
Peptide-Based Biomaterials for Bone and Cartilage Regeneration

 ,
, 
Abstract
1. Introduction
2. Requirements for Osteochondral Tissue Regeneration
3. Peptides for Bone Regeneration
3.1. Osteo-Inducers
3.1.1. Collagen-Mimetic/Derived Peptides
3.1.2. BMP-Mimetic/Derived Peptides
3.1.3. Hormone-Derived Peptides
3.1.4. Circulating Peptides
3.1.5. Other ECM-Derived Peptides
3.2. Biomineralizing Peptides
3.3. Angiogenic Peptides
4. Peptides for Cartilage Regeneration
4.1. Chondroinductive/Chondrogenic Peptides
4.1.1. TGF-β Mimetic Peptides
4.1.2. BMP2-Derived/Mimetic Peptides
5. Other Supporting Peptides
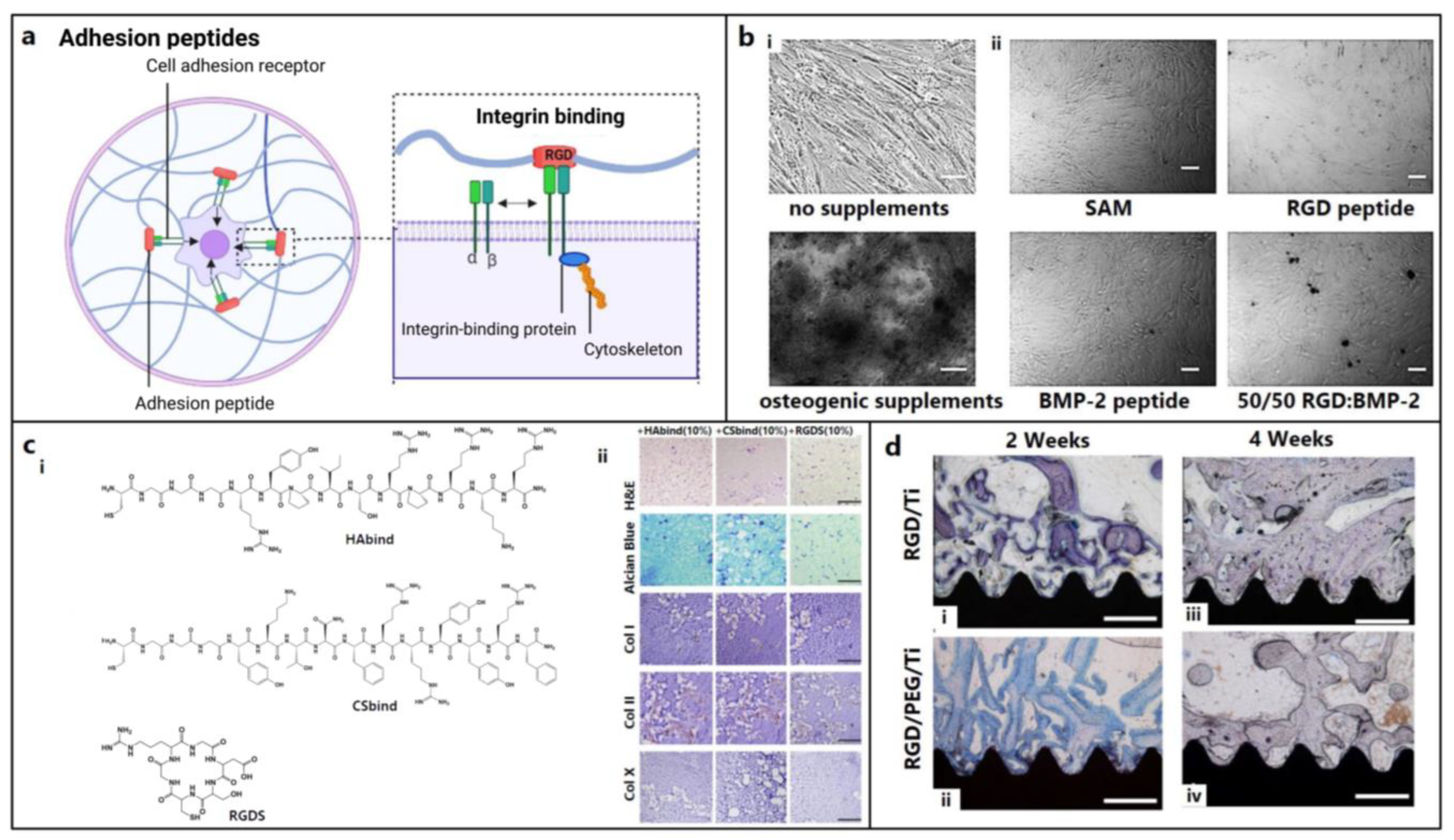
5.1. Adhesion, Binding, or Affinity Peptides
5.2. Cell-Penetrating Peptides (CPPs)
5.3. Peptides Promoting Cell Migration
5.4. Self-Assembly (SA) Peptides
5.5. Degradable Peptides
5.6. Antimicrobial and Immunomodulatory Peptides
6. Peptide-Conjugated Biomaterials
6.1. Osteo-Inductive Scaffold
6.2. Chondro-Inductive Scaffolds
6.3. Multifunctional Scaffolds
7. Summary and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yildirim, N.; Amanzhanova, A.; Kulzhanova, G.; Mukasheva, F.; Erisken, C. Osteochondral interface: Regenerative engineering and challenges. ACS Biomater. Sci. Eng. 2023, 9, 1205–1223. [Google Scholar] [CrossRef]
- Steinmetz, J.D.; Culbreth, G.T.; Haile, L.M.; Rafferty, Q.; Lo, J.; Fukutaki, K.G.; Cruz, J.A.; Smith, A.E.; Vollset, S.E.; Brooks, P.M. Global, regional, and national burden of osteoarthritis, 1990–2020 and projections to 2050: A systematic analysis for the Global Burden of Disease Study 2021. Lancet Rheumatol. 2023, 5, e508–e522. [Google Scholar] [CrossRef]
- Howell, M.; Liao, Q.; Gee, C.W. Surgical management of osteochondral defects of the knee: An educational review. Curr. Rev. Musculoskelet. Med. 2021, 14, 60–66. [Google Scholar] [CrossRef]
- Zhu, M.; Zhong, W.; Cao, W.; Zhang, Q.; Wu, G. Chondroinductive/chondroconductive peptides and their-functionalized biomaterials for cartilage tissue engineering. Bioact. Mater. 2022, 9, 221–238. [Google Scholar] [CrossRef]
- Madry, H. Surgical therapy in osteoarthritis. Osteoarthr. Cartil. 2022, 30, 1019–1034. [Google Scholar] [CrossRef]
- Wei, W.; Dai, H. Articular cartilage and osteochondral tissue engineering techniques: Recent advances and challenges. Bioact. Mater. 2021, 6, 4830–4855. [Google Scholar] [CrossRef]
- Makris, E.A.; Gomoll, A.H.; Malizos, K.N.; Hu, J.C.; Athanasiou, K.A. Repair and tissue engineering techniques for articular cartilage. Nat. Rev. Rheumatol. 2015, 11, 21–34. [Google Scholar] [CrossRef]
- Chen, L.; Wei, L.; Su, X.; Qin, L.; Xu, Z.; Huang, X.; Chen, H.; Hu, N. Preparation and Characterization of Biomimetic Functional Scaffold with Gradient Structure for Osteochondral Defect Repair. Bioengineering 2023, 10, 213. [Google Scholar]
- Rizzo, M.G.; Palermo, N.; D’Amora, U.; Oddo, S.; Guglielmino, S.P.P.; Conoci, S.; Szychlinska, M.A.; Calabrese, G. Multipotential Role of Growth Factor Mimetic Peptides for Osteochondral Tissue Engineering. Int. J. Mol. Sci. 2022, 23, 7388. [Google Scholar] [CrossRef]
- Bullock, G.; Atkinson, J.; Gentile, P.; Hatton, P.; Miller, C. Osteogenic peptides and attachment methods determine tissue regeneration in modified bone graft substitutes. J. Funct. Biomater. 2021, 12, 22. [Google Scholar] [CrossRef]
- Visser, R.; Rico-Llanos, G.A.; Pulkkinen, H.; Becerra, J. Peptides for bone tissue engineering. J. Control. Release 2016, 244, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Pountos, I.; Panteli, M.; Lampropoulos, A.; Jones, E.; Calori, G.M.; Giannoudis, P.V. The role of peptides in bone healing and regeneration: A systematic review. BMC Med. 2016, 14, 103. [Google Scholar]
- Ai, C.; Lee, Y.H.D.; Tan, X.H.; Tan, S.H.S.; Hui, J.H.P.; Goh, J.C.-H. Osteochondral tissue engineering: Perspectives for clinical application and preclinical development. J. Orthop. Transl. 2021, 30, 93–102. [Google Scholar] [CrossRef]
- Reyes, C.D.; Petrie, T.A.; Burns, K.L.; Schwartz, Z.; García, A.J. Biomolecular surface coating to enhance orthopaedic tissue healing and integration. Biomaterials 2007, 28, 3228–3235. [Google Scholar] [CrossRef]
- Bhatnagar, R.S.; Qian, J.J.; Wedrychowska, A.; Sadeghi, M.; Wu, Y.M.; Smith, N. Design of biomimetic habitats for tissue engineering with P-15, a synthetic peptide analogue of collagen. Tissue Eng. 1999, 5, 53–65. [Google Scholar] [CrossRef]
- Uysal, O.; Arslan, E.; Gulseren, G.; Kilinc, M.C.; Dogan, I.; Ozalp, H.; Caglar, Y.S.; Guler, M.O.; Tekinay, A.B. Collagen peptide presenting nanofibrous scaffold for intervertebral disc regeneration. ACS Appl. Bio Mater. 2019, 2, 1686–1695. [Google Scholar]
- Anderson, J.M.; Vines, J.B.; Patterson, J.L.; Chen, H.; Javed, A.; Jun, H.-W. Osteogenic differentiation of human mesenchymal stem cells synergistically enhanced by biomimetic peptide amphiphiles combined with conditioned medium. Acta Biomater. 2011, 7, 675–682. [Google Scholar] [CrossRef]
- Sindrey, D.; Pugh, S.; Smith, T. Connective Tissue Stimulating Peptides. U.S. Patent US20050288229A1, 29 December 2005. [Google Scholar]
- Huang, W.; Yu, K.; Kang, M.; Wang, Q.; Liao, W.; Liang, P.; Liu, G.; Cao, Y.; Miao, J. Identification and functional analysis of three novel osteogenic peptides isolated from tilapia scale collagen hydrolysate. Food Res. Int. 2022, 162, 111993. [Google Scholar] [CrossRef]
- Saito, A.; Suzuki, Y.; Ogata, S.-i.; Ohtsuki, C.; Tanihara, M. Activation of osteo-progenitor cells by a novel synthetic peptide derived from the bone morphogenetic protein-2 knuckle epitope. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2003, 1651, 60–67. [Google Scholar] [CrossRef]
- Saito, A.; Suzuki, Y.; Kitamura, M.; Ogata, S.I.; Yoshihara, Y.; Masuda, S.; Ohtsuki, C.; Tanihara, M. Repair of 20-mm long rabbit radial bone defects using BMP-derived peptide combined with an α-tricalcium phosphate scaffold. J. Biomed. Mater. Res. Part A Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2006, 77, 700–706. [Google Scholar] [CrossRef]
- Lin, Z.-Y.; Duan, Z.-X.; Guo, X.-D.; Li, J.-F.; Lu, H.-W.; Zheng, Q.-X.; Quan, D.-P.; Yang, S.-H. Bone induction by biomimetic PLGA-(PEG-ASP) n copolymer loaded with a novel synthetic BMP-2-related peptide in vitro and in vivo. J. Control. Release 2010, 144, 190–195. [Google Scholar] [CrossRef]
- Duan, Z.; Zheng, Q.; Guo, X.; Yuan, Q.; Chen, S. Experimental research on ectopic osteogenesis of BMP2-derived peptide P24 combined with PLGA copolymers. J. Huazhong Univ. Sci. Technol. 2007, 27, 179–182. [Google Scholar] [CrossRef]
- Saito, A.; Suzuki, Y.; Ogata, S.I.; Ohtsuki, C.; Tanihara, M. Prolonged ectopic calcification induced by BMP-2–derived synthetic peptide. J. Biomed. Mater. Res. Part A Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2004, 70, 115–121. [Google Scholar] [CrossRef]
- Kang, E.-J.; Kim, S.-K.; Eom, T.-G.; Choi, K.-O.; Lee, T.-H. Evaluation of the osteogenic activity of the BMP-2 mimetic peptide, PEP7, in vitro and in vivo. Int. J. Oral Maxillofac. Implant. 2013, 28, 749–756. [Google Scholar] [CrossRef][Green Version]
- Patterson, J. Peptide-functionalized Biomaterials with Osteoinductive or Anti-biofilm Activity. In Racing for the Surface: Antimicrobial and Interface Tissue Engineering; Springer: Morgantown, WV, USA, 2020; pp. 129–168. [Google Scholar]
- Kim, H.K.; Lee, J.S.; Kim, J.H.; Seon, J.K.; Park, K.S.; Jeong, M.H.; Yoon, T.R. Bone-forming peptide-2 derived from BMP-7 enhances osteoblast differentiation from multipotent bone marrow stromal cells and bone formation. Exp. Mol. Med. 2017, 49, e328. [Google Scholar] [CrossRef] [PubMed]
- Akkiraju, H.; Bonor, J.; Nohe, A. CK2. 1, a novel peptide, induces articular cartilage formation in vivo. J. Orthop. Res. 2017, 35, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Peggion, E.; Mammi, S.; Schievano, E.; Silvestri, L.; Schiebler, L.; Bisello, A.; Rosenblatt, M.; Chorev, M. Structure−Function Studies of Analogues of Parathyroid Hormone (PTH)-1–34 Containing β-Amino Acid Residues in Positions 11–13. Biochemistry 2002, 41, 8162–8175. [Google Scholar] [CrossRef] [PubMed]
- de Castro, L.F.; Lozano, D.; Portal-Núñez, S.; Maycas, M.; De la Fuente, M.; Caeiro, J.R.; Esbrit, P. Comparison of the skeletal effects induced by daily administration of PTHrP (1–36) and PTHrP (107–139) to ovariectomized mice. J. Cell. Physiol. 2012, 227, 1752–1760. [Google Scholar] [CrossRef] [PubMed]
- Trejo, C.G.; Lozano, D.; Manzano, M.; Doadrio, J.C.; Salinas, A.J.; Dapía, S.; Gómez-Barrena, E.; Vallet-Regí, M.; García-Honduvilla, N.; Buján, J. The osteoinductive properties of mesoporous silicate coated with osteostatin in a rabbit femur cavity defect model. Biomaterials 2010, 31, 8564–8573. [Google Scholar] [CrossRef] [PubMed]
- Mrak, E.; Guidobono, F.; Moro, G.; Fraschini, G.; Rubinacci, A.; Villa, I. Calcitonin gene-related peptide (CGRP) inhibits apoptosis in human osteoblasts by β-catenin stabilization. J. Cell. Physiol. 2010, 225, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Villa, I.; Melzi, R.; Pagani, F.; Ravasi, F.; Rubinacci, A.; Guidobono, F. Effects of calcitonin gene-related peptide and amylin on human osteoblast-like cells proliferation. Eur. J. Pharmacol. 2000, 409, 273–278. [Google Scholar] [CrossRef]
