
Probiotics as Potential Therapeutic Agents: Safeguarding Skeletal Muscle against Alcohol-Induced Damage through the Gut–Liver–Muscle Axis

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
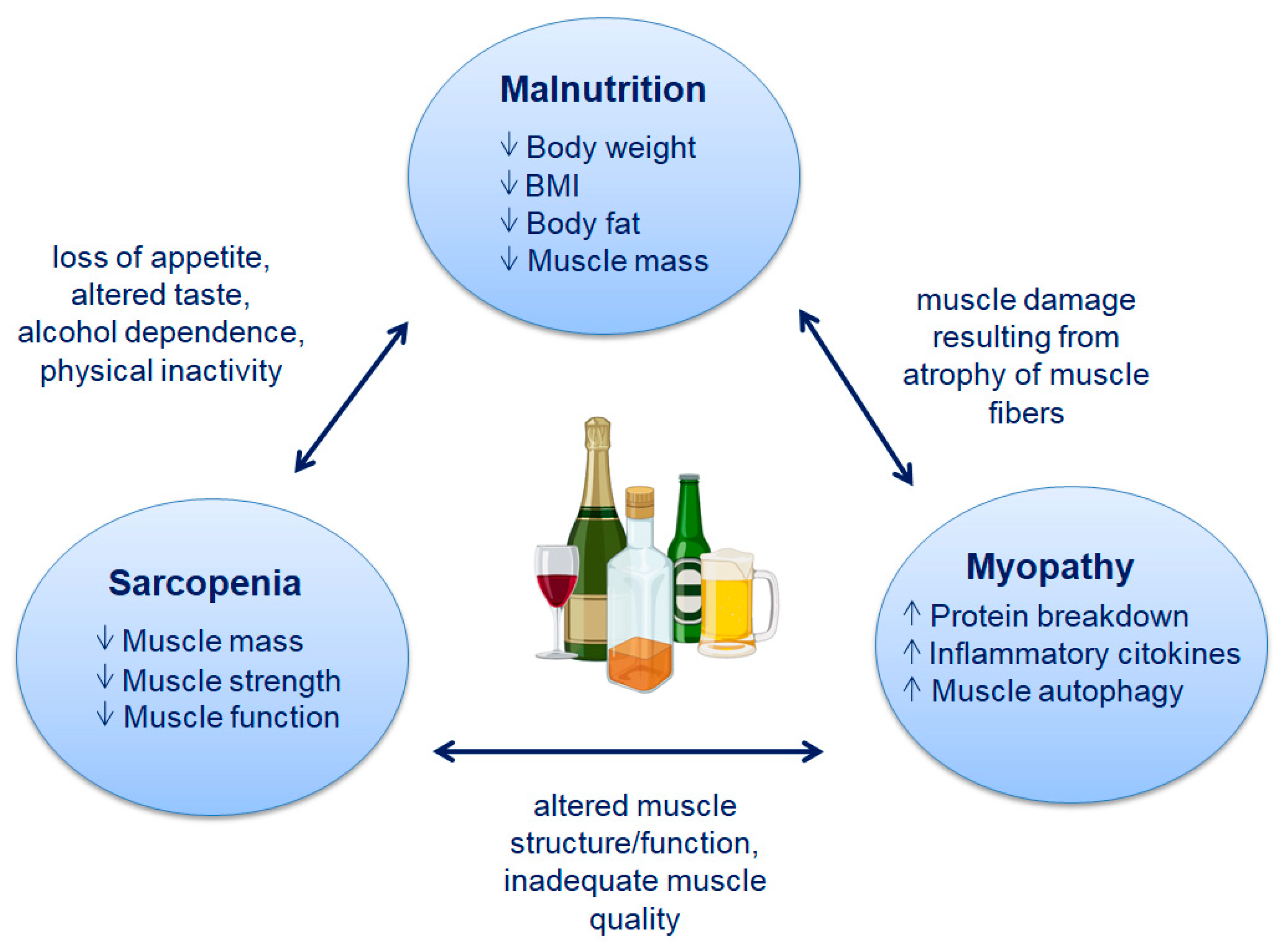
2. Malnutrition and Sarcopenia Associated with Alcoholic Liver Disease
2.1. Malnutrition
- (a)
- Reduction in body mass index (BMI) < 18.5 kg/m2 or underweight as defined by the World Health Organization (WHO);
- (b)
- Combined weight loss and BMI reduction (with age-dependent cut-off);
- (c)
- Sex-dependent reduction in fat mass index (FFMI);
- (d)
- A combination of the six defined parameters (low energy intake, weight loss, muscle mass loss, subcutaneous fat loss, fluid accumulation, and hand grip strength), at least two of which are met.
2.2. Sarcopenia
- (1)
- Criterion 1: Low muscle strength;
- (2)
- Criterion 2: Low muscle quantity or quality;
- (3)
- Criterion 3: Low physical performance.
3. Gut Microbiota and Hypothesis of “Gut-Liver-Muscle Axis” in ALD
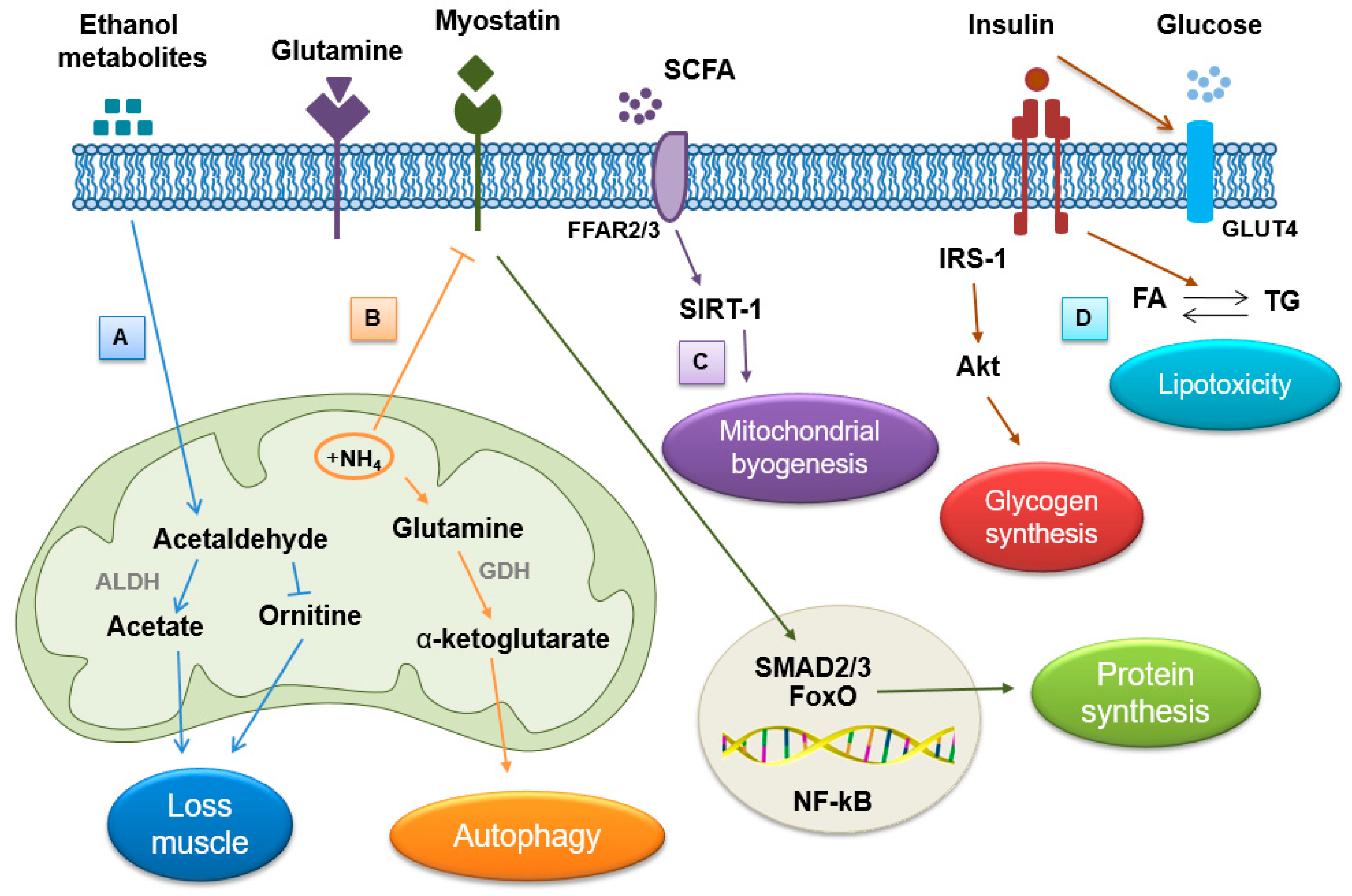
4. Effect of Alcohol on Skeletal Muscle
4.1. Hyperammonemia
4.2. Direct Effects of Ethanol and Its Metabolites
4.3. Endocrine Abnormality
4.4. Myosteatosis
4.5. Gut Dysfunction
5. Effects of Alcohol on the Gut Microbiota
5.1. Alteration of the Composition of the Intestinal Microbiota
5.2. Disfunction of the Intestinal Barrier
5.3. Role of Short Chain Fatty Acids (SCFAs)
5.4. Inflammation
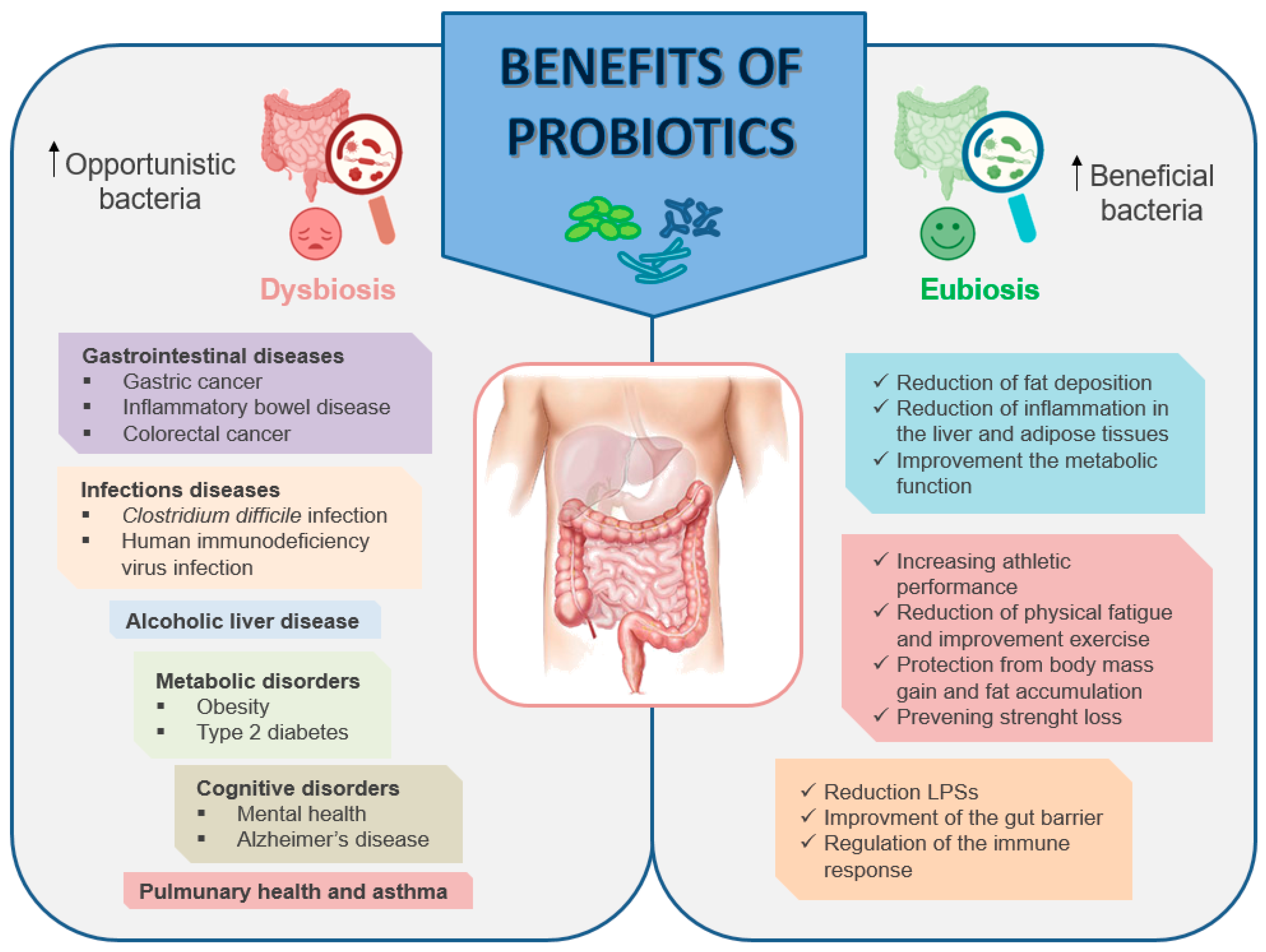
6. Modulation of the Gut Microbiota—Benefits of Probiotics
Mechanisms of Actions of Probiotics
7. Role of Probiotics in Ethanol-Induced Muscle Damage

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subjects (n) | Age | Physical Condition | Dosage | Effects | Reference |
|---|---|---|---|---|---|
| 46 (men and women) | 21–67 years | Moderate alcohol-associated hepatitis | Lactobacillus rhamnosus GG (LGG) for 6 months | Improvement in both liver injury and drinking | [166] |
| 33 men and 39 women | 23–63 years | Non-alcoholic fatty liver disease (NAFLD) | 300 g daily of probiotic or conventional yogurt for 8 weeks | Reduction of serum levels of alanine aminotransferase, aspartate aminotransferase, total cholesterol, and low-density lipoprotein cholesterol | [167] |
| 150 patients | ? | Alcoholic liver injury | Lactobaillus casei Shirota (LcS) (low-dose, 100 mL, and high-dose, 200 mL) for 60 days | Improving lipid metabolism and regulating intestinal flora disorders | [168] |
| 215 patients | ? | Cirrhosis | Probiotics VSL#3, 112.5 billion CFU containing 8 strains of bacteria | Improving nutritional status and reducing severity of liver disease | [169] |
| 66 men | ? | Alcoholic psychosis | Bifidobacterium bifidum and Lactobacillus plantarum 8PA3 for 5 days | Restoration of the bowel flora and great improvement in alcohol-induced liver injury | [170] |
| 10 men | 22.0 ± 2.4 years | Healthy resistance-trained | Bacillus coagulans BC30 + 20 g casein twice daily (500 M) | Increasing athletic performance, with an effect on peak power and fat mass | [171] |
| 30 men | 20–40 years | Healthy without professional exercise training | Lactobacillus plantarum TW10-HK (heat-killed TWK10) 3 × 1010 cells/day for 6 weeks. | Reduced physical fatigue and improved exercise endurance capacity and handrip strength | [172] |
| 17 men | 20.5 ± 0.8 years | Soldiers from an elite combat unit | Bacillus coagulans GBI-30 + Hydroxymethybutyrate calcium (CaHMB) 109 CFU/day for a total of 40 days | Attenuated inflammatory response to intense military training and maintaining muscle integrity | [173] |
| 20 men | 18–30 years | Healthy, non-obese | Streptococcus thermophilus, Lactobacillus acidophilus, Lactobacillus delbrueckii ssp. Bulgaricus, Lactobacillus paracasei, Lactobacillus plantarum, Bifidobacterium longum, Bifidobacterium infantis, and Bifisobacterium breve + high-fat and high-energy diet two sachets (450 billion bacteria per sachet) for 4 weeks | Protection from body mass gain and fat accumulation | [174] |
| 26 men and 12 women | 20–40 years | Healthy | Live Lactobacillus paracasei (L-PS23) or heat-killed Lactobacillus paracasei (HK-PS23) 2 × 1010 cells/day for six weeks | Preventing strength loss after muscle damage and improving blood muscle damage and inflammatory markers, with protective, accelerated recovery and anti-fatigue benefits | [175] |
| 18 (men and women) | 19–26 years | Amateur runners | Lactiplantibacillus plantarum TWK10 1010 CFU/day for 6 weeks | Increasing muscle mass and endurance performance | [176] |
8. Final Considerations
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 1,25(OH)D3 | 1,25 hydroxyvitamin D3 |
| 25(OH)D3 | 25 hydroxyvitamin D3 |
| 4E-BP1 | Eukaryotic translation initiation factor 4E (eIF4E)-binding protein 1 |
| ACTH | Adrenocorticotropic hormone |
| ADH | Alcohol dehydrogenase |
| Akt | Protein kinase B |
| ALD | Alcoholic liver disease |
| ALDH | Aldehyde dehydrogenase |
| AMPK | Adenosine monophosphate-activated protein kinase |
| AMPs | Antimicrobial peptides |
| ATP | Adenosine triphosphate |
| BIA | Bioelectrical impedance analysis |
| BMI | Body mass index |
| CaHMB | Hydroxymethybutyrate calcium |
| COX-2 | Cyclooxygenase 2 |
| CSA | Cross-section area |
| CSFAs | Short chain fatty acids |
| CT | Computer tomography |
| CYP | Cytochrome P450 |
| DCs | Dendritic cells |
| DG | Diacylglycerol |
| EFSA | European Food Safety Authority |
| eIF2 | Eukaryotic initiation factor 2 |
| EWGSOP | European Working Group on Sarcopenia in Older People |
| FA | Fatty acid |
| FDA | Food and Drug Administration |
| FFAs | Free fatty acids |
| FFAR-2 | Free fatty acid receptor 2 |
| FFAR-3 | Free fatty acid receptor 3 |
| FFMI | Fat mass index |
| FGF-21 | Fibroblast growth factor 21 |
| FSH | Follicle stimulating hormone |
| GALT | Gut-associated lymphoid tissue |
| GDH | Glutamate dehydrogenase |
| GLUT-4 | Glucose transporter 4 |
| GSK-3β | Glycogen synthase kinase-3 beta |
| HMW | High molecular weight |
| HPA | Hypothalamic–pituitary–adrenal axis |
| HPG | Hypothalamic–pituitary–gonadal axis |
| HPO | Hypothalamic–pituitary–ovarian axis |
| IBD | Inflammatory bowel disease |
| IEC | Intestinal epithelial cells |
| IFN-γ | Interferon gamma |
| IGF | Insulin-like growth factor |
| IL-1 | Interleukin-1 |
| IL-4 | Interleukin-4 |
| IL-5 | Interleukin-5 |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| IL-10 | Interleukin-10 |
| IL-15 | Interleukin-15 |
| IL-17 | Interleukin-17 |
