçalves, M.A.F.V. The emerging role of viral vectors as vehicles for DMD gene editing. Genome Med. 2016, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Hollinger, K.; Chamberlain, J.S. Viral vector-mediated gene therapies. Curr. Opin. Neurol. 2015, 28, 522–527. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Long, C.; McAnally, J.R.; Shelton, J.M.; Mireault, A.A.; Bassel-Duby, R.; Olson, E.N. Prevention of muscular dystrophy in mice by CRISPR/Cas9–mediated editing of germline DNA. Science 2014, 345, 1184–1188. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.E.; Hakim, C.H.; Ousterout, D.G.; Thakore, P.I.; Moreb, E.A.; Rivera, R.M.C.; Madhavan, S.; Pan, X.; Ran, F.A.; Yan, W.X.; et al. In vivo genome editing improves muscle function in a mouse model of Duchenne muscular dystrophy. Science 2016, 351, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Duan, D. Micro-Dystrophin Gene Therapy Goes Systemic in Duchenne Muscular Dystrophy Patients. Hum. Gene Ther. 2018, 29, 733–736. [Google Scholar] [CrossRef]
- Kornegay, J.N.; Li, J.; Bogan, J.R.; Bogan, D.J.; Chen, C.; Zheng, H.; Wang, B.; Qiao, C.; Howard, J.F.; Xiao, X. Widespread Muscle Expression of an AAV9 Human Mini-dystrophin Vector After Intravenous Injection in Neonatal Dystrophin-deficient Dogs. Mol. Ther. 2010, 18, 1501–1508. [Google Scholar] [CrossRef]
- Sun, C.; Shen, L.; Zhang, Z.; Xie, X. Therapeutic Strategies for Duchenne Muscular Dystrophy: An Update. Genes 2020, 11, 837. [Google Scholar] [CrossRef]
- Judson, R.N.; Rossi, F.M.V. Towards stem cell therapies for skeletal muscle repair. NPJ Regen. Med. 2020, 5, 10. [Google Scholar] [CrossRef]
- Cossu, G.; Previtali, S.C.; Napolitano, S.; Cicalese, M.P.; Tedesco, F.S.; Nicastro, F.; Noviello, M.; Roostalu, U.; Sora, M.G.N.; Scarlato, M.; et al. Intra-arterial transplantation of HLA -matched donor mesoangioblasts in Duchenne muscular dystrophy. EMBO Mol. Med. 2015, 7, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Govoni, A.; Magri, F.; Brajkovic, S.; Zanetta, C.; Faravelli, I.; Corti, S.; Bresolin, N.; Comi, G.P. Ongoing therapeutic trials and outcome measures for Duchenne muscular dystrophy. Cell. Mol. Life Sci. 2013, 70, 4585–4602. [Google Scholar] [CrossRef]
- Rajput, B.S.; Chakrabarti, S.K.; Dongare, V.S.; Ramirez, C.M.; Deb, K.D. Human Umbilical Cord Mesenchymal Stem Cells in the Treatment of Duchenne Muscular Dystrophy: Safety and Feasibility Study in India. J. Stem Cells 2015, 10, 141–156. [Google Scholar] [PubMed]
- Skuk, D.; Tremblay, J.P. Cell therapy in muscular dystrophies: Many promises in mice and dogs, few facts in patients. Expert Opin. Biol. Ther. 2015, 15, 1307–1319. [Google Scholar] [CrossRef] [PubMed]
- Torrente, Y.; Belicchi, M.; Marchesi, C.; D’antona, G.; Cogiamanian, F.; Pisati, F.; Gavina, M.; Giordano, R.; Tonlorenzi, R.; Fagiolari, G.; et al. Autologous Transplantation of Muscle-Derived CD133+ Stem Cells in Duchenne Muscle Patients. Cell Transplant. 2007, 16, 563–577. [Google Scholar] [CrossRef] [PubMed]