- Wang, L.; Shi, X.; Zhao, R.; Halloran, B.P.; Clark, D.J.; Jacobs, C.R.; Kingery, W.S. Calcitonin-gene-related peptide stimulates stromal cell osteogenic differentiation and inhibits RANKL induced NF-κB activation, osteoclastogenesis and bone resorption. Bone 2010, 46, 1369–1379. [Google Scholar] [CrossRef]
- Xu, G.; Jiang, D. The role and mechanism of exogenous calcitonin gene-related peptide on mesenchymal stem cell proliferation and osteogenetic formation. Cell Biochem. Biophys. 2014, 69, 369–378. [Google Scholar] [CrossRef]
- Brager, M.A.; Patterson, M.J.; Connolly, J.F.; Nevo, Z. Osteogenic growth peptide normally stimulated by blood loss and marrow ablation has local and systemic effects on fracture healing in rats. J. Orthop. Res. 2000, 18, 133–139. [Google Scholar] [CrossRef]
- Fei, Q.; Guo, C.; Xu, X.; Gao, J.; Zhang, J.; Chen, T.; Cui, D. Osteogenic growth peptide enhances the proliferation of bone marrow mesenchymal stem cells from osteoprotegerin-deficient mice by CDK2/cyclin A. Acta Biochim. Biophys. Sin. 2010, 42, 801–806. [Google Scholar] [CrossRef]
- An, G.; Xue, Z.; Zhang, B.; Deng, Q.; Wang, Y.; Lv, S. Expressing osteogenic growth peptide in the rabbit bone mesenchymal stem cells increased alkaline phosphatase activity and enhanced the collagen accumulation. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 1618–1624. [Google Scholar]
- Li, G.; Cui, Y.; McILmurray, L.; Allen, W.E.; Wang, H. rhBMP-2, rhVEGF165, rhPTN and thrombin-related peptide, TP508 induce chemotaxis of human osteoblasts and microvascular endothelial cells. J. Orthop. Res. 2005, 23, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Vordemvenne, T.; Paletta, J.R.; Hartensuer, R.; Pap, T.; Raschke, M.J.; Ochman, S. Cooperative effects in differentiation and proliferation between PDGF-BB and matrix derived synthetic peptides in human osteoblasts. BMC Musculoskelet. Disord. 2011, 12, 263. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-J.; Park, Y.-J.; Lee, Y.-M.; Rhyu, I.-C.; Ku, Y. The biological effects of fibrin-binding synthetic oligopeptides derived from fibronectin on osteoblast-like cells. J. Periodontal Implant. Sci. 2012, 42, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Choo, J.-E.; Choi, Y.-S.; Park, J.-B.; Min, D.-S.; Lee, S.-J.; Rhyu, H.K.; Jo, I.-H.; Chung, C.-P.; Park, Y.-J. Assembly of collagen-binding peptide with collagen as a bioactive scaffold for osteogenesis in vitro and in vivo. Biomaterials 2007, 28, 4257–4267. [Google Scholar] [CrossRef] [PubMed]
- Egusa, H.; Kaneda, Y.; Akashi, Y.; Hamada, Y.; Matsumoto, T.; Saeki, M.; Thakor, D.K.; Tabata, Y.; Matsuura, N.; Yatani, H. Enhanced bone regeneration via multimodal actions of synthetic peptide SVVYGLR on osteoprogenitors and osteoclasts. Biomaterials 2009, 30, 4676–4686. [Google Scholar] [CrossRef]
- Hamada, Y.; Yuki, K.; Okazaki, M.; Fujitani, W.; Matsumoto, T.; Hashida, M.K.; Harutsugu, K.; Nokihara, K.; Daito, M.; Matsuura, N. Osteopontin-derived peptide SVVYGLR induces angiogenesis in vivo. Dent. Mater. J. 2004, 23, 650–655. [Google Scholar] [CrossRef]
- Park, K.M.; Lee, Y.; Son, J.Y.; Bae, J.W.; Park, K.D. In situ SVVYGLR peptide conjugation into injectable gelatin-poly (ethylene glycol)-tyramine hydrogel via enzyme-mediated reaction for enhancement of endothelial cell activity and neo-vascularization. Bioconjugate Chem. 2012, 23, 2042–2050. [Google Scholar] [CrossRef]
- Rezania, A.; Healy, K.E. Biomimetic peptide surfaces that regulate adhesion, spreading, cytoskeletal organization, and mineralization of the matrix deposited by osteoblast-like cells. Biotechnol. Prog. 1999, 15, 19–32. [Google Scholar] [CrossRef]
- El Gezawi, M.; Wölfle, U.C.; Haridy, R.; Fliefel, R.; Kaisarly, D. Remineralization, regeneration, and repair of natural tooth structure: Influences on the future of restorative dentistry practice. ACS Biomater. Sci. Eng. 2019, 5, 4899–4919. [Google Scholar] [CrossRef]
- Hsu, C.; Chung, H.; Yang, J.-M.; Shi, W.; Wu, B. Influence of 8DSS peptide on nano-mechanical behavior of human enamel. J. Dent. Res. 2011, 90, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.-Y.; Li, C.C. Microstructure and nanomechanical properties of enamel remineralized with asparagine–serine–serine peptide. Mater. Sci. Eng. C 2013, 33, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Padovano, J.; Ravindran, S.; Snee, P.; Ramachandran, A.; Bedran-Russo, A.; George, A. DMP1-derived peptides promote remineralization of human dentin. J. Dent. Res. 2015, 94, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, K.; Visakan, G.; Phark, J.-H.; Moradian-Oldak, J. Enhancing collagen mineralization with amelogenin peptide: Toward the restoration of dentin. ACS Biomater. Sci. Eng. 2020, 6, 2251–2262. [Google Scholar] [CrossRef]
- Lv, X.; Yang, Y.; Han, S.; Li, D.; Tu, H.; Li, W.; Zhou, X.; Zhang, L. Potential of an amelogenin based peptide in promoting reminerlization of initial enamel caries. Arch. Oral Biol. 2015, 60, 1482–1487. [Google Scholar] [CrossRef]
- Mukherjee, K.; Ruan, Q.; Liberman, D.; White, S.N.; Moradian-Oldak, J. Repairing human tooth enamel with leucine-rich amelogenin peptide–chitosan hydrogel. J. Mater. Res. 2016, 31, 556–563. [Google Scholar] [CrossRef]
- Ding, L.; Han, S.; Peng, X.; Wang, K.; Zheng, S.; Li, H.; Niu, Y.; Li, W.; Zhang, L. Tuftelin-derived peptide facilitates remineralization of initial enamel caries in vitro. J. Biomed. Mater. Res. Part B Appl. Biomater. 2020, 108, 3261–3269. [Google Scholar] [CrossRef]
- Wang, Q.-Q.; Miao, L.; Zhang, H.; Wang, S.Q.; Li, Q.; Sun, W. A novel amphiphilic oligopeptide induced the intrafibrillar mineralisation via interacting with collagen and minerals. J. Mater. Chem. B 2020, 8, 2350–2362. [Google Scholar] [CrossRef] [PubMed]
- Kind, L.; Stevanovic, S.; Wuttig, S.; Wimberger, S.; Hofer, J.; Müller, B.; Pieles, U. Biomimetic remineralization of carious lesions by self-assembling peptide. J. Dent. Res. 2017, 96, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Finetti, F.; Basile, A.; Capasso, D.; Di Gaetano, S.; Di Stasi, R.; Pascale, M.; Turco, C.M.; Ziche, M.; Morbidelli, L.; D’Andrea, L.D. Functional and pharmacological characterization of a VEGF mimetic peptide on reparative angiogenesis. Biochem. Pharmacol. 2012, 84, 303–311. [Google Scholar] [CrossRef]
- Chen, R.R.; Silva, E.A.; Yuen, W.W.; Mooney, D.J. Spatio–temporal VEGF and PDGF delivery patterns blood vessel formation and maturation. Pharm. Res. 2007, 24, 258–264. [Google Scholar] [CrossRef]
- Kang, H.-M.; Kang, Y.; Chun, H.J.; Jeong, J.-W.; Park, C. Evaluation of the in vitro and in vivo angiogenic effects of exendin-4. Biochem. Biophys. Res. Commun. 2013, 434, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Van Hove, A.H.; Burke, K.; Antonienko, E.; Brown, E., III; Benoit, D.S. Enzymatically-responsive pro-angiogenic peptide-releasing poly (ethylene glycol) hydrogels promote vascularization in vivo. J. Control. Release 2015, 217, 191–201. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Tomin, E.; Doty, S.B.; Lane, J.M.; Carney, D.H.; Ryaby, J.T. Thrombin peptide (TP508) promotes fracture repair by up-regulating inflammatory mediators, early growth factors, and increasing angiogenesis. J. Orthop. Res. 2005, 23, 671–679. [Google Scholar] [CrossRef]
- Hardy, B.; Raiter, A.; Weiss, C.; Kaplan, B.; Tenenbaum, A.; Battler, A. Angiogenesis induced by novel peptides selected from a phage display library by screening human vascular endothelial cells under different physiological conditions. Peptides 2007, 28, 691–701. [Google Scholar] [CrossRef]
- Renner, J.N.; Liu, J.C. Investigating the effect of peptide agonists on the chondrogenic differentiation of human mesenchymal stem cells using design of experiments. Biotechnol. Prog. 2013, 29, 1550–1557. [Google Scholar] [CrossRef]
- Zhang, Z.; Gupte, M.J.; Jin, X.; Ma, P.X. Injectable peptide decorated functional nanofibrous hollow microspheres to direct stem cell differentiation and tissue regeneration. Adv. Funct. Mater. 2015, 25, 350–360. [Google Scholar] [CrossRef]
- Akkiraju, H.; Srinivasan, P.P.; Xu, X.; Jia, X.; Safran, C.B.K.; Nohe, A. CK2. 1, a bone morphogenetic protein receptor type Ia mimetic peptide, repairs cartilage in mice with destabilized medial meniscus. Stem Cell Res. Ther. 2017, 8, 82. [Google Scholar] [CrossRef] [PubMed]
- Scholz, M.; Schleicher, P.; Sewing, A.; Gelinsky, M.; Kandziora, F. Cyclic-RGD is as effective as rhBMP-2 in anterior interbody fusion of the sheep cervical spine. Spine 2013, 38, E59–E65. [Google Scholar] [CrossRef] [PubMed]
- Connelly, J.; Petrie, T.; García, A.; Levenston, M. Fibronectin-and Collagen-Mimetic Ligands Regulate BMSC Chondrogenesis in 3D Hydrogels. Eur. Cells Mater. 2011, 22, 168. [Google Scholar] [CrossRef] [PubMed]