| IMAT | Intermuscular adipose tissue |
| IMCL | Intramyocellular lipid |
| iNOS | Inducible nitric oxide synthase |
| LAB | Lactic acid bacteria |
| LC-CoA | Long-chain coenzyme A |
| LH | Luteinizing hormone |
| LPS | Lipopolysaccharide |
| LT | Liver transplantation |
| MAPK | Mitogen-activated protein kinase |
| MEF2A | Myocyte enhancer factor 2A |
| MRI | Magnetic resonance imaging |
| NAFLD | Non-alcoholic fatty liver disease |
| NF-kB | Nuclear factor kappa-light-chain-enhancer |
| NO | Nitric oxide |
| PGC-1α | Peroxisome proliferator-activated receptor γ coactivator 1α |
| PI3K | Phosphatidylinositol 3-kinase |
| PRRs | Pattern recognition receptors |
| ROS | Reactive oxygen species |
| sIgA | Secretory immunoglobulin A |
| SCFAS | Short chain fatty acids |
| SIRT-1 | NAD-dependent deacetylase sirtuin-1 |
| T2D | Type 2 diabetes |
| TCAs | Tricarboxylic acid intermediates |
| TG | Triglyceride |
| TGF-β1 | Transforming growth factor-1 beta |
| Th | T helper |
| TJ | Tight junctions |
| TLRs | Toll-like receptor |
| TNF- α | Tumor necrosis factor alpha |
| TW10-HK | Heat-killed TWK10 |
| UPP | Ubiquitin–preteasome pathway |
| UPS | Ubiquitin–preteasome system |
| WHO | World Health Organization |
| ZO-1 | Zona occludens-1 |
References
- World-Health-Organization. Global Status Report on Alcohol and Health 2018; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- Aslam, A.; Kwo, P.Y. Epidemiology and Disease Burden of Alcohol Associated Liver Disease. J. Clin. Exp. Hepatol. 2023, 13, 88–102. [Google Scholar] [CrossRef]
- NIAAA. What Is a Standard Drink? 2021. Available online: https://www.niaaa.nih.gov/ (accessed on 10 May 2021).
- Manolis, T.A.; Manolis, A.A.; Manolis, A.S. Cardiovascular effects of alcohol: A double-edged sword/how to remain at the nadir point of the J-Curve? Alcohol 2019, 76, 117–129. [Google Scholar] [CrossRef]
- Avila, M.A.; Dufour, J.F.; Gerbes, A.L.; Zoulim, F.; Bataller, R.; Burra, P.; Cortez-Pinto, H.; Gao, B.; Gilmore, I.; Mathurin, P.; et al. Recent advances in alcohol-related liver disease (ALD): Summary of a Gut round table meeting. Gut 2020, 69, 764–780. [Google Scholar] [CrossRef]
- Dasarathy, S. Consilience in sarcopenia of cirrhosis. J. Cachexia Sarcopenia Muscle 2012, 3, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Dubinkina, V.B.; Tyakht, A.V.; Odintsova, V.Y.; Yarygin, K.S.; Kovarsly, B.A.; Pavlenko, A.V.; Ischenko, D.S.; Popenko, A.S.; Alezeev, D.G.; Taraskina, A.Y.; et al. Links of gut microbiota composition with alcohol dependence syndrome and alcoholic liver disease. Microbiome 2017, 5, 141. [Google Scholar] [CrossRef]
- Thapaliya, S.; Runkana, A.; McMullen, M.R.; Nagy, L.E.; McDonald, C.; Naga Prasad, S.V.; Dasarathy, S. Alcohol-induced autophagy contributes to loss in skeletal muscle mass. Autophagy 2014, 10, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Llopis, M.; Cassard, A.M.; Wrzosek, L.; Boschat, L.; Bruneau, A.; Ferrere, G.; Puchois, V.; Martin, J.C.; Lepage, P.; Le Roy, T.; et al. Intestinal microbiota contributes to individual susceptibility to alcoholic liver disease. Gut 2016, 65, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, S.; Matamoros, S.; Cani, P.D.; Neyrinck, A.M.; Jamar, F.; Stärkel, P.; Windey, K.; Tremaroli, V.; Bäckhed, F.; Verbeke, K.; et al. Intestinal permeability, gut-bacterial dysbiosis, and behavioral markers of alcohol-dependence severity. Proc. Natl. Acad. Sci. USA 2014, 21, E4485–E4493. [Google Scholar] [CrossRef]
- Grosicki, G.J.; Fielding, R.A.; Lustgarten, M.S. Gut Microbiota Contribute to Age-Related Changes in Skeletal Muscle Size, Composition, and Function: Biological Basis for a Gut-Muscle Axis. Calcif. Tissue Int. 2018, 102, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Ticinesi, A.; Lauretani, F.; Milani, C.; Nouvenne, A.; Tana, C.; Del Rio, D.; Maggio, M.; Ventura, M.; Meschi, T. Aging Gut Microbiota at the Cross-Road between Nutrition, Physical Frailty, and Sarcopenia: Is There a Gut-Muscle Axis? Nutrients 2017, 30, 1303. [Google Scholar] [CrossRef] [PubMed]
- Lustgarten, M.S. The Role of the Gut Microbiome on Skeletal Muscle Mass and Physical Function: 2019 Update. Front. Physiol. 2019, 26, 1435. [Google Scholar] [CrossRef]
- Kamran, U.; Towey, J.; Khanna, A.; Chauhan, A.; Rajoriya, N.; Holt, A. Nutrition in alcohol-related liver disease: Physiopathology and management. World J. Gastroenterol. 2020, 14, 2916–2930. [Google Scholar] [CrossRef] [PubMed]
- Wing, S.S.; Lecker, S.H.; Jagoe, R.T. Proteolysis in illness-associated skeletal muscle atrophy: From pathways to networks. Crit. Rev. Clin. Lab. Sci. 2011, 48, 49–70. [Google Scholar] [CrossRef]
- Bhanji, R.A.; Moctezuma-Velazquez, C.; Duarte-Rojo, A.; Ebadi, M.; Ghosh, S.; Rose, C.; Montano-Loza, A.J. Myosteatosis and sarcopenia are associated with hepatic encephalopathy in patients with cirrhosis. Hepatol. Int. 2018, 12, 377–386. [Google Scholar] [CrossRef]
- Morley, J.E. Sarcopenia in the elderly. Fam. Pract. 2012, 29, i44–i48. [Google Scholar] [CrossRef]
- Jung, H.N.; Jung, C.H.; Hwang, Y.C. Sarcopenia in youth. Metabolism 2023, 144, 155557. [Google Scholar] [CrossRef]
- Santilli, V.; Bernetti, A.; Mangone, M.; Paoloni, M. Clinical definition of sarcopenia. Clin. Cases Miner. Bone Metab. 2014, 11, 177–180. [Google Scholar] [CrossRef]
- Nagy, H.; Veerapaneni, K.D. Myopathy. [Updated 2023 Aug 14]. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Steiner, J.L.; Lang, C.H. Dysregulation of skeletal muscle protein metabolism by alcohol. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E699–E712. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Cánoves, P.; Scheele, C.; Pedersen, B.K.; Serrano, A.L. Interleukin-6 myokine signaling in skeletal muscle: A double-edged sword? FEBS J. 2013, 280, 4131–4148. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, E.B.; Hamberg, O.; Quistorff, B.; Ott, P. Reduced mitochondrial adenosine triphosphate synthesis in skeletal muscle in patients with Child-Pugh class B and C cirrhosis. Hepatology 2001, 34, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.G.; Ferreira Martins, A.I.; Cunha, C.E.; Anastácio, L.R.; Lima, A.S.; Correia, M.I. Negative energy balance secondary to inadequate dietary intake of patients on the waiting list for liver transplantation. Nutrition 2013, 29, 1252–1258. [Google Scholar] [CrossRef] [PubMed]