- Siemionow, M.; Langa, P.; Harasymczuk, M.; Cwykiel, J.; Sielewicz, M.; Smieszek, J.; Heydemann, A. Human Dystrophin Expressing Chimeric (DEC) Cell Therapy Ameliorates Cardiac, Respiratory, and Skeletal Muscle’s Function in Duchenne Muscular Dystrophy. Stem Cells Transl. Med. 2021, 10, 1406–1418. [Google Scholar] [CrossRef]
- Siemionow, M.; Langa, P.; Brodowska, S.; Kozlowska, K.; Zalants, K.; Budzynska, K.; Heydemann, A. Long-Term Protective Effect of Human Dystrophin Expressing Chimeric (DEC) Cell Therapy on Amelioration of Function of Cardiac, Respiratory and Skeletal Muscles in Duchenne Muscular Dystrophy. Stem Cell Rev. Rep. 2022, 18, 2872–2892. [Google Scholar] [CrossRef]
- Muir, L.A.; Murry, C.E.; Chamberlain, J.S. Prosurvival Factors Improve Functional Engraftment of Myogenically Converted Dermal Cells into Dystrophic Skeletal Muscle. Stem Cells Dev. 2016, 25, 1559–1569. [Google Scholar] [CrossRef]
- Meregalli, M.; Farini, A.; Belicchi, M.; Parolini, D.; Cassinelli, L.; Razini, P.; Sitzia, C.; Torrente, Y. Perspectives of stem cell therapy in Duchenne muscular dystrophy. FEBS J. 2013, 280, 4251–4262. [Google Scholar] [CrossRef]
- Miura, Y.; Sato, M.; Kuwahara, T.; Ebata, T.; Tabata, Y.; Sakurai, H. Transplantation of human iPSC-derived muscle stem cells in the diaphragm of Duchenne muscular dystrophy model mice. PLoS ONE 2022, 17, e0266391. [Google Scholar] [CrossRef]
- Sitzia, C.; Farini, A.; Jardim, L.; Razini, P.; Belicchi, M.; Cassinelli, L.; Villa, C.; Erratico, S.; Parolini, D.; Bella, P.; et al. Adaptive Immune Response Impairs the Efficacy of Autologous Transplantation of Engineered Stem Cells in Dystrophic Dogs. Mol. Ther. 2016, 24, 1949–1964. [Google Scholar] [CrossRef] [PubMed]
- Gussoni, E.; Blau, H.M.; Kunkel, L.M. The fate of individual myoblasts after transplantation into muscles of DMD patients. Nat. Med. 1997, 3, 970–977. [Google Scholar] [CrossRef]
- Nitahara-Kasahara, Y.; Hayashita-Kinoh, H.; Ohshima-Hosoyama, S.; Okada, H.; Wada-Maeda, M.; Nakamura, A.; Okada, T.; Takeda, S. Long-term engraftment of multipotent mesenchymal stromal cells that differentiate to form myogenic cells in dogs with Duchenne muscular dystrophy. Mol. Ther. 2021, 20, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, Y.; Li, Y.; Cao, J.; Zhang, H.; Chen, M.; Wang, L.; Zhang, C. Long-term engraftment of myogenic progenitors from adipose-derived stem cells and muscle regeneration in dystrophic mice. Hum. Mol. Genet. 2015, 24, 6029–6040. [Google Scholar] [CrossRef] [PubMed]
- Cwykiel, J.; Jundzill, A.; Klimczak, A.; Madajka-Niemeyer, M.; Siemionow, M. Donor Recipient Chimeric Cells Induce Chimerism and Extend Survival of Vascularized Composite Allografts. Arch. Immunol. Ther. Exp. 2021, 69, 13. [Google Scholar] [CrossRef] [PubMed]
- Siemionow, M.; Cwykiel, J.; Heydemann, A.; Garcia-Martinez, J.; Szilagyi, E. Creation of Dystrophin Expressing Chimeric Cells of Myoblast Origin as a Novel Stem Cell Based Therapy for Duchenne Muscular Dystrophy. Stem Cell Rev. Rep. 2018, 14, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.D.; Heitzman, J.A.; Townsend, D. Hyperbaric therapy provides no benefit for skeletal muscle and respiratory function and accelerates cardiac injury in mdx mice. Sci. Rep. 2019, 9, 12306. [Google Scholar] [CrossRef] [PubMed]