- Mas-Moruno, C.; Fraioli, R.; Albericio, F.; Manero, J.M.; Gil, F.J. Novel peptide-based platform for the dual presentation of biologically active peptide motifs on biomaterials. ACS Appl. Mater. Interfaces 2014, 6, 6525–6536. [Google Scholar] [CrossRef]
- Sawyer, A.A.; Hennessy, K.M.; Bellis, S.L. The effect of adsorbed serum proteins, RGD and proteoglycan-binding peptides on the adhesion of mesenchymal stem cells to hydroxyapatite. Biomaterials 2007, 28, 383–392. [Google Scholar] [CrossRef]
- Schuler, M.; Hamilton, D.W.; Kunzler, T.P.; Sprecher, C.M.; de Wild, M.; Brunette, D.M.; Textor, M.; Tosatti, S.G. Comparison of the response of cultured osteoblasts and osteoblasts outgrown from rat calvarial bone chips to nonfouling KRSR and FHRRIKA-peptide modified rough titanium surfaces. J. Biomed. Mater. Res. Part B Appl. Biomater. Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2009, 91, 517–527. [Google Scholar] [CrossRef]
- Palchesko, R.N.; Romeo, J.D.; McGowan, K.A.; Gawalt, E.S. Increased osteoblast adhesion on physically optimized KRSR modified calcium aluminate. J. Biomed. Mater. Res. Part A 2012, 100, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Yu, W.; Zhang, Y.; Zhang, F. Increased preosteoblast adhesion and osteogenic gene expression on TiO2 nanotubes modified with KRSR. J. Mater. Sci. Mater. Med. 2013, 24, 1079–1091. [Google Scholar] [CrossRef]
- Nelson, M.; Balasundaram, G.; Webster, T.J. Increased osteoblast adhesion on nanoparticulate crystalline hydroxyapatite functionalized with KRSR. Int. J. Nanomed. 2006, 1, 339. [Google Scholar]
- Balasundaram, G.; Webster, T.J. Increased osteoblast adhesion on nanograined Ti modified with KRSR. J. Biomed. Mater. Res. Part A 2007, 80, 602–611. [Google Scholar] [CrossRef]
- Frith, J.E.; Mills, R.J.; Hudson, J.E.; Cooper-White, J.J. Tailored integrin–extracellular matrix interactions to direct human mesenchymal stem cell differentiation. Stem Cells Dev. 2012, 21, 2442–2456. [Google Scholar] [CrossRef]
- Li, W.; Yu, B.; Li, M.; Sun, D.; Hu, Y.; Zhao, M.; Cui, C.-B.; Hou, S. NEMO-binding domain peptide promotes osteoblast differentiation impaired by tumor necrosis factor alpha. Biochem. Biophys. Res. Commun. 2010, 391, 1228–1233. [Google Scholar] [CrossRef]
- Dai, S.; Hirayama, T.; Abbas, S.; Abu-Amer, Y. The IκB kinase (IKK) inhibitor, NEMO-binding domain peptide, blocks osteoclastogenesis and bone erosion in inflammatory arthritis. J. Biol. Chem. 2004, 279, 37219–37222. [Google Scholar] [CrossRef]
- Kuo, Y.-C.; Wang, C.-C. Surface modification with peptide for enhancing chondrocyte adhesion and cartilage regeneration in porous scaffolds. Colloids Surf. B Biointerfaces 2011, 84, 63–70. [Google Scholar] [CrossRef]
- Yeboah, A.; Cohen, R.I.; Faulknor, R.; Schloss, R.; Yarmush, M.L.; Berthiaume, F. The development and characterization of SDF1α-elastin-like-peptide nanoparticles for wound healing. J. Control. Release 2016, 232, 238–247. [Google Scholar] [CrossRef]
- Torres, P.; Díaz, J.; Arce, M.; Silva, P.; Mendoza, P.; Lois, P.; Molina-Berríos, A.; Owen, G.I.; Palma, V.; Torres, V.A. The salivary peptide histatin-1 promotes endothelial cell adhesion, migration, and angiogenesis. FASEB J. 2017, 31, 4946–4958. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.J.; Xia, C.J.; Wei, Y.; Yao, Y.; Dong, M.W.; Lin, K.Z.; Yu, L.S.; Gao, Y.; Fan, Y.Y. Annexin A1-derived peptide Ac2-26 facilitates wound healing in diabetic mice. Wound Repair Regen. 2020, 28, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Di Grazia, A.; Cappiello, F.; Imanishi, A.; Mastrofrancesco, A.; Picardo, M.; Paus, R.; Mangoni, M.L. The frog skin-derived antimicrobial peptide esculentin-1a (1-21) NH2 promotes the migration of human HaCaT keratinocytes in an EGF receptor-dependent manner: A novel promoter of human skin wound healing? PLoS ONE 2015, 10, e0128663. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Chu, X.; Li, W.; Pan, Q.; You, H. Chondrogenic effect of precartilaginous stem cells following NLS-TAT cell penetrating peptide-assisted transfection of eukaryotic h TGF β3. J. Cell. Biochem. 2013, 114, 2588–2594. [Google Scholar] [CrossRef]
- Pan, Q.; Li, W.; Yuan, X.; Rakhmanov, Y.; Wang, P.; Lu, R.; Mao, Z.; Shang, X.; You, H. Chondrogenic effect of cell-based scaffold of self-assembling peptides/PLGA-PLL loading the hTGFβ3 plasmid DNA. J. Mater. Sci. Mater. Med. 2016, 27, 19. [Google Scholar] [CrossRef] [PubMed]
- Ikeno, M.; Hibi, H.; Kinoshita, K.; Hattori, H.; Ueda, M. Effects of self-assembling peptide hydrogel scaffold on bone regeneration with recombinant human bone morphogenetic protein-2. Oral Craniofacial Tissue Eng. 2011, 1, 91. [Google Scholar] [CrossRef] [PubMed]
- Parmar, P.A.; Chow, L.W.; St-Pierre, J.-P.; Horejs, C.-M.; Peng, Y.Y.; Werkmeister, J.A.; Ramshaw, J.A.; Stevens, M.M. Collagen-mimetic peptide-modifiable hydrogels for articular cartilage regeneration. Biomaterials 2015, 54, 213–225. [Google Scholar] [CrossRef]
- Sridhar, B.V.; Brock, J.L.; Silver, J.S.; Leight, J.L.; Randolph, M.A.; Anseth, K.S. Development of a cellularly degradable PEG hydrogel to promote articular cartilage extracellular matrix deposition. Adv. Healthc. Mater. 2015, 4, 702–713. [Google Scholar] [CrossRef]
- Hao, Z.; Chen, R.; Chai, C.; Wang, Y.; Chen, T.; Li, H.; Hu, Y.; Feng, Q.; Li, J. Antimicrobial peptides for bone tissue engineering: Diversity, effects and applications. Front. Bioeng. Biotechnol. 2022, 10, 1030162. [Google Scholar] [CrossRef] [PubMed]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of action of the antimicrobial peptide buforin II: Buforin II kills microorganisms by penetrating the cell membrane and inhibiting cellular functions. Biochem. Biophys. Res. Commun. 1998, 244, 253–257. [Google Scholar] [CrossRef]
- Sun, Y.; Chan, J.; Bose, K.; Tam, C. Simultaneous control of infection and inflammation with keratin-derived antibacterial peptides targeting TLRs and co-receptors. Sci. Transl. Med. 2023, 15, eade2909. [Google Scholar] [CrossRef]
- de Leeuw, E.; Li, C.; Zeng, P.; Li, C.; Diepeveen-de Buin, M.; Lu, W.-Y.; Breukink, E.; Lu, W. Functional interaction of human neutrophil peptide-1 with the cell wall precursor lipid II. FEBS Lett. 2010, 584, 1543–1548. [Google Scholar] [CrossRef]
- Wenzel, M.; Chiriac, A.I.; Otto, A.; Zweytick, D.; May, C.; Schumacher, C.; Gust, R.; Albada, H.B.; Penkova, M.; Krämer, U. Small cationic antimicrobial peptides delocalize peripheral membrane proteins. Proc. Natl. Acad. Sci. USA 2014, 111, E1409–E1418. [Google Scholar] [CrossRef]
- De La Fuente-Núñez, C.; Korolik, V.; Bains, M.; Nguyen, U.; Breidenstein, E.B.; Horsman, S.; Lewenza, S.; Burrows, L.; Hancock, R.E. Inhibition of bacterial biofilm formation and swarming motility by a small synthetic cationic peptide. Antimicrob. Agents Chemother. 2012, 56, 2696–2704. [Google Scholar] [CrossRef]
- Libardo, M.D.J.; Bahar, A.A.; Ma, B.; Fu, R.; McCormick, L.E.; Zhao, J.; McCallum, S.A.; Nussinov, R.; Ren, D.; Angeles-Boza, A.M. Nuclease activity gives an edge to host-defense peptide piscidin 3 over piscidin 1, rendering it more effective against persisters and biofilms. FEBS J. 2017, 284, 3662–3683. [Google Scholar] [CrossRef]
- Luca, V.; Stringaro, A.; Colone, M.; Pini, A.; Mangoni, M.L. Esculentin (1-21), an amphibian skin membrane-active peptide with potent activity on both planktonic and biofilm cells of the bacterial pathogen Pseudomonas aeruginosa. Cell. Mol. Life Sci. 2013, 70, 2773–2786. [Google Scholar] [CrossRef]
- Wang, B.; Bian, A.; Jia, F.; Lan, J.; Yang, H.; Yan, K.; Xie, L.; Qiao, H.; Chang, X.; Lin, H. “Dual-functional” strontium titanate nanotubes designed based on fusion peptides simultaneously enhancing anti-infection and osseointegration. Biomater. Adv. 2022, 133, 112650. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Han, Y.; Ding, J.; Chen, X.; Huang, S.; Xing, X.; Wu, D.; Chen, J. Regulation of an antimicrobial peptide GL13K-modified titanium surface on osteogenesis, osteoclastogenesis, and angiogenesis base on osteoimmunology. ACS Biomater. Sci. Eng. 2021, 7, 4569–4580. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Le, H.; Wang, Y.; Liu, H.; Li, Z.; Yang, X.; Wang, C.; Ding, J.; Chen, X. Instructive cartilage regeneration modalities with advanced therapeutic implantations under abnormal conditions. Bioact. Mater. 2022, 11, 317–338. [Google Scholar] [CrossRef] [PubMed]
- Bittner, S.M.; Guo, J.L.; Melchiorri, A.; Mikos, A.G. Three-dimensional printing of multilayered tissue engineering scaffolds. Mater. Today 2018, 21, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Kapat, K.; Rameshbabu, A.P.; Maity, P.P.; Mandal, A.; Bankoti, K.; Dutta, J.; Das, D.K.; Dey, G.; Mandal, M.; Dhara, S. Osteochondral defects healing using extracellular matrix mimetic phosphate/sulfate decorated GAGs-agarose gel and quantitative micro-CT evaluation. ACS Biomater. Sci. Eng. 2018, 5, 149–164. [Google Scholar] [CrossRef]
- Bittner, S.M.; Guo, J.L.; Mikos, A.G. Spatiotemporal control of growth factors in three-dimensional printed scaffolds. Bioprinting 2018, 12, e00032. [Google Scholar] [CrossRef]