- Cederholm, T.; Barazzoni, R.; Austin, P.; Ballmer, P.; Biolo, G.; Bischoff, S.C.; Compher, C.; Correia, I.; Higashiguchi, T.; Holst, M.; et al. ESPEN guidelines on definitions and terminology of clinical nutrition. Clin. Nutr. 2017, 36, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Cederholm, T.; Bosaeus, I.; Barazzoni, R.; Bauer, J.; Van Gossum, A.; Klek, S.; Muscaritoli, M.; Nyulasi, I.; Ockenga, J.; Schneider, S.M.; et al. Diagnostic criteria for malnutrition—An ESPEN Consensus Statement. Clin. Nutr. 2015, 34, 335–340. [Google Scholar] [CrossRef] [PubMed]
- White, J.V.; Guenter, P.; Jensen, G.; Malone, A.; Schofield, M.; Academy Malnutrition Work Group; A.S.P.E.N. Malnutrition Task Force; A.S.P.E.N. Board of Directors. Consensus statement: Academy of Nutrition and Dietetics and American Society for Parenteral and Enteral Nutrition: Characteristics recommended for the identification and documentation of adult malnutrition (undernutrition). JPEN J. Parenter. Enteral Nutr. 2012, 36, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Maharshi, S.; Sharma, B.C.; Srivastava, S. Malnutrition in cirrhosis increases morbidity and mortality. J. Gastroenterol. Hepatol. 2015, 30, 1507–1513. [Google Scholar] [CrossRef] [PubMed]
- Plauth, M.; Bernal, W.; Dasarathy, S.; Merli, M.; Plank, L.D.; Schütz, T.; Bischoff, S.C. ESPEN guideline on clinical nutrition in liver disease. Clin. Nutr. 2019, 38, 485–521. [Google Scholar] [CrossRef] [PubMed]
- Buchard, B.; Boirie, Y.; Cassagnes, L.; Lamblin, G.; Coilly, A.; Abergel, A. Assessment of Malnutrition, Sarcopenia and Frailty in Patients with Cirrhosis: Which Tools Should We Use in Clinical Practice? Nutrients 2020, 12, 186. [Google Scholar] [CrossRef]
- Tandon, P.; Low, G.; Mourtzakis, M.; Zenith, L.; Myers, R.P.; Abraldes, J.G.; Shaheen, A.A.; Qamar, H.; Mansoor, N.; Carbonneau, M.; et al. A Model to Identify Sarcopenia in Patients with Cirrhosis. Clin. Gastroenterol. Hepatol. 2016, 14, 1473–1480.e3. [Google Scholar] [CrossRef]
- Sinclair, M.; Hoermann, R.; Peterson, A.; Testro, A.; Angus, P.W.; Hey, P.; Chapman, B.; Gow, P.J. Use of Dual X-ray Absorptiometry in men with advanced cirrhosis to predict sarcopenia-associated mortality risk. Liver Int. 2019, 39, 1089–1097. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Writing Group for the European Working Group on Sarcopenia in Older People 2 (EWGSOP2), and the Extended Group for EWGSOP2. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Baeyens, J.P.; Bauer, J.M.; Boirie, Y.; Cederholm, T.; Landi, F.; Martin, F.C.; Michel, J.P.; Rolland, Y.; Schneider, S.M.; et al. European Working Group on Sarcopenia in Older People. Sarcopenia: European consensus on definition and diagnosis: Report of the European Working Group on Sarcopenia in Older People. Age Ageing 2010, 39, 412–423. [Google Scholar] [CrossRef]
- Schaap, L.A.; Koster, A.; Visser, M. Adiposity, muscle mass, and muscle strength in relation to functional decline in older persons. Epidemiol. Rev. 2013, 35, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Masanés, F.; Rojano, I.; Luque, X.; Salvà, A.; Serra-Rexach, J.A.; Artaza, I.; Formiga, F.; Cuesta, F.; López Soto, A.; Ruiz, D.; et al. Cut-off Points for Muscle Mass—Not Grip Strength or Gait Speed—Determine Variations in Sarcopenia Prevalence. J. Nutr. Health Aging 2017, 21, 825–829. [Google Scholar] [CrossRef]
- Ebadi, M.; Bhanji, R.A.; Mazurak, V.C.; Montano-Loza, A.J. Sarcopenia in cirrhosis: From pathogenesis to interventions. J. Gastroenterol. 2019, 54, 845–859. [Google Scholar] [CrossRef]
- Carey, E.J.; Lai, J.C.; Wang, C.W.; Dasarathy, S.; Lobach, I.; Montano-Loza, A.J.; Dunn, M.A. Fitness, Life Enhancement, and Exercise in Liver Transplantation Consortium. A multicenter study to define sarcopenia in patients with end-stage liver disease. Liver Transpl. 2017, 23, 625–633. [Google Scholar] [CrossRef]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 20, 1647–1651. [Google Scholar] [CrossRef]
- Fulde, M.; Hornef, M.W. Maturation of the enteric mucosal innate immune system during the postnatal period. Immunol. Rev. 2014, 260, 21–34. [Google Scholar] [CrossRef]
- Kamada, N.; Chen, G.Y.; Inohara, N.; Nunez, G. Control of pathogens and pathobionts by the gut microbiota. Nat. Immunol. 2013, 14, 685–690. [Google Scholar] [CrossRef]
- Ijssennagger, N.; Belzer, C.; Hooiveld, G.J.; Dekker, J.; van Mil, S.W.; Müller, M.; Kleerebezem, M.; van der Meer, R. Gut microbiota facilitates dietary heme-induced epithelial hyperproliferation by opening the mucus barrier in colon. Proc. Natl. Acad. Sci. USA 2015, 11, 10038–10043. [Google Scholar] [CrossRef]
- Reinhardt, C.; Bergentall, M.; Greiner, T.U.; Schaffner, F.; Ostergren-Lundén, G.; Petersen, L.C.; Ruf, W.; Bäckhed, F. Tissue factor and PAR1 promote microbiota-induced intestinal vascular remodelling. Nature 2012, 11, 627–631. [Google Scholar] [CrossRef]
- Neuman, H.; Debelius, J.W.; Knight, R.; Koren, O. Microbial endocrinology: The interplay between the microbiota and the endocrine system. FEMS Microbiol. Rev. 2015, 39, 509–521. [Google Scholar] [CrossRef]
- Cho, I.; Yamanishi, S.; Cox, L.; Methé, B.A.; Zavadil, J.; Li, K.; Gao, Z.; Mahana, D.; Raju, K.; Teitler, I.; et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 2012, 30, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J.S. Alcohol, liver disease and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Racanelli, V.; Rehermann, B. The liver as an immunological organ. Hepatology 2006, 43, S54–S62. [Google Scholar] [CrossRef] [PubMed]
- Bode, C.; Bode, J.C. Activation of the innate immune system and alcoholic liver disease: Effects of ethanol per se or increased intestinal translocation of bacterial toxins induced by ethanol? Res. Exp. Clin. Alcohol. 2005, 29, 166S–171S. [Google Scholar] [CrossRef] [PubMed]
- Balmer, M.L.; Slack, E.; de Gottardi, A.; Lawson, M.A.; Hapfelmeier, S.; Miele, L.; Grieco, A.; Van Vlierberghe, H.; Fahrner, R.; Patuto, N.; et al. The liver can act as a firewall mediating mutualism between the host and its intestinal commensal microbiota. Sci. Transl. Med. 2014, 6, 237ra66. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Schnabl, B.; Brenner, D.A. Interactions between the intestinal microbiome and liver diseases. Gastroenterology 2014, 146, 1513–1524. [Google Scholar] [CrossRef] [PubMed]
- Rao, R. Endotoxemia and gut barrier dysfunction in alcoholic liver disease. Hepatology 2009, 50, 638–644. [Google Scholar] [CrossRef]