- Birnkrant, D.J.; Bello, L.; Butterfield, R.J.; Carter, J.C.; Cripe, L.H.; Cripe, T.P.; McKim, D.A.; Nandi, D.; Pegoraro, E. Cardiorespiratory management of Duchenne muscular dystrophy: Emerging therapies, neuromuscular genetics, and new clinical challenges. Lancet Respir. Med. 2022, 10, 403–420. [Google Scholar] [CrossRef]
- Falzarano, M.S.; Scotton, C.; Passarelli, C.; Ferlini, A. Duchenne Muscular Dystrophy: From Diagnosis to Therapy. Molecules 2015, 20, 18168–18184. [Google Scholar] [CrossRef]
- Nowak, K.J.; E Davies, K. Duchenne muscular dystrophy and dystrophin: Pathogenesis and opportunities for treatment. EMBO Rep. 2004, 5, 872–876. [Google Scholar] [CrossRef]
- Valentine, B.A.; Cooper, B.J.; Cummings, J.F.; Lahunta, A. Canine X-linked muscular dystrophy: Morphologic lesions. J. Neurol. Sci. 1990, 97, 1–23. [Google Scholar] [CrossRef]
- Strehle, E.M.; Straub, V. Recent advances in the management of Duchenne muscular dystrophy. Arch. Dis. Child. 2015, 100, 1173–1177. [Google Scholar] [CrossRef]
- Buyse, G.M.; Goemans, N.; Van Den Hauwe, M.; Meier, T. Effects of glucocorticoids and idebenone on respiratory function in patients with duchenne muscular dystrophy. Pediatr. Pulmonol. 2013, 48, 912–920. [Google Scholar] [CrossRef]
- Goemans, N.; Buyse, G. Current treatment and management of dystrophinopathies. Curr. Treat. Options Neurol. 2014, 16, 287. [Google Scholar] [CrossRef]
- Mah, J.K. Current and emerging treatment strategies for Duchenne muscular dystrophy. Neuropsychiatr. Dis. Treat. 2016, 12, 1795–1807. [Google Scholar] [CrossRef]
- D’Ambrosio, E.S.; Mendell, J.R. Evolving Therapeutic Options for the Treatment of Duchenne Muscular Dystrophy. Neurotherapeutics 2023, 20, 1669–1681. [Google Scholar] [CrossRef]
- Markati, T.; Oskoui, M.; A Farrar, M.; Duong, T.; Goemans, N.; Servais, L. Emerging therapies for Duchenne muscular dystrophy. Lancet Neurol. 2022, 21, 814–829. [Google Scholar] [CrossRef]
- Siemionow, M.; Cwykiel, J.; Brodowska, S.; Chambily, L. Human Multi-Chimeric Cell (HMCC) Therapy as a Novel Approach for Tolerance Induction in Transplantation. Stem Cell Rev. Rep. 2023, 19, 2741–2755. [Google Scholar] [CrossRef]
- Niezgoda, A.; Bieganski, G.; Wachowiak, J.; Czarnota, J.; Siemionow, K.; Heydemann, A.; Ziemiecka, A.; Sikorska, M.H.; Bożyk, K.; Siemionow, M. Assessment of Motor Unit Potentials Duration as the Biomarker of DT-DEC01 Cell Therapy Efficacy in Duchenne Muscular Dystrophy Patients up to 12 Months After Systemic–Intraosseous Administration. Arch. Immunol. Ther. Exp. 2023, 71, 24. [Google Scholar] [CrossRef]
- Stevens, C.R.; Berenson, J.; Sledziona, M.; Moore, T.P.; Dong, L.; Cheetham, J. Approach for semi-automated measurement of fiber diameter in murine and canine skeletal muscle. PLoS ONE 2020, 15, e0243163. [Google Scholar] [CrossRef]
- Rüegg, M.A.; Meinen, S. Quantitative Determination of Minimal Feret’s Diameter, Including the Evaluation of the Percentage of Centralized Nuclei, Fiber Numbers, Cross-Sectional Area and the Percentage of Fibrosis. Treat-NMD. 2008. Available online: https://treatnmd.org/wpcontent/uploads/2016/08/cmd-MDC1A_M.1.2.002-66.pdf (accessed on 26 February 2024).