- Guo, J.L.; Diaz-Gomez, L.; Xie, V.Y.; Bittner, S.M.; Jiang, E.Y.; Wang, B.; Mikos, A.G. Three-dimensional printing of click functionalized, peptide patterned scaffolds for osteochondral tissue engineering. Bioprinting 2021, 22, e00136. [Google Scholar] [CrossRef]
- Einhorn, T.A.; Gerstenfeld, L.C. Fracture healing: Mechanisms and interventions. Nat. Rev. Rheumatol. 2015, 11, 45–54. [Google Scholar] [CrossRef]
- Pozos-Guillén, A.; Molina, G.; Soviero, V.; Arthur, R.A.; Chavarria-Bolaños, D.; Acevedo, A.M. Management of dental caries lesions in Latin American and Caribbean countries. Braz. Oral Res. 2021, 35, e055. [Google Scholar] [CrossRef] [PubMed]
- Mehta, M.; Schmidt-Bleek, K.; Duda, G.N.; Mooney, D.J. Biomaterial delivery of morphogens to mimic the natural healing cascade in bone. Adv. Drug Deliv. Rev. 2012, 64, 1257–1276. [Google Scholar] [CrossRef]
- Gerber, H.-P.; Vu, T.H.; Ryan, A.M.; Kowalski, J.; Werb, Z.; Ferrara, N. VEGF couples hypertrophic cartilage remodeling, ossification and angiogenesis during endochondral bone formation. Nat. Med. 1999, 5, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Calori, G.; Albisetti, W.; Agus, A.; Iori, S.; Tagliabue, L. Risk factors contributing to fracture non-unions. Injury 2007, 38, S11–S18. [Google Scholar] [CrossRef]
- Wojtowicz, A.M.; Shekaran, A.; Oest, M.E.; Dupont, K.M.; Templeman, K.L.; Hutmacher, D.W.; Guldberg, R.E.; García, A.J. Coating of biomaterial scaffolds with the collagen-mimetic peptide GFOGER for bone defect repair. Biomaterials 2010, 31, 2574–2582. [Google Scholar] [CrossRef] [PubMed]
- Shekaran, A.; García, J.R.; Clark, A.Y.; Kavanaugh, T.E.; Lin, A.S.; Guldberg, R.E.; García, A.J. Bone regeneration using an alpha 2 beta 1 integrin-specific hydrogel as a BMP-2 delivery vehicle. Biomaterials 2014, 35, 5453–5461. [Google Scholar] [CrossRef]
- Kolambkar, Y.M.; Bajin, M.; Wojtowicz, A.; Hutmacher, D.W.; García, A.J.; Guldberg, R.E. Nanofiber orientation and surface functionalization modulate human mesenchymal stem cell behavior in vitro. Tissue Eng. Part A 2014, 20, 398–409. [Google Scholar] [CrossRef]
- Hestehave Pedersen, R.; Rasmussen, M.; Overgaard, S.; Ding, M. Effects of P-15 peptide coated hydroxyapatite on Tibial defect repair in vivo in Normal and osteoporotic rats. BioMed Res. Int. 2015, 2015, 253858. [Google Scholar] [CrossRef]
- Schmitt, C.M.; Koepple, M.; Moest, T.; Neumann, K.; Weisel, T.; Schlegel, K.A. In vivo evaluation of biofunctionalized implant surfaces with a synthetic peptide (P-15) and its impact on osseointegration. A preclinical animal study. Clin. Oral Implant. Res. 2016, 27, 1339–1348. [Google Scholar] [CrossRef]
- Kumar, V.A.; Taylor, N.L.; Jalan, A.A.; Hwang, L.K.; Wang, B.K.; Hartgerink, J.D. A nanostructured synthetic collagen mimic for hemostasis. Biomacromolecules 2014, 15, 1484–1490. [Google Scholar] [CrossRef]
- Harbers, G.M.; Healy, K.E. The effect of ligand type and density on osteoblast adhesion, proliferation, and matrix mineralization. J. Biomed. Mater. Res. Part A Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2005, 75, 855–869. [Google Scholar] [CrossRef]
- Lam, H.; Li, S.; Lou, N.; Chu, J.; Bhatnagar, R. Synthetic peptides cytomodulin-1 (CM-1) and cytomodulin-2 (CM-2) promote collagen synthesis and wound healing in vitro. In Proceedings of the the 26th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, San Francisco, CA, USA, 1–5 September 2004; pp. 5028–5030. [Google Scholar]
- Moore, N.M.; Lin, N.J.; Gallant, N.D.; Becker, M.L. Synergistic enhancement of human bone marrow stromal cell proliferation and osteogenic differentiation on BMP-2-derived and RGD peptide concentration gradients. Acta Biomater. 2011, 7, 2091–2100. [Google Scholar]
- He, X.; Ma, J.; Jabbari, E. Effect of grafting RGD and BMP-2 protein-derived peptides to a hydrogel substrate on osteogenic differentiation of marrow stromal cells. Langmuir 2008, 24, 12508–12516. [Google Scholar] [CrossRef]
- Niu, X.; Feng, Q.; Wang, M.; Guo, X.; Zheng, Q. Porous nano-HA/collagen/PLLA scaffold containing chitosan microspheres for controlled delivery of synthetic peptide derived from BMP-2. J. Control. Release 2009, 134, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-J.; Yeo, S.-I.; Park, J.-W.; Shin, H.-I.; Bae, Y.-C.; Suh, J.-Y. The effects of synthetic peptide derived from hBMP-2 on bone formation in rabbit calvarial defect. Tissue Eng. Regen. Med. 2008, 5, 488–497. [Google Scholar]
- Lee, J.S.; Lee, J.S.; Murphy, W.L. Modular peptides promote human mesenchymal stem cell differentiation on biomaterial surfaces. Acta Biomater. 2010, 6, 21–28. [Google Scholar] [CrossRef]
- Bragdon, B.; Thinakaran, S.; Moseychuk, O.; Gurski, L.; Bonor, J.; Price, C.; Wang, L.; Beamer, W.G.; Nohe, A. Casein kinase 2 regulates in vivo bone formation through its interaction with bone morphogenetic protein receptor type Ia. Bone 2011, 49, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Akkiraju, H.; Bonor, J.; Olli, K.; Bowen, C.; Bragdon, B.; Coombs, H.; Donahue, L.R.; Duncan, R.; Nohe, A. Systemic injection of CK2. 3, a novel peptide acting downstream of bone morphogenetic protein receptor BMPRIa, leads to increased trabecular bone mass. J. Orthop. Res. 2015, 33, 208–215. [Google Scholar] [CrossRef]
- Alkhiary, Y.M.; Gerstenfeld, L.C.; Krall, E.; Westmore, M.; Sato, M.; Mitlak, B.H.; Einhorn, T.A. Enhancement of experimental fracture-healing by systemic administration of recombinant human parathyroid hormone (PTH 1–34). JBJS 2005, 87, 731–741. [Google Scholar]
- Komrakova, M.; Stuermer, E.K.; Werner, C.; Wicke, M.; Kolios, L.; Sehmisch, S.; Tezval, M.; Daub, F.; Martens, T.; Witzenhausen, P. Effect of human parathyroid hormone hPTH (1–34) applied at different regimes on fracture healing and muscle in ovariectomized and healthy rats. Bone 2010, 47, 480–492. [Google Scholar] [CrossRef]
- Mognetti, B.; Marino, S.; Barberis, A.; Bravo Martin, A.-S.; Bala, Y.; Di Carlo, F.; Boivin, G.; Portigliatti Barbos, M. Experimental stimulation of bone healing with teriparatide: Histomorphometric and microhardness analysis in a mouse model of closed fracture. Calcif. Tissue Int. 2011, 89, 163–171. [Google Scholar] [CrossRef]
- Chintamaneni, S.; Finzel, K.; Gruber, B. Successful treatment of sternal fracture nonunion with teriparatide. Osteoporos. Int. 2010, 21, 1059–1063. [Google Scholar] [CrossRef]
- Whitfield, J.F.; Motley, P.; Willick, G.E. Parathyroid hormone, its fragments and their analogs for the treatment of osteoporosis. Treat. Endocrinol. 2002, 1, 175–190. [Google Scholar] [CrossRef]
- Calland, J.W.; Harris, S.E.; Carnes, D.L., Jr. Human pulp cells respond to calcitonin gene-related peptide in vitro. J. Endod. 1997, 23, 485–489. [Google Scholar] [CrossRef]
- Vignery, A.; McCarthy, T. The neuropeptide calcitonin gene-related peptide stimulates insulin-like growth factor I production by primary fetal rat osteoblasts. Bone 1996, 18, 331–335. [Google Scholar] [CrossRef]
- Ballica, R.; Valentijn, K.; Khachatryan, A.; Guerder, S.; Kapadia, S.; Gundberg, C.; Gilligan, J.; Flavell, R.A.; Vignery, A. Targeted expression of calcitonin gene-related peptide to osteoblasts increases bone density in mice. J. Bone Miner. Res. 1999, 14, 1067–1074. [Google Scholar] [CrossRef]
- Bab, I.; Gazit, D.; Chorev, M.; Muhlrad, A.; Shteyer, A.; Greenberg, Z.; Namdar, M.; Kahn, A. Histone H4-related osteogenic growth peptide (OGP): A novel circulating stimulator of osteoblastic activity. EMBO J. 1992, 11, 1867–1873. [Google Scholar] [CrossRef]
- Gabet, Y.; Müller, R.; Regev, E.; Sela, J.; Shteyer, A.; Salisbury, K.; Chorev, M.; Bab, I. Osteogenic growth peptide modulates fracture callus structural and mechanical properties. Bone 2004, 35, 65–73. [Google Scholar] [CrossRef]
- Olszewska-Pazdrak, B.; Carney, D.H. Systemic administration of thrombin peptide TP508 enhances VEGF-stimulated angiogenesis and attenuates effects of chronic hypoxia. J. Vasc. Res. 2013, 50, 186–196. [Google Scholar] [CrossRef]
- Hanratty, B.M.; Ryaby, J.T.; Pan, X.-H.; Li, G. Thrombin related peptide TP508 promoted fracture repair in a mouse high energy fracture model. J. Orthop. Surg. Res. 2009, 4, 1. [Google Scholar] [CrossRef]
- Li, X.; Wang, H.; Touma, E.; Qi, Y.; Rousseau, E.; Quigg, R.J.; Ryaby, J.T. TP508 accelerates fracture repair by promoting cell growth over cell death. Biochem. Biophys. Res. Commun. 2007, 364, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-A.; Ku, Y.; Rhyu, I.-C.; Chung, C.-P.; Park, Y.-J. Effects of fibrin-binding oligopeptide on osteopromotion in rabbit calvarial defects. J. Periodontal Implant. Sci. 2010, 40, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Martino, M.M.; Tortelli, F.; Mochizuki, M.; Traub, S.; Ben-David, D.; Kuhn, G.A.; Müller, R.; Livne, E.; Eming, S.A.; Hubbell, J.A. Engineering the growth factor microenvironment with fibronectin domains to promote wound and bone tissue healing. Sci. Transl. Med. 2011, 3, 100ra189. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Choo, J.-E.; Park, H.-J.; Park, J.-B.; Lee, S.-C.; Jo, I.; Lee, S.-J.; Chung, C.-P.; Park, Y.-J. Injectable gel with synthetic collagen-binding peptide for enhanced osteogenesis in vitro and in vivo. Biochem. Biophys. Res. Commun. 2007, 357, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.K.; Kim, M.-K.; Bae, Y.-S.; Jo, I.; Lee, S.-J.; Chung, C.-P.; Park, Y.-J. A novel collagen-binding peptide promotes osteogenic differentiation via Ca2+/calmodulin-dependent protein kinase II/ERK/AP-1 signaling pathway in human bone marrow-derived mesenchymal stem cells. Cell. Signal. 2008, 20, 613–624. [Google Scholar] [CrossRef]