- Wang, Y.; Tong, J.; Chang, B.; Wang, B.; Zhang, D.; Wang, B. Effects of alcohol on intestinal epithelial barrier permeability and expression of tight junction-associated proteins. Mol. Med. Rep. 2014, 9, 2352–2356. [Google Scholar] [CrossRef]
- Hartmann, P.; Seebauer, C.T.; Schnabl, B. Alcoholic liver disease: The gut microbiome and liver cross talk. Alcohol. Clin. Exp. Res. 2015, 39, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Muniz, L.R.; Knosp, C.; Yeretssian, G. Intestinal antimicrobial peptides during homeostasis, infection, and disease. Front. Immunol. 2012, 3, 310. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.K.; Meena, A.S.; Rao, V.; Rao, R.G.; Balazs, L.; Rao, R. Human Defensin-5 Blocks Ethanol and Colitis-Induced Dysbiosis, Tight Junction Disruption and Inflammation in Mouse Intestine. Sci. Rep. 2018, 2, 16241. [Google Scholar] [CrossRef] [PubMed]
- Bellot, P.; Francés, R.; Such, J. Pathological bacterial translocation in cirrhosis: Pathophysiology, diagnosis and clinical implications. Liver Int. 2013, 33, 31–39. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef]
- Kuo, H.K.; Liao, K.C.; Leveille, S.G.; Bean, J.F.; Yen, C.J.; Chen, J.H.; Yu, Y.H.; Tai, T.Y. Relationship of homocysteine levels to quadriceps strength, gait speed, and late-life disability in older adults. J. Gerontol. A Biol. Sci. Med. Sci. 2007, 62, 434–439. [Google Scholar] [CrossRef]
- Lin, R.; Liu, W.; Piao, M.; Zhu, H. A review of the relationship between the gut microbiota and amino acid metabolism. Amino Acids 2017, 49, 2083–2090. [Google Scholar] [CrossRef]
- Dukes, A.; Davis, C.; El Refaey, M.; Upadhyay, S.; Mork, S.; Arounleut, P.; Johnson, M.H.; Hill, W.D.; Isales, C.M.; Hamrick, M.W. The aromatic amino acid tryptophan stimulates skeletal muscle IGF1/p70s6k/mTor signaling in vivo and the expression of myogenic genes in vitro. Nutrition 2015, 31, 1018–1024. [Google Scholar] [CrossRef]
- Elkina, Y.; Von Haehling, S.; Anker, S.D.; Springer, J. The role of myostatin in muscle wasting: An overview. J. Cachexia Sarcopenia Muscle 2011, 2, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Beulens, J.W.; van Loon, L.J.; Kok, F.J.; Pelsers, M.; Bobbert, T.; Spranger, J.; Helander, A.; Hendriks, H.F. The effect of moderate alcohol consumption on adiponectin oligomers and muscle oxidative capacity: A human intervention study. Diabetologia 2007, 50, 1388–1392. [Google Scholar] [CrossRef] [PubMed]
- González-Reimers, E.; Fernández-Rodríguez, C.M.; Santolaria-Fernández, F.; de la Vega-Prieto, M.J.; Martín-González, C.; Gómez-Rodríguez, M.Á.; Alemán-Valls, M.R.; Rodríguez-Gaspar, M. Interleukin-15 and other myokines in chronic alcoholics. Alcohol. Alcohol. 2011, 46, 529–533. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shenkman, B.S.; Zinovyeva, O.E.; Belova, S.P.; Mirzoev, T.M.; Vilchinskaya, N.A.; Vikhlyantsev, I.M.; Ulanova, A.D.; Turtikova, O.V.; Samkhaeva, N.D.; Parfenov, V.A.; et al. Cellular and molecular signatures of alcohol-induced myopathy in women. Am. J. Physiol. Endocrinol. Metab. 2019, 1, E967–E976. [Google Scholar] [CrossRef] [PubMed]
- Maneesh, M.; Dutta, S.; Chakrabarti, A.; Vasudevan, D.M. Alcohol abuse-duration dependent decrease in plasma testosterone and antioxidants in males. Indian. J. Physiol. Pharmacol. 2006, 50, 291–296. [Google Scholar] [PubMed]
- Smiles, W.J.; Parr, E.B.; Coffey, V.G.; Lacham-Kaplan, O.; Hawley, J.A.; Camera, D.M. Protein coingestion with alcohol following strenuous exercise attenuates alcohol-induced intramyocellular apoptosis and inhibition of autophagy. Am. J. Physiol. Endocrinol. Metab. 2016, 1, E836–E849. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Thapaliya, S.; Runkana, A.; Yang, Y.; Tsien, C.; Mohan, M.L.; Narayanan, A.; Eghtesad, B.; Mozdziak, P.E.; McDonald, C.; et al. Hyperammonemia in cirrhosis induces transcriptional regulation of myostatin by an NF-κB-mediated mechanism. Proc. Natl. Acad. Sci. USA 2013, 5, 18162–18167. [Google Scholar] [CrossRef]
- Dasarathy, S.; Hatzoglou, M. Hyperammonemia and proteostasis in cirrhosis. Curr. Opin. Clin. Nutr. Metabol. Care 2018, 21, 30–36. [Google Scholar] [CrossRef]
- Davuluri, G.; Allawy, A.; Thapaliya, S.; Rennison, J.H.; Singh, D.; Kumar, A.; Sandlers, Y.; Van Wagoner, D.R.; Flask, C.A.; Hoppel, C.; et al. Hyperammonaemia-induced skeletal muscle mitochondrial dysfunction results in cataplerosis and oxidative stress. J. Physiol. 2016, 15, 7341–7360. [Google Scholar] [CrossRef]
- Owen, O.E.; Kalhan, S.C.; Hanson, R.W. The key role of anaplerosis and cataplerosis for citric acid cycle function. J. Biol. Chem. 2002, 23, 30409–30412. [Google Scholar] [CrossRef]
- Ábrigo, J.; Elorza, A.A.; Riedel, C.A.; Vilos, C.; Simon, F.; Cabrera, D.; Estrada, L.; Cabello-Verrugio, C. Role of Oxidative Stress as Key Regulator of Muscle Wasting during Cachexia. Oxid. Med. Cell Longev. 2018, 28, 2063179. [Google Scholar] [CrossRef]
- Bonet-Ponce, L.; Saez-Atienzar, S.; da Casa, C.; Flores-Bellver, M.; Barcia, J.M.; Sancho-Pelluz, J.; Romero, F.J.; Jordan, J.; Galindo, M.F. On the mechanism underlying ethanol-induced mitochondrial dynamic disruption and autophagy response. Biochim. Biophys. Acta 2015, 1852, 1400–1409. [Google Scholar] [CrossRef]
- Holmuhamedov, E.L.; Czerny, C.; Beeson, C.C.; Lemasters, J.J. Ethanol suppresses ureagenesis in rat hepatocytes: Role of acetaldehyde. J. Biol. Chem. 2012, 2, 7692–7700. [Google Scholar] [CrossRef]
- Tsien, C.; Davuluri, G.; Singh, D.; Allawy, A.; Ten Have, G.A.; Thapaliya, S.; Schulze, J.M.; Barnes, D.; McCullough, A.J.; Engelen, M.P.; et al. Metabolic and molecular responses to leucine-enriched branched chain amino acid supplementation in the skeletal muscle of alcoholic cirrhosis. Hepatology 2015, 61, 2018–2029. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.L.; Lang, C.H. Alcohol impairs skeletal muscle protein synthesis and mTOR signaling in a time-dependent manner following electrically stimulated muscle contraction. J. Appl. Physiol. 2014, 117, 1170–1179. [Google Scholar] [CrossRef] [PubMed]
- Rachdaoui, N.; Sarkar, D.K. Effects of alcohol on the endocrine system. Endocrinol. Metab. Clin. N. Am. 2013, 42, 593–615. [Google Scholar] [CrossRef] [PubMed]
- Patrelli, T.S.; Gizzo, S.; Franchi, P.; Franchi, L.; Volpi, L.; Esposito, F.; Pedrazzi, G.; Degli Antoni, A.; Bacchi Modena, A. Impact of Highly Active Antiretroviral Therapy on the Natural History of Cervical Precancerous Lesions: A 17-Year Institutional Longitudinal Cohort Study. Repr. Sci. 2014, 21, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Duca, Y.; Aversa, A.; Condorelli, R.A.; Calogero, A.E.; La Vignera, S. Substance Abuse and Male Hypogonadism. J. Clin. Med. 2019, 22, 732. [Google Scholar] [CrossRef] [PubMed]