- Ambroise, M.M.; Jothilingam, P.; Ramdas, A. Utility of nuclear morphometry in effusion cytology. Asian Pac. J. Cancer Prev. 2014, 15, 6919–6922. [Google Scholar] [CrossRef][Green Version]
- Tyagi, S.; Beqollari, D.; Lee, C.S.; Walker, L.A.; Bannister, R.A. Semi-automated Analysis of Mouse Skeletal Muscle Morphology and Fiber-type Composition. J. Vis. Exp. 2017, 126, 56024. [Google Scholar] [CrossRef]
- Gollapudi, S.K.; Michael, J.J.; Chandra, M. Striated Muscle Dynamics. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Sieck, D.C.; Zhan, W.; Fang, Y.; Ermilov, L.G.; Sieck, G.C.; Mantilla, C.B. Structure-Activity Relationships in Rodent Diaphragm Muscle Fibers vs. Neuromuscular Junctions. Respir. Physiol. Neurobiol. 2012, 180, 88–96. [Google Scholar] [CrossRef]
- Deconinck, N.; Dan, B. Pathophysiology of duchenne muscular dystrophy: Current hypotheses. Pediatr. Neurol. 2007, 36, 1–7. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Group * | Number of Images | Number of Fibers ** |
|---|---|---|---|
| Heart | WT | 32 | 3225 |
| Vehicle | 36 | 3932 | |
| MBN/MBDMD 0.5 × 106 | 36 | 3361 | |
| MBN/MBDMD 1 × 106 | 36 | 3390 | |
| Diaphragm | WT | 29 | 4997 |
| Vehicle | 30 | 4006 | |
| MBN/MBDMD 0.5 × 106 | 36 | 4706 | |
| MBN/MBDMD 1 × 106 | 35 | 4445 | |
| Gastrocnemius | WT | 24 | 2126 |
| Vehicle | 40 | 4427 | |
| MBN/MBDMD 0.5 × 106 | 35 | 3122 | |
| MBN/MBDMD 1 × 106 | 36 | 3414 | |
| Total | 405 | 45,151 |
| Organ | Group * | Number of Images | Number of Fibers ** |
|---|---|---|---|
| Heart | WT | 32 | 3225 |
| Vehicle | 36 | 2297 | |
| MBN/MBDMD 1 × 106 | 36 | 2012 | |
| MBN/MBDMD 5 × 106 | 36 | 1950 | |
| Diaphragm | WT | 29 | 4997 |
| Vehicle | 30 | 2126 | |
| MBN/MBDMD 1 × 106 | 25 | 2322 | |
| MBN/MBDMD 5 × 106 | 31 | 2471 | |
| Gastrocnemius | WT | 24 | 2126 |
| Vehicle | 34 | 2120 | |
| MBN/MBDMD 1 × 106 | 36 | 2219 | |
| MBN/MBDMD 5 × 106 | 36 | 2122 | |
| Total | 385 | 29,987 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siemionow, M.; Budzynska, K.; Zalants, K.; Langa, P.; Brodowska, S.; Siemionow, K.; Heydemann, A. Amelioration of Morphological Pathology in Cardiac, Respiratory, and Skeletal Muscles Following Intraosseous Administration of Human Dystrophin Expressing Chimeric (DEC) Cells in Duchenne Muscular Dystrophy Model. Biomedicines 2024, 12, 586. https://doi.org/10.3390/biomedicines12030586
Siemionow M, Budzynska K, Zalants K, Langa P, Brodowska S, Siemionow K, Heydemann A. Amelioration of Morphological Pathology in Cardiac, Respiratory, and Skeletal Muscles Following Intraosseous Administration of Human Dystrophin Expressing Chimeric (DEC) Cells in Duchenne Muscular Dystrophy Model. Biomedicines. 2024; 12(3):586. https://doi.org/10.3390/biomedicines12030586
Chicago/Turabian StyleSiemionow, Maria, Katarzyna Budzynska, Kristina Zalants, Paulina Langa, Sonia Brodowska, Krzysztof Siemionow, and Ahlke Heydemann. 2024. "Amelioration of Morphological Pathology in Cardiac, Respiratory, and Skeletal Muscles Following Intraosseous Administration of Human Dystrophin Expressing Chimeric (DEC) Cells in Duchenne Muscular Dystrophy Model" Biomedicines 12, no. 3: 586. https://doi.org/10.3390/biomedicines12030586
APA StyleSiemionow, M., Budzynska, K., Zalants, K., Langa, P., Brodowska, S., Siemionow, K., & Heydemann, A. (2024). Amelioration of Morphological Pathology in Cardiac, Respiratory, and Skeletal Muscles Following Intraosseous Administration of Human Dystrophin Expressing Chimeric (DEC) Cells in Duchenne Muscular Dystrophy Model. Biomedicines, 12(3), 586. https://doi.org/10.3390/biomedicines12030586