- Stile, R.A.; Healy, K.E. Thermo-responsive peptide-modified hydrogels for tissue regeneration. Biomacromolecules 2001, 2, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Yarbrough, D.K.; Hagerman, E.; Eckert, R.; He, J.; Choi, H.; Cao, N.; Le, K.; Hedger, J.; Qi, F.; Anderson, M. Specific binding and mineralization of calcified surfaces by small peptides. Calcif. Tissue Int. 2010, 86, 58–66. [Google Scholar] [CrossRef]
- Tang, S.; Dong, Z.; Ke, X.; Luo, J.; Li, J. Advances in biomineralization-inspired materials for hard tissue repair. Int. J. Oral Sci. 2021, 13, 42. [Google Scholar] [CrossRef]
- Deshpande, A.S.; Fang, P.-A.; Simmer, J.P.; Margolis, H.C.; Beniash, E. Amelogenin-collagen interactions regulate calcium phosphate mineralization in vitro. J. Biol. Chem. 2010, 285, 19277–19287. [Google Scholar] [CrossRef]
- Van Hove, A.H.; Benoit, D.S. Depot-based delivery systems for pro-angiogenic peptides: A review. Front. Bioeng. Biotechnol. 2015, 3, 102. [Google Scholar] [CrossRef]
- Hamada, Y.; Nokihara, K.; Okazaki, M.; Fujitani, W.; Matsumoto, T.; Matsuo, M.; Umakoshi, Y.; Takahashi, J.; Matsuura, N. Angiogenic activity of osteopontin-derived peptide SVVYGLR. Biochem. Biophys. Res. Commun. 2003, 310, 153–157. [Google Scholar] [CrossRef]
- Hamada, Y.; Egusa, H.; Kaneda, Y.; Hirata, I.; Kawaguchi, N.; Hirao, T.; Matsumoto, T.; Yao, M.; Daito, K.; Suzuki, M. Synthetic osteopontin-derived peptide SVVYGLR can induce neovascularization in artificial bone marrow scaffold biomaterials. Dent. Mater. J. 2007, 26, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Lane, T.F.; Iruela-Arispe, M.L.; Johnson, R.S.; Sage, E.H. SPARC is a source of copper-binding peptides that stimulate angiogenesis. J. Cell Biol. 1994, 125, 929–943. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Rigueur, D.; Lyons, K.M. TGFβ signaling in cartilage development and maintenance. Birth Defects Res. Part C Embryo Today Rev. 2014, 102, 37–51. [Google Scholar] [CrossRef]
- Bhatnagar, R.S.; Qian, J.J. Peptide Compositions with Growth Factor-like Activity. U.S. Patent 5,661,127, 26 August 1997. [Google Scholar]
- Mittal, A.; Kumar, R.; Parsad, D.; Kumar, N. Cytomodulin-functionalized porous PLGA particulate scaffolds respond better to cell migration, actin production and wound healing in rodent model. J. Tissue Eng. Regen. Med. 2014, 8, 351–363. [Google Scholar] [CrossRef]
- Zhao, B.; Yang, D.; Wong, J.H.; Wang, J.; Yin, C.; Zhu, Y.; Fan, S.; Ng, T.B.; Xia, J.; Li, Z. A Thioether-Stabilized d-Proline–l-Proline-Induced β-Hairpin Peptide of Defensin Segment Increases Its Anti-Candida albicans Ability. ChemBioChem 2016, 17, 1416–1420. [Google Scholar] [CrossRef] [PubMed]
- Ruczynski, J.; Wierzbicki, P.M.; Kogut-Wierzbicka, M.; Mucha, P.; Siedlecka-Kroplewska, K.; Rekowski, P. Cell-penetrating peptides as a promising tool for delivery of various molecules into the cells. Folia Histochem. Cytobiol. 2014, 52, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Renner, J.N.; Kim, Y.; Liu, J.C. Bone morphogenetic protein-derived peptide promotes chondrogenic differentiation of human mesenchymal stem cells. Tissue Eng. Part A 2012, 18, 2581–2589. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, K.M.; Pollot, B.E.; Clem, W.C.; Phipps, M.C.; Sawyer, A.A.; Culpepper, B.K.; Bellis, S.L. The effect of collagen I mimetic peptides on mesenchymal stem cell adhesion and differentiation, and on bone formation at hydroxyapatite surfaces. Biomaterials 2009, 30, 1898–1909. [Google Scholar] [CrossRef]
- Park, J.-W.; Kurashima, K.; Tustusmi, Y.; An, C.-H.; Suh, J.-Y.; Doi, H.; Nomura, N.; Noda, K.; Hanawa, T. Bone healing of commercial oral implants with RGD immobilization through electrodeposited poly (ethylene glycol) in rabbit cancellous bone. Acta Biomater. 2011, 7, 3222–3229. [Google Scholar] [CrossRef] [PubMed]
- Paredes, V.; Salvagni, E.; Rodríguez-Castellon, E.; Gil, F.; Manero, J. Study on the use of 3-aminopropyltriethoxysilane and 3-chloropropyltriethoxysilane to surface biochemical modification of a novel low elastic modulus Ti–Nb–Hf alloy. J. Biomed. Mater. Res. Part B Appl. Biomater. 2015, 103, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Benoit, D.S.; Anseth, K.S. The effect on osteoblast function of colocalized RGD and PHSRN epitopes on PEG surfaces. Biomaterials 2005, 26, 5209–5220. [Google Scholar] [CrossRef] [PubMed]
- Dee, K.C.; Andersen, T.T.; Bizios, R. Design and function of novel osteoblast-adhesive peptides for chemical modification of biomaterials. J. Biomed. Mater. Res. 1998, 40, 371–377. [Google Scholar] [CrossRef]
- Mohammed, M.; Lai, T.-S.; Lin, H.-C. Substrate stiffness and sequence dependent bioactive peptide hydrogels influence the chondrogenic differentiation of human mesenchymal stem cells. J. Mater. Chem. B 2021, 9, 1676–1685. [Google Scholar] [CrossRef]
- Li, R.; Xu, J.; Wong, D.S.H.; Li, J.; Zhao, P.; Bian, L. Self-assembled N-cadherin mimetic peptide hydrogels promote the chondrogenesis of mesenchymal stem cells through inhibition of canonical Wnt/β-catenin signaling. Biomaterials 2017, 145, 33–43. [Google Scholar] [CrossRef]
- Zhu, M.; Lin, S.; Sun, Y.; Feng, Q.; Li, G.; Bian, L. Hydrogels functionalized with N-cadherin mimetic peptide enhance osteogenesis of hMSCs by emulating the osteogenic niche. Biomaterials 2016, 77, 44–52. [Google Scholar] [CrossRef]
- Ligorio, C.; Mata, A. Synthetic extracellular matrices with function-encoding peptides. Nat. Rev. Bioeng. 2023, 1, 518–536. [Google Scholar] [CrossRef]
- Lin, H.; Zhou, J.; Shen, L.; Ruan, Y.; Dong, J.; Guo, C.; Chen, Z. Biotin-conjugated anti-CD44 antibody–avidin binding system for the improvement of chondrocyte adhesion to scaffolds. J. Biomed. Mater. Res. Part A 2014, 102, 1140–1148. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.S.; Lui, J.C.; Baron, J. Identification of chondrocyte-binding peptides by phage display. J. Orthop. Res. 2013, 31, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Zhang, X.; Pi, Y.; Wang, X.; Jia, Z.; Zhu, J.; Dai, L.; Chen, W.; Yin, L.; Chen, H. Polycaprolactone electrospun mesh conjugated with an MSC affinity peptide for MSC homing in vivo. Biomaterials 2012, 33, 3375–3387. [Google Scholar] [CrossRef]
- Webb, D.J.; Roadcap, D.W.; Dhakephalkar, A.; Gonias, S.L. A 16-amino acid peptide from human α2-macroglobulin binds transforming growth factor-β and platelet-derived growth factor-BB. Protein Sci. 2000, 9, 1986–1992. [Google Scholar] [CrossRef]
- Shah, R.N.; Shah, N.A.; Del Rosario Lim, M.M.; Hsieh, C.; Nuber, G.; Stupp, S.I. Supramolecular design of self-assembling nanofibers for cartilage regeneration. Proc. Natl. Acad. Sci. USA 2010, 107, 3293–3298. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, S.; Fernandes, H.; Moroni, L. Decellularized extracellular matrix scaffolds for cartilage regeneration. In Cartilage Tissue Engineering Methods and Protocols; Humana: New York, NY, USA, 2015; pp. 133–151. [Google Scholar]
- Masters, K.S.; Shah, D.N.; Leinwand, L.A.; Anseth, K.S. Crosslinked hyaluronan scaffolds as a biologically active carrier for valvular interstitial cells. Biomaterials 2005, 26, 2517–2525. [Google Scholar] [CrossRef]
- Ruoslahti, E.; Pierschbacher, M.D. Arg-Gly-Asp: A versatile cell recognition signal. Cell 1986, 44, 517–518. [Google Scholar] [CrossRef]
- Kim, H.D.; Heo, J.; Hwang, Y.; Kwak, S.-Y.; Park, O.K.; Kim, H.; Varghese, S.; Hwang, N.S. Extracellular-matrix-based and Arg-Gly-Asp–modified photopolymerizing hydrogels for cartilage tissue engineering. Tissue Eng. Part A 2015, 21, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, B.; Meyer, J.; Jonczyk, A.; Kessler, H.; Adamietz, P.; Meenen, N.M.; Kantlehner, M.; Goepfert, C.; Nies, B. RGD-peptides for tissue engineering of articular cartilage. Biomaterials 2002, 23, 3455–3463. [Google Scholar] [CrossRef]
- Kim, I.L.; Khetan, S.; Baker, B.M.; Chen, C.S.; Burdick, J.A. Fibrous hyaluronic acid hydrogels that direct MSC chondrogenesis through mechanical and adhesive cues. Biomaterials 2013, 34, 5571–5580. [Google Scholar] [CrossRef]
- Homandberg, G.A.; Hui, F. Arg-Gly-Asp-Ser peptide analogs suppress cartilage chondrolytic activities of integrin-binding and nonbinding fibronectin fragments. Arch. Biochem. Biophys. 1994, 310, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wen, F.; Wu, Y.; Goh, G.S.H.; Ge, Z.; Tan, L.P.; Hui, J.H.P.; Yang, Z. Cross-talk between TGF-beta/SMAD and integrin signaling pathways in regulating hypertrophy of mesenchymal stem cell chondrogenesis under deferral dynamic compression. Biomaterials 2015, 38, 72–85. [Google Scholar] [CrossRef]