- Svartberg, J.; Midtby, M.; Bønaa, K.H.; Sundsfjord, J.; Joakimsen, R.M.; Jorde, R. The associations of age, lifestyle factors and chronic disease with testosterone in men: The Tromsø Study. Eur. J. Endocrinol. 2003, 149, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.K.; Swan, S.; Jørgensen, N.; Toppari, J.; Redmon, B.; Punab, M.; Drobnis, E.Z.; Haugen, T.B.; Zilaitiene, B.; Sparks, A.E.; et al. Alcohol and male reproductive health: A cross-sectional study of 8344 healthy men from Europe and the USA. Hum. Reprod. 2014, 29, 1801–1809. [Google Scholar] [CrossRef] [PubMed]
- Badrick, E.; Bobak, M.; Britton, A.; Kirschbaum, C.; Marmot, M.; Kumari, M. The relationship between alcohol consumption and cortisol secretion in an aging cohort. J. Clin. Endocrinol. Metab. 2008, 93, 750–757. [Google Scholar] [CrossRef]
- Glass, D.J. Signaling pathways perturbing muscle mass. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 225–229. [Google Scholar] [CrossRef]
- Kvorning, T.; Andersen, M.; Brixen, K.; Madsen, K. Suppression of endogenous testosterone production attenuates the response to strength training: A randomized, placebo-controlled, and blinded intervention study. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E1325–E1332. [Google Scholar] [CrossRef]
- Liu, W.; Thomas, S.G.; Asa, S.L.; Gonzalez-Cadavid, N.; Bhasin, S.; Ezzat, S. Myostatin is a skeletal muscle target of growth hormone anabolic action. J. Clin. Endocrinol. Metab. 2003, 88, 5490–5496. [Google Scholar] [CrossRef]
- Lakshman, K.M.; Bhasin, S.; Corcoran, C.; Collins-Racie, L.A.; Tchistiakova, L.; Forlow, S.B.; St Ledger, K.; Burczynski, M.E.; Dorner, A.J.; Lavallie, E.R. Measurement of myostatin concentrations in human serum: Circulating concentrations in young and older men and effects of testosterone administration. Mol. Cell Endocrinol. 2009, 10, 26–32. [Google Scholar] [CrossRef] [PubMed]
- García, P.S.; Cabbabe, A.; Kambadur, R.; Nicholas, G.; Csete, M. Elevated Myostatin Levels in Patients with Liver Disease. Anesth. Analg. 2010, 111, 707–709. [Google Scholar] [CrossRef] [PubMed]
- Wilkes, J.J.; Lloyd, D.J.; Gekakis, N. Loss-of-Function Mutation in Myostatin Reduces Tumor Necrosis Factor α Production and Protects Liver Against Obesity-Induced Insulin Resistance. Diabetes 2009, 58, 1133–1143. [Google Scholar] [CrossRef] [PubMed]
- Delogu, W.; Caligiuri, A.; Provenzano, A.; Rosso, C.; Bugianesi, E.; Coratti, A.; Macias-Barragan, J.; Galastri, S.; Di Maira, G.; Marra, F. Myostatin regulates the fibrogenic phenotype of hepatic stellate cells via c-jun N-terminal kinase activation. Dig. Liver Dis. 2019, 51, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- Manari, A.P.; Preedy, V.R.; Peters, T.J. Nutritional intake of hazardous drinkers and dependent alcoholics in the UK. Addict. Biol. 2003, 8, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Shankar, K.; Liu, X.; Singhal, R.; Chen, J.R.; Nagarajan, S.; Badger, T.M.; Ronis, M.J. Chronic ethanol consumption leads to disruption of vitamin D3 homeostasis associated with induction of renal 1,25 dihydroxyvitamin D3-24-hydroxylase (CYP24A1). Endocrinology 2008, 149, 1748–1756. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Lester, R.; Akima, H.; Gorgey, A. Quantification of intermuscular and intramuscular adipose tissue using magnetic resonance imaging after neurodegenerative disorders. Neural Regen. Res. 2017, 12, 2100. [Google Scholar]
- Perdomo, G.; Commerford, S.R.; Richard, A.M.; Adams, S.H.; Corkey, B.E.; O’Doherty, R.M.; Brown, N.F. Increased beta-oxidation in muscle cells enhances insulin-stimulated glucose metabolism and protects against fatty acid-induced insulin resistance despite intramyocellular lipid accumulation. J. Biol. Chem. 2004, 25, 27177–27186. [Google Scholar] [CrossRef]
- Gueugneau, M.; Coudy-Gandilhon, C.; Théron, L.; Meunier, B.; Barboiron, C.; Combaret, L.; Taillandier, D.; Polge, C.; Attaix, D.; Picard, B.; et al. Skeletal muscle lipid content and oxidative activity in relation to muscle fiber type in aging and metabolic syndrome. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Z.; Zhao, M.; Nie, Y.; Liu, P.; Zhu, Y.; Zhang, X. Skeletal muscle lipid droplets and the athlete’s paradox. Cells 2019, 8, 249. [Google Scholar] [CrossRef] [PubMed]
- Gumucio, J.P.; Qasawa, A.H.; Ferrara, P.J.; Malik, A.N.; Funai, K.; McDonagh, B.; Mendias, C.L. Reduced mitochondrial lipid oxidation leads to fat accumulation in myosteatosis. FASEB J. 2019, 33, 7863–7881. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, L.P.; Fisher, J.S. Skeletal Muscle Insulin Resistance: Roles of Fatty Acid Metabolism and Activity. Phys. Ther. 2008, 88, 1279–1296. [Google Scholar] [CrossRef] [PubMed]
- Coleman, S.K.; Rebalka, I.A.; D’Souza, D.M.; Hawke, T.J. Skeletal muscle as a therapeutic target for delaying complications of type 1 diabetes. World Diabetes J. 2015, 6, 1323–1336. [Google Scholar] [CrossRef] [PubMed]
- Straczkowski, M.; Kowalska, I.; Baranowski, M.; Nikolahus, A.; Otziomek, E.; Zabielski, P.; Adamska, A.; Blachnio, A.; Gorski, J.; Gorska, M. Increased ceramide level in skeletal muscle in men at risk of developing type 2 diabetes. Diabetologia 2007, 50, 2366–2373. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Kahn, C.R. Insulin signaling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Engen, P.A.; Green, S.J.; Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. The Gastrointestinal Microbiome: Alcohol Effects on the Composition of Intestinal Microbiota. Alcohol. Res. 2015, 37, 223–236. [Google Scholar]
- Dunagan, M.; Chaudhry, K.; Samak, G.; Rao, R.K. Acetaldehyde disrupts tight junctions in Caco-2 cell monolayers by a protein phosphatase 2A-dependent mechanism. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1356–G1364. [Google Scholar] [CrossRef]
- Bala, S.; Marcos, M.; Gattu, A. Acute binge drinking increases serum endotoxin and bacterial DNA levels in healthy individuals. PLoS ONE 2014, 9, e96864. [Google Scholar] [CrossRef]
- Gordon, B.S.; Kelleher, A.R.; Kimball, S.R. Regulation of muscle protein synthesis and the effects of catabolic states. Int. J. Biochem. Cell Biol. 2013, 45, 2147–2157. [Google Scholar] [CrossRef]
- Lowe, P.P.; Gyongyosi, B.; Satishchandran, A.; Iracheta-Vellve, A.; Cho, Y.; Ambade, A.; Szabo, G. Reduced gut microbiome protects from alcohol-induced neuroinflammation and alters intestinal and brain inflammasome expression. J. Neuroinflamm. 2018, 27, 298. [Google Scholar] [CrossRef]
- Xiao, H.W.; Ge, C.; Feng, G.X.; Li, Y.; Luo, D.; Dong, J.L.; Li, H.; Wang, H.; Cui, M.; Fan, S.J. Gut microbiota modulates alcohol withdrawal-induced anxiety in mice. Toxicol. Lett. 2018, 1, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Posteraro, B.; Paroni Sterbini, F.; Petito, V.; Rocca, S.; Cubeddu, T.; Graziani, C.; Arena, V.; Vassallo, G.A.; Mosoni, C.; Lopetuso, L.; et al. Liver Injury, Endotoxemia, and Their Relationship to Intestinal Microbiota Composition in Alcohol-Preferring Rats. Alcohol. Clin. Exp. Res. 2018, 42, 2313–2325. [Google Scholar] [CrossRef] [PubMed]
- Bjørkhaug, S.T.; Aanes, H.; Neupane, S.P.; Bramness, J.G.; Malvik, S.; Henriksen, C.; Skar, V.; Medhus, A.W.; Valeur, J. Characterization of gut microbiota composition and functions in patients with chronic alcohol overconsumption. Gut Microbes 2019, 10, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Mutlu, E.A.; Gillevet, P.M.; Rangwala, H.; Sikaroodi, M.; Naqvi, A.; Engen, P.A.; Kwasny, M.; Lau, C.K.; Keshavarzian, A. Colonic microbiome is altered in alcoholism. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 1, G966–G978. [Google Scholar] [CrossRef] [PubMed]
- Fucarino, A.; Burgio, S.; Paladino, L.; Caruso Bavisotto, C.; Pitruzzella, A.; Bucchieri, F.; Cappello, F. The Microbiota Is Not an Organ: Introducing the Muco-Microbiotic Layer as a Novel Morphofunctional Structure. Anatomia 2022, 1, 186–203. [Google Scholar] [CrossRef]
- Cappello, F.; Saguto, D.; Burgio, S.; Paladino, L.; Bucchieri, F. Does Intestine Morphology Still Have Secrets to Reveal? A Proposal about the “Ghost” Layer of the Bowel. Appl. Biosci. 2022, 1, 95–100. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Bishehsari, F.; Magno, E.; Swanson, G.; Desai, V.; Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. Alcohol and Gut-Derived Inflammation. Alcohol. Res. 2017, 38, 163–171. [Google Scholar]
- Liu, Y.; Yu, X.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. The role of MUC2 mucin in intestinal homeostasis and the impact of dietary components on MUC2 expression. Int. J. Biol. Macromol. 2020, 1, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Grewal, R.K.; Mahmood, A. Ethanol induced changes in glycosylation of mucins in rat intestine. Ann. Gastroenterol. 2009, 22, 178–183. [Google Scholar]
- Elamin, E.E.; Masclee, A.A.; Dekker, J.; Jonkers, D.M. Ethanol metabolism and its effects on the intestinal epithelial barrier. Nutr. Rev. 2013, 71, 483–499. [Google Scholar] [CrossRef]
- Chiang, C.P.; Wu, C.W.; Lee, S.P.; Ho, J.L.; Lee, S.L.; Nieh, S.; Yin, S.J. Expression pattern, ethanol-metabolizing activities, and cellular localization of alcohol and aldehyde dehydrogenases in human small intestine. Alcohol. Clin. Exp. Res. 2012, 36, 2047–2058. [Google Scholar] [CrossRef] [PubMed]
- Ceni, E.; Mello, T.; Galli, A. Pathogenesis of alcoholic liver disease: Role of oxidative metabolism. World J. Gastroenterol. 2014, 20, 17756–17772. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.; Mach, N. The crosstalk between the gut microbiota and mitochondria during exercise. Front. Physiol. 2017, 8, 319. [Google Scholar] [CrossRef] [PubMed]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef]
- Kimura, I.; Inoue, D.; Hirano, K.; Tsujimoto, G. The SCFA receptor GPR43 and energy metabolism. Front. Endocrinol. 2014, 5, 85. [Google Scholar] [CrossRef]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Mottawea, W.; Chiang, C.K.; Mühlbauer, M.; Starr, A.E.; Butcher, J.; Abujamel, T.; Deeke, S.A.; Brandel, A.; Zhou, H.; Shokralla, S.; et al. Altered intestinal microbiota-host mitochondria crosstalk in new onset Crohn’s disease. Nat. Commun. 2016, 23, 13419. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, S.; Kim, H.; Garcia-Perez, I.; Reza, M.M.; Martin, K.A.; Kundu, P.; Cox, L.M.; Selkrig, J.; Posma, J.M.; Zhang, H.; et al. The gut microbiota influences skeletal muscle mass and function in mice. Sci. Transl. Med. 2019, 24, eaan5662. [Google Scholar] [CrossRef]
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panourgia, M.P.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and anti-inflammaging: A systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev. 2007, 128, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Hussey, S.E.; Sanchez-Avila, A.; Tantiwong, P.; Musi, N. Effect of lipopolysaccharide on inflammation and insulin action in human muscle. PLoS ONE 2013, 21, e63983. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Malhotra, S.; Kumar, A. Nuclear factor-kappa B signaling in skeletal muscle atrophy. J. Mol. Med. 2008, 86, 1113–1126. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, S.; de Timary, P.; Delzenne, N.M.; Starkel, P. The link between inflammation, bugs, the intestine and the brain in alcohol dependence. Transl. Psychiatry 2017, 7, e1048. [Google Scholar] [CrossRef]
- Webster, J.M.; Kempen, L.J.A.P.; Hardy, R.S.; Langen, R.C.J. Inflammation and skeletal muscle wasting during cachexia. Front. Physiol. 2020, 11, 597675. [Google Scholar] [CrossRef]
- Li, Y.P.; Chen, Y.; John, J.; Moylan, J.; Jin, B.; Mann, D.L.; Reid, M.B. TNF-alpha acts via p38 MAPK to stimulate expression of the ubiquitin ligase atrogin1/MAFbx in skeletal muscle. FASEB J. 2005, 19, 362–370. [Google Scholar] [CrossRef]
- Lee, J.S.; Oh, T.Y.; Kim, Y.K.; Baik, J.H.; So, S.; Hahm, K.B.; Surh, Y.J. Protective effects of green tea polyphenol extracts against ethanol-induced gastric mucosal damages in rats: Stress-responsive transcription factors and MAP kinases as potential targets. Mutat. Res. 2005, 11, 214–224. [Google Scholar] [CrossRef]
- Na, H.K.; Lee, J.Y. Molecular basis of alcohol-related gastric and colon cancer. Int. J. Mol. Sci. 2017, 18, 1116. [Google Scholar] [CrossRef]
- Rao, P.S.; Kumar, S. Chronic effects of ethanol and/or Darunavir/Ritonavir on u937 monocytic cells: Regulation of cytochrome p450 and antioxidant enzymes, oxidative stress, and cytotoxicity. Alcohol. Clin. Exp. Res. 2016, 40, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Bluemel, S.; Williams, B.; Knight, R.; Schnabl, B. Precision medicine in alcoholic and nonalcoholic fatty liver disease via modulating the gut microbiota. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G1018–G1036. [Google Scholar] [CrossRef] [PubMed]