- Tavella, S.; Raffo, P.; Tacchetti, C.; Cancedda, R.; Castagnola, P. N-CAM and N-cadherin expression during in vitro chondrogenesis. Exp. Cell Res. 1994, 215, 354–362. [Google Scholar] [CrossRef]
- Bian, L.; Guvendiren, M.; Mauck, R.L.; Burdick, J.A. Hydrogels that mimic developmentally relevant matrix and N-cadherin interactions enhance MSC chondrogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 10117–10122. [Google Scholar] [CrossRef]
- Jo, J.; Song, H.; Park, S.G.; Lee, S.H.; Ko, J.J.; Park, J.H.; Jeong, J.; Cheon, Y.P.; Lee, D.R. Regulation of Differentiation Potential of Human Mesenchymal Stem Cells by Intracytoplasmic Delivery of Coactivator-Associated Arginine Methyltransferase 1 Protein Using Cell-Penetrating Peptide. Stem Cells 2012, 30, 1703–1713. [Google Scholar] [CrossRef]
- Suh, J.S.; Lee, J.Y.; Choi, Y.J.; You, H.K.; Hong, S.-D.; Chung, C.P.; Park, Y.J. Intracellular delivery of cell-penetrating peptide-transcriptional factor fusion protein and its role in selective osteogenesis. Int. J. Nanomed. 2014, 9, 1153–1166. [Google Scholar]
- Park, S.; Doh, J.; Park, S.; Lim, J.; Kim, S.; Youn, J.; Jin, H.; Seo, S.; Song, M.; Sung, S. Branched oligomerization of cell-permeable peptides markedly enhances the transduction efficiency of adenovirus into mesenchymal stem cells. Gene Ther. 2010, 17, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Trepat, X.; Chen, Z.; Jacobson, K. Cell migration. Compr. Physiol. 2012, 2, 2369. [Google Scholar] [PubMed]
- Shen, W.; Chen, X.; Chen, J.; Yin, Z.; Heng, B.C.; Chen, W.; Ouyang, H.-W. The effect of incorporation of exogenous stromal cell-derived factor-1 alpha within a knitted silk-collagen sponge scaffold on tendon regeneration. Biomaterials 2010, 31, 7239–7249. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Okumura, K.; Isogai, H.; Isogai, E. The human cathelicidin antimicrobial peptide LL-37 and mimics are potential anticancer drugs. Front. Oncol. 2015, 5, 144. [Google Scholar] [CrossRef]
- Rubert Pérez, C.M.; Álvarez, Z.; Chen, F.; Aytun, T.; Stupp, S.I. Mimicking the bioactivity of fibroblast growth factor-2 using supramolecular nanoribbons. ACS Biomater. Sci. Eng. 2017, 3, 2166–2175. [Google Scholar] [CrossRef]
- Zhang, S. Emerging biological materials through molecular self-assembly. Biotechnol. Adv. 2002, 20, 321–339. [Google Scholar] [CrossRef]
- Ryan, D.M.; Nilsson, B.L. Self-assembled amino acids and dipeptides as noncovalent hydrogels for tissue engineering. Polym. Chem. 2012, 3, 18–33. [Google Scholar] [CrossRef]
- Avinash, M.; Govindaraju, T. Nanoarchitectonics of biomolecular assemblies for functional applications. Nanoscale 2014, 6, 13348–13369. [Google Scholar] [CrossRef]
- Lee, J.Y.; Choo, J.E.; Choi, Y.S.; Lee, K.Y.; Min, D.S.; Pi, S.H.; Seol, Y.J.; Lee, S.J.; Jo, I.H.; Chung, C.P. Characterization of the surface immobilized synthetic heparin binding domain derived from human fibroblast growth factor-2 and its effect on osteoblast differentiation. J. Biomed. Mater. Res. Part A Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2007, 83, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Furuya, Y.; Inagaki, A.; Khan, M.; Mori, K.; Penninger, J.M.; Nakamura, M.; Udagawa, N.; Aoki, K.; Ohya, K.; Uchida, K. Stimulation of bone formation in cortical bone of mice treated with a receptor activator of nuclear factor-κB ligand (RANKL)-binding peptide that possesses osteoclastogenesis inhibitory activity. J. Biol. Chem. 2013, 288, 5562–5571. [Google Scholar] [CrossRef]
- Recha-Sancho, L.; Semino, C.E. Heparin-based self-assembling peptide scaffold reestablish chondrogenic phenotype of expanded de-differentiated human chondrocytes. J. Biomed. Mater. Res. Part A 2016, 104, 1694–1706. [Google Scholar] [CrossRef]
- Yamaoka, H.; Asato, H.; Ogasawara, T.; Nishizawa, S.; Takahashi, T.; Nakatsuka, T.; Koshima, I.; Nakamura, K.; Kawaguchi, H.; Chung, U.i. Cartilage tissue engineering using human auricular chondrocytes embedded in different hydrogel materials. J. Biomed. Mater. Res. Part A Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2006, 78, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kisiday, J.; Jin, M.; Kurz, B.; Hung, H.; Semino, C.; Zhang, S.; Grodzinsky, A. Self-assembling peptide hydrogel fosters chondrocyte extracellular matrix production and cell division: Implications for cartilage tissue repair. Proc. Natl. Acad. Sci. USA 2002, 99, 9996–10001. [Google Scholar] [CrossRef]
- Bin Hafeez, A.; Jiang, X.; Bergen, P.J.; Zhu, Y. Antimicrobial peptides: An update on classifications and databases. Int. J. Mol. Sci. 2021, 22, 11691. [Google Scholar] [CrossRef]
- He, Y.; Jin, Y.; Wang, X.; Yao, S.; Li, Y.; Wu, Q.; Ma, G.; Cui, F.; Liu, H. An antimicrobial peptide-loaded gelatin/chitosan nanofibrous membrane fabricated by sequential layer-by-layer electrospinning and electrospraying techniques. Nanomaterials 2018, 8, 327. [Google Scholar] [CrossRef]
- Tsai, P.-W.; Yang, C.-Y.; Chang, H.-T.; Lan, C.-Y. Human antimicrobial peptide LL-37 inhibits adhesion of Candida albicans by interacting with yeast cell-wall carbohydrates. PLoS ONE 2011, 6, e17755. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lu, Z.; Wang, F.; Wang, Y. Cathelicidin-WA polarizes E. coli K88-induced M1 macrophage to M2-like macrophage in RAW264. 7 cells. Int. Immunopharmacol. 2018, 54, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Vallet-Regí, M.; Lozano, D.; González, B.; Izquierdo-Barba, I. Biomaterials against bone infection. Adv. Healthc. Mater. 2020, 9, 2000310. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Sui, B.; Xue, Y.; Liu, X.; Sun, J. Cartilage repair in degenerative osteoarthritis mediated by squid type II collagen via immunomodulating activation of M2 macrophages, inhibiting apoptosis and hypertrophy of chondrocytes. Biomaterials 2018, 180, 91–103. [Google Scholar] [CrossRef]
- Zhang, H.; Lin, C.; Zeng, C.; Wang, Z.; Wang, H.; Lu, J.; Liu, X.; Shao, Y.; Zhao, C.; Pan, J. Synovial macrophage M1 polarisation exacerbates experimental osteoarthritis partially through R-spondin-2. Ann. Rheum. Dis. 2018, 77, 1524–1534. [Google Scholar] [CrossRef]
- Liu, H.-W.; Wei, D.-X.; Deng, J.-Z.; Zhu, J.-J.; Xu, K.; Hu, W.-H.; Xiao, S.-H.; Zhou, Y.-G. Combined antibacterial and osteogenic in situ effects of a bifunctional titanium alloy with nanoscale hydroxyapatite coating. Artif. Cells Nanomed. Biotechnol. 2018, 46, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Kumar, M.; Chansuria, J.; Singh, T.B.; Bhatnagar, R.; Shukla, V.K. Effect of Cytomodulin-10 (TGF-ß1 analogue) on wound healing by primary intention in a murine model. Int. J. Surg. 2009, 7, 460–465. [Google Scholar] [CrossRef][Green Version]
- Boda, S.K.; Wang, H.; John, J.V.; Reinhardt, R.A.; Xie, J. Dual delivery of alendronate and E7-BMP-2 peptide via calcium chelation to mineralized nanofiber fragments for alveolar bone regeneration. ACS Biomater. Sci. Eng. 2020, 6, 2368–2375. [Google Scholar] [CrossRef] [PubMed]
- Laboy-López, S.; Méndez Fernández, P.O.; Padilla-Zayas, J.G.; Nicolau, E. Bioactive Cellulose Acetate Electrospun Mats as Scaffolds for Bone Tissue Regeneration. Int. J. Biomater. 2022, 2022, 3255039. [Google Scholar] [CrossRef]
- Lukasova, V.; Buzgo, M.; Sovkova, V.; Dankova, J.; Rampichova, M.; Amler, E. Osteogenic differentiation of 3D cultured mesenchymal stem cells induced by bioactive peptides. Cell Prolif. 2017, 50, e12357. [Google Scholar] [CrossRef]
- Gentile, P.; Ferreira, A.M.; Callaghan, J.T.; Miller, C.A.; Atkinson, J.; Freeman, C.; Hatton, P.V. Multilayer nanoscale encapsulation of biofunctional peptides to enhance bone tissue regeneration in vivo. Adv. Healthc. Mater. 2017, 6, 1601182. [Google Scholar] [CrossRef]
- Gentile, P.; Ghione, C.; Tonda-Turo, C.; Kalaskar, D. Peptide functionalisation of nanocomposite polymer for bone tissue engineering using plasma surface polymerisation. RSC Adv. 2015, 5, 80039–80047. [Google Scholar] [CrossRef]
- Shekaran, A.; García, A.J. Extracellular matrix-mimetic adhesive biomaterials for bone repair. J. Biomed. Mater. Res. Part A 2011, 96, 261–272. [Google Scholar] [CrossRef]
- Hasenbein, M.; Andersen, T.; Bizios, R. Micropatterned surfaces modified with select peptides promote exclusive interactions with osteoblasts. Biomaterials 2002, 23, 3937–3942. [Google Scholar] [CrossRef]
- Posadowska, U.; Storm, I.; de Wolf, F.; van den Beucken, J.; Leeuwenburgh, S. Nanofibrillar hydrogel scaffolds from recombinant protein-based polymers with integrin-and proteoglycan-binding domains. Prod. Protein-Based Polym. Pichia Pastor. 2016, 1, 135. [Google Scholar]