- Metchnikoff, E.; Mitchell, P. The Prolongation of Life: Optimistic Studies; W. Heinemann: London, UK; G.P. Putnam’s Sons: New York, NY, USA, 1907. [Google Scholar]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front. Immunol. 2021, 26, 578386. [Google Scholar] [CrossRef] [PubMed]
- Hempel, S.; Newberry, S.J.; Maher, A.R.; Wang, Z.; Miles, J.N.; Shanman, R.; Johnsen, B.; Shekelle, P.G. Probiotics for the prevention and treatment of antibiotic-associated diarrhea: A systematic review and meta-analysis. JAMA 2012, 307, 1959–1969. [Google Scholar] [PubMed]
- Plaza-Díaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Immune-mediated mechanisms of action of probiotics and synbiotics in treating pediatric intestinal diseases. Nutrients 2018, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Berings, M.; Karaaslan, C.; Altunbulakli, C.; Gevaert, P.; Akdis, M.; Bachert, C.; Akdis, C.A. Advances and highlights in allergen immunotherapy: On the way to sustained clinical and immunologic tolerance. J. Allergy Clin. Immunol. 2017, 140, 1250–1267. [Google Scholar] [CrossRef] [PubMed]
- Rather, I.A.; Bajpai, V.K.; Kumar, S.; Lim, J.; Paek, W.K.; Park, Y.H. Probiotics and atopic dermatitis: An overview. Front. Microbiol. 2016, 7, 507. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Guidance on the scientific requirements for health claims related to the immune system, the gastrointestinal tract and defence against pathogenic microorganisms. EFSA J. 2016, 14, 4369. [Google Scholar]
- FDA Dietary Supplements: New Dietary Ingredient Notifications and Related Issues: Guidance for Industry, August 2016. Available online: http://www.fda.gov/downloads/Food/GuidanceRegulation/GuidanceDocumentsRegulatoryInformation/UCM515733.pdf (accessed on 10 October 2017).
- Government of Canada Health Canada. Available online: https://inspection.canada.ca/food-labels/labelling/industry/health-claims-on-food-labels/eng/1392834838383/1392834887794?chap=10 (accessed on 31 October 2019).
- Hempel, S.; Newberry, S.; Ruelaz, A.; Wang, Z.; Miles, J.N.; Suttorp, M.J.; Johnsen, B.; Shanman, R.; Slusser, W.; Fu, N.; et al. Safety of probiotics used to reduce risk and prevent or treat disease. Evid. Rep. Technol. Assess. (Full Rep.) 2011, 200, 1–645. [Google Scholar]
- Sanders, M.E.; Akkermans, L.M.; Haller, D.; Hammerman, C.; Heimbach, J.; Hormannsperger, G.; Huys, G. Safety assessment of probiotics for human use. Gut Microbes 2010, 1, 164–185. [Google Scholar] [CrossRef]
- Sheyholislami, H.; Connor, K.L. Are probiotics and prebiotics safe for use during pregnancy and lactation? A systematic review and meta-analysis. Nutrients 2021, 13, 2382. [Google Scholar] [CrossRef]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The first microbial colonizers of the human gut: Composition, activities, and health implications of the infant gut microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, 81. [Google Scholar] [CrossRef]
- O’mahony, S.M.; Clarke, G.; Dinan, T.G.; Cryan, J.F. Early-life adversity and brain development: Is the microbiome a missing piece of the puzzle? Neuroscience 2017, 342, 37–54. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, S.E.; Hickey, L.; Donath, S.; Opie, G.F.; Anderson, P.J.; Garland, S.M.; Cheong, J.L.Y. Probiotics, prematurity and neurodevelopment: Follow-up of a randomised trial. BMJ Paediatr. Open 2017, 1, e000176. [Google Scholar] [CrossRef] [PubMed]
- Sazawal, S.; Hiremath, G.; Dhingra, U.; Malik, P.; Deb, S.; Black, R.E. Efficacy of Probiotics in Prevention of Acute Diarrhoea: A Meta-Analysis of Masked, Randomised, Placebo-Controlled Trials. Lancet Infect. Dis. 2006, 6, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [PubMed]
- Fantinato, V.; Camargo, H.R.; Sousa, A.L.O.P. Probiotics study with Streptococcus salivarius and its ability to produce bacteriocins and adherence to KB cells. Rev. Odontol. UNESP 2019, 48, 1–9. [Google Scholar] [CrossRef]
- Chang, Y.; Jeong, C.H.; Cheng, W.N.; Choi, Y.; Shin, D.M.; Lee, S.; Han, S.G. Quality characteristics of yogurts fermented with short-chain fatty acid-producing probiotics and their effects on mucin production and probiotic adhesion onto human colon epithelial cells. J. Dairy. Sci. 2021, 104, 7415–7425. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. 2019, 10 (Suppl. 1), S49–S66. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, E.F.; Cotter, P.D.; Stanton, C.; Ross, R.P.; Hill, C. Production of bioactive substances by intestinal bacteria as a basis for explaining probiotic mechanisms: Bacteriocins and conjugated linoleic acid. Int. J. Food Microbiol. 2012, 16, 189–205. [Google Scholar] [CrossRef]
- Collado, M.; Meriluoto, J.; Salminen, S. Role of commercial probiotic strains against human pathogen adhesion to intestinal mucus. Lett. Appl. Microbiol. 2007, 45, 454–460. [Google Scholar] [CrossRef]
- Lee, B.; Bak, Y. Irritable bowel syndrome, gut microbiota and probiotics. J. Neurogastroenterol. Motil. 2011, 17, 252–266. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Plaza-Diaz, J. Beneficial effects of rosemary, sage and propolis on human health. Nutrients 2018, 10, 1629. [Google Scholar]
- Maruta, H.; Yoshimura, Y.; Araki, A.; Kimoto, M.; Takahashi, Y.; Yamashita, H. Activation of AMP-Activated Protein Kinase and Stimulation of Energy Metabolism by Acetic Acid in L6 Myotube Cells. PLoS ONE 2016, 27, e0158055. [Google Scholar] [CrossRef]
- Lee, J.H.; Jun, H.-S. Role of myokines in regulating skeletal muscle mass and function. Front. Physiol. 2019, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, W.K.; Williams, J.; Atherton, P.; Larvin, M.; Lund, J.; Narici, M. Sarcopenia, dynapenia, and the impact of advancing age on human skeletal muscle size and strength; a quantitative review. Front. Physiol. 2012, 11, 260. [Google Scholar] [CrossRef]
- Philp, A.; Hargreaves, M.; Baar, K. More than a store: Regulatory roles for glycogen in skeletal muscle adaptation to exercise. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1343–E1351. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Li, N.; Shi, J.; Li, H.; Yue, Y.; Jiao, W.; Wang, N.; Song, Y.; Huo, G.; Li, B. Lactobacillus acidophilus alleviates type 2 diabetes by regulating hepatic glucose, lipid metabolism and gut microbiota in mice. Food Funct. 2019, 1, 15804–15815. [Google Scholar] [CrossRef]