- Reyes, C.D.; García, A.J. α2β1 integrin-specific collagen-mimetic surfaces supporting osteoblastic differentiation. J. Biomed. Mater. Res. Part A Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2004, 69, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Yukna, R.A.; Callan, D.P.; Krauser, J.T.; Evans, G.H.; Aichelmann-Reidy, M.E.; Moore, K.; Cruz, R.; Scott, J.B. Multi-center clinical evaluation of combination anorganic bovine-derived hydroxyapatite matrix (ABM)/cell binding peptide (P-15) as a bone replacement graft material in human periodontal osseous defects. 6-month results. J. Periodontol. 1998, 69, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Thorwarth, M.; Schultze-Mosgau, S.; Wehrhan, F.; Srour, S.; Wiltfang, J.; Neukam, F.W.; Schlegel, K.A. Enhanced bone regeneration with a synthetic cell-binding peptide—In vivo results. Biochem. Biophys. Res. Commun. 2005, 329, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Thorwarth, M.; Schultze-Mosgau, S.; Wehrhan, F.; Kessler, P.; Srour, S.; Wiltfang, J.; Schlegel, K.A. Bioactivation of an anorganic bone matrix by P-15 peptide for the promotion of early bone formation. Biomaterials 2005, 26, 5648–5657. [Google Scholar] [CrossRef] [PubMed]
- Matos, S.; Guerra, F.; Krauser, J.T.; Figueiredo, H.; Marcelino, J.P.; Sanz, M. Evaluation of an anorganic bovine-derived mineral with P-15 hydrogel bone graft: Preliminary study in a rabbit cranial bone model. Clin. Oral Implant. Res. 2012, 23, 698–705. [Google Scholar] [CrossRef]
- Culpepper, B.K.; Phipps, M.C.; Bonvallet, P.P.; Bellis, S.L. Enhancement of peptide coupling to hydroxyapatite and implant osseointegration through collagen mimetic peptide modified with a polyglutamate domain. Biomaterials 2010, 31, 9586–9594. [Google Scholar] [CrossRef]
- Wang, V.; Misra, G.; Amsden, B. Immobilization of a bone and cartilage stimulating peptide to a synthetic bone graft. J. Mater. Sci. Mater. Med. 2008, 19, 2145–2155. [Google Scholar] [CrossRef]
- Shuqiang, M.; Kunzheng, W.; Xiaoqiang, D.; Wei, W.; Mingyu, Z.; Daocheng, W. Osteogenic growth peptide incorporated into PLGA scaffolds accelerates healing of segmental long bone defects in rabbits. J. Plast. Reconstr. Aesthetic Surg. 2008, 61, 1558–1560. [Google Scholar] [CrossRef]
- Jung, R.E.; Hämmerle, C.H.; Kokovic, V.; Weber, F.E. Bone regeneration using a synthetic matrix containing a parathyroid hormone peptide combined with a grafting material. Int. J. Oral Maxillofac. Implant. 2007, 22, 258–266. [Google Scholar]
- Sheller, M.R.; Crowther, R.S.; Kinney, J.H.; Yang, J.; Di Jorio, S.; Breunig, T.; Carney, D.H.; Ryaby, J.T. Repair of rabbit segmental defects with the thrombin peptide, TP508. J. Orthop. Res. 2004, 22, 1094–1099. [Google Scholar] [CrossRef]
- Rammelt, S.; Illert, T.; Bierbaum, S.; Scharnweber, D.; Zwipp, H.; Schneiders, W. Coating of titanium implants with collagen, RGD peptide and chondroitin sulfate. Biomaterials 2006, 27, 5561–5571. [Google Scholar] [CrossRef]
- Ferris, D.; Moodie, G.; Dimond, P.; Giorani, C.; Ehrlich, M.; Valentini, R. RGD-coated titanium implants stimulate increased bone formation in vivo. Biomaterials 1999, 20, 2323–2331. [Google Scholar] [CrossRef]
- Schneiders, W.; Reinstorf, A.; Pompe, W.; Grass, R.; Biewener, A.; Holch, M.; Zwipp, H.; Rammelt, S. Effect of modification of hydroxyapatite/collagen composites with sodium citrate, phosphoserine, phosphoserine/RGD-peptide and calcium carbonate on bone remodelling. Bone 2007, 40, 1048–1059. [Google Scholar] [CrossRef]
- Zhang, L.; Hemraz, U.D.; Fenniri, H.; Webster, T.J. Tuning cell adhesion on titanium with osteogenic rosette nanotubes. J. Biomed. Mater. Res. Part A 2010, 95, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Balasundaram, G.; Shimpi, T.M.; Sanow, W.R.; Storey, D.M.; Kitchell, B.S.; Webster, T.J. Molecular plasma deposited peptides on anodized nanotubular titanium: An osteoblast density study. J. Biomed. Mater. Res. Part A 2011, 98, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Kim, S.H.; Jung, Y. In situ chondrogenic differentiation of bone marrow stromal cells in bioactive self-assembled peptide gels. J. Biosci. Bioeng. 2015, 120, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Y.; Wang, B.; Yang, J.; Heng, B.C.; Yang, Z.; Ge, Z.; Lin, J. TGF-β1 affinity peptides incorporated within a chitosan sponge scaffold can significantly enhance cartilage regeneration. J. Mater. Chem. B 2018, 6, 675–687. [Google Scholar] [CrossRef]
- Ju, X.; Liu, X.; Zhang, Y.; Chen, X.; Chen, M.; Shen, H.; Feng, Y.; Liu, J.; Wang, M.; Shi, Q. A photo-crosslinked proteinogenic hydrogel enabling self-recruitment of endogenous TGF-β1 for cartilage regeneration. Smart Mater. Med. 2022, 3, 85–93. [Google Scholar] [CrossRef]
- Hsieh, Y.-H.; Hsieh, M.-F.; Fang, C.-H.; Jiang, C.-P.; Lin, B.; Lee, H.-M. Osteochondral regeneration induced by TGF-β loaded photo cross-linked hyaluronic acid hydrogel infiltrated in fused deposition-manufactured composite scaffold of hydroxyapatite and poly (ethylene glycol)-block-poly (ε-caprolactone). Polymers 2017, 9, 182. [Google Scholar] [CrossRef]
- Ding, X.; Gao, J.; Yu, X.; Shi, J.; Chen, J.; Yu, L.; Chen, S.; Ding, J. 3D-printed porous scaffolds of hydrogels modified with TGF-β1 binding peptides to promote in vivo cartilage regeneration and animal gait restoration. ACS Appl. Mater. Interfaces 2022, 14, 15982–15995. [Google Scholar] [CrossRef]
- Burdick, J.A.; Anseth, K.S. Photoencapsulation of osteoblasts in injectable RGD-modified PEG hydrogels for bone tissue engineering. Biomaterials 2002, 23, 4315–4323. [Google Scholar] [CrossRef]
- Zhang, X.; Gu, J.; Zhang, Y.; Tan, Y.; Zhou, J.; Zhou, D. Immobilization of RGD peptide onto the surface of apatite-wollastonite ceramic for enhanced osteoblast adhesion and bone regeneration. J. Wuhan Univ. Technol.-Mater. Sci. Ed. 2014, 29, 626–634. [Google Scholar] [CrossRef]
- Pan, H.; Zheng, Q.; Yang, S.; Guo, X.; Wu, B.; Zou, Z.; Duan, Z. A novel peptide-modified and gene-activated biomimetic bone matrix accelerating bone regeneration. J. Biomed. Mater. Res. Part A 2014, 102, 2864–2874. [Google Scholar] [CrossRef]
- Wang, C.; Yue, H.; Huang, W.; Lin, X.; Xie, X.; He, Z.; He, X.; Liu, S.; Bai, L.; Lu, B. Cryogenic 3D printing of heterogeneous scaffolds with gradient mechanical strengths and spatial delivery of osteogenic peptide/TGF-β1 for osteochondral tissue regeneration. Biofabrication 2020, 12, 025030. [Google Scholar] [CrossRef]
- Zhang, S.; Zhou, X.; Liu, T.; Huang, Y.; Li, J. The effects of Peptide Mel4-coated titanium plates on infection rabbits after internal fixation of open fractures. Arch. Orthop. Trauma Surg. 2022, 142, 729–734. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, L.; Wu, D.; Huang, W.; Lin, Y.; Zhou, B.; Chen, J. The effects of titanium surfaces modified with an antimicrobial peptide GL13K by silanization on polarization, anti-inflammatory, and proinflammatory properties of macrophages. BioMed Res. Int. 2020, 2020, 2327034. [Google Scholar] [CrossRef]
- Fischer, N.G.; Chen, X.; Astleford-Hopper, K.; He, J.; Mullikin, A.F.; Mansky, K.C.; Aparicio, C. Antimicrobial and enzyme-responsive multi-peptide surfaces for bone-anchored devices. Mater. Sci. Eng. C 2021, 125, 112108. [Google Scholar] [CrossRef]
- Hoyos-Nogues, M.; Velasco, F.; Ginebra, M.-P.; Manero, J.M.; Gil, F.J.; Mas-Moruno, C. Regenerating bone via multifunctional coatings: The blending of cell integration and bacterial inhibition properties on the surface of biomaterials. ACS Appl. Mater. Interfaces 2017, 9, 21618–21630. [Google Scholar] [CrossRef]
- Cheng, H.; Yue, K.; Kazemzadeh-Narbat, M.; Liu, Y.; Khalilpour, A.; Li, B.; Zhang, Y.S.; Annabi, N.; Khademhosseini, A. Mussel-inspired multifunctional hydrogel coating for prevention of infections and enhanced osteogenesis. ACS Appl. Mater. Interfaces 2017, 9, 11428–11439. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, S.; Li, J.; Zhang, L.; Wang, Y.; Gao, H.; Cao, J. Three-dimensional printed hydroxyapatite bone tissue engineering scaffold with antibacterial and osteogenic ability. J. Biol. Eng. 2021, 15, 21. [Google Scholar]
- Liu, Z.; Yuan, X.; Liu, M.; Fernandes, G.; Zhang, Y.; Yang, S.; Ionita, C.N.; Yang, S. Antimicrobial peptide combined with BMP2-modified mesenchymal stem cells promotes calvarial repair in an osteolytic model. Mol. Ther. 2018, 26, 199–207. [Google Scholar] [CrossRef]
- Li, N.; Bai, J.; Wang, W.; Liang, X.; Zhang, W.; Li, W.; Lu, L.; Xiao, L.; Xu, Y.; Wang, Z. Facile and versatile surface functional polyetheretherketone with enhanced bacteriostasis and osseointegrative capability for implant application. ACS Appl. Mater. Interfaces 2021, 13, 59731–59746. [Google Scholar] [CrossRef]
- Yuan, X.; Ouyang, L.; Luo, Y.; Sun, Z.; Yang, C.; Wang, J.; Liu, X.; Zhang, X. Multifunctional sulfonated polyetheretherketone coating with beta-defensin-14 for yielding durable and broad-spectrum antibacterial activity and osseointegration. Acta Biomater. 2019, 86, 323–337. [Google Scholar] [CrossRef]
- Trzcińska, Z.; Bruggeman, M.; Ijakipour, H.; Hodges, N.J.; Bowen, J.; Stamboulis, A. Polydopamine linking substrate for amps: Characterisation and stability on ti6al4v. Materials 2020, 13, 3714. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, J.; Chen, J.; Nie, B.; Yue, B.; Zhang, W.; Lyu, Z.; Long, T.; Wang, Y. KR-12 coating of polyetheretherketone (PEEK) surface via polydopamine improves osteointegration and antibacterial activity in vivo. J. Mater. Chem. B 2020, 8, 10190–10204. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types | Source Molecule | Mode of Delivery | In Vitro/In Vivo Activity | [Refs] |
|---|---|---|---|---|
| a. Osteogenic peptides | ||||
| (i) Osteo-inductive peptides | ||||
| GFOGER | Col | Ti implant | Osteogenic differentiation of BMSCs, mineralization in rat | [14] |
| GTPGQGIAGQRGVV (P15 peptide) | HAP scaffold | ↑ Osteogenic effect in rat | [15] | |
| ((PKG)4-(POG)4-(DOG)4) (KOD peptide) | Peptide scaffold | ↑ GAG and Col deposition in rabbit | [16] | |
| DGEA | PA scaffold | Osteogenic differentiation of hMSC | [17] | |
| NGLPGPIGP (BCSP™-1) | Injection | In vitro mineralization of bone marrow-derived osteoblasts and in rats | [18] | |
| GPAGPHGPVG, APDPFRMY, TPERYY | Culture medium | ↑ Osteogenic activity in MC3T3-E1 cells | [19] | |
| KIPKASSVPTELSAISTLYL | BMP-2 | Culture medium, TCP scaffold | ↑ ALP activity in murine osteoprogenitor cells and bone formation in rabbit | [20,21] |
| SKIPKASSVPTGLSAISTLYLAAA (P24) | PLGA/PEG-ASP scaffold | Bone formation in Wistar rats | [22,23,24] | |
| CKIPKPSSVP-TELSAISMLYL (PEP7) | Ti implant | Bone formation in human osteoblast-like cell | [25] | |
| KIPKASSVPTELSAISTLYL | Alginate gel | Osteogenic differentiation of MSC, C3H10T1/2 and bone formation in rat | [20] | |
| NSVNSKIPKACCVPTELSAI, KIPKASSVPTELSAISTLYL, DWIVA | Alginate gel | ↑ Osteogenic activity in C3H10T1/2 cells and bone formation in rat | [11] | |
| RKKNPNCRRH | BMP-4 | Alginate gel | ↑ Osteogenic activity in hMSCs and bone formation in rabbit | [26] |
| VEHDKEFFHPRYHH (BFP-2) | BMP-7 | BFP-2-treated BMSCs | ↑ Osteogenic activity in BMSCs and ectopic bone formation in mice | [27] |
| TVPKPSSAPTQLNAISTLYF, GQGFSYPYKAVFSTQ, ETLDGQSINPKLAGL | PET sheet | ↑ Osteogenic activity of BMSCs and bone formation in mice | [26] | |
| KVGKACCVPTKLSPISVLY | BMP-9 | PET sheet | ↑ Osteogenic activity and in vivo bone formation | [26] |
| CK2.2, CK2.3 | Synthetic | Tail vein injection | ↑ BMP signaling in C3H10T1/2 cells and mice model | [28] |
| PTHrP1–34, PTHrP1–36, PTHrP107–111 | PTH | s.c. injection | Osteogenic differentiation of BMSCs, bone formation in ovariectomized mice | [29,30,31] |
| CGRP–α and β-CGRP | CGRP | Culture medium | Osteogenic effects in hOBs | [32,33,34,35] |
| ALKRQGRTLYGFGG (OGP) | Mammalian blood | Culture medium | ↑ Osteogenic effects in BMSCs | [36,37,38] |
| AGYKPDEGKRGDACEGDSGGPFV (TP508) | Thrombin | Culture medium | ↑ Proliferation, osteogenic differentiation (osteoblast) | [39,40] |
| FN III9-10/12-14 | FN | Coating on Petri plate | ↑ Osteoblast activity in human osteoblasts | [41] |
| CBM | OPN | Collagen scaffold | Osteogenic differentiation of BMSCs and bone formation in rabbit | [42] |
| SVVYGLR | OPN | Gelatin–PEG Tyr hydrogel | ↑ Neovascularization in HUVEC and s.c. injection in mice | [43,44,45] |
| FHRRIKA | BSP | Quartz surface | ↑ Osteoblast activity in osteoblast-like cells | [46] |
| (ii) Biomineralization peptides | ||||
| 8DSS | DPP | Soaking in solution | ↑ Remineralization of enamel ex vivo | [47,48] |
| 3NSS | DSS | Soaking in solution | ↑ Remineralization of human enamel ex vivo | [49] |
| DSESSEEDR | DMP-1 | Soaking in solution | ↑ Mineralization of demineralized dentin | [50] |
| shADP5, QP5, P26, P32 | Amelogenin | Soaking in solution | ↑ Remineralization of the dentin ex vivo | [51,52] |
| LRAP | Amelogenin | Chitosan hydrogel | ↑ Mineralization of human tooth ex vivo | [53] |
| TDP (DRNLGDSLHRQEI) | Tuftelin | Soaking in solution | Remineralization of enamel caries ex vivo | [54] |
| NNCCCCRRES(p) | CEMP1 | Soaking in solution | ↑ Remineralization of enamel | [55] |
| P11-4 (Ac-QQRFEWEFEQQ-NH2) | - | Hydrogel | ↑ Remineralization of dentinal collagen | [56] |
| (iii) Angiogenic peptides | ||||
| QK | VEGF | Gelatin-coated dish | ↑ Migration and proliferation of EC | [57] |
| PBA2-1c | PDGF-BB | PLG scaffold | Angiogenesis in mice | [58] |
| Exendin-4 | Exendin-4 | s.c. injection of matrigel plug | ↑ Tube formation in HUVECs and angiogenesis in mice | [59] |
| SPARC113, SPARC118 | OPN | PEG hydrogel | ↑ Angiogenesis in mouse and rat (s.c. injection) | [60] |
| TP508 | Thrombin | Percutaneous injection | ↑ Neo-angiogenesis in rat bone fracture | [61] |
| RoY | Synthetic | Culture medium; s.c. injection | ↑ Proliferation, sprouting of HUVEC, in vivo angiogenesis in mice | [62] |
| b. Chondroinductive peptides | ||||
| CMs | TGF-β | PHEMA-g-PLLA-acrylic microsphere | ↑ Chondrogenic differentiation in BMSCs | [63,64] |
| CK2.1 | Synthetic | HA hydrogel | ↑ Chondrogenesis in mice | [65] |
| c. Other supporting peptides | ||||
| (i) Adhesion, binding, affinity peptides | ||||
| Cyclic RGD | Col, FN, VN | Ti cage | ↑ Spinal fusion in sheep | [66] |
| PHSRN | FN | Ti implant | ↑ Adhesion of osteoblast-like cells | [67,68] |
| FHRRIKA | BSP | Ti implant | ↑ Mineralization in osteoblast and rat model | [69,70] |
| KRSR | BSP, FN, VN, OPN | Calcium aluminate scaffolds | ↑ Adhesion, osteogenic gene expression in mouse C3T3 fibroblasts and osteoblasts | [71,72,73,74] |
| HAV | N-Cadherin | PS-PEO surface | ↑ Adhesion of hMSCs | [75] |
| NEMO-binding domain (NBD) peptide | Synthetic | Culture medium | ↑ Osteoblast differentiation in C2C12 cells | [76,77] |
| CDPGYIGSR | Laminin | PEO/chitosan scaffold | ↑ Adhesion and ECM deposition in bovine knee chondrocytes (BKCs) | [78] |
| (ii) Peptides supporting cell migration | ||||
| SDF1-ELP | SDF-1 | Self-assembled nanoparticle | EC migration, vascularization in diabetic mice | [79] |
| Histatin-1 | Saliva | In vitro | Adhesion, migration, and angiogenesis of EC | [80] |
| Ac2-26 | Annexin A1 | Injection | Cell migration in diabetic mice | [81] |
| Esculentin 1-21 | Frog skin | Culture medium | Migration of HaCaT cells | [82] |
| (ii) Cell penetrating peptides (CPPs) | ||||
| NLS-TAT | HIV-1 Tat protein | PLGA-PLL scaffold | ↑ Chondrogenic differentiation in PSC | [83,84] |
| (iii) Self-assembly (SA) peptides | ||||
| RADA16-I | Synthetic | Ti cylinder | ↑ Mineralization in rabbit | [85] |
| (iv) Degradable peptides | ||||
| KCGPQGIWGQCK | MMP-derived | PEG hydrogel | ↑ GAGs and collagen deposition in MSCs | [86,87] |
| (v) Antimicrobial and immunomodulatory peptides | ||||
| Melamine | Synthetic | In vitro | Destabilization of cell membrane in vivo | [88] |
| LL-37 | Human cathelicidin | In vitro | Inhibits cell wall synthesis | [89] |
| Pexiganan | Synthetic analog of magainin-2 | In vivo | Disrupts bacterial membrane in diabetic foot ulcer patient | [88] |
| Magainin-1 | Frog skin | In vitro | Destabilizes or disrupts bacterial membrane | [88] |
| PLG0206 | Synthetic | In vivo | Disrupts cell membrane | [88] |
| Protegrin-1 | Porcine leukocytes | In vitro | Disrupts cell membrane | [88] |
| Bactenecin | Bovine neutrophils | In vitro | Inhibits protein synthesis | [88] |
| PEP8R | Synthetic | Hydrogel | Disrupts bacterial cell membrane | [88] |
| Indolicidin | Bovine neutrophil | In vitro | Increase cell membrane permeability | [88] |
| ILRWPWWPWRRK (Omiganan) | Synthetic | Topical application | Destabilizes cell membrane in vivo | [88] |
| RAIGGGLSSVGGGSSTIKY (KAMP-19) | Keratin | In vitro | Pore formation in bacterial cell membrane | [90] |
| HNP-1 | Human neutrophil | In vitro | Destabilizes cell wall integrity | [91] |
| RWRWRW-NH2 | Synthetic | In vitro | Delocalizes peripheral membrane proteins | [92] |
| AMP 1037 | Synthetic | In vitro | ↓ Swimming and motility of bacteria, gene expression | [93] |
| Piscidin and sculentin 1–21 | Fish | In vitro | Degrades biofilm via DNA damage in vitro | [94,95] |
| β defensin 3 | Human | Strontium titanate nanotubes | Antibacterial activity | [96] |
| GKIIKLKASLKLL (GL13K) | Salivary protein | Ti implant surface | Delocalization of bacterial cell membrane | [97] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapat, K.; Kumbhakarn, S.; Sable, R.; Gondane, P.; Takle, S.; Maity, P. Peptide-Based Biomaterials for Bone and Cartilage Regeneration. Biomedicines 2024, 12, 313. https://doi.org/10.3390/biomedicines12020313
Kapat K, Kumbhakarn S, Sable R, Gondane P, Takle S, Maity P. Peptide-Based Biomaterials for Bone and Cartilage Regeneration. Biomedicines. 2024; 12(2):313. https://doi.org/10.3390/biomedicines12020313
Chicago/Turabian StyleKapat, Kausik, Sakshi Kumbhakarn, Rahul Sable, Prashil Gondane, Shruti Takle, and Pritiprasanna Maity. 2024. "Peptide-Based Biomaterials for Bone and Cartilage Regeneration" Biomedicines 12, no. 2: 313. https://doi.org/10.3390/biomedicines12020313
APA StyleKapat, K., Kumbhakarn, S., Sable, R., Gondane, P., Takle, S., & Maity, P. (2024). Peptide-Based Biomaterials for Bone and Cartilage Regeneration. Biomedicines, 12(2), 313. https://doi.org/10.3390/biomedicines12020313