- Vatsalya, V.; Feng, W.; Kong, M.; Hu, H.; Szabo, G.; McCullough, A.; Dasarathy, S.; Nagy, L.E.; Radaeva, S.; Barton, B.; et al. The Beneficial Effects of Lactobacillus GG Therapy on Liver and Drinking Assessments in Patients with Moderate Alcohol-Associated Hepatitis. Am. J. Gastroenterol. 2023, 118, 1457–1460. [Google Scholar] [CrossRef]
- Nabavi, S.; Rafraf, M.; Somi, M.H.; Homayouni-Rad, A.; Asghari-Jafarabadi, M. Effects of probiotic yogurt consumption on metabolic factors in individuals with nonalcoholic fatty liver disease. J. Dairy. Sci. 2014, 97, 7386–7393. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Y.; Guo, X.; Ma, Y.; Zhang, H.; Liang, H. Effect of Lactobacillus casei on lipid metabolism and intestinal microflora in patients with alcoholic liver injury. Eur. J. Clin. Nutr. 2021, 75, 1227–1236. [Google Scholar] [CrossRef]
- Ramachandran, G.; Pottakkat, B.; Basu, S.; Mohan, P. Effect of probiotics on nutritional status, biochemical parameters, and disease severity in cirrhotic patients referred for liver transplantation-A randomised double blind, placebo-controlled trial. Clin. Nutr. ESPEN 2023, 57, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Kirpich, I.A.; Solovieva, N.V.; Leikhter, S.N.; Shidakova, N.A.; Lebedeva, O.V.; Sidorov, P.I.; Bazhukova, T.A.; Soloviev, A.G.; Barve, S.S.; McClain, C.J.; et al. Probiotics restore bowel flora and improve liver enzymes in human alcohol-induced liver injury: A pilot study. Alcohol 2008, 42, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Georges, J.; Lowery, R.P.; Yaman, G.; Kerio, C.; Ormes, J.; McCleary, S.A.; Sharp, M.; Shields, K.; Rauch, J.; Silva, J.; et al. The effects of probiotic supplementation on lean body mass, strength, and power, and health indicators in resistance trained males: A pilot study. J. Int. Soc. Sports Nutr. 2014, 1, 11. [Google Scholar] [CrossRef]
- Cheng, Y.C.; Lee, C.C.; Lee, M.C.; Hsu, H.Y.; Lin, J.S.; Huang, C.C.; Watanabe, K. Effects of heat-killed Lactiplantibacillus plantarum TWK10 on exercise performance, fatigue, and muscle growth in healthy male adults. Physiol. Rep. 2023, 11, e15835. [Google Scholar] [CrossRef] [PubMed]
- Gepner, Y.; Hoffman, J.R.; Shemesh, E.; Stout, J.R.; Church, D.D.; Varanoske, A.N.; Zelicha, H.; Shelef, I.; Chen, Y.; Frankel, H.; et al. Combined effect of Bacillus coagulans GBI-30, 6086 and HMB supplementation on muscle integrity and cytokine response during intense military training. J. Appl. Physiol. 2017, 1, 11–18. [Google Scholar] [CrossRef]
- Osterberg, K.L.; Boutagy, N.E.; McMillan, R.P.; Stevens, J.R.; Frisard, M.I.; Kavanaugh, J.W.; Davy, B.M.; Davy, K.P.; Hulver, M.W. Probiotic supplementation attenuates increases in body mass and fat mass during high-fat diet in healthy young adults. Obesity 2015, 23, 2364–2370. [Google Scholar] [CrossRef]
- Lee, M.C.; Ho, C.S.; Hsu, Y.J.; Huang, C.C. Live and Heat-Killed Probiotic Lactobacillus paracasei PS23 Accelerated the Improvement and recovery of strength and damage biomarkers after exercise-induced muscle damage. Nutrients 2022, 14, 4563. [Google Scholar] [CrossRef]
- Huang, W.C.; Lee, M.C.; Lee, C.C.; Ng, K.S.; Hsu, Y.J.; Tsai, T.Y.; Young, S.L.; Lin, J.S.; Huang, C.C. Effect of Lactobacillus plantarum TWK10 on Exercise Physiological Adaptation, Performance, and Body Composition in Healthy Humans. Nutrients 2019, 19, 2836. [Google Scholar] [CrossRef]




| Subjects (n) | Age | Physical Condition | Dosage | Effects | Reference |
|---|---|---|---|---|---|
| 19 men | 18–40 years | Healthy moderate-drinking | 100 mL whisky (32 g alcohol per day) or mineral water daily for 4 weeks | Increase in adiponectin concentration, and, in particular, HMW (high molecular weight) adiponectin, associated with skeletal muscle oxidative capacity | [63] |
| 31 men and 4 women | 53.3 ± 11.58 years | Alcoholic patients | >150 g alcohol/day during a prolonged period (>5 years), with an estimated total, lifelong consumption of 24 ± 16 kg ethanol/kg body weight | Increase in IL-15 correlated with increased protein content in muscle fibers, promoting myogenic differentiation and muscle growth | [64] |
| 30 women | 43 ± 5 years | Alcoholic patients hospitalized with a regular intake of alcohol of 3 years | During hospitalization (2–3 weeks), the patients abstained from alcohol, but before they consumed 4 units/day ethanol and 14 units/week ethanol | Atrophy both fast and slow muscle fibers, impairments in IGF-1 dependent signaling and pathways controlling translation initiation (AMPK/mTor/4E-BP1), increase in level of calpain-1 and ubiquitinated proteins | [65] |
| 46 men | 29.6 ± 4.2 years | Alcohol abusers | 3–10 years history of alcohol abuse | Low plasma testosterone accompanied by a low LH and FSH | [66] |
| 8 men | 21.4 ± 4.8 years | Healthy and trained | 60 mL of vodka across 3 h period (12 ± 2 drinks consumed) every 30 min and 1 h post-exercise | Pro-apoptotic effects in skeletal muscle following exercise | [67] |
| 4 men and 1 woman | 49.2 ± 11.4 | Alcoholic cirrhosis | ? | Skeletal muscle autophagy | [8] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sausa, M.; Fucarino, A.; Paladino, L.; Zummo, F.P.; Fabbrizio, A.; Di Felice, V.; Rappa, F.; Barone, R.; Marino Gammazza, A.; Macaluso, F. Probiotics as Potential Therapeutic Agents: Safeguarding Skeletal Muscle against Alcohol-Induced Damage through the Gut–Liver–Muscle Axis. Biomedicines 2024, 12, 382. https://doi.org/10.3390/biomedicines12020382
Sausa M, Fucarino A, Paladino L, Zummo FP, Fabbrizio A, Di Felice V, Rappa F, Barone R, Marino Gammazza A, Macaluso F. Probiotics as Potential Therapeutic Agents: Safeguarding Skeletal Muscle against Alcohol-Induced Damage through the Gut–Liver–Muscle Axis. Biomedicines. 2024; 12(2):382. https://doi.org/10.3390/biomedicines12020382
Chicago/Turabian StyleSausa, Martina, Alberto Fucarino, Letizia Paladino, Francesco Paolo Zummo, Antonio Fabbrizio, Valentina Di Felice, Francesca Rappa, Rosario Barone, Antonella Marino Gammazza, and Filippo Macaluso. 2024. "Probiotics as Potential Therapeutic Agents: Safeguarding Skeletal Muscle against Alcohol-Induced Damage through the Gut–Liver–Muscle Axis" Biomedicines 12, no. 2: 382. https://doi.org/10.3390/biomedicines12020382
APA StyleSausa, M., Fucarino, A., Paladino, L., Zummo, F. P., Fabbrizio, A., Di Felice, V., Rappa, F., Barone, R., Marino Gammazza, A., & Macaluso, F. (2024). Probiotics as Potential Therapeutic Agents: Safeguarding Skeletal Muscle against Alcohol-Induced Damage through the Gut–Liver–Muscle Axis. Biomedicines, 12(2), 382. https://doi.org/10.3390/biomedicines12020382